









-
The APETALA2/ETHYLENE-RESPONSIVE FACTOR (AP2/ERF) gene family represents one of the largest and most functionally diverse transcription factors, orchestrating a wide range of biological processes throughout the plant life cycle[1,2]. These factors are defined by a highly conserved 60–70 amino acid AP2/ERF DNA-binding domain, which directly recognizes specific cis-regulatory elements, enabling precise spatiotemporal control of gene expression. Based on the number of AP2/ERF domains and overall sequence similarity, the family is systematically divided into several subfamilies: the AP2 subfamily (containing two AP2 domains), the ERF and DEHYDRATION-RESPONSIVE ELEMENT BINDING (DREB) subfamilies (each with a single AP2 domain), the RELATED TO ABI3/VP1 (RAV) subfamily (possessing both an AP2 and a B3 domain), and the distinct Soloist group[3−6]. AP2 genes have been identified across numerous plant species, including Arabidopsis (Arabidopsis thaliana), rice (Oryza sativa), wheat (Triticum aestivum), strawberry (Fragaria vesca), grape (Vitis vinifera), and exhibit high evolutionary conservation[7−11]. The AP2 subfamily is further subdivided into the euAP2, eu-AINTEGUMENTA (euANT), and basalANT subgroups based on variations in the amino acid sequence and nuclear localization signal within its double AP2 domain, with each subgroup associated with distinct functional roles[12−14].
The AP2/ERF transcription factor family plays a dual role in plant growth, development, and stress adaptation, with its members governing processes ranging from floral organogenesis, meristem maintenance, and secondary metabolite biosynthesis, to biotic and abiotic stress responses[15−22]. Within this family, the ERF and AP2 subfamilies represent the two largest lineages yet exhibit markedly distinct functional trajectories: ERF subfamily members (including DREB) are primarily involved in stress responses, whereas AP2 subfamily members are mainly associated with developmental regulation. For instance, MdERF38 modulates drought-induced anthocyanin biosynthesis in apple (Malus × domestica)[23], while overexpression of PtoERF15 enhances drought tolerance in poplar (Populus tomentosa) by maintaining stem water potential[24]. Similarly, natural allelic variation in the DREB transcription factor gene TaDTG6-B confers enhanced drought tolerance in wheat[25]. Conversely, other members may play negative roles; overexpression of the pearl millet (Pennisetum glaucum) gene PgRAV_01 was found to negatively affect drought tolerance in tobacco by impairing antioxidant capacity and osmotic adjustment[26]. In contrast to the well-characterized roles of the ERF, DREB, and RAV subfamilies in stress signaling, most functionally characterized AP2 subfamily members in plants are involved in developmental regulation, particularly floral organogenesis[15,16,21]. The direct role and molecular mechanisms of the AP2 subfamily in stress adaptation, particularly drought tolerance, remain largely unexplored.
Methyl jasmonate (MeJA), the active form of jasmonic acid (JA), is a key phytohormone that regulates defense responses, plant development, and secondary metabolite biosynthesis[27−32]. In recent years, its function in regulating drought tolerance has drawn increasing attention. Transcription factors belonging to the AP2/ERF family mediate diverse plant biological processes via the JA signaling pathway. For example, in sugarcane (Saccharum officinarum), ScAIL1 enhances plant disease resistance by activating JA biosynthesis, thereby balancing defense responses and growth through the JA signaling cascade[33]. In poplar, PtoERF15 confers drought tolerance by activating PtoMYC2b, a core component of the MeJA signaling pathway, to coordinate stress-responsive xylem development[24]. Additionally, in Cymbidium faberi, the MeJA-inducible transcription factor CfMYC2-like directly interacts with and represses CfRAP2.4, an AP2-family activator, thereby suppressing floral MeJA biosynthesis and fine-tuning scent production between vegetative and flowering tissues[34]. Given the critical role of AP2 transcription factors in MeJA-mediated physiological processes, identifying MeJA-responsive AP2 genes and deciphering their regulatory mechanisms in MeJA-induced drought tolerance are of paramount importance for advancing targeted breeding of drought-resistant crop varieties and plant germplasm.
Blueberry (Vaccinium spp.), a perennial shrub of high economic and nutritional value, is renowned for its flavonoid-rich fruits[35,36]. However, as a shallow-rooted species with a relatively underdeveloped root system, it is particularly vulnerable to environmental stresses such as winter temperature fluctuations, late-spring frosts, and seasonal drought[37]. Recent molecular studies have begun to elucidate the genetic basis of blueberry drought tolerance, revealing a core regulatory network that integrates flavonoid biosynthesis and plant hormone signaling[38]. Genome-wide analyses have further identified multiple stress-responsive transcription factor families (e.g., MYB, WRKY, NF-YA, DELLA), some of which regulate ROS or exhibit drought-responsive expression[39−42]. For instance, while VcTT2 (an MYB transcription factor) enhances drought tolerance via antioxidant activation[43], VcMYB4a increases stress sensitivity, illustrating the divergent roles within a single family[44]. Root-specific studies further highlight genes in carbon metabolism and terpenoid biosynthesis that shape drought-responsive metabolites[45].
Despite these advances, the role of the AP2 subfamily in blueberry drought tolerance remains unclear. In this study, we conducted a genome-wide identification of 32 VcAP2 genes (VcAP2-1 to VcAP2-32) in blueberry using published 'Draper' genomic data. VcAP2-20 was highly induced by MeJA and drought stress. Overexpression of VcAP2-20 enhanced drought tolerance in blueberry calli and seedlings by reducing ROS accumulation, MDA content, and electrolyte leakage, while increasing proline content and antioxidant enzyme activities. VcAP2-20 positively regulated drought tolerance by upregulating drought-responsive genes (e.g., VcSOD1, VcPODs, and VcAPX1). Our findings establish a foundational understanding of the role of VcAP2-20 in drought adaptation, providing a theoretical basis for further exploration of its molecular mechanisms.
-
Thirty-day-old seedlings of the northern highbush blueberry (Vaccinium corymbosum L. cv. 'Legacy') were cultivated in a controlled greenhouse environment under a 16 h light/8 h dark photoperiod at 24 ± 1 °C with 60%−70% relative humidity. The seedling growth substrate was a mixture of nutrient soil, vermiculite, and perlite in equal volume proportions. The substrate was regularly irrigated with 0.1% (w/v) ferrous sulfate solution to stably maintain the rhizosphere pH within the range of 5.2–5.5. To induce drought stress, seedlings were treated with a 10% (w/v) polyethylene glycol 6000 (PEG6000) solution for 0, 3, and 12 h. For root treatment, each pot was irrigated with 50 mL of 10% PEG6000 solution, with clean water irrigation as the control; for leaf treatment, a 10% PEG6000 solution was evenly sprayed on the leaf surface until liquid dripped, with clean water spraying as the synchronous control. Following each treatment duration, leaves and roots were rapidly harvested, immediately frozen in liquid nitrogen, and stored at −80 °C for subsequent RNA extraction.
Blueberry 'Northland' calli for genetic transformation were cultured on Woody Plant Medium (WPM) supplemented with 1.0 mg·L−1 2,4-dichlorophenoxyacetic acid (2,4-D) and 0.4 mg·L−1 6-benzylaminopurine (6-BA) at 24 °C in darkness and subcultured every month. Apple 'Orin' calli for transient transformation were cultured on Murashige and Skoog (MS) medium supplemented with 1.5 mg·L−1 2,4-D and 0.4 mg·L−1 6-BA at 24 °C in darkness and subcultured every 20 d.
Blueberry 'Legacy' tissue-cultured seedlings used for genetic transformation were cultured in WPM medium containing 1.0 mg·L−1 ZT in a controlled greenhouse environment under a 16 h light/8 h dark photoperiod at 24 ± 1 °C with 60%−70% relative humidity.
Identification of AP2 subfamily genes in blueberry
-
The genome sequence of the highbush blueberry cv. 'Draper' was downloaded from the Vaccinium Genome Database (GDV) (
www.vaccinium.org )[46]. The amino acid sequences of AP2 subfamily proteins in Arabidopsis were downloaded from the TAIR website (www.arabidopsis.org ) and used as a query to perform blastp alignment (E-value < 1e−5) in the TBtools software (v2.3.3.1)[47]. The Hidden Markov Model (HMM) for AP2 (PF00847) was downloaded from Pfam (http://pfam.xfam.org ) and used to determine the presence of the AP2 domain. Furthermore, all candidate sequences were checked by the NCBI-CDD website (www.ncbi.nlm.nih.gov/cdd ), SMART website (https://smart.embl.de ), CD-HIT website (www.bioinformatics.org/cd-hit ), and DNAMAN software to remove sequences without two AP2 domains and redundant sequences.Bioinformatics analysis
-
To assess the physicochemical properties of VcAP2 subfamily members, the ExPASy ProtParam website (
https://web.expasy.org/protparam ) was employed. Subcellular localization of VcAP2 proteins was predicted using the WoLF PSORT website (https://wolfpsort.hgc.jp ). Conserved domains were predicted by the Batch web CD-Search tools (www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) and visualized by TBtools software (v2.3.3.1).For phylogenetic analysis, the amino acid sequences of 14 AtAP2 proteins from Arabidopsis and 32 VcAP2 proteins from blueberry were aligned using CLUSTALW in MEGA 7.0 under default parameters. The resulting multiple sequence alignment was visualized using the ESPript online tool (
https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi ). A phylogenetic tree was then constructed based on the alignment using the neighbor-joining method with 1,000 bootstrap replicates, and subsequently visualized and annotated using the Evolview website (www.evolgenius.info/evolview ).To investigate regulatory elements, the 2,000 bp upstream sequences from the initiation codon were retrieved as promoter sequences, and then submitted to the PlantCARE database (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) to predict the cis-acting regulatory elements. The predicted results were counted in Excel and visualized in TBtools software (v2.3.3.1).GUS staining and activity analysis
-
The promoter sequence of VcAP2-20 was cloned and inserted into the pBI121-GUS vector to generate a proVcAP2-20:GUS transcriptional fusion construct. This construct was subsequently transformed into blueberry calli and apple calli via Agrobacterium tumefaciens-mediated transient transformation as previously described[23,48]. Following transformation, excess bacterial suspension was removed from the calli, which were then plated onto medium containing 100 μM MeJA, abscisic acid (ABA), or salicylic acid (SA), while calli cultured on medium without hormone served as the control. After 48 h of incubation in darkness, the blueberry and apple calli were stained using a GUS staining kit (Hua Yue Yang), respectively. GUS activity was quantitatively measured with a GUS reporter gene quantitative detection kit (Coolaber) according to the manufacturer's instructions. All primers used in the experiment are listed in Supplementary Table S1.
Subcellular localization
-
The coding sequence (CDS) of VcAP2-20 was cloned and inserted into the pBI121-eGFP vector to generate a 35S:VcAP2-20-eGFP fusion construct. This construct was mixed in equal proportion with an Agrobacterium tumefaciens suspension carrying the nuclear localization AtWRKY40-mCherry, and the mixture was infiltrated into epidermal cells of tobacco (Nicotiana benthamiana). Fluorescence was detected using a laser scanning confocal microscope (Leica TCS SP8). All primers used in this experiment are listed in Supplementary Table S1.
RNA extraction and Quantitative real-time PCR (RT-qPCR)
-
Total RNA was extracted using the HiPure Plant RNA Kits (Magen, China). The first-strand cDNA was synthesized using the 5× All-In-One RT MasterMix (Abm, China). An RT-qPCR assay was conducted using the PowerUp™ SYBR™ Green Premix (Applied Biosystems) on an ABI StepOnePlus™ Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). VcUBC28 was used as the internal reference gene for normalization[49,50]. The 2−ΔΔCᴛ method was used to calculate the relative expression. All primers used in the experiment are listed in Supplementary Table S1.
Generation of transgenic blueberry calli and seedlings
-
The 35S:VcAP2-20-eGFP recombinant vector was infiltrated into blueberry 'Northland' calli to obtain VcAP2-20-OE blueberry calli using the method described previously[48]. To construct 35S:VcAP2-20-RNAi recombinant vector, the sense and antisense of 320 bp gene-specific sequence of VcAP2-20 were amplified and inserted into the modified pRI101 vector, respectively. VcAP2-20-OE and VcAP2-20-RNAi transgenic seedlings were obtained using the Agrobacterium-mediated leaf disc transformation method[51,52]. All primers used in the experiment are listed in Supplementary Table S1.
Drought tolerance testing of transgenic materials
-
Wild-type (WT) and transgenic blueberry materials, including calli and seedlings, were used as experimental subjects to evaluate their drought tolerance. For the calli-level test, WT and VcAP2-20-OE calli with robust growth and soft texture after 25 d of subculture were selected as materials. Calli samples of 0.1 g were accurately weighed and placed on subculture medium supplemented with 5% and 10% PEG6000, respectively, then incubated in the dark at 25 °C for 10 d. WT and VcAP2-20-OE calli cultured on standard subculture medium without PEG6000 were set as controls. For the whole-plant level test, 6-month-old WT and transgenic seedlings (including VcAP2-20-OE and VcAP2-20-RNAi lines) were subjected to natural dehydration treatment for 25 d. After the treatment, phenotypic changes of all tested seedlings were observed, and leaf samples were collected for subsequent physiological analysis.
To detect superoxide anions, calli from WT and transgenic lines under both normal and drought-stress conditions were stained with 3,3'-Nitrotetrazolium blue chloride (NBT) solution (Coolaber, China) for 20 min. The experiment was conducted with three independent biological replicates, and each biological replicate included three technical replicates.
For physiological measurements, calli and leaves from WT and transgenic seedlings were harvested after drought treatment, snap-frozen in liquid nitrogen, and ground to a fine powder. A 0.1 g aliquot of the powdered sample was used to determine relative electrical conductivity, hydrogen peroxide (H2O2), malondialdehyde (MDA), and proline content, as well as the activities of peroxidase (POD), superoxide dismutase (SOD), and ascorbate peroxidase (APX), following previously reported methods[43,53]. All experiments were performed with three independent biological replicates, each with three technical replicates.
Statistical analysis
-
Experimental data are presented as mean ± standard deviation (SD) from at least three independent biological replicates. Statistical analyses were performed using IBM SPSS Statistics 27.0.1. Differences between the two groups were analyzed by Student's t-test, with significance levels denoted as * p < 0.05, ** p < 0.01, *** p < 0.001.
-
The AP2 subfamily is characterized by two AP2 domains, as reported for all 14 members in Arabidopsis[1,7]. In this study, we initially identified 52 candidate AP2 subfamily members in the blueberry genome using Hidden Markov Model (HMM) and blastp analyses. After removing redundant sequences, 32 non-redundant proteins containing two AP2 domains were retained and designated VcAP2-1 to VcAP2-32 (Fig. 1a). Their physicochemical properties were predicted using ExPASy ProtParam. These proteins ranged in length from 259 (VcAP2-28) to 721 (VcAP2-18) amino acids, with corresponding molecular weights between 29,725.22 Da and 78,973.23 Da. Theoretical isoelectric point (pI) values varied from 4.8 (VcAP2-3) to 9.74 (VcAP2-28), classifying 20 proteins as acidic (pI < 7) and 12 as basic (pI > 7). According to the instability index, with a threshold of 40 used to distinguish stable (< 40) from unstable (> 40) proteins, 29 VcAP2 proteins were classified as stable and three as unstable. The aliphatic index ranged from 50.14 (VcAP2-20) to 79.92 (VcAP2-28). All VcAP2 proteins exhibited negative grand average of hydropathicity (GRAVY) values, indicating hydrophilic character and suggesting favorable thermal stability. Subcellular localization predictions using WoLF PSORT indicated that all 32 VcAP2 proteins are predicted to be localized to the nucleus, which is fully consistent with the functional properties of AP2 transcription factors (Supplementary Table S2). Among them, 11 VcAP2 proteins are predicted to be exclusively localized to the nucleus, while the remaining 21 VcAP2 proteins have a multi-compartment localization pattern, with the nucleus as one of their core localization sites and additional predicted distribution in other cellular compartments (Supplementary Table S2).
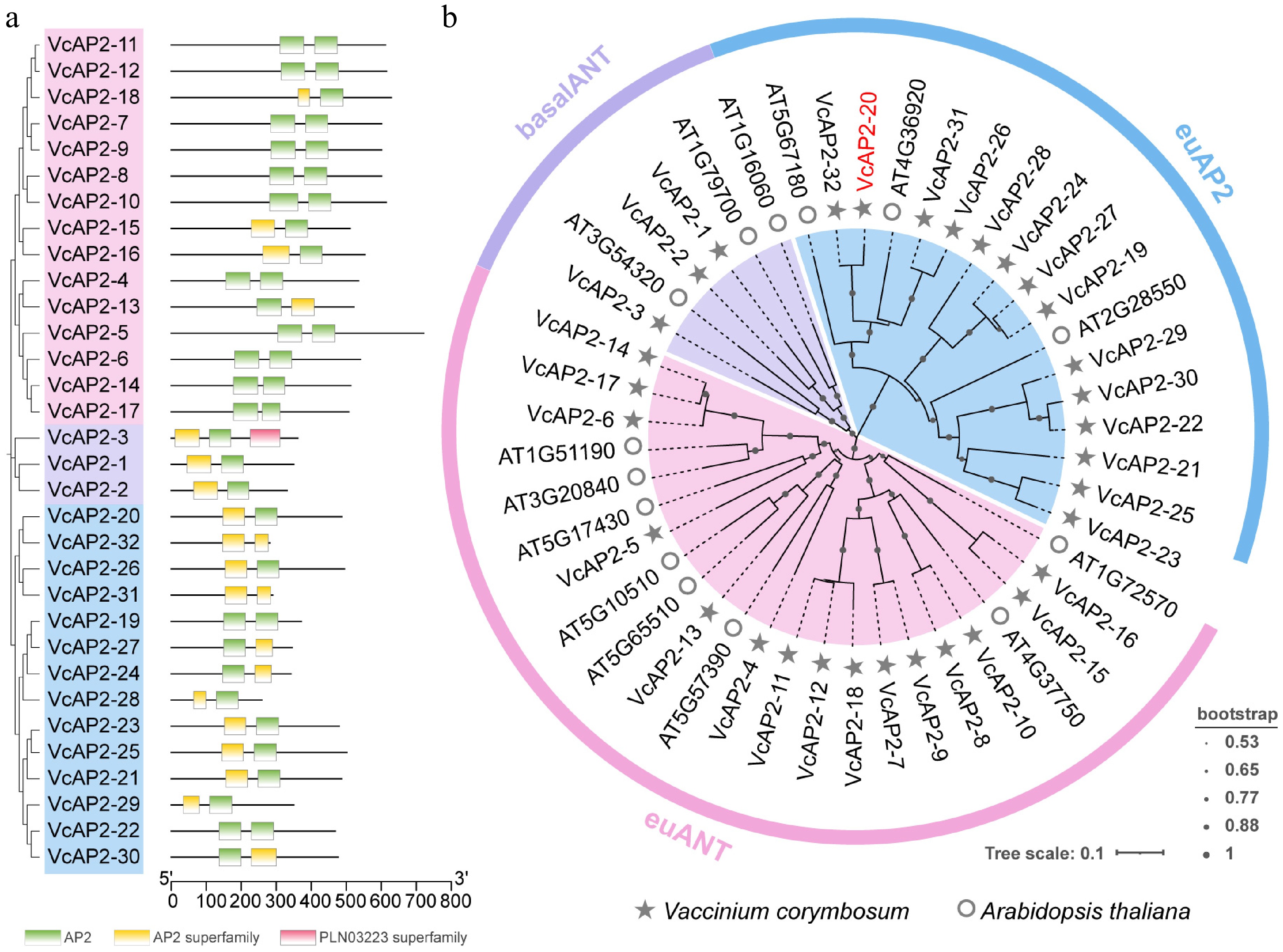
Figure 1.
Genome-wide identification of VcAP2 subfamily members in blueberry. (a) Conserved domains of the VcAP2 subfamily members in blueberry. The AP2 subfamily proteins were clustered into three subgroups: euAP2 (blue), euANT (pink), and basalANT (purple). (b) Phylogenetic analysis of the AP2 proteins from blueberry (Vaccinium corymbosum) and Arabidopsis (Arabidopsis thaliana). Proteins from blueberry and Arabidopsis are marked by a star, and a circle, respectively. VcAP2-20 belongs to the euAP2 subgroup.
To elucidate the evolutionary relationships among the VcAP2 proteins in blueberry and their homologs in the model plant Arabidopsis, a phylogenetic tree was constructed using 32 VcAP2 proteins from blueberry and 14 AtAP2 proteins from Arabidopsis. The resulting phylogeny classified the VcAP2 proteins into three distinct subgroups, euAP2, euANT, and basalANT, consistent with the subgroup organization previously established in Arabidopsis. Among these, the euANT subgroup contained the highest number of members (15), followed by euAP2 (14), while the basalANT subgroup comprised only three members (Fig. 1b, Supplementary Fig. S1). Notably, VcAP2-26 and VcAP2-31 were most closely related to AtAP2 (AT4G36920), whereas VcAP2-8 and VcAP2-10 showed the highest sequence similarity to AtANT (AT4G37750), suggesting potential functional conservation between these orthologous pairs (Fig. 1b).
Analysis of cis-acting elements in the promoter of blueberry VcAP2 genes
-
Cis-acting elements in plant promoters can bind to transcription factors, thereby activating genes involved in plant development, hormone regulation, and stress responses. To identify potential regulatory elements, we analyzed the promoters of all 32 VcAP2 genes using PlantCARE. The predicted elements were classified into five functional categories: core elements, light-responsive elements, development-related elements, hormone-responsive elements, and stress-responsive elements. All 32 VcAP2 promoters harbored numerous core promoter elements (e.g., CAAT-box and TATA-box). Among light-responsive elements, the G-box and Box4 were the most widespread. With the exception of VcAP2-9, every promoter contained multiple hormone-responsive elements. These included motifs responsive to abscisic acid (ABA)-responsive elements (ABRE), auxin-responsive elements (AuxRR-core and TGA-element), gibberellin (GA)-responsive elements (GARE-motif, P-box, and TATC-box), methyl jasmonate (MeJA)-responsive elements (TGACG-motif and CGTCA-motif), and salicylic acid (SA)-responsive elements (TCA-element). ABA-responsive elements were the most abundant hormone-related motifs; interestingly, the VcAP2-14 promoter contained ABREs exclusively. Many promoters also harbored elements responsive to MeJA or auxin.
Furthermore, we identified several abiotic stress-responsive elements, including drought and salt-inducible elements (DRE-core, MBS, MYC, etc.), low-temperature-responsive elements (LTR), and oxidative stress-responsive elements (ARE and GC-motif), as well as biotic stress-related elements such as W-box and WUN-motif. The widespread presence of abiotic stress-related elements in multiple VcAP2 promoters implies a potential role for these genes in mediating abiotic stress adaptation (Fig. 2).
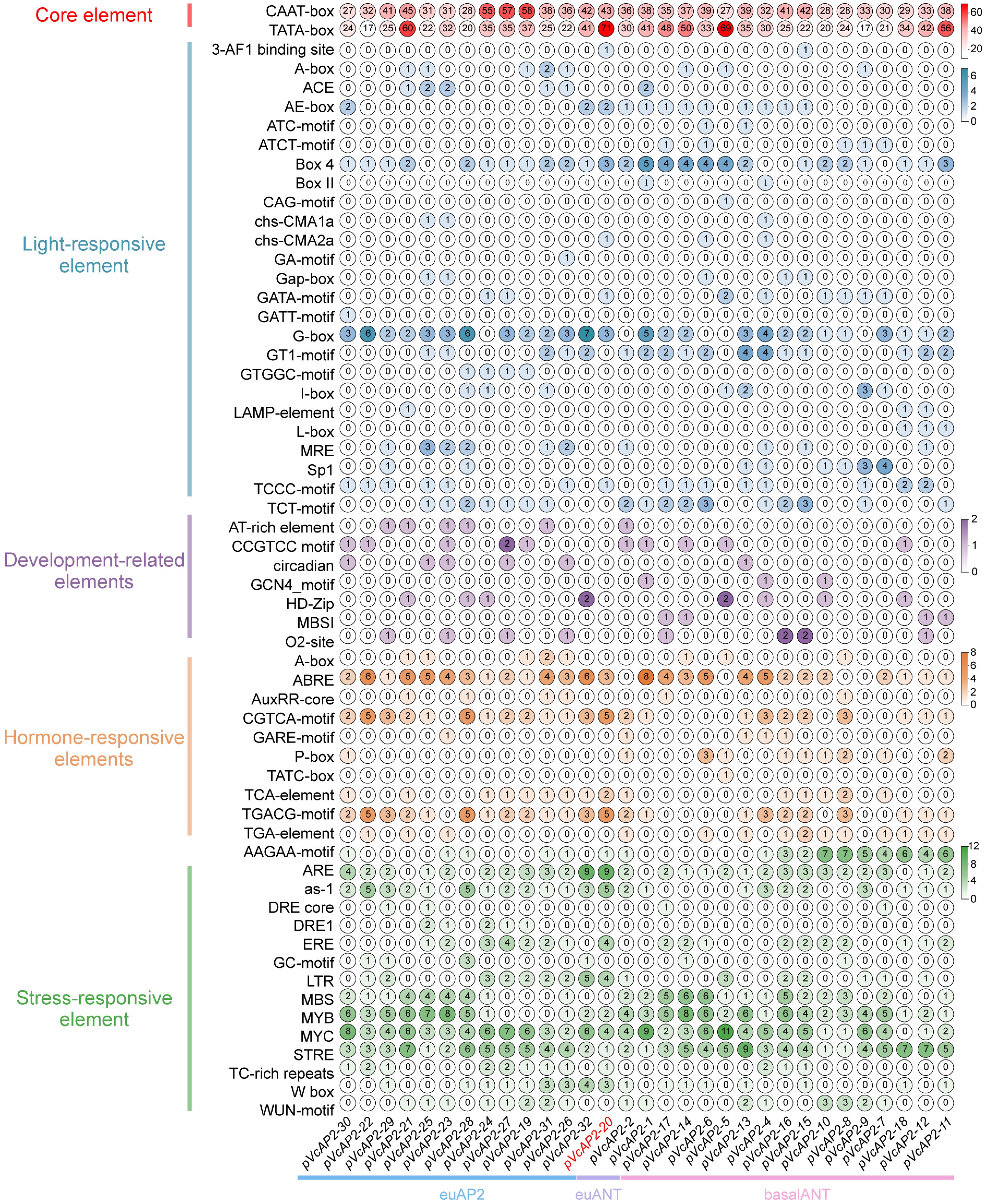
Figure 2.
Cis-acting elements analysis in the promoters of VcAP2 subfamily members. Promoter sequences were analyzed using the PlantCARE database to predict cis-acting regulatory elements, which were categorized into five functional types: core elements (red), light-responsive elements (blue), development-related elements (purple), hormone-responsive elements (orange), and stress-responsive elements (green). The numbers within the circle indicate the count of each element in each VcAP2 promoter.
VcAP2 is an MeJA- and drought-responsive transcription factor
-
Based on the presence of MeJA-responsive elements (TGACG-motif and CGTCA-motif) in the promoters of multiple VcAP2 genes, we analyzed published transcriptome data to examine their expression patterns under MeJA treatment[46]. The resulting heatmap revealed that the expression of VcAP2-20 was specifically upregulated by MeJA, peaking at 12 h post-treatment with the highest induction level among all VcAP2 genes (Fig. 3a). To investigate whether this transcriptional response is mediated by promoter activity, we cloned the VcAP2-20 promoter and fused it to the GUS reporter gene in the pBI121 vector. The proVcAP2-20:GUS construct was transformed into blueberry calli. Following treatment with 100 μM MeJA, GUS staining showed that calli harboring proVcAP2-20:GUS exhibited markedly darker blue staining compared to the untreated control. Wild-type (WT) calli appeared light yellow, while those transformed with a 35S:GUS construct (positive control) stained dark blue (Fig. 3b). Quantitative measurement of GUS activity confirmed that MeJA treatment significantly enhanced the activity of the VcAP2-20 promoter (Fig. 3c). Since the VcAP2-20 promoter also contains predicted SA- and ABA-responsive elements, we sought to determine whether these elements are also functional. To do so, we further treated proVcAP2-20:GUS apple calli with 100 μM MeJA, 100 μM SA, or 100 μM ABA. Although SA and ABA treatments slightly intensified GUS staining compared to the control, the effect was considerably weaker than that induced by MeJA (Supplementary Fig. S2), consistent with the predominant role of MeJA in regulating VcAP2-20 expression. Taken together, these results demonstrate that MeJA activates VcAP2-20 expression via its promoter.
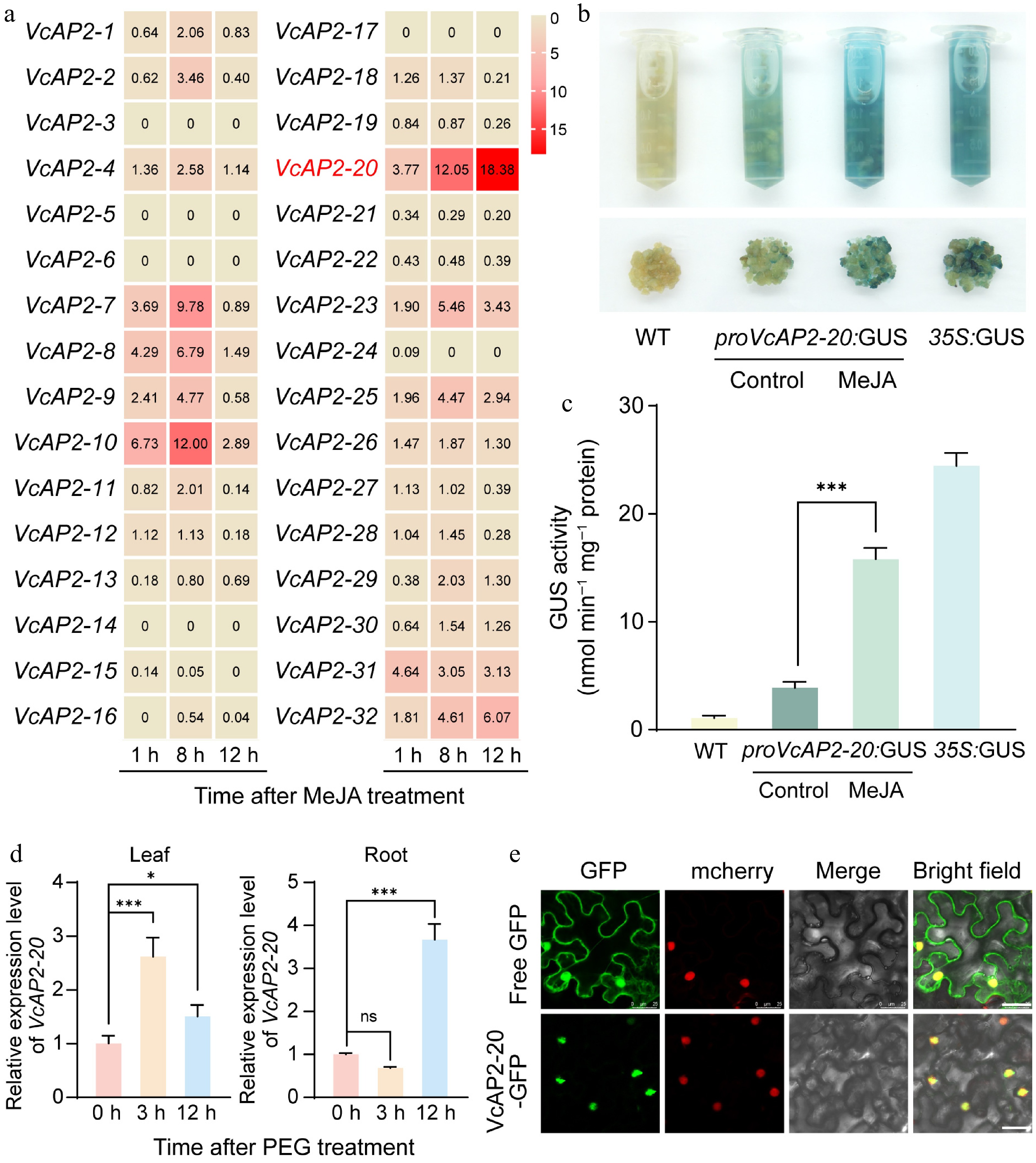
Figure 3.
VcAP2-20 expression is induced by methyl jasmonate (MeJA) treatment and drought stress. (a) Heatmap showing the expression profiles of VcAP2s in blueberry leaves under MeJA treatment. Transcriptome data were downloaded from GigaDB Dataset (
https://gigadb.org/dataset/100537 ). (b) Confirmation of 100 μM MeJA treatment enhanced the promoter activity of VcAP2-20 using the GUS staining in blueberry calli. Calli grown under normal conditions served as the control. (c) Bar charts showing the quantification of the GUS activity in (b). (d) RT-qPCR analysis of relative expression levels of VcAP2-20 in blueberry leaves and roots under drought stress simulated by 10% PEG6000. Values are presented as mean ± SD from three biological replicates. Statistical significance was determined by Student's t-test: * p < 0.05, ** p < 0.01, *** p < 0.001. (e) Subcellular localization of the 35S:VcAP2-20-GFP fusion protein in tobacco (Nicotiana benthamiana) leaf epidermal cells. The WRKY40-mCherry fusion protein served as a nuclear localization marker, while free GFP was used as the control. Scale bar = 25 μm.Given that MeJA is a key signaling molecule in plants' drought responses, we hypothesized that VcAP2-20 might also be involved in drought stress adaptation. To test this, we subjected 30-day-old blueberry seedlings to drought stress simulated by watering with 10% PEG6000. We analyzed VcAP2-20 expression in leaves and roots at 0, 3, and 12 h after PEG treatment. The results revealed that VcAP2-20 expression in leaves peaked at 3 h post-treatment, whereas in roots, it reached a maximum at 12 h (Fig. 3d), indicating that drought stress induces VcAP2-20 expression in a tissue-specific manner.
To determine the subcellular localization of VcAP2-20, its coding sequence was cloned into the pBI121 vector, generating a 35S:VcAP2-20-eGFP fusion construct. This construct, along with an mCherry-labelled nuclear localization marker (AtWRKY40-mCherry), was transiently co-transfected into tobacco epidermal cells. Confocal microscopy revealed that the eGFP signal completely overlapped with the mCherry signal, demonstrating that VcAP2-20 is a nuclear protein (Fig. 3e).
VcAP2-20 enhances drought tolerance in blueberry calli
-
Given that VcAP2-20 expression was upregulated by both MeJA and drought treatments, we hypothesized that this gene contributes to drought tolerance in blueberry. To investigate its role, we generated overexpression lines by transforming wild-type (WT) blueberry calli with a 35S:VcAP2-20-eGFP construct. RT-qPCR confirmed that VcAP2-20 transcript levels were approximately 8-fold higher in the transgenic calli than in the WT (Fig. 4b). Furthermore, the detection of eGFP signals by stereo fluorescence microscopy confirmed that the VcAP2-20-OE calli were transgenic-positive (Supplementary Fig. S3).
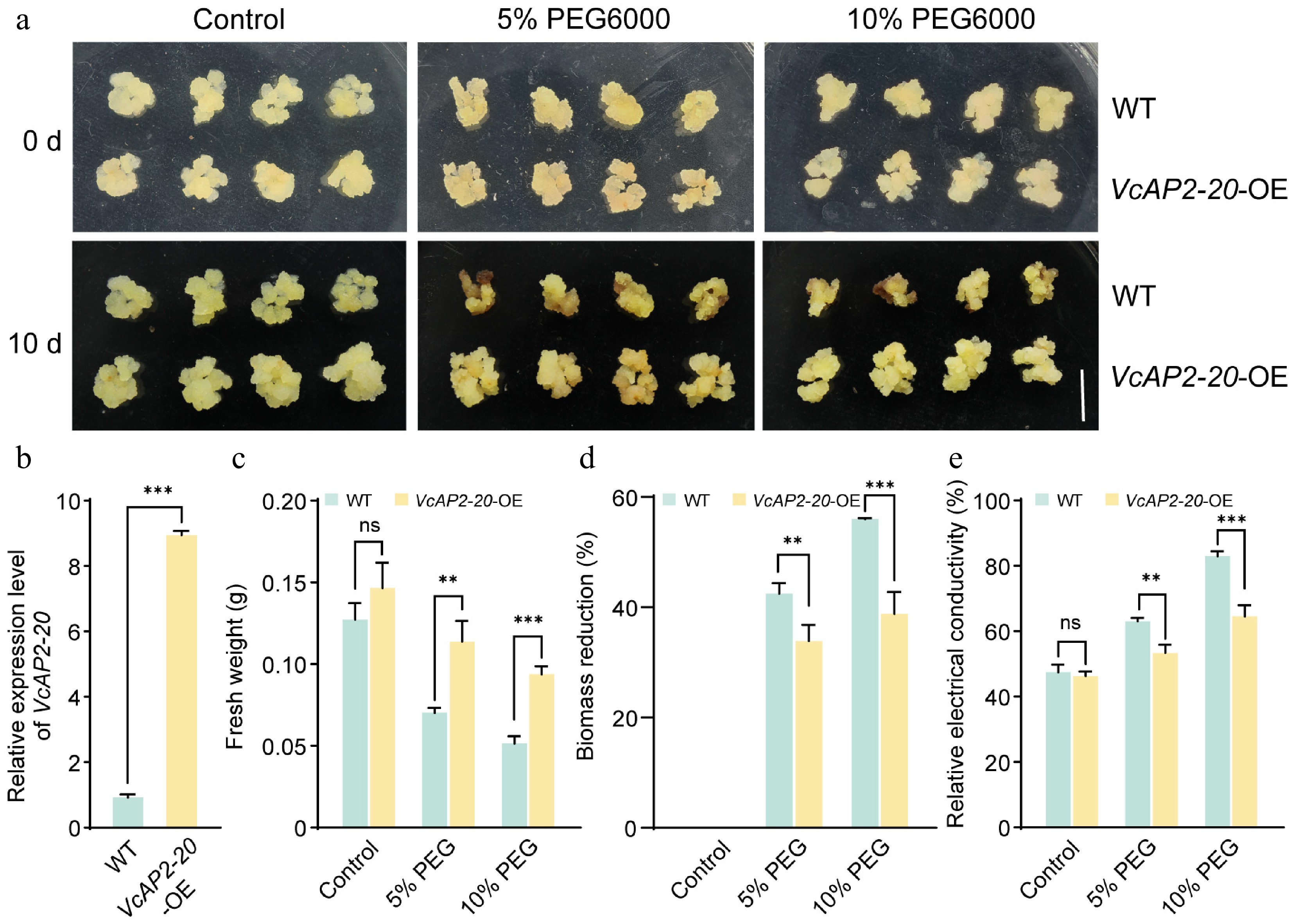
Figure 4.
Overexpression of VcAP2-20 enhances drought tolerance in blueberry calli. (a) Phenotypes of wild-type (WT), and VcAP2-20-overexpressing (VcAP2-20-OE) blueberry calli after 0 and 10 d of drought stress, simulated by 5% or 10% PEG6000. Calli grown under normal conditions served as controls. Scale bar = 1 cm. (b) RT-qPCR analysis of relative expression levels of VcAP2-20 in WT and VcAP2-20-OE blueberry calli. Physiological indices of WT and VcAP2-20-OE calli under drought stress: (c) fresh weight, (d) biomass reduction, and (e) relative electrical conductivity. Values are presented as mean ± SD from three biological replicates. Statistical significance was determined by Student's t-test: * p < 0.05, ** p < 0.01, *** p < 0.001.
Under normal growth conditions, WT and VcAP2-20-overexpressing (VcAP2-20-OE) calli exhibited similar phenotypes. However, when subjected to drought stress simulated by 5% or 10% PEG6000, WT calli exhibited severe browning and reduced viability, whereas VcAP2-20-OE calli remained healthy (Fig. 4a). Consistently, the fresh weight of VcAP2-20-OE calli was 1.62- and 1.82-fold higher than that of WT under 5% and 10% PEG6000, respectively (Fig. 4c). Similarly, the biomass reduction of VcAP2-20-OE calli was significantly lower than that of WT calli following PEG6000 treatment, indicating that overexpression of VcAP2-20 mitigates drought stress-induced water loss (Fig. 4d). Furthermore, although the relative electrical conductivity increased with drought intensity, it was consistently lower in VcAP2-20-OE calli than in WT under drought stress, while no difference was detected under control conditions (Fig. 4e). Collectively, these results demonstrate that overexpression of VcAP2-20 enhances drought tolerance in blueberry calli.
VcAP2-20 promotes ROS scavenging in blueberry calli
-
The cellular redox balance, governed by the production and scavenging of reactive oxygen species (ROS), is pivotal for plant stress adaptation. To assess superoxide anion (O2−) accumulation, we subjected WT and VcAP2-20-OE calli to drought stress simulated with 5% or 10% PEG6000, and then performed nitroblue tetrazolium (NBT) staining. NBT forms an insoluble blue formazan precipitate in the presence of O2−. Under control conditions, both genotypes showed faint, comparable staining. With increasing PEG6000 concentration, staining intensified in all calli; however, VcAP2-20-OE calli consistently exhibited markedly lighter staining than the WT (Fig. 5a). Consistent with this, quantitative measurement of hydrogen peroxide (H2O2) revealed that its levels remained significantly lower in VcAP2-20-OE calli than in the WT after PEG6000 treatment (Fig. 5b). Together, these results indicate that overexpression of VcAP2-20 alleviates drought-induced ROS accumulation.
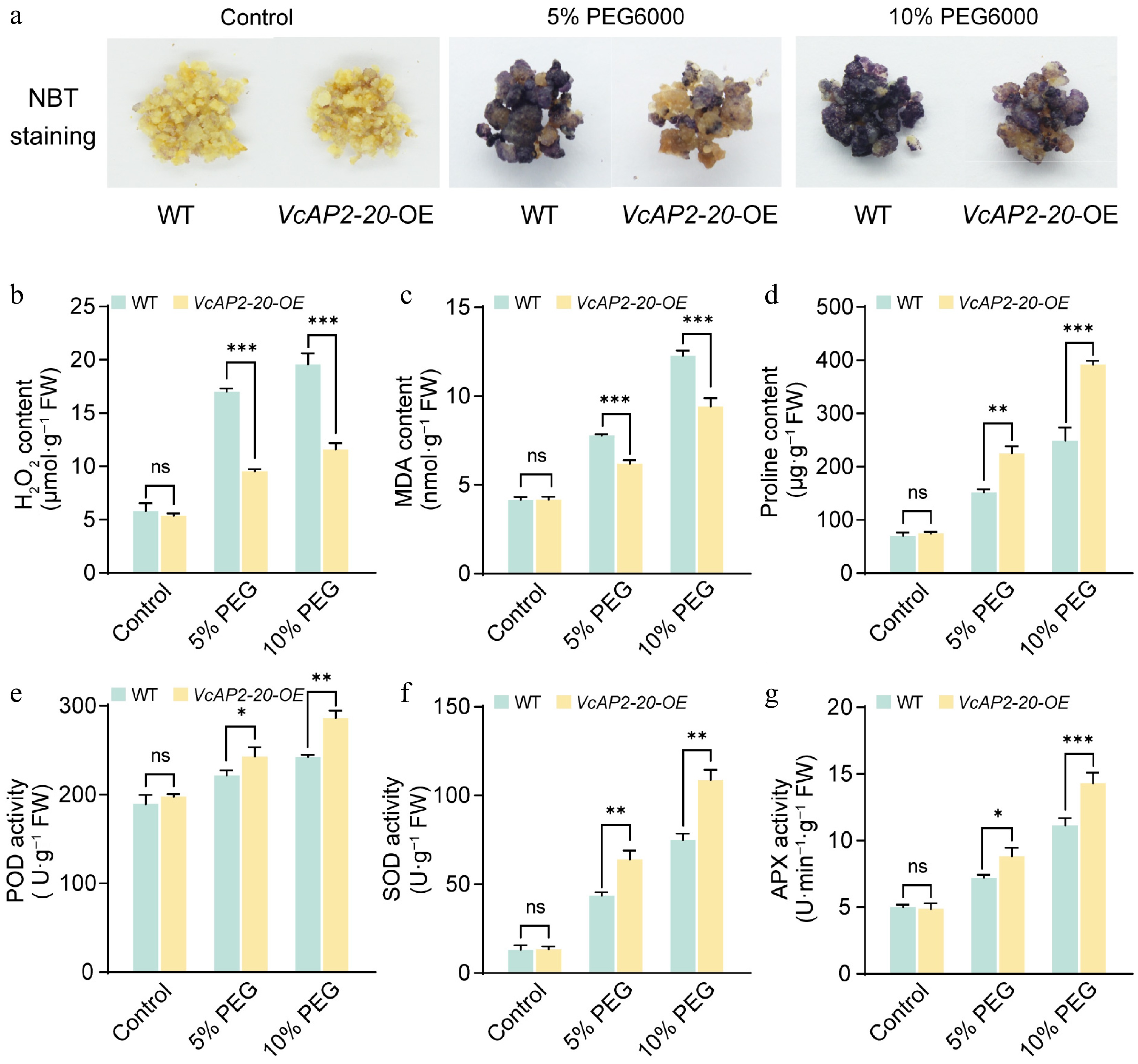
Figure 5.
Overexpression of VcAP2-20 enhances reactive oxygen species (ROS) scavenging in blueberry calli. (a) NBT staining indicating superoxide accumulation in wild-type (WT) and VcAP2-20-overexpressing (VcAP2-20-OE) blueberry calli after 10-d treatment with 5% or 10% PEG6000. Untreated calli under normal conditions served as controls. Contents of (b) H2O2, (c) malondialdehyde (MDA), and (d) proline, as well as activities of antioxidant enzymes; (e) peroxidase (POD), (f) superoxide dismutase (SOD), and (g) ascorbate peroxidase (APX) in WT and VcAP2-20-OE blueberry calli following the same treatments. FW, fresh weight. Values are presented as mean ± SD from three biological replicates. Statistical significance was determined by Student's t-test: * p < 0.05, ** p < 0.01, *** p < 0.001.
We next evaluated membrane lipid peroxidation by measuring the content of malondialdehyde (MDA), a terminal product of this process. Under control conditions, MDA levels did not differ significantly between VcAP2-20-OE and WT calli. However, following PEG6000 treatment, the MDA content in VcAP2-20-OE calli was reduced to approximately 80% of that in the WT (Fig. 5c). To determine whether VcAP2-20 also contributes to osmotic adjustment, we quantified proline levels. Drought stress triggered pronounced proline accumulation in VcAP2-20-OE calli, reaching levels 1.48- and 1.57-fold higher than those in the WT under 5% and 10% PEG6000, respectively (Fig. 5d). These results demonstrate that VcAP2-20 overexpression mitigates membrane damage and enhances osmotic adjustment under drought stress.
Furthermore, the activities of peroxidase (POD), superoxide dismutase (SOD), and ascorbate peroxidase (APX), were measured as indicators of ROS scavenging capacity. Analysis revealed that under 5% PEG6000 treatment, the activities of POD, SOD, and APX in VcAP2-20-OE calli were 1.10, 1.47, and 1.22 times higher, respectively, than those in WT calli. When the PEG6000 concentration was increased to 10%, the activities of these three antioxidant enzymes further rose to 1.18-, 1.45-, and 1.29-fold those in the WT, respectively (Fig. 5e−g). These results demonstrate that VcAP2-20 enhances the drought tolerance of blueberry calli by elevating the activity of key antioxidant enzymes. Collectively, these findings indicate that overexpression of VcAP2-20 improves ROS scavenging under drought stress, thereby effectively increasing the drought tolerance of blueberry calli.
VcAP2-20 positively regulates drought tolerance in blueberry seedlings
-
To further investigate the role of VcAP2-20 in drought tolerance in blueberry plants, we generated transgenic seedlings with either overexpression (VcAP2-20-OE) or RNA interference-mediated suppression (VcAP2-20-RNAi) of the gene. Semi-quantitative RT-PCR was first conducted to preliminarily evaluate the expression of VcAP2-20 in WT and transgenic seedlings, revealing that all four VcAP2-20-OE lines (VcAP2-20-OE#1, #2, #3, #4) exhibited stronger amplification bands than WT, while all four tested VcAP2-20-RNAi lines (VcAP2-20-RNAi#1, #2, #3, #4) showed weaker band intensity compared with the WT control (Supplementary Fig. S4). RT-qPCR analysis confirmed that VcAP2-20 expression was upregulated more than fourfold in all VcAP2-20-OE lines, while it was downregulated over twofold in all VcAP2-20-RNAi lines tested (Fig. 6b). WT and transgenic seedlings were then subjected to natural drought stress, with well-watered seedlings serving as the control. After 25 d, most leaves of the VcAP2-20-RNAi lines showed severe yellowing and wilting compared with the WT, with some leaves even turning red. In contrast, leaves of the VcAP2-20-OE lines exhibited only mild water loss and slight yellowing (Fig. 6a). Physiological measurements corroborated these phenotypic observations. Under drought conditions, the relative water content of leaves from VcAP2-20-RNAi lines decreased to approximately 46 % of that in the WT (Fig. 6c), and chlorophyll content dropped below 40 % of the WT level (Fig. 6d). In contrast, relative electrical conductivity was about 1.20-fold higher than in WT (Fig. 6e). Conversely, leaves from VcAP2-20-OE lines retained significantly higher relative water content and chlorophyll content, along with lower relative electrical conductivity, compared with WT plants (Fig. 6c−e). These results demonstrated that VcAP2-20 mitigates drought-induced damage in blueberry plants.
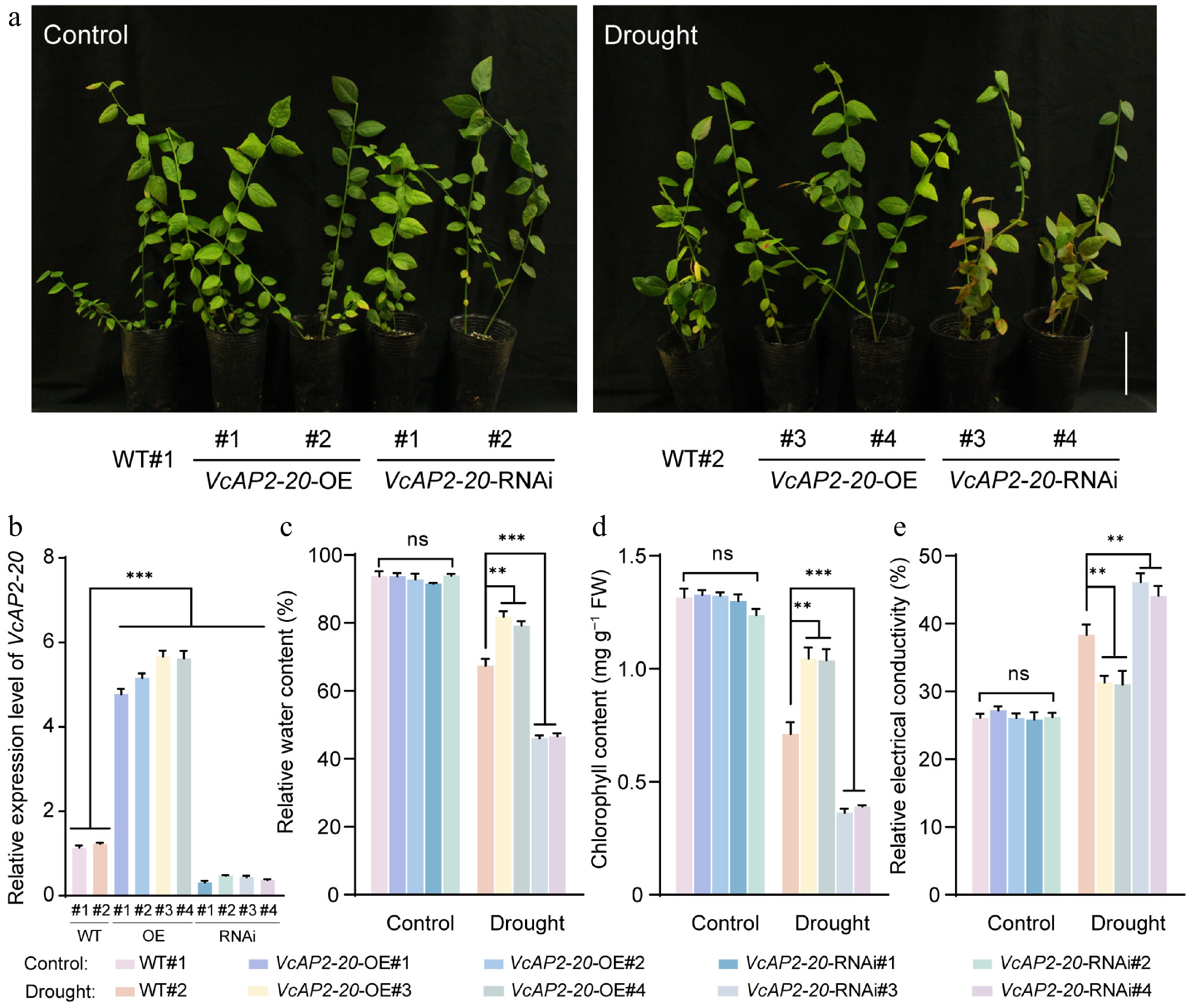
Figure 6.
VcAP2-20 promotes drought tolerance in blueberry seedlings. (a) Phenotypes of wild-type (WT) and transgenic blueberry seedlings after 25 d of natural drought treatment, with normally watered plants serving as controls. VcAP2-20-OE: VcAP2-20-overexpression line; VcAP2-20-RNAi: RNA interference-mediated VcAP2-20 suppression line. Scale bar = 10 cm. (b) RT-qPCR analysis of relative expression levels of VcAP2-20 in WT and transgenic blueberry seedlings. Physiological parameters of seedlings under drought stress; (c) relative water content, (d) chlorophyll content, and (e) relative electrical conductivity. Values are presented as mean ± SD from three biological replicates. Statistical significance was determined by Student's t-test: * p < 0.05, ** p < 0.01, *** p < 0.001.
VcAP2-20 upregulates drought-responsive genes to promote ROS homeostasis under drought stress
-
To further elucidate the role of VcAP2-20 in modulating ROS scavenging capacity under drought stress, we conducted quantitative analyses of key ROS-related physiological parameters. Under drought conditions, the H2O2 and MDA contents in leaves of VcAP2-20-OE lines were approximately 80% of those measured in WT plants (Fig. 7a, b). Concurrently, proline accumulation in VcAP2-20-OE lines increased by more than 1.7-fold relative to WT, whereas VcAP2-20-RNAi lines showed an approximate 25% reduction in proline content under the same stress conditions (Fig. 7c). To determine whether the enhanced drought tolerance of VcAP2-20-OE blueberry seedlings is associated with improved antioxidant enzyme activity, we measured the activities of three key antioxidant enzymes, namely APX, SOD, and POD, in leaf tissues. Under drought stress, VcAP2-20-OE leaves exhibited significantly higher APX, SOD, and POD activities compared with WT leaves, while VcAP2-20-RNAi lines exhibited a marked decrease in the activities of these enzymes (Fig. 7d−f). Collectively, these results suggest that VcAP2-20 enhances ROS scavenging by upregulating antioxidant enzyme activities, thereby contributing to improved drought tolerance in blueberry.
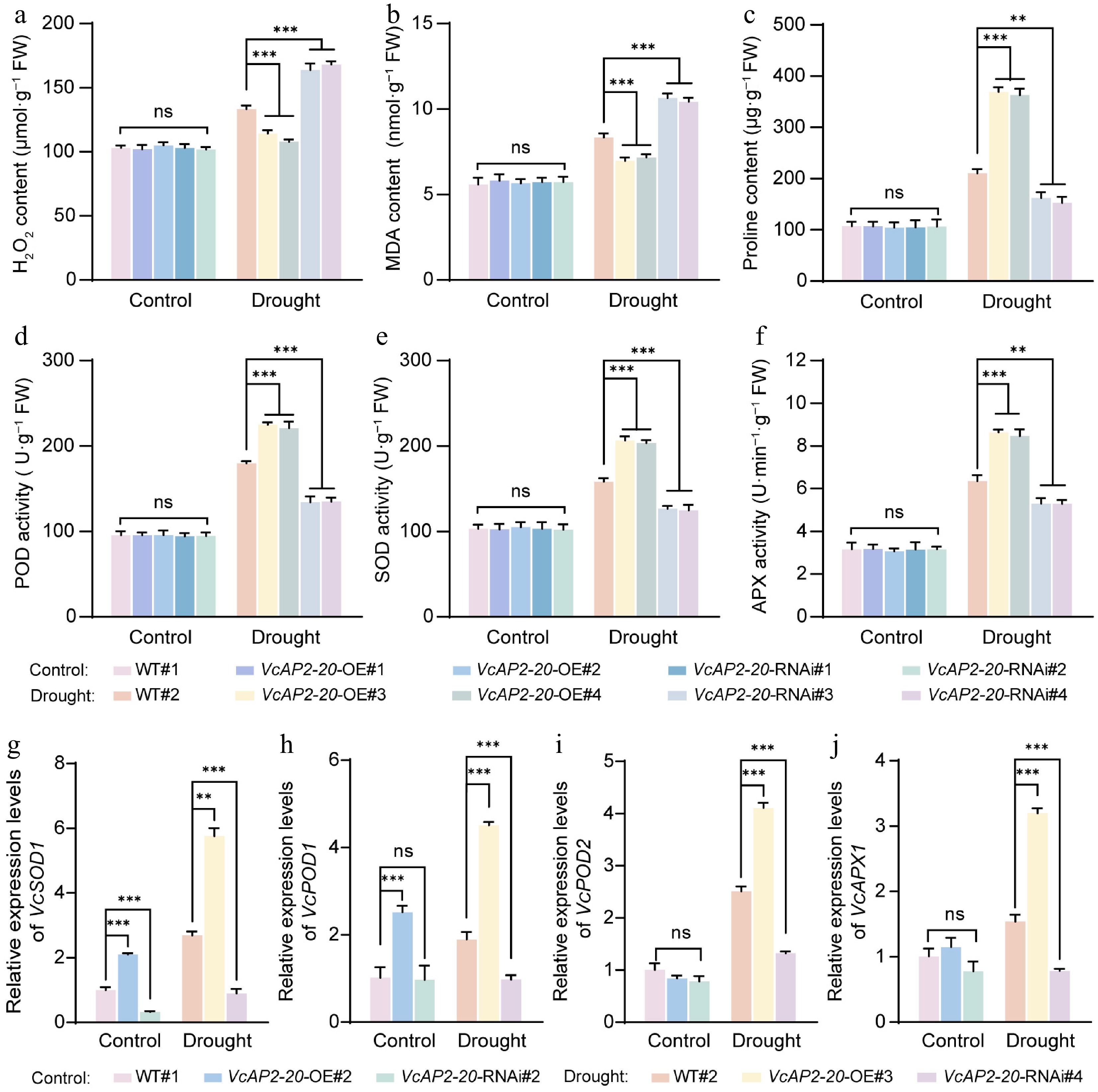
Figure 7.
VcAP2-20 enhances ROS scavenging capacity by activating drought-responsive genes. Contents of (a) H2O2, (b) MDA, and (c) proline, as well as activities of antioxidant enzymes; (d) POD, (e) SOD, and (f) APX in WT and transgenic blueberry seedlings following the same treatments. FW, fresh weight. (g)−(j) RT-qPCR analysis of antioxidant gene expression in leaves; (g) VcSOD1, (h) VcPOD1, (i) VcPOD2, and (j) VcAPX1. Values are presented as mean ± SD from three biological replicates. Statistical significance was determined by Student's t-test: * p < 0.05, ** p < 0.01, *** p < 0.001.
To explore the mechanism by which VcAP2-20 enhances drought tolerance, we examined the expression levels of key drought-responsive genes VcSOD1, VcPOD1, VcPOD2, and VcAPX1 in the leaves of WT and transgenic seedlings under natural drought treatment. Under well-watered conditions, the expression levels of VcSOD1 and VcPOD1 in VcAP2-20-OE#2 leaves were significantly higher than those in the WT, while only VcSOD1 expression in VcAP2-20-RNAi#2 showed a significant increase compared to WT. No significant differences were observed in the expression of VcPOD2 or VcAPX1 between WT and transgenic seedlings under these conditions (Fig. 7g−j). Notably, under drought stress, the expression of all four enzyme-encoding genes was significantly upregulated in VcAP2-20-OE#3 leaves, while it was downregulated in VcAP2-20-RNAi#4 leaves (Fig. 7g−j). These results indicate that under drought conditions, VcAP2-20 enhances drought tolerance in blueberry by upregulating the expression of key drought-responsive genes.
-
AP2 transcription factors play crucial roles in regulating diverse physiological and biochemical processes, particularly in the development of vegetative tissues and reproductive organs. This study systematically identified 32 VcAP2 subfamily transcription factors in blueberry and investigated the key role of VcAP2-20 in regulating drought stress responses. Our findings expand the understanding of AP2 transcription factors in plant stress response, and reveal a critical functional link between VcAP2-20, a specific AP2 family member, and enhanced drought resistance in blueberry via the regulation of antioxidant defense and osmotic homeostasis in response to MeJA signaling. This work provides important candidate genes and a theoretical basis for the genetic improvement of drought tolerance in blueberry.
With the availability of high-quality genomic data, this study presents the first genome-wide identification and comprehensive bioinformatic analysis of VcAP2s in blueberry. The AP2 subfamily is widely distributed across plant species. For instance, 14, 24, 22, 13, and 42 members have been identified in Arabidopsis, rice, bayberry (Myrica rubra), melon (Cucumis melo), and loquat (Eriobotrya japonica), respectively[7,9,54−56]. Here, we identified 32 non-redundant VcAP2 genes from the blueberry 'Draper' genome, which were consecutively named VcAP2-1 to VcAP2-32 (Supplementary Table S2). Phylogenetic analysis classified these 32 genes into the conserved euAP2, euANT, and basalANT subgroups, consistent with the evolutionary framework established in model plants such as Arabidopsis (Fig. 1).
Under drought stress, elevated endogenous MeJA levels contribute to alleviating reductions in relative water content and net photosynthetic rate, while improving water use efficiency. For example, in poplar (Populus euphratica), PeJAZ2, a key component of the MeJA signaling pathway, enhances drought tolerance and water use efficiency by promoting abscisic acid (ABA)-induced stomatal closure[57]. Beyond water regulation, MeJA also strengthens the antioxidant defense system. Numerous studies have demonstrated that MeJA significantly upregulates antioxidant enzyme activities, thereby reducing the accumulation of malondialdehyde (MDA) and hydrogen peroxide (H2O2), diminishing electrolyte leakage, and ultimately alleviating membrane lipid peroxidation. In Reaumuria trigyna, drought resistance is modulated by RtNAC055 in a MeJA-dependent manner, a mechanism that helps maintain the equilibrium between the antioxidant system and intracellular H2O2 levels[58]. Similarly, in pepper (Capsicum annuum), the calcium sensor-interacting protein kinase CaCIPK3 enhances drought tolerance by upregulating MeJA-related genes and reinforcing antioxidant defense capacity[59]. In this study, promoter analysis revealed that VcAP2 genes are widely involved in potential regulatory networks for hormone and stress responses. Importantly, VcAP2-20 emerged as a research focus because its promoter is enriched with MeJA-responsive and drought-responsive cis-elements, and its expression is strongly induced by both signals. Experiments confirmed that MeJA can activate the VcAP2-20 promoter, a finding consistent with the presence of TGACG/CGTCA motifs, which reveals the molecular basis for its upstream regulation by the MeJA signaling pathway. Given that MeJA is a core signaling molecule in plant drought response, our data support a functional association between VcAP2-20 and the crosstalk between MeJA signaling and drought adaptation in blueberry, highlighting its potential as a candidate regulatory node in this process.
While ERF subfamily members are widely recognized as master regulators of stress responses, direct involvement of the AP2 subfamily in abiotic stress tolerance has been less frequently reported. In tobacco (Nicotiana tabacum), NtERF172 directly activates the NtCAT gene to regulate H2O2 homeostasis and enhance drought tolerance[60]. PtrERF109, an ERF transcription factor in trifoliate orange (Poncirus trifoliata), acts as a positive regulator that enhances cold tolerance by directly activating the peroxidase gene PtrPrx1 to strengthen antioxidant capacity and scavenge ROS[61]. TaERF3, induced by salt and drought stress, enhances tolerance by activating downstream stress-related genes, leading to reduced H2O2 accumulation and improved cellular homeostasis[62]. Our work on VcAP2-20 helps bridge this knowledge gap. Functional validation demonstrated that VcAP2-20 is a nuclear-localized transcription factor that positively regulates drought tolerance (Fig. 3). Overexpression of VcAP2-20 in blueberry calli and seedlings significantly improved survival under drought and effectively alleviated stress-induced physiological damage, including reduced water loss, decreased membrane lipid peroxidation (MDA content), and lower electrolyte leakage. The core protective mechanism lies in the enhanced ROS scavenging capacity mediated by VcAP2-20 (Figs. 5, 7). Specifically, overexpression of VcAP2-20 led to significant upregulation of drought-responsive genes such as VcSOD1, VcPOD1, VcPOD2, and VcAPX1 (Fig. 7g−j), which in turn promoted the activity of key antioxidant enzymes POD, SOD, and APX (Figs. 5, 7a−f). This coordinated boost in enzymatic activity directly correlated with a marked reduction in O2− and H2O2 accumulation under drought, as visualized by NBT staining and quantitative assays, thereby preventing oxidative damage to cellular macromolecules (Fig. 5a, b). Proline serves as a compatible solute, stabilizing proteins and cellular structures, and contributes to maintaining turgor pressure[63]. The substantial increase in proline accumulation in VcAP2-20-OE calli under drought indicates enhanced osmotic adjustment capacity (Fig. 5d). Concurrently, the lower MDA levels in transgenic calli directly demonstrate reduced lipid peroxidation, a key indicator of preserved membrane integrity (Fig. 5c). This protection likely results from the combined effects of diminished ROS attack and enhanced osmotic homeostasis.
Although the positive regulatory function of VcAP2-20 in blueberry drought tolerance has been characterized at both the physiological and transcriptional levels, its direct downstream target genes and precise regulatory network remain to be fully elucidated. Insights from functionally homologous genes in other plant species offer valuable clues. In poplar, AP2/ERF transcription factors such as ERF194 enhance drought tolerance by upregulating ROS-scavenging enzyme genes (POD, SOD), suggesting a conserved regulatory module linking this family to antioxidant defense[64]. In rice, OsERF71 directly binds to the promoter of OsCCD1, a key gene in lignin biosynthesis, to improve drought resistance[65]. Additionally, OsERF71 regulates the expression of ABA-responsive genes (OsABI5, OsPP2C68, OsRAB16C, and OsRAB16D) and proline biosynthesis genes (OsP5CS1 and OsP5CS2) under drought stress[66]. Given that overexpression of VcAP2-20 significantly upregulates the expression of VcSOD1 in blueberry (Fig. 7g), it is plausible that these genes are direct transcriptional targets of VcAP2-20. These inferences highlight promising candidates for downstream regulation, and future studies will be essential to validate these direct regulatory relationships and fully elucidate the precise molecular network through which VcAP2-20 mediates drought tolerance in blueberry.
-
In summary, we propose a working model: drought stress directly induces VcAP2-20 expression and simultaneously stimulates endogenous MeJA accumulation, which in turn activates VcAP2-20 transcription; this transcription factor then promotes the expression of a series of drought-responsive genes, strengthening the ROS scavenging system, thereby mitigating oxidative damage and maintaining cellular homeostasis, ultimately conferring enhanced drought tolerance to blueberry. This study reveals a novel mechanism in blueberry whereby an AP2 transcription factor regulates drought response by integrating MeJA signaling and the antioxidant pathway. It provides genetic resources for improving drought tolerance in blueberry and related crops through molecular breeding.
-
The authors confirm their contributions to the paper as follows: conceived the project: Zhang L, Cao Y; designed the study: Zhang L, Xu Y; performed the experiments and collected the data: Xu Y, Qiu X, Du B, Li X, Yang H; improved the charts: Cao Y; drafted the manuscript: Xu Y, Cao Y; revised the manuscript: Zhang L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
This work was supported by the Agriculture Science and Technology Major Project (Grant No. NK202201020408), the Key R&D Program of Shandong Province (2024LZGCQY025), and the 5·5 Engineering Research & Innovation Team Project of Beijing Forestry University (Grant No. BLRC2023B08).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/frures-0026-0016.
- Supplementary Table S1 All primers used in this study.
- Supplementary Table S2 Physicochemical properties analysis of VcAP2 proteins in blueberry.
- Supplementary Fig. S1 Multiple sequence alignment of the amino acid sequence of euAP2 subgroup members in Arabidopsis and blueberry.
- Supplementary Fig. S2 VcAP2-20 expression is induced by multiple plant hormones, with the strongest response observed under methyl jasmonate (MeJA) treatment.
- Supplementary Fig. S3 The observation of GFP fluorescence in VcAP2-20-overexpressing blueberry calli.
- Supplementary Fig. S4 Semi-quantitative RT-PCR detection of VcAP2-20 expression in WT and transgenic plants.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu Y, Qiu X, Li X, Du B, Yang H, et al. 2026. Methyl jasmonate-responsive VcAP2-20, a novel transcription factor, promotes drought tolerance in blueberry. Fruit Research 6: e024 doi: 10.48130/frures-0026-0016 

Methyl jasmonate-responsive VcAP2-20, a novel transcription factor, promotes drought tolerance in blueberry
- Received: 04 February 2026
- Revised: 18 March 2026
- Accepted: 20 March 2026
- Published online: 03 July 2026
Abstract: Blueberries (Vaccinium spp.) are small berry-bearing shrubs that boast high nutritional value and exhibit extensive development prospects. However, their shallow root systems render them sensitive to drought stress. Although several APETALA2/ETHYLENE-RESPONSIVE FACTOR (AP2/ERF) subfamilies are implicated in drought stress responses, the specific role of the AP2 subfamily, particularly in conferring drought tolerance in blueberry, remains elusive. In this study, we identified 32 non-redundant VcAP2 genes encoding proteins containing two AP2 domains. Cis-acting element analysis of the VcAP2 promoters revealed a widespread presence of MeJA-responsive and drought-responsive elements. Transcriptome data and GUS activity assays confirmed that VcAP2-20 is MeJA-inducible. VcAP2-20 expression was also induced under drought stress. Overexpression of VcAP2-20 in blueberry calli and seedlings significantly enhanced drought tolerance, as evidenced by reduced water loss, lower electrolyte leakage and ROS accumulation, decreased malondialdehyde (MDA) content, and increased proline levels and antioxidant enzyme activities. Conversely, RNAi-mediated suppression of VcAP2-20 increased drought sensitivity. Furthermore, overexpression of VcAP2-20 upregulated the expression of key antioxidant genes, including VcSOD1, VcPOD1, VcPOD2, and VcAPX1, under drought stress. Our findings indicate that VcAP2-20, a MeJA- and drought-responsive AP2 transcription factor, enhances blueberry drought tolerance via reinforcing the antioxidant defense system. This work provides valuable genetic resources and supports the development of blueberry cultivars with improved drought resilience.
-
Key words:
- Blueberry /
- APETALA2 /
- VcAP2-20 /
- Methyl jasmonate /
- Drought tolerance