








-
Genome editing refers to the targeted and precise modification of the genetic material of living organisms[1]. The technology exploits nucleases to introduce site-specific double-strand breaks (DSBs). These breaks are then repaired by non-homologous end joining (NHEJ) or homology-directed repair (HDR)[1], thereby achieving gene editing. Technologies such as meganucleases[2], zinc finger nucleases (ZFNs)[3], and transcription activator-like effector nucleases (TALENs)[4] have laid the foundation for genome editing. However, these technologies are difficult to be utilized, which limits their wide application.
Clustered Regularly Interspaced Short Palindromic Repeats/ CRISPR-associated proteins (CRISPR/Cas) represent a revolutionary gene‐editing system that employs RNA‐guided Cas nucleases to introduce double‐strand breaks at specific genomic loci[5]. The CRISPR/Cas system was demonstrated to be an adaptive genomic defense mechanism of bacteria that can recognize and target foreign nucleic acids derived from viruses (phages), or other mobile genetic elements[6]. The CRISPR array is transcribed into CRISPR RNA (crRNA), which pairs with trans-acting crRNA (tracrRNA) to form a binary complex[7,8]. This complex guides the Cas9 to cleave the foreign DNA sequence matching the crRNA, eliminating the invader. The Cas9 from Streptococcus pyogenes (SpCas9) was the first Cas nuclease repurposed for genome editing. Due to its high activity and specificity, it remains the most widely used gene editing nuclease[9].
Biochemical studies published in 2012 revealed that Cas9 specificity is dictated by the dual-RNA guide structure, composed of crRNA and tracrRNA[5]. To facilitate application in eukaryotes, crRNA and tracrRNA were fused into a single-stranded guide RNA (sgRNA), yielding a fully programmable one-nuclease-one-guide-RNA design that underpins modern CRISPR genome editing. Owing to its design simplicity and operational efficiency compared with meganucleases, ZFNs, and TALENs, the CRISPR/Cas system has been widely adopted for diverse applications in plant gene-function analysis, and crop improvement[10−13]. Recent innovations, including the development of base editors[14], prime editors[15], and diverse novel CRISPR systems[16,17] continue to consolidate its pivotal role in both fundamental plant research and precision molecular breeding.
Conventional CRISPR/Cas9 delivery methods rely on genetic transformation like Agrobacterium‐mediated transformation[18] or microprojectile bombardment[19], which frequently integrate foreign DNA into the host genome (Fig. 1). These foreign DNA such as Cas9, sgRNA cassettes, and selectable markers lead to stable transgene inheritance[20], and are called transgenes or T-DNA. However, the persistent activity of T-DNA can elevate off‐target effects, and promote the development of chimeric tissues[21,22], ultimately leading to genomic instability that necessitates extensive screening to identify stable germplasm. Secondly, the unintended dissemination of transgenes into wild plant populations pose risks to ecosystem biodiversity[23]. Although transgene segregation via self‐crossing or backcrossing can eventually eliminate T-DNA[15,17,24], these methods are labor‐intensive and time‐consuming, particularly in crops with complex ploidy and prolonged growth cycles, such as cotton[25] and wheat[26]. Moreover, these approaches are not feasible for asexually propagated crops like potatoes[27], peanuts[28], and strawberries[29]. In conclusion, conventional gene editing methods suffer from inherent limitations due to T-DNA integration into the genome. Thus, developing T-DNA-free gene-editing methods is of considerable importance.
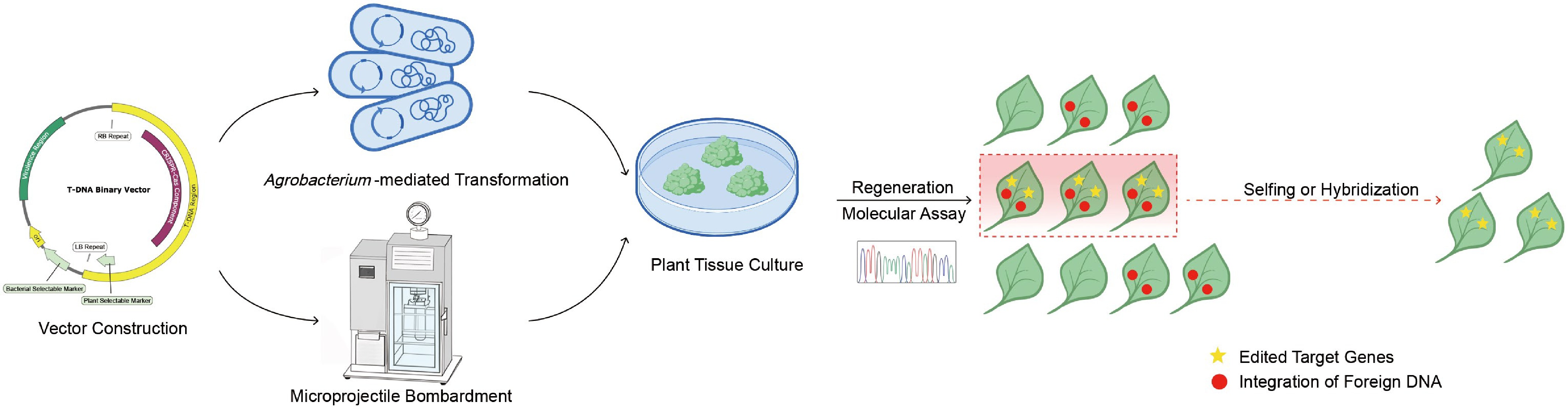
Figure 1.
Schematic overview of conventional CRISPR/Cas9-mediated gene editing in plants.
Plasmid constructs encoding the CRISPR/Cas components are introduced into plant explants or embryos via Agrobacterium‐mediated transformation or particle bombardment. Following tissue culture and regeneration, transgenic plants are obtained in which the CRISPR/Cas elements are stably integrated into the genome. Molecular assays, such as PCR and next‐generation sequencing, are then employed to identify plants harboring target site modifications. Subsequent self‐pollination or outcrossing is performed to segregate out the transgenic sequences, ultimately yielding T‐DNA-free, gene‐edited progeny.Transgene‐free (T-DNA-free) genome editing refers to a technology that targets and modifies genomic sites to obtain gene-edited products without incorporating T-DNA sequences[30], thereby reducing regulatory burdens and detection costs associated with traditional transgenic methods and improving breeding efficiency[30,31]. In this context, T‐DNA-free genome editing offers a promising strategy to significantly enhance crop breeding efficiency, and contribute to sustainable agricultural development and food security. This review delineates the fundamental mechanisms and recent applications of T‐DNA-free genome editing methodologies (Table 1), encompassing visualized screening[32−36], the Transgene Killer CRISPR (TKC) system[37−40], transient DNA/RNA expression[41−44], ribonucleoprotein (RNP)‐mediated editing[45−52], virus‐mediated editing[53−61], pollen‐mediated editing[62−65], and graft‐mobile‐mediated editing[66]. The advantages and limitations of each approach are critically evaluated, and the challenges associated with their broader implementation in crop improvement are discussed in this review.
Table 1. Cases of T-DNA-free genome editing in plants.
Techniques Species Target genes Efficiency of T-DNA-free gene editing Ref. Visualized screening Arabidopsis thaliana AtABP1 About 50% [33] Rice IAMT 64.5% [35] Tomato 37.9% Arabidopsis thaliana 19.6% Transgene Killer CRISPR
(TKC) systemRice OsLAZY1 100% [37] OsCLE 66.67%–95.37% [38] Rice OsSE5 100% [39] OsYSA 100% 84K poplar GLK1 and GLK2 54.5% [40] Transient expression of DNA Wheat TaGASR7 2.19% (Bobwhite); 1.61% (Kenong199) [41] TaGW2 Unidentified TaDEP1 1.08% TaNAC2 1.50% TaPIN1 0.625% TaLOX2 8.24% TdGASR7 0.76% (Shimai11); 0.87% (Yumai4) Tobacco NtPDS 8.20% [42] Transient expression of RNA Wheat TaGW2 1.10% [41] Tomato SlALS1; SlER; SlRbohD; SlDMR6; SlINVINH1 12%–36.3% [43] Tobacco NtALS 1.9% Potato StDMR6 Unidentified Citrus CsLOB1 1.9% Lettuce LsFT Up to 50.0% [44] LsALS With the treatment of benazolin, up to 96.4% (callus), and 51.2% (regenerated plants) LsPDS Up to 3.53% Ribonucleoprotein (RNP)-mediated editing Arabidopsis thaliana AtBRI1 44% [45] AtPHYB 16%–19% Lettuce LsBIN2 46% (callus); 13.5% (regenerated plants) Rice OsAOC 8.4% OsDWD1 17% Tomato SlP450 19%–23% Wheat TaGASR7 1.8% [50] TaLOX2 33.6% TaCER9 23.8% Maize ZmLIG 9.7% [48] ZmALS2 Unidentified ZmMS26 2.4% ZmMS45 3.7% Soybean Gly m Bd 30K 0.4%–4.6% [52] Virus-mediated editing Tobacco NbPDS 43%–61% [53] Tobacco NbPDS 56% (inoculated leaves) [54] NbPCNA 30% (systematic leaves) Tobacco NbPDS 65% [55] NbAG 89% Arabidopsis thaliana AtPDS3 2.9% and 38% [57] AtCHLI1 14.8% and 2.1% Tobacco NbPDS; NbTOM1; NbAG; NbFLS2 0.8%–13.2% [58] Tobacco NbPDS Over 90% [59] NbRDR6 90% NbSGS3 100% NbGFP 77% and 91% Tobacco PDS 26.4%–78.2% [60] Tomato FucT Pepper DCL2 Peanut RDR6 Physalis alkekengi SGS3;BBLd; ER; AHAS1 Wheat TaPDS 65.2% [61] Pollen-mediated editing Maize ZmVLHP 12.5% [63] ZmGW2 6.25% Wheat TaGT1 16.9% Arabidopsis thaliana AtGL1 Up to 16.9% Cole crops BoFAD2 1.89%–12.11% [65] Brassica napus BnFAD2 12.92%–12.97% Graft-mobile-mediated editing Brassica napus BrNIA1 0.57%–0.50% (TLS1 and TLS2) [66] -
To streamline the isolation of non-transgenic mutants, visual-screening systems were established. Independent promoter-driven visual screening markers (such as fluorescent proteins or pigment genes) are added to the CRISPR/Cas9 vector. During the genetic transformation process, transgenic sequences, including visual screening markers, may be integrated into the plant genome. Due to the presence of visual screening markers in transgenic mutants, they can be identified by fluorescence microscopy or the naked eye, thereby retaining non-transgenic mutants. Gao et al. introduced an mCherry expression cassette driven by the strong pAt2S3 promoter into a CRISPR/Cas9 vector. Following transformation, T-DNA-free seeds were identified by the absence of cherry red fluorescence during microscopic screening[33]. Similarly, Aliaga-Franco et al. engineered a CRISPR/Cas9 vector containing a DsRED expression cassette to enable fluorescence-based selection of transgenic seeds. This system was validated in tomato, rice, and Arabidopsis, where fluorescence-negative T2 plants lacked Cas9-specific sequences, confirming the absence of transgenes[35]. In addition, Liu et al.[36] integrated the Flowering Locus T (FT) and PAP1 genes into a CRISPR/Cas9 vector, called the Cas9-PF system. Overexpression of FT accelerates flowering[34], while PAP1 overexpression results in anthocyanin accumulation, producing a purple leaf phenotype[32]. In Arabidopsis, green-leaved T1 plants, which lack the anthocyanin-induced coloration, were selected based on visual observation, and subsequently confirmed as transgene-free by PCR analysis[36]. Notably, the Cas9-PF system reduced the overall breeding cycle, significantly decreasing both time and labor requirements.
This strategy effectively distinguishes transgenic from non-transgenic individuals through visual markers, significantly reducing the workload of molecular testing. Furthermore, visual selection markers pose fewer environmental and health concerns than antibiotic and/or herbicide resistance markers. However, it is important to note that exceptions such as silencing of visual marker gene expression, and T-DNA fragment integration require molecular testing to ensure screening accuracy.
Transgene Killer CRISPR (TKC) system
-
The first-generation Transgene Killer-CRISPR (TKC) system comprises two coordinated genetic cassettes designed to automatically eliminate transgenic elements during plant reproduction. The first component encodes CMS2, a mitochondrial-targeted cytotoxic protein derived from rice, which when constitutively expressed under the CaMV 35S promoter, induces male sterility by disrupting mitochondrial dynamics in developing pollen grains[67]. The second component employs BARNASE, a non-specific ribonuclease driven by the rice REG2 promoter, highly active during early embryogenesis, to selectively ablate embryos that retain T‐DNA inserts[68,69]. He et al. integrated both the 35S::CMS2 and REG2::BARNASE cassettes into a conventional CRISPR/Cas9 binary vector and targeted the rice LAZY1 locus, whose loss-of-function mutations manifest as an increased tillering angle. In the T1 generation, progeny displaying the altered tillering phenotype were confirmed by sequencing to harbor biallelic mutations at LAZY1 and, by PCR and Southern blot analyses, to be devoid of any transgenic sequences, thereby validating the self‐eliminating capability of the TKC system[37]. Building on this strategy, Liu et al. combined the TKC system with multi-gene editing to develop a rice custom assembly and simplified editing (CASE) toolkit, achieving T-DNA-free editing of multiple genes in rice[38].
Notably, the TKC system automatically eliminates the transgene via a 'suicide cassette' after gene editing in the T0 generation, but 1%–5% of T1 plants still retain the transgene. Zhu et al. combined the RUBY reporter gene with the TKC system to develop the TKC2 system[39]. The RUBY gene is a synthetic betalain biosynthesis pathway gene, consisting of three key enzyme genes in series, which can efficiently convert tyrosine into red-purple betalain, visible to the naked eye in plants. TKC2 tracks the presence of transgenes in real time through color changes, further reducing the risk of transgene escape and achieving a 100% efficiency in generating transgene-free progeny in the T0 generation. Similar to the principles of the TKC2 system, Wu et al. constructed the Visual Monitoring DNA-Free Multi-Gene Editing System (VMDFGE), a system that integrates a Ruby visual screening system, multi-gene editing, and a heat-induced exogenous gene removal system (LoxP::FRT/FLP)[40]. They inserted LoxP::FRT double recognition sites on the left and right borders (LB and RB) of the vector T-DNA. The vector also contains FLP recombinase driven by an HSP promoter. Under appropriate temperature induction, FLP recombinase cleaves genomic DNA at the recognition sites, leaving only the LB-LoxP::FRT-RB structure, which is then excised and degraded from the host genome.
Overall, the TKC and TKC2 system offers a robust mechanism for the automatic purging of transgenic material, thereby streamlining the recovery of T‐DNA-free, genome‐edited plants, and obviating laborious molecular screening. However, because its mode of action depends on meiotic segregation and seed development, its application is currently restricted to sexually propagated species and is not directly transferable to asexually propagated crops. The LoxP::FRT/FLP system is suitable for perennial woody plants and asexual species, but after excision, a LoxP::FRT scar of less than 138 bp remains in the genome. Although the fragment is extremely small, its impact on plant growth and development, and biosafety needs to be systematically evaluated.
CRISPR/Cas DNA transient expression for genome editing
-
Conventional plant genome editing often involves the stable integration of CRISPR/Cas DNA constructs into the plant genome through methods such as Agrobacterium-mediated transformation or particle bombardment[18,19]. These techniques can also facilitate transient expression of CRISPR/Cas vectors, enabling the regeneration of genome-edited plants without the incorporation of foreign DNA (Fig. 2). In this approach, the introduced plasmid DNA is gradually degraded by endogenous nucleases, resulting in T-DNA-free, genome-edited T0 plants.
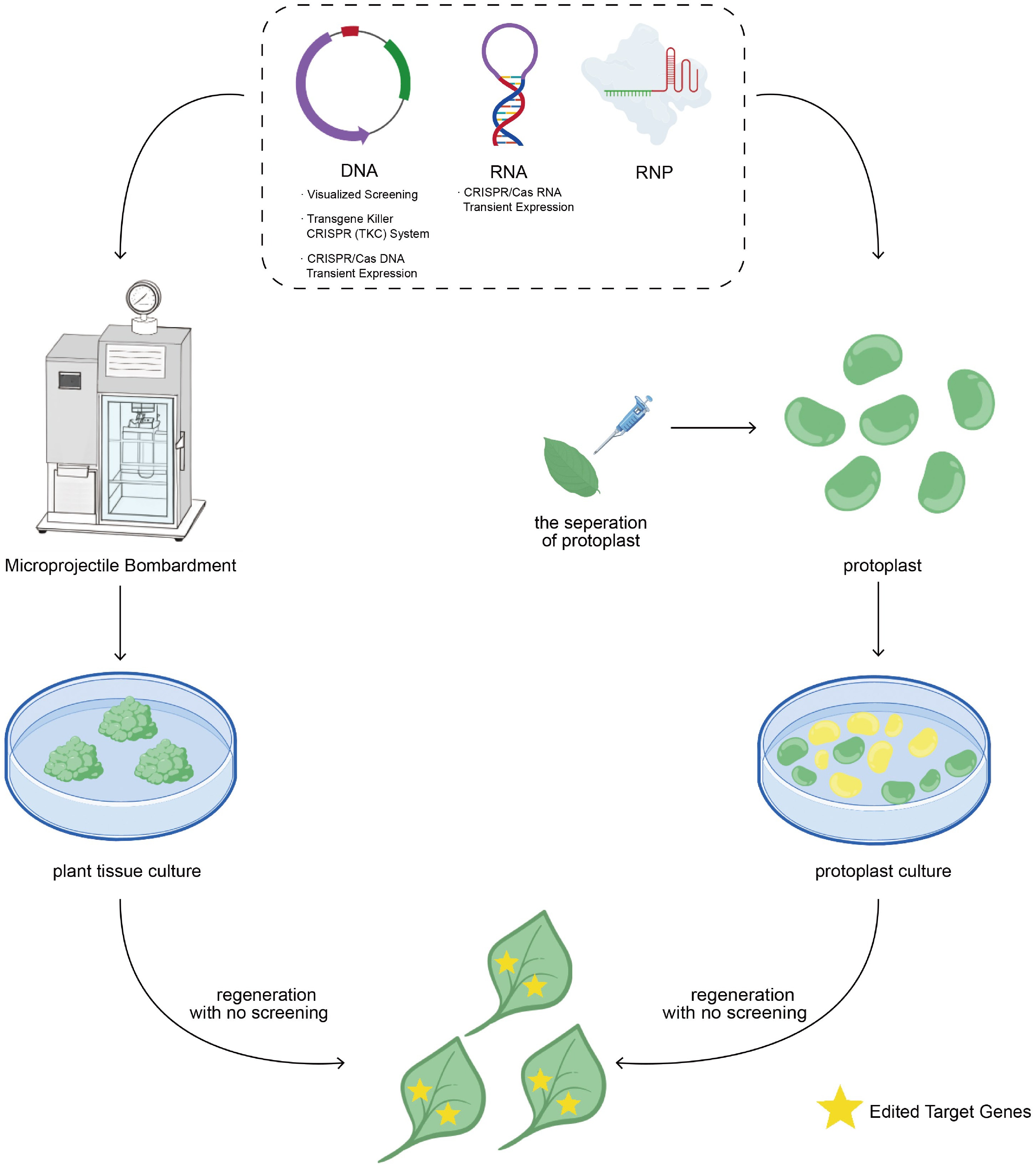
Figure 2.
Schematic representation of CRISPR/Cas delivery and regeneration of T-DNA-free, genome-edited plants.
CRISPR/Cas DNA, RNA, or RNP is introduced into plant tissues via particle bombardment, or into protoplasts using polyethylene glycol (PEG)-mediated transformation. Regenerated plants are obtained without reliance on antibiotic or herbicide selection, and subsequent molecular analyses confirm the absence of transgenic sequences in the edited plants.Zhang et al. developed the Transiently Expressing CRISPR/Cas9 DNA (TECCDNA) method, wherein CRISPR/Cas9 DNA constructs were directly delivered into immature wheat embryos via particle bombardment. This approach targeted seven genes across four wheat varieties, achieving editing efficiencies in T0 plants ranging from 1.0% to 9.5%, with 43.8% to 86.8% of the edited plants being T-DNA-free[41]. Similarly, Chen et al. targeted the phytoene desaturase (PDS) gene in tobacco using Agrobacterium-mediated transient expression of CRISPR/Cas9 constructs. Their strategy produced PDS mutants with an editing efficiency of 47.5%, of which 17.2% were confirmed to be T-DNA-free, representing 8.2% of the total T0 plants[42].
CRISPR/Cas RNA transient expression for genome editing
-
Although the TECCDNA method can generate T-DNA-free mutants, a substantial fraction of gene-edited plants still harbors transgenic components. Additionally, degraded vector fragments may occasionally integrate into the host genome, evading detection by standard PCR assays. To overcome these limitations, Zhang et al. advanced the approach by developing TECCRNA, which relies on the transient expression of in vitro transcribed (IVT) Cas9 and sgRNA. In this method, particle bombardment is used to deliver IVT reagents into immature wheat embryos, thereby regenerating plants, without the need for antibiotic or herbicide selection. When targeting the GW2 gene in the wheat variety Kenong199, no transgenic sequences were detectable in the T0 mutants, although the editing efficiency was relatively low (1.1%). The reduced efficiency is attributed to the inherent instability of RNA compared to DNA, rendering it more susceptible to degradation. Despite this limitation, TECCRNA offers a robust strategy for obtaining transgene-free, genome-edited plants[41] (Fig. 2).
In a complementary approach, Huang et al. combined CRISPR/Cas RNA transient expression with the visual screening system. With the Agrobacterium-mediated transient expression platform, cytosine base editors (CBE), CRISPR/Cas12a RNA, and green fluorescent protein (GFP) were co-expressed. In this configuration, the CBE component was employed to edit the acetolactate synthase (ALS) gene, thereby conferring resistance to the herbicide chlorsulfuron and establishing a selectable phenotype; CRISPR/Cas12a RNA enabled target gene modification; and GFP expression facilitated the rapid identification of transformants lacking integrated T-DNA. In experiments on tomato, tobacco, potato, and citrus, the pure homozygous or biallelic editing efficiencies in T-DNA-free plants ranged from 1.9% to 42.1%. Whole-genome sequencing confirmed the absence of transgenic sequences, and off-target mutations in predicted regions. Notably, the integration of herbicide resistance selection and GFP screening substantially improved the recovery of T-DNA-free, genome-edited plants in tomato and citrus, compared with previous approaches, that did not incorporate these markers[43].
Lee et al. further advanced RNA-based genome editing by developing a base editing system that exploits RNA transient expression. Their system employed a T7 promoter-driven vector harboring both adenine base editor (ABE), and CBE elements. The corresponding mRNA and sgRNA were delivered into lettuce protoplasts via polyethylene glycol (PEG)-mediated transformation, to achieve transgene-free genome editing. In this system, the ABE-sgRNA complex induced A-to-G conversions at frequencies of up to 37.6% in callus, and 50.0% in stem tissues, while the CBE-sgRNA complex mediated C-to-T transitions with efficiencies of up to 20.8% and 14.2% in calli and stems, respectively. Furthermore, glyphosate treatment further increased the overall gene editing efficiency to 51.2%[44]. Collectively, these results underscore the efficacy of RNA-based ABE and CBE systems in performing precise, transgene-free genome editing in plants, providing a valuable tool for functional genomics and crop improvement.
Ribonucleoprotein(RNP)-mediated genome editing
-
Ribonucleoprotein (RNP) is a ribonucleoprotein complex preassembled in vitro by Cas9 protein and guide RNA. RNP‐mediated genome editing entails the direct introduction of RNP complexes into plant cells, obviating the need for plasmid vectors that encode these components (Fig. 2). This approach minimizes the risk of integration of plasmid-derived expression elements into the host genome, thereby facilitating the generation of T‐DNA-free mutant plants. RNPs have been delivered via multiple methods, including polyethylene glycol (PEG)‐mediated transformation[45−47,49−51], particle bombardment[48,52], and electroporation[70]. Recipient tissues include protoplasts[46,47,49−51], immature embryos[51], and meristematic tissues[52].
Woo et al. pioneered the PEG‐mediated delivery of RNP complexes into protoplasts isolated from Arabidopsis, tobacco, lettuce, and rice, achieving editing efficiencies of up to 46% in regenerated plants[45]. Subsequent applications of this method have extended to species such as apple[46], grape[47], petunia[47], wheat[49], and reishi (Ganoderma lucidum)[51]. Liang et al. further expanded this strategy by delivering CRISPR‐Cpf1 RNPs and TALEN proteins into wheat and rice protoplasts, demonstrating the versatility of the PEG‐mediated approach[50].
Svitashev et al. employed particle bombardment to deliver RNP complexes into immature corn embryos, in conjunction with DNA vectors expressing cell division–promoting transcription factors (ODP2 and WUS), and a marker gene (MOPAT‐DSRED) for visual screening. Control experiments using embryos transformed solely with the Cas9 protein (negative control), and those receiving DNA vectors encoding Cas9, and four sgRNAs (positive control) confirmed mutations at all target sites following RNP or DNA vector delivery, while no mutations were detected in the negative controls. Notably, the RNP‐based approach yielded T‐DNA-free plants with editing efficiencies ranging from 2.4% to 9.7%, and exhibited a significant reduction in off‐target effects compared with plasmid‐based methods[48]. Similarly, Kuwabara et al. achieved T-DNA‐free genome editing in soybean by delivering CRISPR/Cas9 RNP complexes directly into the shoot apical meristem (SAM) cells via particle bombardment (iPB‐RNP), thereby circumventing tissue culture. In various soybean varieties, heritable editing efficiencies in the T1 generation were approximately 0.4%–4.6% of T0 plants[52].
In summary, RNP-mediated genome editing is a DNA-free system. Preassembled in vitro RNP is directly introduced into plant cells, eliminating the need for any transcription or translation processes. Editing and degradation are completed within hours, eliminating the risk of exogenous DNA integration. By precisely regulating the protein-to-RNA molar ratio, it enables dose-dependent editing, avoiding excessive or sustained expression. This significantly reduces the risk of off-target editing and plant immune responses, significantly shortening the time required to obtain gene-edited plants. However, the preparation of protoplasts and subsequent tissue culture regeneration remain challenging for certain plant species. Although co‐expression of developmental regulatory genes can enhance regeneration efficiency, this strategy carries an increased risk of transgene integration.
Virus-mediated genome editing
-
Viral-mediated plant genome editing exploits the natural infectivity of viruses to deliver CRISPR/Cas components into plant cells, where they replicate and spread to achieve targeted genome modification. Among current viral vectors, positive-strand RNA viruses are the most widely adopted. However, their limited cargo capacity—typically < 1,000 bp—restricts the size of exogenous sequences they can carry[70]. Because CRISPR/Cas is a two-component system, one common strategy is to generate transgenic lines that constitutively overexpress Cas9 via genetic transformation. SgRNA is then introduced into these Cas9-expressing plants by viral vectors, such as tobacco rattle virus (TRV), or barley stripe mosaic virus (BSMV)[53−55,61]. This approach bypasses tissue culture during the editing step, and ultimately yields gene-edited, transgene-free progeny. Although this approach enables infinite rounds of gene editing with only one round of tissue culture, it cannot completely eliminate the problem of genotype limitations[56].
Recent advances have shown that positive-strand RNA viral vectors can simultaneously deliver both sgRNA and RNA-guided endonucleases to wild-type plants, enabling heritable, T-DNA-free editing without any tissue-culture step. Weiss et al. modified TRV to carry carry the compact RNA-guided TnpB enzyme ISYmu1 and its sgRNA, overcoming the limitations of TRV loading capacity[57]. They also added a tRNA Ileu sequence to the vector to enhance its transmissibility. Whole-genome sequencing of the progeny of TRV-infected plants confirmed that the edits were stably inherited by subsequent generations, with no transgene or viral residues detected in the plants. Yoshida et al. delivered Cas9 protein and sgRNA to the shoot apical meristem of Nicotiana benthamiana with tobacco ringspot virus (TRSV) vectors introduced via Agrobacterium tumefaciens[58]. Editing efficiency was improved by knockdown of NbRDR6, or expression of the TRV 16K suppressor. They also used a TRSV vector, and a TRV vector to deliver SpCas9 and sgRNA, respectively, to overcome sgRNA sequence bias. Plants inoculated with the viral vectors occasionally produced gene-edited lateral shoots, with a transgenic gene editing frequency of up to 100%.
Negative-strand RNA viruses offer larger payloads and can accommodate complete CRISPR/Cas cassettes, removing the need for pre-transformed Cas9 lines. Ma et al. inserted Cas9 and sgRNA between the N and P genes of Sonchus yellow net rhabdovirus (SYNV) genome[59]. The gRNA employed a tRNA-gRNA-tRNA (tgtRNA) structure, utilizing endogenous tRNA processing enzymes for precise release of the mature gRNA. Plants were infected by agrobacterium infiltration or mechanical friction inoculation, allowing systemic spread throughout the plant. The virus does not integrate into the host genome, ensuring non-transgenic progeny. Liu et al. developed a transient delivery system based on the broad-host-range tomato spotted wilt virus (TSWV), which can be used to stably deliver a series of large CRISPR/Cas nuclease molecules to a variety of crop varieties[60].
Pollen-mediated genome editing
-
Pollen-mediated genome editing leverages pollen from haploid inducer (HI), or doubled haploid (DH) inducer lines to deliver CRISPR/Cas genome-editing vectors, facilitating efficient trait development in various crops[62−65]. In HI-mediated delivery, CRISPR/Cas vectors are introduced via paternal pollen. Following fertilization, paternal chromosomes are typically lost during haploid induction. Subsequent chromosome doubling of haploid progeny generates transgene-free, homozygous edited plants, thereby significantly accelerating breeding cycles and eliminating transgene persistence[62−64].
In contrast, DH inducer lines enable direct production of diploid homozygous edited offspring from maternal plants, circumventing the need for artificial chromosome doubling[65]. In Brassica species, DH inducer lines have been engineered to deliver CRISPR/Cas9 vectors via pollination, targeting FAD2 homologs in cabbage (Brassica oleracea var. capitata), and rapeseed (Brassica napus). Edited progeny exhibited stable, multiplex mutations in FAD2 homologs, with maternal ploidy levels maintained, thus circumventing genotype-dependent transformation barriers, expediting functional genomics, and trait development[65].
Graft-mobile-mediated editing
-
Yang et al. developed a graft-mobile CRISPR/Cas9 system by appending mobile tRNA-like sequence (TLS) to Cas9 and guide RNA (gRNA) transcripts, facilitating the intercellular movement of these RNA molecules from transgenic rootstocks to wild-type scions upon grafting[66] (Fig. 3). This strategy enabled heritable, transgene-free gene editing in progeny, with mutation efficiencies of 0.57% for TLS1, and 0.5% for TLS2. Phenotypic analysis identified homozygous nia1 mutant traits in 0.117% of TLS1 seeds, and 0.141% of TLS2 seeds, confirming germline transmission of the edits[66]. The method obviates the need for tissue culture-based regeneration, offering significant advantages for species with recalcitrant in vitro regeneration, and potential applicability to clonally propagated crops. However, the observed editing efficiencies are currently suboptimal for widespread agricultural adoption, necessitating further optimization. While cross-species editing between Arabidopsis thaliana rootstocks, and Brassica napus scions has been demonstrated, the system's efficacy across diverse plant taxa remains unassessed. Potential physiological and genetic barriers, such as differences in vascular connectivity and RNA/protein trafficking mechanisms, may limit interspecies editing efficiency, highlighting the need for comprehensive evaluation across various crop species.
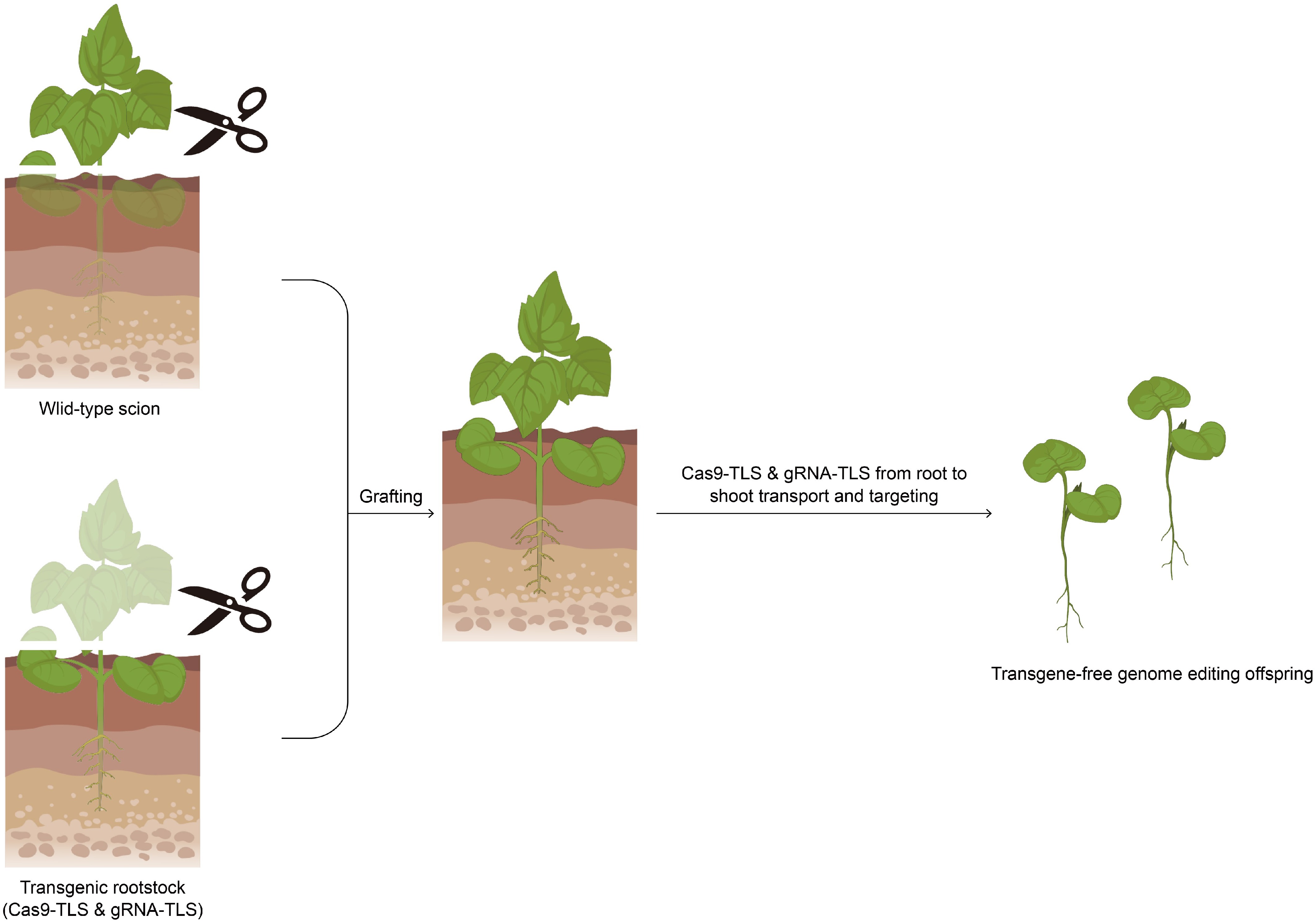
Figure 3.
Schematic diagram of graft-mobile-mediated editing.
-
Globally, the regulation of genome-edited crops varies significantly, influenced by differing approaches to biotechnology oversight. In 2020, the United States Department of Agriculture (USDA) updated its guidelines, exempting certain genome-edited crops from regulatory oversight if they do not contain foreign DNA. This product-based approach focuses on the characteristics of the final product, rather than the process used to create it. Similarly, countries such as Canada, Brazil, Argentina, Australia, Chile, Colombia, and Israel[71] have adopted product-based regulatory frameworks, assessing crops based on their end traits irrespective of the methods used for their development. In contrast, the European Union has historically employed a process-based regulatory approach, classifying gene-edited plants as genetically modified organisms (GMOs), subject to stringent regulations. However, in 2023, the European Court of Justice ruled that certain gene-editing methods, with established safety profiles, might be excluded from GMO restrictions, suggesting a move towards a more nuanced regulatory framework[71]. China has taken significant steps by approving gene-edited crops, including soybeans, wheat, corn, and rice, to enhance yields and ensure food security. The Ministry of Agriculture and Rural Affairs granted safety certificates to these crops, distinguishing gene-edited varieties from traditional GMOs[72−74].
-
T-DNA-free genome editing technology offers significant advantages. Several countries have implemented streamlined regulatory approval processes for gene-edited crops without exogenous DNA insertion, facilitating the rapid development of plant varieties with specific traits, and significantly shortening breeding cycles.
Despite the significant advantages of T-DNA-free genome editing, several limitations and challenges remain. First, methods using visible screening markers (such as fluorescent markers or pigments), are simple and intuitive to use, eliminating the need for antibiotic or herbicide resistance genes. However, these methods often require self-pollination to screen for transgene-free plants in T1 or subsequent generations, a time-consuming process that is unsuitable for perennial or asexually propagated crops. Furthermore, the expression of fluorescent markers or pigments can be easily affected by genotype or tissue background. This method is more suitable as a real-time monitoring tool, and can be combined with other strategies to improve reliability.
Second, the TKC system significantly reduces the effort required to obtain transgene-free plants, by automatically killing gametes or zygotes carrying T-DNA. However, it still requires generations of propagation to remove the transgene, and there is the issue of T-DNA 'escape'. The improved TKC2 system, incorporating visible markers, reduces the probability of escape. In contrast, the heat-induced transgene eradication system, LoxP::FRT/FLP, eliminates the transgene without propagation, making it particularly suitable for asexually propagated and perennial species, but it often leaves short remnants.
Third, transient DNA/RNA/RNP expression systems can overcome the bottlenecks of traditional stable transformation, which often require long cycles or difficult transformation. CRISPR/Cas elements exist only transiently within cells and typically do not integrate into the genome, resulting in T-DNA-free plants. However, transient DNA expression still has a small chance of integration, whereas RNA or RNP expression completely avoids this. These methods are applicable to plant species that are difficult to genetically transform through tissue culture, including perennial woody plants (such as trees), asexually propagated crops (such as bananas, potatoes), and superior inbred lines, etc. Their greatest limitations lie in delivery and regeneration efficiency. Protoplast-based methods rely on efficient regeneration systems. However, many crops struggle to regenerate from protoplasts. Particle bombardment and electroporation have limited efficiency, and they can also easily cause cell damage. Expressing regeneration-related genes such as WUS[75], BBM[76], and GRF-GIF[77] can improve regeneration efficiency to a certain extent, while delivery via viruses, pollen, or grafting can help reduce cell damage and regeneration dependency. Meanwhile, the short duration of transient expression can limit editing efficiency, which can be improved by optimizing Cas protein activity, using strong promoters, adding nuclear localization signals, and simultaneously expressing multiple sgRNAs.
Fourth, virus-mediated, pollen-mediated, and graft-mediated editing systems offer new approaches for crops with difficult regeneration or lacking stable transformation systems. Previous studies have shown that viruses can simultaneously carry Cas9 and sgRNA to achieve gene editing in wild-type plants, but in most cases, the editing is localized in somatic cells, and is difficult to transmit to germ cells. Fusion of gRNA to mobile RNA elements (such as FT mRNA or tRNA) can facilitate its entry into meristems and enhance heritability, but in some contexts, this can actually reduce editing efficiency[55]. Using smaller Cas enzymes (e.g., ISYmu1 or CasΦ) is an effective strategy for delivering sgRNA into germ cells[78]. Furthermore, agricultural applications of viral vectors still face biosafety and regulatory issues. Pollen-mediated gene editing, a cutting-edge technology, can bypass somatic cell regeneration, but its efficiency is limited by the ability of the gene editing elements to effectively penetrate the pollen wall and enter the zygote. Pollen wall structure, viability, and processing conditions vary significantly across species and have a significant impact on transfection efficiency. Using nanoparticles or liposomes to assist delivery, along with optimizing electroporation or PEG-assisted conditions, is expected to improve pollen transfection and editing success rates.
Finally, CRISPR/Cas9 is the mainstream tool for editing nuclear genes, but delivering it to organelles poses significant challenges. Cas9 and sgRNA have difficulty effectively passing through the double-layer membrane structure of mitochondria and chloroplasts. However, alternative tools that do not rely on CRISPR have been developed. Mitochondrial-targeted TALENs (mitoTALENs) have been used to edit the mitochondrial genomes of plants such as rice, rapeseed, and tomato, where male sterility exists[79]. The modular base editing system CyDENT can successfully edit the nuclear genomes of plant cells and also observed editing activity in the plant chloroplast genomes, although the efficiency in the chloroplasts is still relatively low at present[79].
-
T-DNA-free genome editing technologies offer promising avenues for developing plant varieties with specific genetic enhancements without introducing foreign DNA. This approach not only significantly shortens breeding cycles but also enables precise manipulation of traits such as herbicide and pest resistance, thereby reducing dependency on chemical pesticides and mitigating environmental pollution. Moreover, by avoiding the integration of transgenes, this technology inherently minimizes off-target effects, thereby enhancing the overall safety profile of genome editing. Such improvements directly address public concerns regarding environmental integrity and food safety, potentially facilitating broader market acceptance of gene-edited crops[80]. Combining the T-DNA-free genome editing technology (such as RNP-mediated genome editing) with base editors, prime editors, multi-gene editing systems, and large fragment genome editing systems can further expand the application scope of T-DNA-free genome editing technology, while achieving more precise and efficient gene editing. This is expected to enhance the adaptability of crops to adverse environmental conditions and contribute to the implementation of ecological restoration efforts.
However, despite these advantages, it remains essential to implement rigorous regulatory frameworks and robust detection methods to verify that genome-edited materials are devoid of unintended genetic alterations. Transparent and evidence-based policies will be crucial in building public trust and ensuring the technology's responsible integration into global agricultural practices[81], a necessity given the projected rise in global food demand to meet the needs of an estimated 9.7 billion people by 2050 (
www.fao.org/faostat/en/#data/OA , accessed from March 14, 2025). -
The authors confirm their contributions to the paper as follows: study conception and design: Xu Z, Li Y (corresponding author); draft manuscript preparation: Xu Z, Wang C; data collection and organization: Li Y (the third author), Tong Y, Zhang K, Xia X, Zhao J, Zhao Q, Xiang S, Xu L; manuscript revision and editing: Xu Z, Wang C, Li Y (the third author). All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
#Authors contributed equally: Zhangke Xu, Chunping Wang
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Yunnan Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu Z, Wang C, Li Y, Tong Y, Zhang K, et al. 2026. T-DNA-free genome editing in plants. Agrobiodiversity 3(1): 11−20 doi: 10.48130/abd-0026-0001 

T-DNA-free genome editing in plants
- Received: 20 May 2025
- Revised: 30 November 2025
- Accepted: 31 December 2025
- Published online: 25 February 2026
Abstract: Genome editing enables precise genetic modification in plants, advancing functional genomics and crop improvement. Conventional CRISPR/Cas methods often integrate foreign DNA, such as Cas9 and selection markers, into the host genome, raising biosafety and regulatory concerns. T-DNA-free genome editing technologies overcome these issues by generating edited plants without exogenous DNA integration. This review summarizes recent advances in T-DNA-free approaches, including visualized screening, Transgene Killer CRISPR (TKC) systems, transient DNA/RNA expression, ribonucleoprotein (RNP)-mediated editing, virus-mediated delivery, pollen-mediated editing, and graft-mobile systems. These strategies simplify regulation and accelerate breeding cycles. However, challenges remain in editing efficiency, delivery, and heritability, especially in asexually propagated or transformation-recalcitrant crops. The rational application and continued innovation of T-DNA-free genome editing will have significant implications for the future development of agriculture and food security.
-
Key words:
- Plants /
- Genome editing /
- CRISPR/Cas /
- T-DNA-free /
- Transformation