






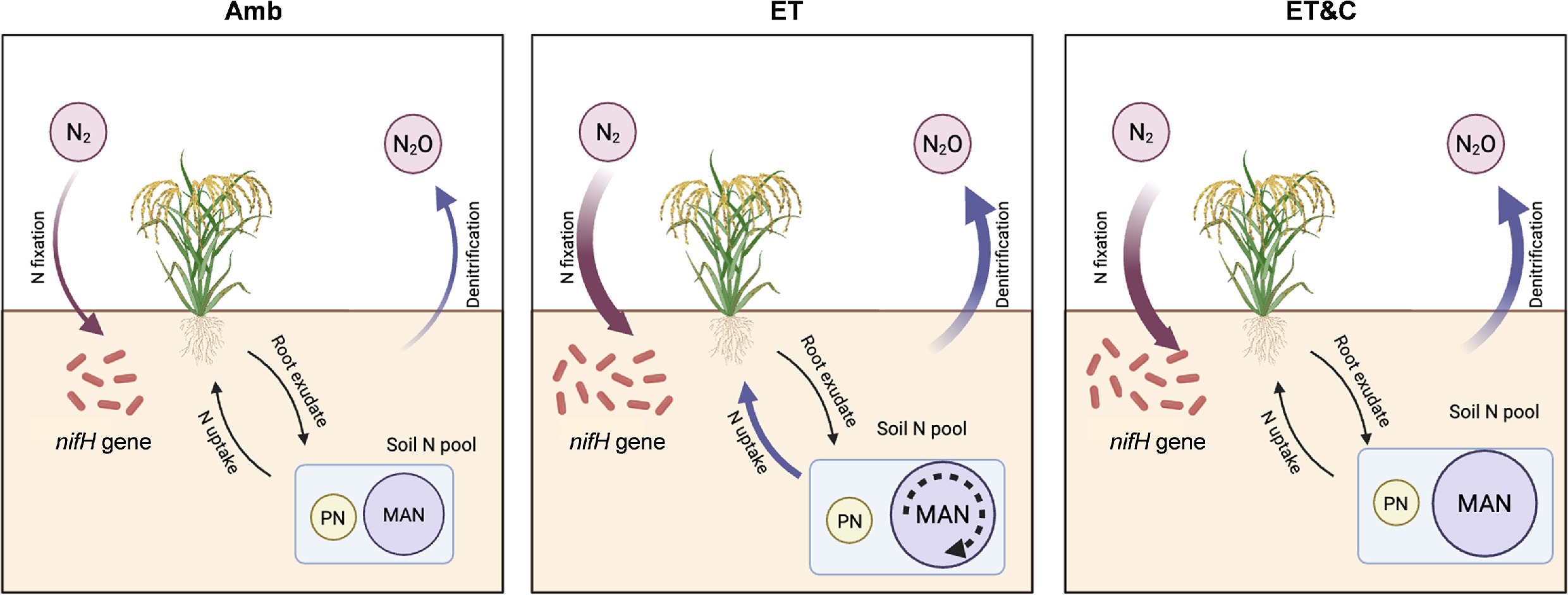



-
By the end of the 21st century, the moderate emission scenario (RCP 4.5) of the Intergovernmental Panel on Climate Change projects atmospheric CO2 to reach ~600 ppm and global mean temperature to rise by 1.5–2.0 °C[1]. These changes will shift climatic patterns and vegetation distributions, with direct and indirect impacts on agricultural productivity[2−5]. At the same time, population growth, particularly in Asia and Africa, where rice is a dietary staple, will intensify pressure on rice production[6,7]. Meeting this rising demand under a changing climate will require strategies that sustain yields while maintaining soil health[6,7].
While the effects of climate change on soil organic carbon (C) have been extensively documented across ecosystems[5,8−11], its influence on soil nitrogen (N), a fundamental nutrient for plant growth and the main component of fertilizers, remains less understood[12]. Nitrogen cycling is driven by microbial processes such as mineralization, immobilization, biological N fixation, plant uptake, and leaching[13−17]. The balance between inputs (e.g., fixation, root exudate) and outputs (e.g., plant uptake, gaseous loss, leaching, and run-off) determines long-term soil N stocks[18,19]. Elevated CO2 can stimulate plant growth and C inputs to soil, but without additional N inputs, it may exacerbate microbial N limitation, alter mineralization rates, and increase immobilization[20]. Warming generally accelerates microbial turnover and enzymatic activity, enhancing N mineralization and nitrification, although these responses depend on soil moisture, substrate supply, and microbial adaptation[21−23]. Such changes can shift the availability of inorganic N forms (NH4+, NO3−) and the magnitude of N losses[24].
Flooded rice paddies present a unique agroecosystem where anaerobic conditions suppress nitrification and favor ammonium accumulation as the dominant plant-available N form[25]. Climate change could intensify N cycling dynamics: warming may accelerate mineralization, while elevated CO2 could alter plant–microbe competition and root–soil interactions[23]. However, the fate of stabilized N in particulate organic matter (POM) and mineral-associated organic matter (MAOM) under these conditions remains poorly understood. Likewise, the potential of microbial functional gene abundance and stable isotope tracers to reveal shifts in N partitioning across soil fractions is underexplored.
Here, we examine how elevated temperature and CO2 affect N dynamics in rice agroecosystems by: (1) monitoring nitrous oxide (N2O) emissions as an indicator of gaseous N loss and assessing plant N uptake; (2) quantifying labile and stable soil N pools (particulate N and mineral-associated N) coupled with 15N analysis; and (3) assessing N-fixing microbial functional genes and root biomass and exudates as indicators of N input. By integrating plant, soil, and microbial responses, we identify mechanisms governing N stabilization and availability under future climate conditions. Our findings advance understanding of soil N resilience and inform strategies to optimize N use efficiency and sustainability in rice systems facing global change.
-
Three treatments were established in temperate rice paddy fields using an open-top chamber (OTC) system, with two replicates per treatment: (1) ambient atmospheric conditions (Amb; 0 °C temperature increase and +0 ppm CO2), (2) elevated temperature conditions (ET; +2 °C and +0 ppm CO2), and (3) elevated temperature combined with elevated CO2 (ET&C; +2 °C and +200 ppm CO2). For the ET and ET&C treatments, chamber air temperature and CO2 concentration were automatically controlled every 5 s based on real-time comparisons with the ambient (Amb) conditions. Temperature and CO2 sensors installed at the center of each chamber continuously monitored environmental conditions to guide the adjustment system. To reduce atmospheric interference among chambers, the experimental plots were evenly arranged with dimensions of 4 m × 2 m.
The OTC chambers consisted of circular stainless-steel frames with a base diameter of 5.3 m and a height of 3 m. The structures were covered with transparent tetrafluoroethylene polymer film, allowing approximately 93% UV transmission. An opening at the chamber roof (3.4 m diameter) allowed natural precipitation and wind flow to enter the chambers, thereby maintaining more realistic field conditions. The chamber walls were constructed as double layers; perforations in the inner layer prevented direct CO2 fumigation into the crop canopy and promoted gradual gas mixing and cooling within the chamber.
For CO2 enrichment, perforated distribution tubes with holes spaced at 3 cm intervals were installed between the double layers and connected to high-purity liquid CO2 cylinders (> 99.95%). These cylinders were equipped with pressure regulators and a vaporization unit. CO2 supply operated as pulsed fumigation regulated through real-time comparison with ambient conditions by offsetting 200 ppm above ambient. Chamber temperature control and ventilation were maintained using blowers directed toward the chamber center together with resistance heating elements.
Due to the substantial infrastructure and operational costs associated with constructing and maintaining active open-top chamber systems with real-time CO2 control over multiple growing seasons, two replicate plots were established per treatment. This is consistent with the plot-level replication used in our companion study conducted on the same OTC platform[4]. To improve the precision of treatment effect estimates within this design, three soil subsamples were collected from spatially distributed positions within each plot at every sampling event and treated as analytical replicates (n = 3) to characterize within-plot variability. We acknowledge that because plot-level replication is limited to n = 2 per treatment, these values reflect within-plot rather than between-plot variance, and all statistical findings should therefore be interpreted as indicative rather than conclusive.
Rice cultivation and biomass N uptake measurement
-
The soil inside the OTC plots was classified as Entisol according to the USDA Soil Taxonomy system, with a texture consisting of 40% sand, 32% silt, and 28% clay. The soil fertility level was relatively low; the surface soil layer (0–20 cm) contained 14.6 g kg−1 of soil organic C, 0.9 g kg−1 of total N, and 11 mg kg−1 of available phosphate (Lancaster-P). All chambers were managed uniformly following standard rice cultivation practices used in the Republic of Korea for three consecutive years. During the winter fallow period (late October to early May), the plots remained unmanaged without crop cover. At the beginning of May, soils were manually tilled to a depth of 20 cm and irrigated in preparation for rice transplanting.
Chemical fertilizers were applied to all treatments at the recommended rates (N–P–K = 90–19–47 kg ha−1) according to the Rural Development Administration guidelines[26]. The rice cultivar used in this study was Sae-ilmi, a japonica variety widely cultivated in Southern Korea. Seedlings were transplanted manually with a spacing of 30 cm × 15 cm. Floodwater depth was maintained automatically at 5–10 cm starting in early May and was drained in September prior to harvest in mid-October. At maturity, all aboveground biomass was harvested and dried in an oven at 60 °C for 72 h. The dried biomass was then separated into grain and straw fractions. Each fraction was weighed and finely ground before determining nitrogen concentration using an elemental analyzer (CHNS-932 Analyzer, Leco, St. Joseph, MI, USA).
N2O flux investigation
-
Nitrous oxide emissions were measured to quantify nitrogen losses from the soil under different atmospheric treatments using the static chamber method[27] during the 2nd and 3rd years. Opaque cylindrical chambers (0.3 m diameter × 0.3 m height) equipped with covers were installed on the soil surface between rice plants. To eliminate plant uptake effects within the chambers, any tiny, germinated vegetation enclosed inside the chamber area was manually removed prior to gas sampling. Two thermometers were installed to monitor air and soil temperature during sampling. An electric fan attached to the chamber lid ensured proper mixing of air within the chamber headspace during gas collection. Chambers remained open except during the sampling period. Gas samples were collected at 14-h intervals. The collected gas was analyzed using Fourier-transform infrared spectroscopy (FT-IR; CIC Photonics, NM, USA) equipped with a 9-m optical cell to determine changes in N2O concentration in the chamber headspace.
Nitrous oxide emission rates were calculated based on the increase in gas concentration within the chamber headspace based on the following equation[27]:
$ {\rm Emission\;rate\;(mg\;m^{-2}\;{h}^{-1})}=\{(C_2/T_2)-(C_1/T_1)\}\times (273/\Delta t) \times (V/A)\times \rho $ where, C1 and C2 (m3 m−3) are the inner chamber N2O concentrations before and after cover closing, respectively. T1 and T2 (K) are the absolute temperature of inside chamber before and after cover closing, respectively. ∆t (h) is closing time (0.5 h). V (m3) is the volume of the chamber. A (m2) is the surface area of the chamber. ρ (g cm−3) indicates N2O density at a standardized state.
Cumulative N2O fluxes were estimated by adding daily emission rates[28]:
$ {\rm Cumulative\; N_{2}O\; flux\; (kg\; ha^{-1})} = \sum\nolimits_{i}^{n}\left({R}_{i}\times D_{i}\right) $ where, Di is interval days between the ith and (i–1)th samplings. n is the total sampling number.
Soil sampling and N pool fractionation with isotope analysis
-
Surface soil samples (0–20 cm) were collected monthly during the rice growing season (May–October) in the 3rd year. Collected soils were freeze-dried, ground, and passed through a 2 mm sieve. A subsample (5 g) was extracted with 30 mL of ultra-deionized water to determine dissolved nitrogen concentrations. Nitrogen concentrations in the extracts were analyzed using a TOC-TN analyzer (TOC-L, Shimadzu, Japan). Another subsample (5 g) was extracted using 2 M potassium chloride (KCl) to measure inorganic N forms (NH4+ and NO3−). Soil ammonium and nitrate concentrations were quantified monthly using the salicylate method and cadmium reduction method, respectively.
At the harvest stage, soil samples were further fractionated into four operational N pools based on density and particle size: free light particulate N (FLPN), occluded particulate N (OPN), sand-associated N, and silt- and clay-associated N[29−31]. For density separation, dried and sieved soil was suspended in 100 mL sodium iodide solution (NaI; density = 1.85 g cm−3) and shaken at 30 rpm for 30 min. Floating materials were collected as light particulate N using membrane filters (0.45 µm), washed three times with deionized water to remove NaI residues, and dried at 60 °C for 72 h. The remaining soil pellet was dispersed using 5% sodium hexametaphosphate to break aggregates. Floating materials released after dispersion were collected as occluded particulate N using membrane filtration and dried at 60 °C for 72 h. The remaining soil fraction was subsequently separated into sand-associated N (> 53 µm) and silt- and clay-associated N (< 53 µm) through wet sieving. These fractions were oven-dried at 105 °C for 24 h. Each fraction was finely ground and analyzed using elemental analysis–isotope ratio mass spectrometry (EA-IRMS; Vision-EA, Isoprime, UK) to determine N concentrations and stable N isotope ratios.
Root biomass and exudates measurements
-
At the rice maturity stage, three rice plants from each plot were carefully excavated to avoid damaging the root system in the 3rd year. Aboveground biomass was separated from roots by cutting at the stem base. Roots were thoroughly washed with tap water to remove adhering soil particles. After washing, roots were gently blotted with tissue paper to remove excess moisture before weighing. Samples were then stored in labeled bags and dried in an oven at 60 °C for 72 h. After drying, samples were cooled and weighed to determine dry root biomass.
To quantify root exudates, a modified hydroponic extraction technique was employed. Intact rice plants were carefully removed from the field to minimize root damage and gently washed to remove residual soil. Roots were then transferred into glass bottles containing 250 mL of triple-distilled water. The bottles were incubated in darkness at 25 °C for 4 h to allow the release of root exudates. Following incubation, the solution was filtered through a 0.22 µm membrane filter to remove plant debris and microorganisms. The filtered solution was analyzed for total N using a TOC-TN analyzer (TOC-L, Shimadzu, Japan).
Gene quantification
-
To assess microbial functional potential related to biological nitrogen fixation, we quantified the abundance of the nifH gene, which encodes the nitrogenase reductase subunit and serves as the standard functional marker for diazotrophic microorganisms in soil[32].
Soil samples for DNA extraction were collected at the rice maturity stage (October) in the 3rd year, concurrent with soil fractionation and isotope analyses, to ensure consistency across nitrogen pool measurements. From each plot, surface soil (0–20 cm) was collected using a stainless-steel auger after removing surface litter. Samples were immediately placed on ice in the field, transported to the laboratory within 6 h, and stored at −80 °C prior to DNA extraction to minimize post-sampling microbial activity.
Prior to DNA extraction, frozen soil samples were gently homogenized under sterile conditions. Genomic DNA was extracted from 0.5 g of soil using the FastDNA™ Spin Kit for Soil (MP Biomedicals, USA) following the manufacturer's instructions. Quantification of the nifH and bacterial 16S rRNA genes was performed using StepOnePlus Real-Time PCR System (Applied Biosystems™, USA) with SYBR™ Green Universal Master Mix (Applied Biosystems™, USA) detection chemistry. Each 20 µL reaction contained 50 ng of template DNA and 10 pmol of each primer specific for nifH or 16S rRNA genes. The nifH fragments were amplified with primers PolF (50-TGCGAYCCSAARGCBGACTC-30) and PolR (50-ATSGCC-ATCATYTCRCCGGA-30)[33], and 16S rRNA fragments were amplified with primers 1055YF (ATGGYTGTCGTCAGCT) and 1392R (ACGGGCGGTGTGTAC)[34]. qPCR amplification for the nifH gene was conducted under the following thermal cycling conditions: initial denaturation at 94 °C for 5 min, followed by 40 cycles of denaturation at 94 °C for 20 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s. A melt curve analysis was performed with the following settings: 95 °C for 15 s, 60 °C for 1 min, a gradual temperature increase at 0.05 °C s−1 from 60 to 95 °C, and a final hold at 95 °C for 15 s. The qPCR conditions for bacterial 16S rRNA gene amplification were identical, except for the annealing temperature, which was set to 57 °C.
Statistical analysis
-
The effects of atmospheric treatments on measured variables were evaluated using one-way analysis of variance (ANOVA) in R. When significant treatment effects were detected, Fisher's least significant difference (LSD) test was applied for pairwise comparisons at a significance threshold of p < 0.05.
Treatment responses were further summarized using log response ratios (lnRR = ln[Treatment/Baseline mean]) to quantify effect sizes across N-related variables. The ambient treatment (Amb) was used as the baseline for warming and combined warming + CO2 effects, whereas the ET treatment served as the baseline when evaluating additional CO2 effects under warming. Because of the small sample size (n = 3), uncertainty around mean lnRR values was estimated using bootstrap resampling (5,000 iterations). The resulting 95% confidence intervals were used only for visualization purposes and do not constitute a test of statistical significance; all inference of treatment effects was based on one-way ANOVA with Fisher's LSD post-hoc comparisons (p < 0.05). We acknowledge that n = 3 replicates limit statistical power, and findings should be interpreted accordingly.
-
Nitrogen uptake by rice increased significantly under elevated temperature (ET) compared to ambient conditions (Amb), whereas the effect of additional elevated [CO2] (ET vs ET&C) was inconsistent (Fig. 1; Supplementary Figs S1 and S2). Over the past two years, ET consistently enhanced grain and straw N contents, resulting in higher total N uptake, despite no significant changes in biomass yield. In contrast, the combined treatment (ET&C) reduced N uptake relative to ET and showed values comparable to Amb, suggesting that elevated [CO2] offset the temperature-driven enhancement of N uptake. Notably, in 2021, elevated [CO2] alone suppressed grain yield, further diminishing N uptake.
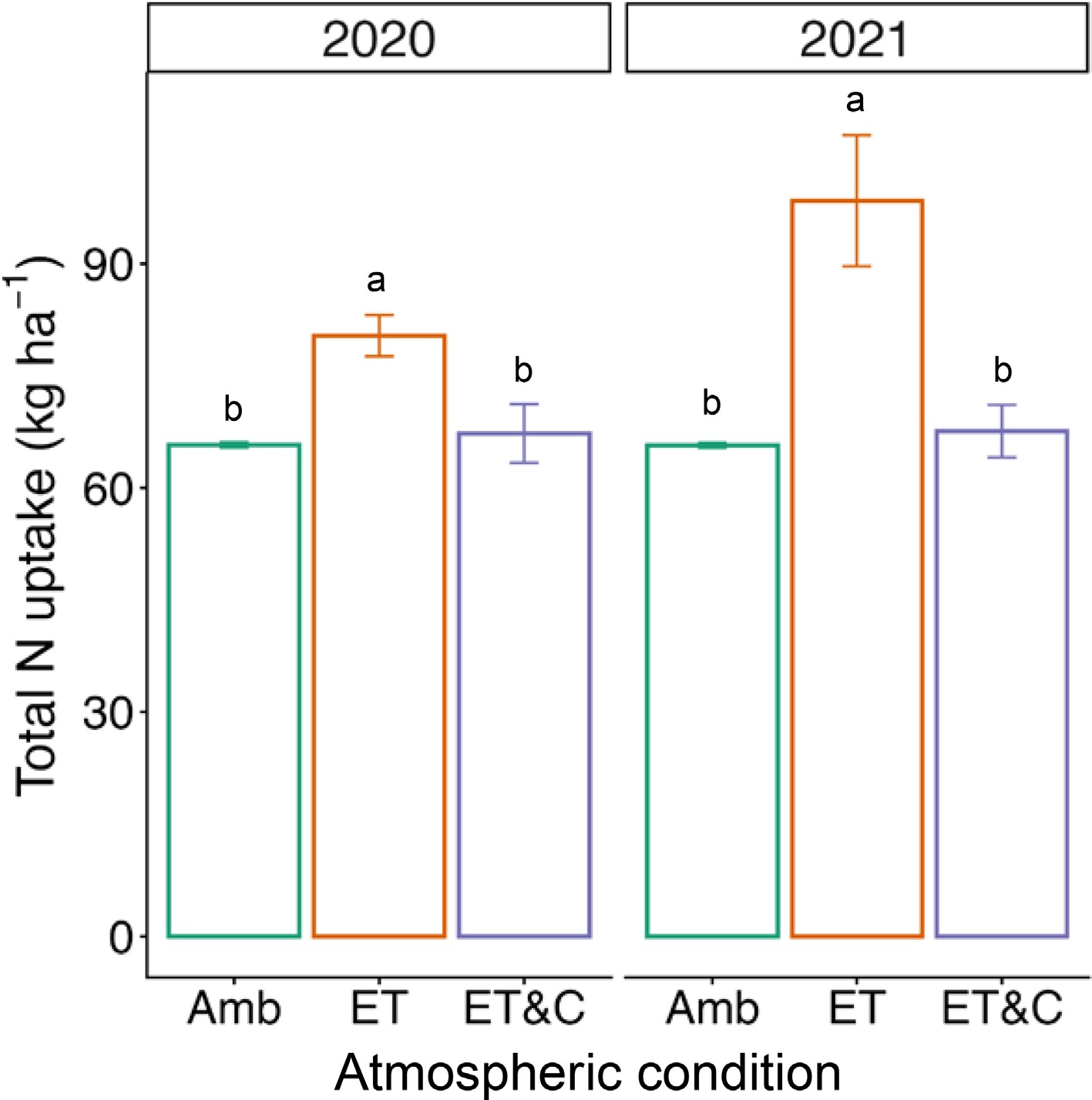
Figure 1.
Nitrogen (N) uptake of rice under different atmospheric conditions for 2 years. Error bars indicate standard deviation (n = 3 within-plot subsamples). Different letters represent statistical difference at 0.05.
N2O flux and N availability in soil
-
Elevated temperature stimulated N2O emissions, with the strongest increases observed under ET&C relative to Amb (Fig. 2; Supplementary Fig. S2). While ET alone significantly enhanced N2O fluxes, the effect of [CO2] addition was less consistent across years. Seasonal dynamics of soil inorganic N (NH4+, NO3−) and water-extractable N (WEN) showed pronounced peaks during the mid-growing season (July–August) (Supplementary Fig. S3). These peaks were particularly elevated under ET, indicating accelerated N mineralization and turnover.
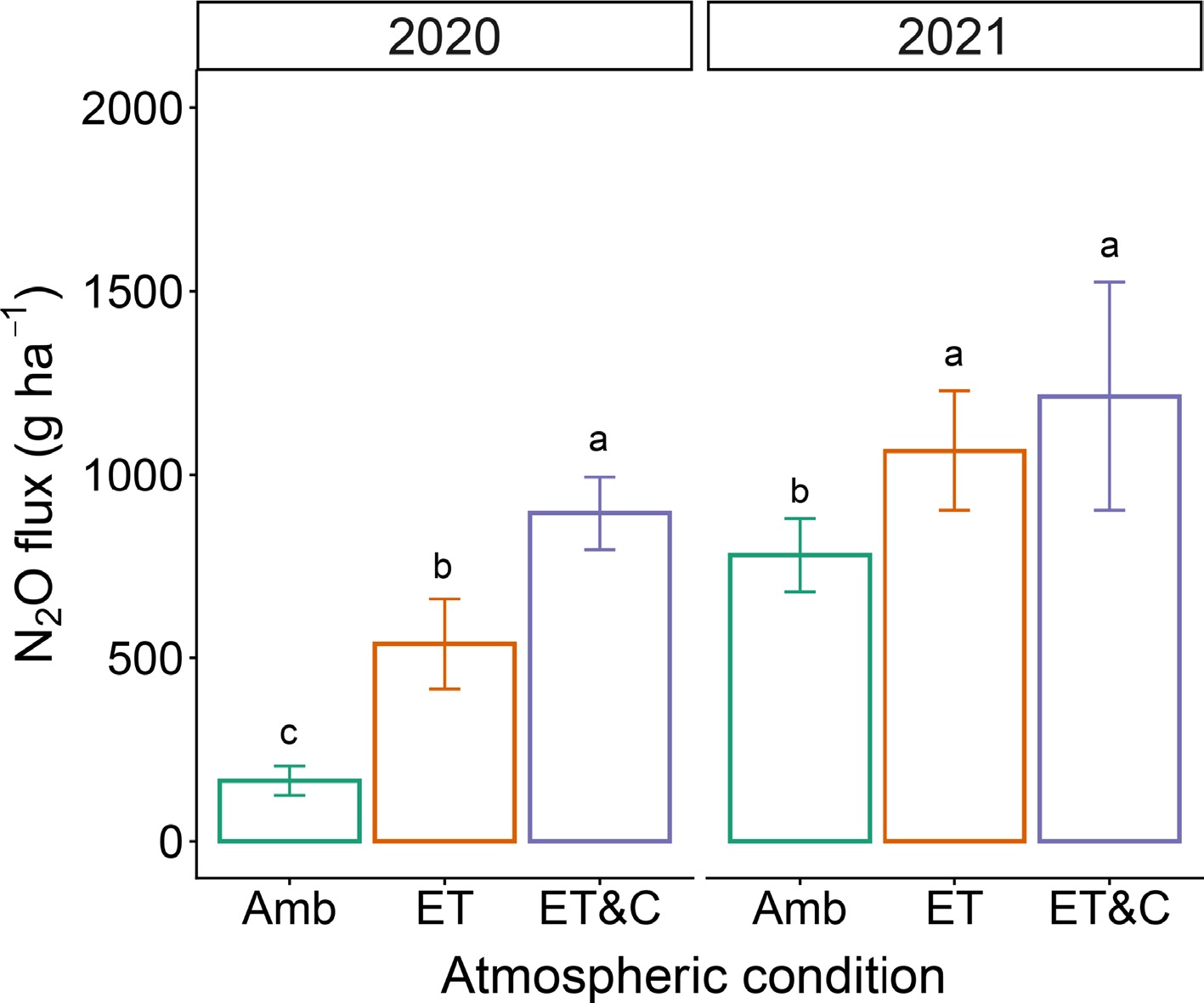
Figure 2.
Soil N2O fluxes under different atmospheric conditions for 2 years. Error bars indicate standard deviation (n = 3 within-plot subsamples). Different lowercases represent statistical difference at 0.05.
Soil N pool fractions and N isotopic values
-
Despite higher N losses via plant uptake and gaseous emissions, bulk soil N content was highest under ET&C, with ET showing comparable levels (Fig. 3; Supplementary Fig. S2). Although bulk soil δ15N values did not differ significantly among treatments, fraction-specific responses were evident. Free light particulate N increased under Amb and ET&C, indicating that elevated [CO2] stimulated particulate N formation. Occluded light N was strongly enhanced under ET and ET&C and showed marked 15N enrichment, suggesting intensified microbial processing and stabilization. Sand-associated N showed minimal treatment effects, and silt- and clay-associated N concentrations did not differ significantly among treatments; however, δ15N values were significantly elevated under ET relative to Amb.
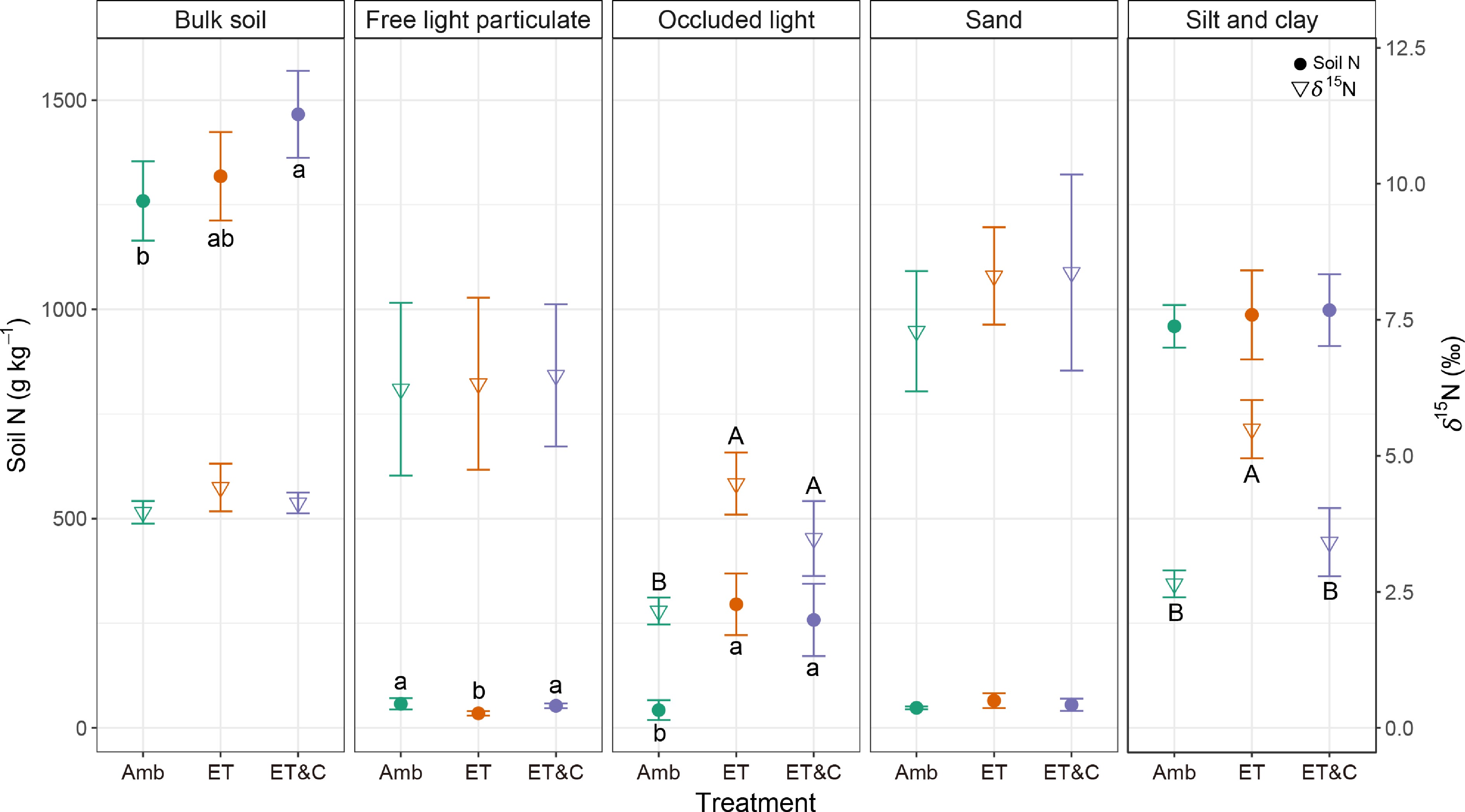
Figure 3.
Soil N content and δ15N values in each fraction under different atmospheric conditions in the 3rd year. Closed circles and open triangles represent soil N content and δ15N values, respectively. Error bars indicate standard deviation (n = 3 within-plot subsamples). Different lowercase and uppercase letters represent statistical differences at 0.05 in N content and δ15N, respectively.
Root biomass, root exudates, and N-fixing gene abundance
-
Root biomass and root exudates were not significantly affected by atmospheric treatments (Fig. 4a, b). In contrast, nifH gene abundance increased significantly under ET and ET&C relative to Amb (Fig. 4c). Similarly, the ratio of nifH to bacterial 16S rRNA gene copies was higher under ET and ET&C (Fig. 4d).
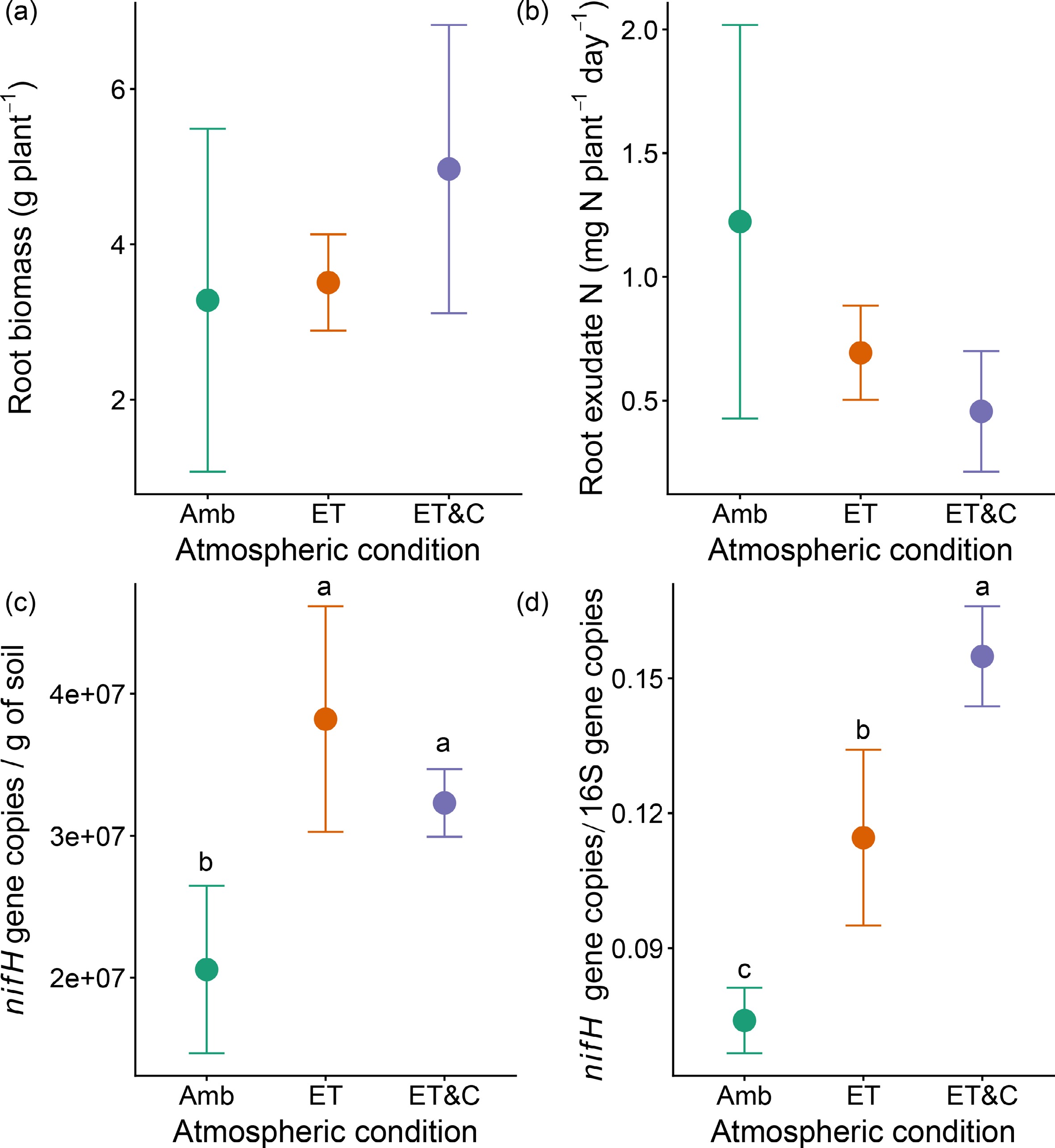
Figure 4.
(a) Root biomass, and (b) exudate N under different atmospheric conditions in the 3rd year. (c) Gene copy numbers of nifH, and (d) ratio between gene copy numbers of nifH and 16S gene under different atmospheric conditions in the 2nd year. Error bars indicate standard deviation (n = 3 within-plot subsamples). Different lowercase letters represent statistical differences at 0.05, respectively.
-
Across 2 years, warming (ET) consistently increased plant N uptake and N2O emissions relative to ambient conditions, indicating greater N availability during the growing season. Warmer conditions accelerate N mineralization and substrate supply, which enhances N uptake by rice and stimulates gaseous losses. Although NH4+ dominates under anaerobic paddy conditions and nitrification is constrained[25], elevated temperature can still enhance NH4+ supply and promote N2O production through both direct pathways and residual nitrification[24].
However, the combined treatment (ET&C) did not further increase N uptake beyond ET, even though grain and straw N contents rose. This suggests that elevated CO2 partially offsets the temperature effect on yield. Elevated CO2 is typically associated with a 'CO2 fertilization effect' in C3 crops, improving tiller numbers and grain formation through enhanced carboxylation and stomatal regulation[35,36]. Yet, co-elevation can negate these benefits by intensifying heat stress and raising canopy temperature, thereby suppressing biomass production[4,37,38].
Microbial functional potential associated with nitrogen retention under warming
-
Despite greater N outputs, bulk soil N was maintained or even increased under ET and ET&C. Because neither root biomass nor exudates differed across treatments, enhanced plant-derived N inputs are unlikely to explain this pattern. Microbial processes associated with nitrogen retention represent a plausible pathway contributing to the maintenance of soil nitrogen pools.
The nifH gene encodes the nitrogenase reductase subunit and is the standard functional marker for diazotrophic potential. A critical concern when interpreting increases in nifH copy numbers is whether the signal reflects a general expansion of total microbial biomass rather than a specific enrichment of diazotrophs. Our data directly address this: the ratio of nifH to bacterial 16S rRNA gene copies, a community-normalized index of diazotrophic relative abundance, increased significantly under ET and ET&C (Fig. 4d). This indicates that diazotrophs became more abundant relative to the broader bacterial community, not simply as a passive consequence of warming-driven microbial biomass growth. Warming likely favored diazotrophs by accelerating SOC turnover (increasing C and energy availability) and by sustaining the anoxic microsites characteristic of flooded soils that shelter oxygen-sensitive nitrogenase activity. Linking this functional shift to our isotope data provides further mechanistic support. Newly fixed N, isotopically close to atmospheric N2 (δ15N ≈ 0‰), would be expected to dilute the 15N signal in pools receiving fresh microbial inputs[39,40]. Yet we observed the opposite: δ15N enrichment was elevated in occluded particulate N and silt- and clay-associated N under ET. This apparent paradox is reconcilable when the fractionation pathway is considered. Biological N fixation products are rapidly processed through microbial biomass turnover; as microbes die and their necromass undergoes repeated cycles of mineralization and re-immobilization, isotopically lighter N is preferentially lost as NH3 or N2O, leaving 15N-enriched residues in stable organo-mineral pools[41]. The co-occurrence of elevated nifH/16S ratios and 15N enrichment in protected fractions is thus consistent with a scenario in which warming stimulates diazotrophic input that is subsequently stabilized through microbial processing[42]. We acknowledge that δ15N enrichment in mineral-associated organic matter can arise from multiple pathways and that gene abundance data cannot quantify fixation fluxes directly. Future studies coupling 15N2 tracer incubations with diazotroph community profiling will be needed to partition these contributions quantitatively.
Stabilization in mineral-associated pools under future climate
-
The fraction-resolved data reveal two mechanistically distinct pathways of N retention. Occluded particulate N, physically protected within soil aggregates, and silt- and clay-associated N, chemically stabilized through organo-mineral bonding[31], both increased or remained stable under warming, yet their δ15N responses and governing processes differ in important ways. Occluded N showed the strongest quantitative increase under ET and ET&C, accompanied by marked 15N enrichment. This fraction represents organic matter entrapped within aggregates, shielded from enzymatic access by physical occlusion rather than chemical bonding[31]. Its 15N enrichment under warming is most consistent with intensified microbial processing prior to occlusion: as fresh organic N passes through successive cycles of mineralization and microbial re-immobilization within aggregate interiors, isotopically light 14N is preferentially lost at each turnover step, and 15N-enriched microbial necromass accumulates and becomes physically entrapped—consistent with the microbial carbon pump framework[39,43]. Silt- and clay-associated N showed minimal quantitative change but pronounced 15N enrichment under ET. This dissociation suggests active turnover within what is often assumed to be a largely inert fraction. Chemical stabilization via mineral sorption preferentially retains more complex, processed organic N compounds carrying higher δ15N signatures[41,44]. Warming may accelerate replacement of older, isotopically lighter mineral-associated organic N with freshly processed, 15N-enriched microbial products without net pool accumulation. A critical alternative interpretation must be acknowledged. Kinetic isotope fractionation during N loss, denitrification, N2O production, and NH3 volatilization preferentially removes 14N, passively enriching residual soil N in 15N regardless of stabilization mechanisms[40]. Given increased N2O emissions under ET and ET&C, some portion of the observed δ15N enrichment may reflect this loss-driven effect. Our dataset cannot fully partition these two drivers. However, three lines of evidence suggest stabilization contributes independently: (i) bulk soil δ15N did not differ significantly among treatments, implying the enrichment is fraction-specific rather than a whole-soil loss signal; (ii) enrichment is strongest in the most microbially processed pools; and (iii) occluded N increased quantitatively under warming, indicating genuine accumulation. Together, these observations support a dual interpretation: warming simultaneously intensifies N loss and promotes stabilization of microbially processed N in protected pools.
Reconciling temperature–CO2 interactions
-
The combined ET&C treatment did not further increase plant N uptake beyond ET alone, and in some cases, values approached Amb levels, suggesting that elevated CO2 offset rather than amplified the warming-driven increase in N demand. Notably, rhizodeposit N did not differ significantly across atmospheric treatments (Fig. 4b), indicating that CO2-driven changes in belowground C inputs and associated microbial N immobilization are unlikely to explain this pattern. Instead, the following mechanism is proposed. Elevated CO2 reduces stomatal conductance, which decreases transpirational cooling of the rice canopy. Under the already-elevated temperatures of the ET treatment, this stomatal closure can raise canopy and panicle temperatures above the optimum for photosynthesis and spikelet fertility in japonica rice, suppressing biomass accumulation and N demand even as carboxylation efficiency per se is improved by higher atmospheric CO2. Together, these findings suggest that under combined warming and elevated CO2, the canopy-level thermal penalty of stomatal closure counteracts the photosynthetic benefit of CO2 enrichment, resulting in no net gain in plant N demand relative to ET alone[4].
While elevated temperature strengthened fast N cycling and fixation, co-elevation with CO2 (ET&C) did not always amplify responses. Elevated CO2 can increase belowground C inputs yet simultaneously intensify microbial N demand and immobilization, producing context-dependent net effects on plant uptake and gaseous loss (N2O)[45,46]. From our simplified N budget data, ET&C exhibited similar rice biomass N uptake to Amb but 10-fold higher N2O loss. Based on the data, higher bulk N content would be reported, but the trend was opposite to our expectation. The fraction-specific and isotope data and N-fixing gene abundance data suggest that under ET&C, stimulated diazotrophic activity introduced more N retention in soil and a larger portion of a microbially processed N became stabilized in soil aggregates, partially decoupling bulk N retention from immediate plant acquisition. This helps explain why bulk soil N remained high under ET&C even when plant uptake was similar to Amb.
Implications for climate-smart rice management
-
Our results suggest that warming promotes microbial pathways that enhance nitrogen retention and stabilization in protected soil pools, thereby moderating the sensitivity of soil nitrogen stocks to increased turnover under climate change. Practically, this mechanism could: (i) sustain soil fertility under warming; (ii) reduce dependence on mineral N fertilizer where fixation is enhanced; and (iii) lower vulnerability to N leaching by favoring protected pools. Management strategies that support diazotrophs and mineral association—e.g., maintaining moderate organic C inputs, optimizing flood regimes to preserve anoxic microsites, and avoiding practices that disrupt aggregation—may increase N-use efficiency. For policy and sustainability framing, these findings suggest that leveraging biological N inputs is a viable key to climate-resilient rice cultivation, especially in low-fertility paddies where warming pressure on N cycling is rising.
Limitations and generalizability
-
A limitation of this study is the low number of experimental plots per treatment (n = 2), which reflects the substantial costs of constructing and maintaining open-top chamber infrastructure across multiple growing seasons rather than a deliberate methodological choice. Statistical analyses were conducted on within-plot subsamples (n = 3 per treatment), which reflect within-plot rather than between-plot variance; all findings should therefore be interpreted as indicative rather than conclusive. The small plot-level replication limits our ability to fully distinguish treatment effects from plot-specific spatial variation[4]. We therefore interpret our results as indicative rather than conclusive and encourage replication across additional sites and soil types to confirm the observed patterns.
We acknowledge that attributing δ15N enrichment in mineral-associated fractions to microbial processing and stabilization is an inference based on isotope patterns and is not directly confirmed by measurements of microbial community composition or enzyme activity in this study. This interpretation is consistent with established frameworks linking microbial necromass to organo-mineral associations, but direct supporting evidence such as phospholipid fatty acid analysis, microbial community profiling, or extracellular enzyme assays was not collected here. Future studies should integrate these measurements alongside isotope fractionation analysis to mechanistically confirm the stabilization pathways hypothesized here.
Open-top chambers imitate field conditions while controlling climate forcings, but lateral exchanges and landscape heterogeneity are constrained. We focused on nifH as a functional marker; future work should couple process-rate measurements of N2 fixation (e.g., 15N2 assimilation) with diazotroph community profiling to resolve taxa and pathways. Finally, testing across soil textures, flood regimes, and cultivar × management combinations will refine the boundary conditions under which fixation-driven buffering dominates.
-
Warming appeared to intensify N cycling in flooded rice soils, with results suggesting increased plant N uptake and elevated N2O emissions under elevated temperature conditions. Despite these higher N outputs, bulk soil N stocks were maintained or increased under warming treatments, a pattern consistent with redistribution of N among soil pools rather than a straightforward imbalance between inputs and losses, though the mechanisms driving this redistribution require further investigation. Fraction-specific analyses indicated that warming may promote the retention of N in occluded particulate and mineral-associated pools, accompanied by δ15N enrichment patterns consistent with intensified microbial processing and stabilization, although multiple processes, including N loss fractionation, may contribute to the observed isotopic signatures. The observed increase in nifH gene abundance under warming and combined warming–CO2 conditions suggests a shift in microbial diazotrophic potential; however, as nifH abundance does not directly quantify biological N fixation rates, this finding should be interpreted as indicative of changing functional genetic potential rather than confirmed fixation activity. Collectively, these results suggest that climate warming may reshape N partitioning and stabilization pathways in flooded rice systems, with N redistribution toward more protected soil pools as a potential outcome, a pattern that warrants replication across contrasting soil types and climate zones before broader conclusions can be drawn.
We acknowledge the support of the National Research Foundation for this study.
-
It accompanies this paper at: https://doi.org/10.48130/nc-0026-0010.
-
Not applicable.
-
The authors confirm contributions to the paper as follows: Hyeon Ji Song: data collection, analysis and interpretation of results, and draft manuscript preparation; Young Ho Seo: data collection, analysis and interpretation of results; Ho Jun Shin: data collection, analysis and interpretation of results; Jae Jin Lee: analysis and interpretation of results and draft manuscript preparation; Snowie Jane G. Ampode: draft manuscript preparation; Pil Joo Kim: study conception and design and draft manuscript preparation. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are not publicly available as they are part of ongoing research but are available from the corresponding author upon reasonable request.
-
This work was supported by the National Research Foundation, Ministry of Education, Korea (NRF-2023R1A2C3004842). Hyeon Ji Song, Young Ho Seo, Ho Jun Shin, and Snowie Jane C. Ampode were supported by scholarships from the BK21+ program of the Ministry of Education and Human Resources Development, Korea.
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
Full list of author information is available at the end of the article.
- The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Song HJ, Seo YH, Shin HJ, Lee JJ, Ampode SJG, et al. 2026. Warming reshapes nitrogen partitioning and stabilization pathways in flooded rice soils. Nitrogen Cycling 2: e023 doi: 10.48130/nc-0026-0010 

Warming reshapes nitrogen partitioning and stabilization pathways in flooded rice soils
- Received: 20 March 2026
- Revised: 08 May 2026
- Accepted: 03 June 2026
- Published online: 30 June 2026
Abstract: Climate change, through elevated temperature and atmospheric carbon dioxide (CO2) concentration, is expected to alter soil quality, which is directly linked to crop productivity. While the effects of climate change on soil carbon (C) dynamics have been extensively studied across various agroecosystems, shifts in nitrogen (N), an essential nutrient for crops, have received far less attention. In this study, we conducted a 3-year field experiment using three types of open-top chambers to simulate present conditions (Amb), elevated temperature (+2 °C; ET), and combined elevated temperature and CO2 (+2 °C and +200 ppm; ET&C) in a rice cultivation system. We examined N dynamics by evaluating inputs and losses, as well as changes in soil N pools, integrating 15N isotopic analysis. In terms of N output, rice N uptake (grain + straw) and N2O emissions increased significantly under ET compared with Amb. Interestingly, the response of each soil N pool fraction varied across treatments: the free light fraction, representing particulate N, showed no significant difference, whereas aggregate-associated N pools (occluded N), relatively stable forms of N, increased substantially under both future climate conditions (ET and ET&C). Correspondingly, greater 15N enrichment in mineral-associated N (occluded N and silt and clay-associated N) under ET indicated enhanced microbial processing and stabilization under warming conditions. Although root biomass and exudate inputs did not differ significantly among treatments, the abundance of the nifH gene, associated with microbial N fixation, increased markedly under ET and ET&C. Together, these results show that warming reshapes soil N partitioning by promoting retention of N in protected pools, partially offsetting increased N losses. While nifH gene abundance does not directly quantify N2 fixation rates, its increase is consistent with altered microbial N-fixing potential under warming. Our findings highlight how climate warming can modify pathways of soil N retention and stabilization in flooded rice systems, with implications for soil fertility and N management under future climate conditions.
-
Key words:
- Nitrogen cycle /
- Climate change /
- Open top chamber /
- Nitrogen isotopic signature