








-
With the continuing growth of the global population and the escalating impacts of climate change, conventional plant breeding practices are becoming insufficient to secure food supplies. Consequently, plant genetic transformation technologies have emerged as indispensable tools in modern agriculture. The pioneering work in 1983, demonstrating the regeneration of transgenic tobacco via Agrobacterium-mediated transformation, marked a significant milestone by transitioning these techniques from experimental research to practical application[1,2]. Concurrently, Chinese researchers achieved exogenous DNA integration into cotton using the pollen tube-mediated method[3]. These breakthroughs laid the foundation for the broad application of Agrobacterium-mediated transformation across diverse plant species[4]. In parallel, alternative approaches, including virus-mediated transformation[5], particle bombardment[6,7], microinjection[8], PEG-mediated transformation[9,10], and nanomaterial-mediated transformation[11,12], have been developed to further diversify and improve transformation strategies.
Despite the capability of these methods to stably integrate target genes into the nuclear genomes of various plants, achieving high transformation efficiency has remained a longstanding challenge[13]. Recent advancements have focused on refining Agrobacterium-mediated techniques to minimize or even eliminate the need for extensive tissue culture, thereby reducing genotype-dependent constraints[14−16]. These technical improvements not only broaden the applicability of existing transformation platforms but also highlight a growing shift in research priorities—from merely expanding methodological diversity to actively overcoming biological and technical bottlenecks that limit transformation success. Moreover, the inefficiencies associated with explant regeneration via organogenesis or somatic embryogenesis have been addressed through the overexpression of developmental regulators, such as WUSCHEL (WUS)[17−20], BABY BOOM (BBM)[17−19,21], WOUND-INDUCED DEDIFFERENTIATION 1 (WIND1)[22], GROWTH-REGULATING FACTOR (GRF) and chimeric gene GRF-INTERACTING FACTOR (GIF)[23], GRF5[24], DNA binding with One Finger (DOF)[25], WUSCHEL-RELATED HOMEOBOX 5 (WOX5)[26], and AINTEGUMENTA-LIKE 5 (AIL5)[27], which not only enhance transformation efficiency but also extend the spectrum of transformable species. Complementary improvements have been achieved through modifications of Agrobacterium strains[28−30], the optimization of nanomaterials[31−34], and the techniques to increase plant cell permeability[35,36].
The application of plant genetic transformation technologies for crop improvement are multifaceted, leading not only to increased yields[37−44], but also to enhanced resistance against fungal pathogens[45−47] and insect pests[46,48], and improved tolerance to abiotic stresses such as drought[49−51], salinity[49,50,52], high temperature[53,54], and low temperature[55]. A comprehensive timeline that charts the major developments in plant genetic transformation technologies is provided in Fig. 1, which serves to contextualize the rapid evolution and expanding application of these methodologies.
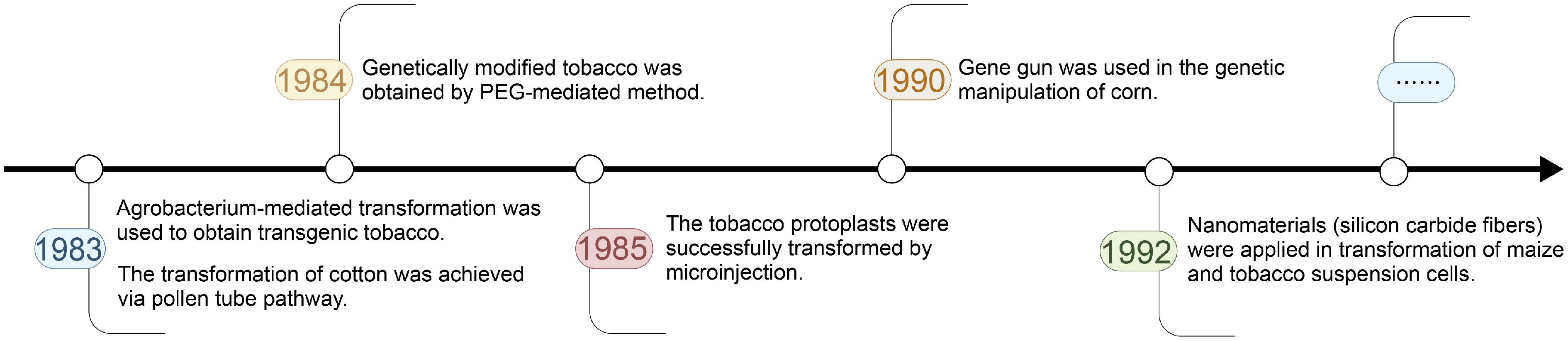
Figure 1.
The timeline of the development of plant genetic transformation technologies.
-
Genetic transformation technologies serve as critical tools for functional genomic studies in plants, enabling elucidation of target gene roles through controlled integration of exogenous DNA. Over recent decades, these methodologies have evolved into two principal categories, biologically mediated and non-biologically mediated transformation systems.
Biologically mediated transformation
-
Biologically mediated transformation predominantly includes Agrobacterium- and virus-mediated methods (Fig. 2), with Agrobacterium-mediated transformation being particularly popular due to its high transformation efficiency, low transgene copy number, compatibility with large DNA inserts, and seamless detection via selectable markers[4]. Virus-mediated transformation utilizes viruses as vectors, in which the gene of interest is assembled into the viral genome, and the recombinant virus infects host cells to accomplish transformation. This system can circumvent conventional tissue culture and plant regeneration procedures[5].
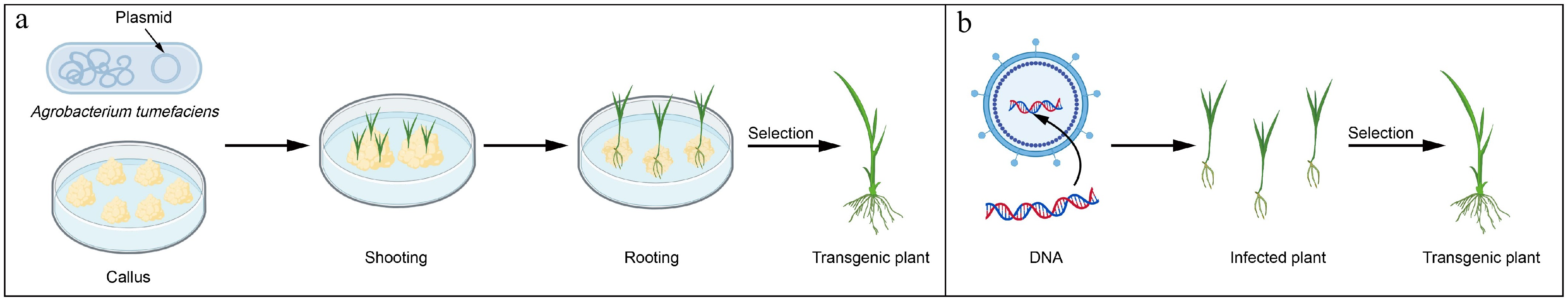
Figure 2.
Schematic diagram of biologically mediated transformation.
(a) Agrobacterium-mediated transformation. (b) Virus-mediated transformation. Part of the graphical materials were obtained fromBioGDP.com .Traditional Agrobacterium-mediated transformation protocols rely on ex planta tissue culture, in which transformed explants such as leaf discs or calli are regenerated under sterile conditions, a process that is often constrained by species-specific barriers and substantial labor requirements. Recent methodological innovations have addressed these bottlenecks through alternative strategies. In tillering species, infection of root-stem junctions with Agrobacterium rhizogenes induces transgenic root formation, enabling direct shoot regeneration from transformed root primordia[14]. Similarly, tissue culture-independent transformation in rice (Oryza sativa) has been achieved via Agrobacterium inoculation of embryonic coleoptiles, capitalizing on the pluripotency of meristematic cells[15]. For soybean (Glycine max), the Genotype-independent Fast Transformation (GiFT) method combines ultrasonic permeabilization of germinated embryos with Agrobacterium co-culture, followed by direct soil transplantation, enabling transgenic line production within 35 d while minimizing tissue culture reliance[16].
In addition, genomic data have been used to engineer free-living or endosymbiotic rhizobia with the requisite machinery for T-DNA delivery into plant cells. This approach may help to attenuate host defense responses during transformation, as plants are generally less prone to mount strong immune reactions against these typically non-pathogenic bacteria[56]. Many examples of T-DNA delivery by rhizobial strains have been reported, including for Rhizobium trifolii, Phyllobacterium myrsinacearum, Sinorhizobium meliloti, and Mesorhizobium loti[57].
Non-biologically mediated transformation
-
Non-biological transformation methods circumvent the reliance on living vectors by employing physical or chemical mechanisms to deliver exogenous DNA into plant cells (Fig. 3). While these approaches offer genotype flexibility, they often result in higher transgene complexity, including rearrangements, multi-copy insertions, or integration of fragmented DNA.
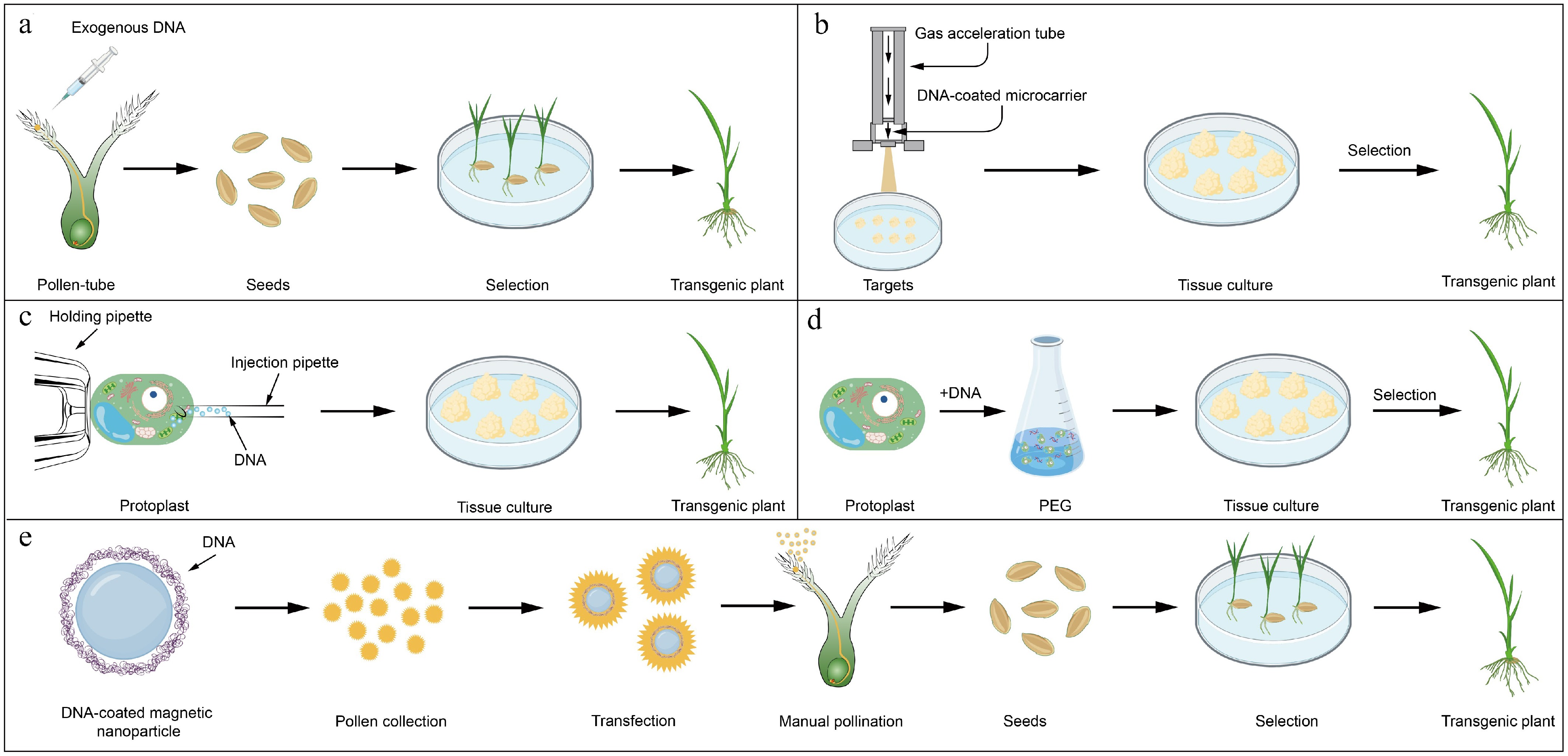
Figure 3.
Schematic diagram of non-biologically mediated transformation.
(a) Pollen tube-mediated transformation. (b) Particle bombardment. (c) Microinjection-mediated transformation. (d) PEG-mediated transformation. (e) Nanomaterial-mediated transformation. Part of the graphical materials were obtained fromBioGDP.com .Pollen tube-mediated transformation
-
The pollen tube-mediated method leverages the natural fertilization process to introduce exogenous DNA into developing embryos. Following pollination, DNA applied to stigmatic surfaces infiltrates ovules through microchannels formed by elongating pollen tubes. During syngamy, the exogenous DNA integrates into zygotic or early embryonic genomes, bypassing cell wall barriers at developmental stages characterized by active DNA replication and recombination. Although this method eliminates tissue culture requirements and accommodates diverse genotypes, its efficiency remains variable due to stochastic DNA uptake during double fertilization[3]. The plant genetic transformation through the pollen tube pathway does not involve cell culture or plant regeneration processes, making it suitable for species with poor regeneration capabilities or genotype limitations.
Particle bombardment
-
Genetic transformation by particle bombardment utilizes pressurized helium to accelerate DNA-coated metallic microparticles (0.6–1.2 µm gold or tungsten) into target tissues. The transient pores generated in cell membranes and walls permit nuclear entry of exogenous DNA, which integrates via non-homologous recombination. Transformation efficiency is influenced by several parameters, including microparticle velocity (controlled by rupture disk pressure, 450–2,200 psi), target distance (6–12 cm), and the regenerative capacity of bombarded tissues. Originally demonstrated in maize (Zea mays)[6], this method has been adapted for recalcitrant species such as wheat (Triticum aestivum), soybean (Glycine max), and poplar (Populus spp.), where genotype-independent delivery is critical[7]. When applying to different species, parameters, such as the number of bombardments, pressure, and distance, need to be optimized, all of which affect the final transformation efficiency.
Microinjection-mediated transformation
-
Microinjection employs glass capillaries to deliver femtoliter-scale volumes of DNA directly into subcellular compartments, including nuclei of protoplasts, zygotes, or meristematic cells. This technique enables precise co-delivery of multiplexed biomolecules, such as CRISPR-Cas9 ribonucleoproteins or synthetic mRNA, into single cells. However, its utility is constrained by the need for specialized micromanipulation systems, high-throughput imaging to monitor injection success, and the risk of cell lysis during membrane penetration[8]. Notably, it also allows for the simultaneous delivery of diverse substances, including nucleic acids, proteins, and synthetic materials, into the same cell, enabling efficient transformation with multiple agents. However, since the transformation occurs at the cellular level, it requires high standards for specialized equipment and technical expertise.
PEG-mediated transformation
-
PEG-mediated transformation exploits the polymer's ability to destabilize protoplast membranes through charge neutralization, facilitating endocytosis of plasmid DNA-calcium phosphate co-precipitates. Following DNA uptake, osmotic stabilization and cell wall regeneration are induced in protoplasts cultured on auxin- and cytokinin-supplemented media, enabling subsequent plant regeneration[9]. With ongoing refinements, PEG-mediated transformation has become a standard technique in plant genetic engineering[10].
Nanomaterial-mediated transformation
-
Engineered nanomaterials enable targeted DNA delivery through physicochemical interactions, including electrostatic adsorption, van der Waals forces, or ligand-receptor binding. For instance, silicon carbide whiskers, vortex-agitated in cell suspensions, create localized cell wall abrasions, allowing plasmid delivery into regenerable tissues, as first demonstrated in tobacco and maize suspension cultures cell[11]. Similarly, superparamagnetic iron oxide nanoparticles (SPIONs) conjugated with plasmid DNA have been utilized in magnetofection, where oscillating magnetic fields drive nanoparticle internalization into pollen grains. Subsequently, stable genetic transformation is achieved by obtaining transgenic seeds directly through artificial pollination during the plant's natural reproductive process[12]. The combination of nano-magnetic transformation with pollen-mediated delivery circumvents major constraints of traditional genetic transformation—especially the requirements for tissue culture regeneration and the narrow host range. This strategy substantially enhances transformation efficiency, accelerates transgenic plant production, and supports high-throughput and multiplex gene delivery.
Although Agrobacterium-mediated transformation is often the more reliable choice for generating stable transgenic lines due to its technical maturity and high genetic stability, nanomaterial-based systems offer greater potential and flexibility for recalcitrant species or applications requiring large-scale, high-throughput gene function screening.
Plant genetic transformation technologies have made substantial contributions to societal development and economic advancement. Although a range of transformation methods has been widely applied in functional genomics research and crop improvement, significant challenges and limitations remain. Agrobacterium-mediated transformation is widely employed due to its stability and well-established operational framework; however, its host range is limited and often constrained by genotype dependence[10]. Virus-mediated transformation can circumvent tissue culture and plant regeneration processes, but it is largely restricted to transient expression and raises potential biosafety concerns. Some plant viruses can be transmitted to the next generation through seeds, leading to adverse effects on plant growth and yield[5]. Although hybridization-based detoxification is effective, it is time-consuming and labor-intensive, and thus, rapid and efficient virus elimination strategies remain to be developed. The pollen-tube pathway, constrained by the natural flowering period, has achieved success in introducing exogenous genes only in a few monocotyledonous and dicotyledonous species[13]. Microinjection enables direct gene delivery; nevertheless, its high technical complexity limits its suitability for large-scale applications. Particle bombardment is prone to generating high chimerism and reduced genetic stability[58]. PEG-mediated transformation is applicable to diverse plant species, yet it frequently results in cellular damage. Recently, nanomaterial-mediated strategies have emerged, offering improved transformation efficiency, shortened cultivation cycles for transgenic plants, and enabling high-throughput as well as multigene delivery. However, nanomaterial-mediated plant transformation also raises biosafety concerns, particularly regarding their environmental persistence, potential toxicity, unintended dissemination, off-target genetic delivery, and the current lack of comprehensive regulatory frameworks to evaluate their long-term ecological and health impacts. Despite the need for additional optimization to expand their applicability across a broader range of species, nanomaterial-based approaches represent a revolutionary delivery method[59]. In summary, future advancements in plant transformation technologies must achieve a balance between efficiency and stability, while simultaneously addressing ethical and biosafety considerations, thereby providing more reliable and responsible tools for both crop improvement and fundamental research.
-
Recent advances have demonstrated that the manipulation of developmental regulators can markedly enhance plant regeneration in both monocotyledonous and dicotyledonous species. By promoting cellular dedifferentiation, establishing totipotency, and stimulating somatic embryogenesis, these regulators significantly improve transformation efficiencies and broaden the range of transformable genotypes.
WUSCHEL and BABY BOOM
-
The synergistic action of key transcription factors such as WUSCHEL (WUS) and BABY BOOM (BBM) has been pivotal in advancing plant transformation protocols. WUS maintains cells in an undifferentiated state, thereby facilitating the integration of exogenous DNA, while BBM actively triggers somatic embryogenesis and cell proliferation[60,61]. Their efficacy in enhancing plant regeneration has been verified in multiple species, including maize[17−19], rice[17], sorghum[17,20], and apple[21]. For instance, the overexpression of these genes in maize enabled the transformation of previously recalcitrant inbred lines, such as the Pioneer line PHH5G, which yielded transgenic calli from over 40% of explants, a substantial improvement over traditional methods like particle bombardment or Agrobacterium-mediated transformation[17]. To mitigate the adverse developmental effects associated with their constitutive expression, strategies such as employing inducible promoters (e.g., Hsp17.7 and Hsp26) to drive early expression of site-specific recombinases (e.g., Cre) and using tissue- and time-specific promoters (Axig1 and PLTP) have been developed. These innovations allow for genotype-independent transformation that eliminates the need for a callus culture stage[18,19].
WOUND-INDUCED DEDIFFERENTIATION 1
-
The AP2/ERF transcription factor WOUND-INDUCED DEDIFFERENTIATION 1 (WIND1) plays an essential role in initiating cellular dedifferentiation and callus formation[62]. WIND1 activates key components of the cytokinin signaling pathway, particularly ARR1 and ARR12, which are critical for callus induction. In Arabidopsis, its ectopic expression has been shown to enable bud regeneration from explants without the need for conventional stimuli such as wounding or auxin application. Moreover, the sequential activation of WIND1 followed by the embryonic regulator LEAFY COTYLEDON2 further enhances embryogenic callus production, thereby demonstrating the value of combined transcription factor strategies for organ-specific regeneration[22,63].
GRF-GIF Chimeras
-
Chimeras combining the GROWTH-REGULATING FACTOR (GRF) with its cofactor GRF-INTERACTING FACTOR (GIF) have proven effective in enhancing plant regeneration and transformation efficiencies. These constructs leverage the conserved QLQ and WRC domains of GRF to mediate protein-protein interactions and DNA binding, facilitating the formation of a transcriptional activation complex essential for plant development. In soybean, the overexpression of the GRF3-GIF1 chimera significantly improved regeneration and transformation efficiency across various cultivars, including those traditionally recalcitrant to transformation, such as DN252, SN14, and ZJ602[23]. Similar successes have been observed in other species; for instance, the GRF4-GIF1 chimera enhanced regeneration in wheat, maize, and watermelon, demonstrating its broad applicability across different plant species[64].
GRF5
-
Similarly, the overexpression of GRF5 alone has been found to accelerate bud regeneration and significantly increase genetic transformation efficiency across multiple species. For example, the overexpression of AtGRF5 in sugar beet facilitates the stable transformation of cultivars that have been historically difficult to transform. Introducing AtGRF5 or its homologs into canola, soybean, and sunflower markedly enhances the genetic transformation efficiency of explant tissues. Moreover, the transformation of two putative AtGRF5 homologs into maize significantly increases transformation efficiency and enables the regeneration of fully fertile transgenic plants. Collectively, these findings indicate that GRF5 overexpression improves genetic transformation efficiency across both monocotyledonous and dicotyledonous plants[24].
DOF Family
-
Transcription factors in the DOF family, characterized by a conserved zinc finger domain, are involved in regulating hormone signaling as well as key processes such as tissue differentiation and dormancy. In wheat, the overexpression of DOF family members, specifically TaDOF5.6 and TaDOF3.4, has been shown to effectively enhance callus induction rates and improve genetic transformation efficiency in cultivars, such as Fielder, KN199, and Jimai22[25].
WOX5
-
WOX5 is crucial for maintaining the stem cell niche within the root apical meristem, thereby playing a significant role in cell differentiation and regeneration. Studies have demonstrated that the expression of TaWOX5 in various wheat cultivars, such as Fielder, CB037, Zhongmai895, Jimai22, Lunxuan987, and Jing411, can overcome genotype-dependent limitations and significantly enhance transformation efficiency. Further investigations have indicated that TaWOX5 can also improve the genetic transformation efficiency in other monocotyledons, such as Triticum monococcum, rye, triticale, barley, and maize[26].
AINTEGUMENTA-LIKE 5
-
AINTEGUMENTA-LIKE 5 (AIL5) transcription factors are integral to regulating embryo development, organogenesis, and meristem maintenance (Table 1). Overexpression of MdAIL5 in apple has been shown to markedly increase the regeneration efficiency of adventitious shoots from leaf explants, even in tissues exhibiting a high degree of maturity[27].
Table 1. Developmental regulators applied in plant genetic transformation.
Genes Species Delivery context Expression strategy Transformation effects Ref. WUS, BBM Maize Agrobacterium Constitutive expression Significantly increases transformation efficiency and reduces genotype dependence [17] Rice Sorghum Maize Agrobacterium Specific expression Transformation frequency varied between inbred lines with averages ranging from 9% for one ear from B73 to 224% for one ear from PHH5G [18] Maize Agrobacterium Specific expression The transformation frequency in the T0 generation is 29%–69% [19] WUS Sorghum Agrobacterium Specific expression Wus2 enhances both transformation efficiency and CRISPR editing frequency in sorghum [20] BBM Apple Agrobacterium Constitutive expression The transformation efficiency was 3.68%–28.15% [21] WIND1 Arabidopsis Agrobacterium Constitutive expression Greatly enhances de novo shoot regeneration [22] Rapeseed GRF3-GIF1 Soybean Agrobacterium Constitutive expression The average transformation efficiency with the GRF3-GIF1 chimera was 2.74-fold higher than for the empty vector control [23] GRF5 Sugar beet Agrobacterium Constitutive expression Transformation efficiency was increased 6-fold [24] Canola Promoted the proliferation of transgenic callus cells Soybean Promoted the production of transgenic shoots Sunflower Promoted the production of transgenic shoots Maize Transformation frequencies increase by three times for inbred line A188 TaDOF5.6, TaDOF3.4 Wheat Agrobacterium Constitutive expression The transformation efficiency was improved by about 17%–34% [25] WOX5 Wheat Agrobacterium Constitutive expression Successfully transformed 31 common wheat cultivars, overcoming genotype dependency [26] Triticum monococcum PI428182 showed a significant increase in transformation efficiency to 94.5% ± 8.2% Rye Transformation efficiency of 7.8% in Lanzhou Heimai Triticale Transformation efficiency of 16.4%–53.3% in Linfen45, ZS3297, ZS1257, and ZS3224 Barley Transformation efficiency of 10.4%–88.4% in barley Maize Transformation efficiency of 19.2%–38.4% in B73 and A188 AIL5 Apple Agrobacterium Constitutive expression Significantly improved the regeneration efficiency of adventitious shoot [27] Modification of Agrobacterium
-
Agrobacterium-mediated transformation remains the cornerstone of plant genetic engineering, yet its efficacy is constrained by the innate resistance of many species and varieties to T-DNA delivery and integration[29]. Recent efforts have thus focused on augmenting A. tumefaciens virulence and mitigating plant defense activation via expanded virulence loci and heterologous secretion systems[28,65]. Ternary vector systems incorporating a third helper plasmid enriched for vir gene clusters have markedly improved transformation outcomes in recalcitrant species. One exemplar is the pKL2299 helper plasmid, which harbors an expanded complement of vir genes from the Ti plasmid pTiBo542 and utilizes an RK2 replication origin for compatibility; its deployment in A. tumefaciens LBA4404 has more than doubled maize transformation frequencies in multiple elite inbred lines[28]. Such enhanced vir gene expression compensates for the limitations of binary vectors and streamlines high-throughput plant transformation workflows. Engineering A. tumefaciens to express a functional Type III Secretion System (T3SS) derived from Pseudomonas syringae enables the direct translocation of Type III effectors (T3Es) into plant cells, thereby suppressing basal immunity and elevating transformation efficiencies[29]. Moreover, this secretion platform is versatile enough to export eukaryotic proteins: translocation of histone H2A-1 into plant nuclei further amplifies T-DNA integration rates, illustrating the potential to deliver non‐canonical transformation enhancers[29]. Finally, recognition of the bacterial elongation factor Tu (EF-Tu) by plant pattern‐recognition receptors—particularly the EF-Tu receptor (EFR) in Arabidopsis—activates PAMP‐triggered immunity that diminishes AMT efficiency[66]. To evade this defense, endogenous A. tumefaciens EF-Tu genes have been replaced with DC3000 EF-Tu variants harboring divergent epitopes that escape EFR detection; engineered strains exhibit significantly elevated transformation rates in Arabidopsis thaliana and Camelina sativa compared to wild-type controls[30].
Improvement of nanomaterials
-
Recent advances in nanotechnology have yielded a suite of nanoparticle platforms capable of bypassing the physical barrier of the plant cell wall to deliver DNA, RNA, and proteins with high efficiency and minimal toxicity[67]. Among these, carbon‐based nanocarriers, magnetic nanoparticles, and mesoporous silica nanoparticles (MSNs) have been most extensively validated for both transient and heritable gene modulation in diverse crop species, offering genotype‐independent and tissue‐culture-free transformation modalities[67,68]. Polyethyleneimine-modified single-walled carbon nanotubes (PEI-SWCNTs) exploit electrostatic adsorption of nucleic acids to their cationic surface, protecting cargo from nuclease degradation while facilitating cellular uptake via passive diffusion and endocytosis. In protoplasts of Nicotiana benthamiana, Eruca sativa, Triticum aestivum, and Gossypium hirsutum, PEI‐SWCNT-DNA complexes achieved transient gene expression, with protein levels comparable to or greater than PEG‐mediated transfection, and without compromising cell viability[31]. Modified carbon dots (MCDs) synthesized via the electrochemical etching method have been shown to adsorb and condense plasmid DNA into stable complexes that penetrate seed coats, enabling whole‐seed transformation in wheat and its wild relatives. By co‐incubating wheat seeds with MCD-DNA conjugates under mild agitation, germinated seedlings exhibited robust, transient expression of reporter genes (e.g., GFP, LUC, and GUS) in plants, without detectable phenotypic abnormalities or growth retardation[34]. This method supports rapid functional genomics and epigenomics assays in recalcitrant genotypes, circumventing both Agrobacterium dependence and protoplast isolation[34]. Magnetofection utilizes superparamagnetic Fe3O4 nanoparticles coated with plasmid DNA to transfect pollen grains through their germination pores. In maize inbred lines traditionally recalcitrant to tissue culture, pollen pretreated to open the operculum was incubated with DNA-MNP complexes under a magnetic field at low temperatures. Resulting T1 progeny harbored transgenes (e.g., EGFP, GUS) at frequencies of 5%–16%, and exhibited stable, heritable expression without callus regeneration, demonstrating a fully genotype‐independent, culture‐free transformation system[32]. Finally, mesoporous silica nanoparticles (MSNs) have emerged as versatile carriers for topical RNA interference (RNAi) in rice. Foliar spraying of MSN-siRNA complexes effectively delivered siRNA, resulting in up to 70% gene silencing of the PDS gene and 75% silencing of the transgenic Ruby gene. Additionally, foliar application of MSNs loaded with small interfering RNAs (siRNAs) against seed shattering genes in wild rice Oryza alta achieved gene silencing efficiencies up to 49.4%. This spray‐on approach enabled transient, nonpermanent trait modulation without genetic integration or tissue damage, providing a sustainable tool for trait engineering in field settings[33].
Increase of plant cell permeability
-
Surface-active agents are widely employed to lower the interfacial tension between bacterial suspensions and plant tissues, thereby enhancing Agrobacterium infiltration and T-DNA transfer. Non-ionic surfactants such as Silwet® L-77 and Tween-20 facilitate the penetration of bacterial cells through the waxy cuticle into the apoplast by reducing water surface tension[69]. In transient assays, Triton X-100 at 0.005%–0.01% has been shown to increase reporter gene expression in both herbaceous and woody species, with optimal concentrations dependent on the recipient tissue[70,71]. Moreover, the addition of acetosyringone and Silwet-L77 to Agrobacterium inoculation media significantly enhanced transformation efficiency in wheat, underscoring the utility of finely tuned surfactant treatments for recalcitrant cereal genotypes[35]. Physical treatments employing ultrasonic energy, collectively termed sonication-assisted Agrobacterium-mediated transformation (SAAT), exploit cavitational effects to produce micro-lesions in the plant cell wall and plasma membrane. In rubber tree (Hevea brasiliensis), brief sonication of embryogenic calli prior to co-culture increased GUS reporter activity and stable integration events by up to threefold, as cavitation-enabled Agrobacterium to access inner cell layers more effectively[36,72]. Mechanistic studies indicate that ultrasound frequencies of 20–40 kHz transiently disrupt cell wall architecture without compromising viability, facilitating both bacterial ingress and T-DNA uptake[73,74]. Building on these principles, the soybean GiFT method incorporates a controlled sonication step to wound-imbibed seed explants, yielding genotype-independent transformation with a 35-d turnaround from infection to plant establishment[16]. By integrating optimized surfactant concentrations with targeted ultrasonic treatments, researchers can transiently modulate plant cell permeability, resulting in substantial improvements in both transient expression and stable transgenic event frequencies. These combined strategies offer a versatile, tissue-culture-free means to overcome host barriers and broaden the applicability of Agrobacterium-mediated transformation across a wider array of plant species and genotypes (Table 2).
Table 2. Other methods to improve the efficiency of plant genetic transformation.
Methods Details Species Ref. Modification of Agrobacterium Introducing helper plasmids Maize [28] Modifying Agrobacterium tumefaciens with T3SS Arabidopsis thaliana, Nicotiana benthamiana, Wheat, Alfalfa, Switchgrass [29] Expressing the DC3000 EF-Tu variants Arabidopsis thaliana, Camelina sativa [30] Improvement of nanomaterials Polyethyleneimine (PEI)-modified single-walled carbon nanotubes (PEI-SWCNTs) Nicotiana benthamiana, Arugula, Wheat, Cotton [31] Modified carbon dots (MCDs) Wheat [34] Ferroferric oxide (Fe₃O₄) magnetic nanoparticles (MNPs) Maize [32] Mesoporous silica nanoparticles (MSNs) Oryza alta [33] Increase of plant cell permeability Surfactants Wheat [35] Sonication Soybean, Rubber tree [16,36] -
Genetic enhancement of photosynthetic capacity, nutrient uptake, stress resilience, and developmental timing through targeted transgene expression has yielded substantial increases in grain yield across staple cereals and legumes. In rice (Oryza sativa), upregulation of the plasma-membrane H+-ATPase gene OSA1 enhances both nitrogen and carbon assimilation, boosting nitrogen fertilizer utilization by 46% and increasing grain yield by 33% relative to wild-type controls in multi-location trials[37]. Similarly, constitutive overexpression of the dehydration-responsive element-binding transcription factor OsDREB1C drives a transcriptional program that augments photosynthetic capacity and nitrogen use efficiency, resulting in field-measured yield gains of 41%–68% and shorter growth duration under both irrigated and water-limiting conditions[38,75]. Maize (Zea mays) yield has also been improved through modulation of both cell-wall dynamics and hormonal signaling. Transgenic elevation of the α-expansin ZmEXPA5 accelerates cell-wall loosening, shortens the anthesis-silking interval, and confers drought resilience; under both water-deficit and well-watered regimes, ZmEXPA5-overexpressing lines exhibit significant grain yield improvements[39]. Likewise, overexpression of the strigolactone biosynthesis enzyme ZmCCD8 increases kernel weight as well as sucrose and free amino acid concentrations in harvested grain, reflecting enhanced assimilate allocation during grain fill[42,76]. In soybean (Glycine max), overexpression of the chloroplast protease subunit GmFtsH25 strengthens thylakoid grana stacking and elevates photosynthetic efficiency, leading to marked increases in seed yield and biomass accumulation in field assays[40]. Parallel work on the CONSTANS-like transcription factor GmCOL3a demonstrates that its ectopic expression increases branch number and delays floral transition, thereby extending the vegetative phase and amplifying per‐plant yield in diverse genetic backgrounds[41]. In wheat (Triticum aestivum), the CONSTANS‐like gene TaCOL-B5 has been shown to modulate spike architecture by increasing spikelet node number per spike, thereby substantially elevating grain yield in both greenhouse and field evaluations[43]. Heterologous expression of the soybean DREB1 transcription factor GmDREB1 enhances osmotic adjustment, photosynthetic efficiency, and melatonin‐mediated stress signaling, translating to grain yield increases of 4.8%–18.4% under limited-irrigation field conditions[44].
Enhancing resistance to biotic stresses
-
Transgenic deployment of broad‐spectrum defense genes has yielded notable successes across diverse crop species. Overexpression of rice BROAD-SPECTRUM RESISTANCE2 (BSR2/CYP78A15) in tomato 'Micro-Tom' conferred robust tolerance to the necrotrophic fungi Botrytis cinerea and Rhizoctonia solani, as well as to the bacterial pathogens Pseudomonas syringae and Ralstonia pseudosolanacearum, without observable developmental abnormalities[45]. In soybean (Glycine max), overexpression of the novel plasma membrane protein GmDR1 imparts broad‐spectrum immunity against both pathogens and pests. Transgenic lines exhibited significantly enhanced resistance to Fusarium virguliforme (sudden death syndrome), soybean cyst nematode (Heterodera glycines), spider mites (Tetranychus urticae, Koch), and soybean aphids (Aphis glycines, Matsumura)[46]. In rice (Oryza sativa), the transcription factor OsGRF6, originally characterized for its role in yield enhancement, has been repurposed to bolster disease resistance. Constitutive expression of OsGRF6 resulted in a greater than 50% reduction in lesion length following inoculation with Xanthomonas oryzae pv. oryzae (Xoo), the causal agent of bacterial blight, and mechanistic analyses revealed that OsGRF6 directly upregulates OsYUCCA1 and OsWRKY82, thereby synchronizing auxin biosynthesis and defense signaling pathways to enhance resistance[47]. Targeting chewing insects, the jasmonate signaling repressor GhJAZ24 was engineered for damage‐induced expression in cotton, resulting in potent activity against Helicoverpa armigera (cotton bollworm) and Spodoptera frugiperda (fall armyworm)[48]. This iJAZ strategy has also been translated into rice and maize, where GhJAZ24‐based constructs conferred effective insecticidal activity against lepidopteran pests, demonstrating the broad applicability of JAZ‐mediated defense modules across monocot and dicot crops[77].
Enhancing tolerance to abiotic stresses
-
Transgenic overexpression of transcription factors (e.g., GmNF-YC9, TaASR1-D, GhABF3, SlWRKY81, TaNACα18, OsEDS1) and key enzymes (GmSQE1, ScGolS1) has been demonstrated to confer enhanced tolerance to drought, salinity, heat, and cold across major crop species. These modifications improve antioxidant defenses, osmotic adjustment, hormonal signaling, and reactive oxygen species (ROS) scavenging, resulting in greater physiological resilience and yield stability under stress conditions[49−55].
In soybean, constitutive expression of the CCAAT-box transcription complex subunit GmNF-YC9 and the sterol-biosynthesis enzyme GmSQE1 significantly enhances drought tolerance by mitigating oxidative damage, with field trials across two seasons showing increased yield per plant under water‐deficit conditions[51]. Similarly, in wheat, overexpression of the abscisic acid-stress-ripening protein TaASR1-D improves osmotic adjustment and antioxidant capacity under both drought and saline conditions, as evidenced by higher relative water content and reduced ROS accumulation in transgenic lines compared to wild-type controls[49]. In upland cotton, GhABF3, an ABA-responsive element‐binding factor, when overexpressed, upregulates downstream stress-responsive genes, reduces cellular oxidation, decreases leaf water loss, and leads to longer roots and less wilting under drought and salinity stress[50]. SlWRKY81 in tomato functions as a positive regulator of proline biosynthesis and antioxidant defenses; its overexpression significantly enhances tomato tolerance to saline-alkali stress[52]. High‐temperature tolerance has been achieved in wheat through TaNACα18, a NAC family transcription factor whose overexpression modulates cell‐wall remodeling and defense gene expression, conferring thermotolerance without developmental penalties[54,78]. In rice, the immune activator OsEDS1 promotes H2O2 scavenging by stimulating catalase activity via direct catalase association, thereby enhancing heat‐stress tolerance and safeguarding yield traits under elevated temperatures[53]. Finally, cold tolerance in potato has been markedly improved by overexpressing ScGolS1, a galactinol synthase from Solanum commersonii. Transgenic Solanum tuberosum lines exhibit reduced electrolyte leakage, lower malondialdehyde content, decreased oxidative staining, and higher survival rates following freezing stress, underscoring the role of raffinose‐family oligosaccharides in cellular protection during low‐temperature exposure[55].
-
The first commercial release of the Flavr Savr tomato marked a significant milestone in crop biotechnology, and by the mid-1990s, a suite of transgenic events, including Bt cotton, insect-resistant maize, Bt potatoes, Roundup Ready soybean, and herbicide-tolerant canola, had been deregulated for field cultivation in major agricultural markets[79]. Over the following 28 years, the global area devoted to biotech crops increased substantially, with genetically modified (GM) crops becoming among the most rapidly adopted technologies in modern agriculture[80].
By 2023, GM varieties were cultivated in 27 countries, covering approximately 206.3 million hectares, while a total of 76 countries and regions had approved at least one biotech crop for cultivation, import, or processing[80,81]. National regulatory agencies continued to expand approval portfolios: in 2023 alone, over 80 new cultivation approvals, predominantly for maize and soybean events, were granted across key agricultural markets. The trait spectrum of approved GM events remains dominated by herbicide tolerance and insect resistance, which together account for the majority of commercialized transgenic traits globally[82].
In major producing countries, adoption rates are notably high: in the United States, GM varieties constitute over 90% of maize, upland cotton, and soybean acreage; Canada reports that more than 80% of its canola, corn, and soybean plantings are GM; and in Brazil, major commodity crops such as soybean, cotton, and maize are predominantly transgenic under commercial production.
The integration of GM crops into international trade networks has been associated with improved land-use efficiency, reductions in pesticide application, and greater stability in commodity supply, thereby contributing to food security in importing countries[83]. Regulatory harmonization, exemplified by the European Union's approval of 13 new GM events for food and feed in 2023, illustrates increasing alignment among major markets, facilitating smoother cross-border flows of biotech commodities.
Recent advances in CRISPR/Cas9 transformation technologies have further expanded the toolbox for crop improvement. For instance, the newly developed pHNRhCas9NG system exhibited nearly 100% genome editing efficiency in the tested species, tomato[84]. The FrCas9-based CBE V2.1 has been demonstrated to be a highly efficient cytosine base editor, capable of achieving precise C-to-T conversions in plants[85]. Moreover, studies have indicated that selecting appropriate promoters for CRISPR/Cas systems is critical for ensuring efficient gene editing. Recent studies have demonstrated that the promoter of the PCE8 gene can drive the Cas9 nuclease to produce efficient editing in dicotyledons[86].
Recent advances in delivery technologies have substantially accelerated progress in plant genome editing. Engineered RNA viruses have emerged as efficient CRISPR carriers: tomato spotted wilt virus (TSWV)–derived vectors enable the delivery of Cas9, Cas12a, and base editors, generating heritable, virus-free mutants across multiple crops[87]. Similarly, the barley yellow striate mosaic virus (BYSMV) system supports transgene-free, culture-independent heritable editing in wheat[88]. In addition, nanoparticle complexes composed of cationic polymers and nucleic acids have successfully delivered Cas9-sgRNA into maize cells to achieve gene knockout[89]. Collectively, these studies highlight the convergence of next-generation editing tools with innovative transformation strategies.
Looking forward, the convergence of CRISPR/Cas-based genome editing with traditional transgenic approaches promises rapid trait stacking and precision improvements, including drought resilience, enhanced nutrient profiles, and resistance to insect pests. Continued innovation aims to develop 'smart' crops combining high yield, broad-spectrum biotic stress resistance, and robust abiotic stress tolerance, thereby delivering the next generation of resilient cultivars to meet the twin challenges of a growing population and an increasingly unpredictable climate.
-
The authors confirm their contributions to the paper as follows: study conception and design: Zhang K, Li Y; draft manuscript preparation: Zhang K, Wang C; data collection and organization: Li Y, Tong Y, Xu Z, Xia X, Zhao J, Zhao Q, Xiang S, Xu L; manuscript revision and editing: Zhang K, Wang C, Li Y. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
#Authors contributed equally: Kefei Zhang, Chunping Wang
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Yunnan Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang K, Wang C, Li Y, Tong Y, Xu Z, et al. 2026. The applications of plant genetic transformation technology and progress of relevant research. Agrobiodiversity 3(1): 1−10 doi: 10.48130/abd-0025-0013 

The applications of plant genetic transformation technology and progress of relevant research
- Received: 29 May 2025
- Revised: 25 November 2025
- Accepted: 18 December 2025
- Published online: 20 January 2026
Abstract: This review provides a systematic overview of the principles and application scope of plant genetic transformation technologies, with particular emphasis on strategies to improve transformation efficiency and the potential applications of these technologies in crop genetic improvement. The paper focuses on two main categories of transformation methods: biologically mediated and non-biologically mediated ones. Various genetic transformation techniques, including Agrobacterium-mediated transformation, virus-mediated transformation, particle bombardment (gene gun), pollen tube-mediated method, microinjection-mediated transformation, polyethylene glycol (PEG)-mediated transformation, and nanomaterial-mediated transformation, are discussed in detail. In terms of improving genetic transformation efficiency, specific techniques, such as the regulation of developmental regulators, the modification of Agrobacterium strains, the optimization of nanomaterials, and the increase of plant cell permeability, can significantly improve plant regeneration efficiency, enabling the transformation of recalcitrant plant genotypes. Finally, the applications of genetic transformation technology in improving crop yield and tolerance to biotic and abiotic stresses are highlighted. This article aims to provide valuable theoretical references and practical insights for the in-depth investigation and innovative application of plant genetic transformation systems.