










-
The ovary is a vital component of the chicken's reproductive system, with egg-laying rates depending on follicle development and ovulation. Post-maturity, the ovary harbors a spectrum of follicle sizes, categorized as follows: primordial follicles (< 0.08 mm), primary follicles (0.08−1 mm), undifferentiated pre-hierarchical follicles (1−8 mm), and preovulatory/hierarchical follicles (> 9 mm)[1]. Despite the abundance of follicles, most undergo atresia, with only a select few maturing into preovulatory status. Follicle selection is critical as it determines the fate of follicles in the ovary—either advancing in the follicular hierarchy or undergoing atresia. The small yellow follicle (SYF) stage marks a critical juncture in follicle selection, with granulosa cells (GCs), the oocyte-adjacent somatic cells, playing a nurturing role in follicle maturation. GC differentiation is a hallmark of follicle selection, as they synthesize and secrete progesterone and express high levels of FSHR[2], which plays an indispensable role in follicle selection.
Follicle selection in chickens is a complex process coordinated by multiple factors, including the neuroendocrine system, hypothalamic-pituitary-gonadal axis, and the oocytes themselves. The hypothalamus secretes GnRH, prompting the pituitary to release FSH, which then targets FSHR on ovarian GCs, igniting downstream pathways and enhancing gene transcription vital for steroidogenesis, including cytochrome P450 family 11 subfamily A member 1 (CYP11A1), steroidogenic acute regulatory protein (STAR). The expression of these genes plays an important role in follicle selection and GC differentiation by regulating steroid hormone synthesis in the ovary, including progesterone (P4), and estradiol (E2)[3]. FSH and its receptor (FSHR) are closely related to chicken follicle development and egg-laying rate[4]. The forkhead box L2 (FOXL2) gene also plays an important regulatory role in follicle selection and hierarchical follicles development[5], and is significantly associated with egg-laying traits[6]. Anti-Mullerian hormone (AMH) can inhibit the downstream signaling pathway of FSHR in GCs, regulating GC differentiation and follicle selection[7]. Additionally, the oocyte itself plays a crucial role in regulating follicle selection. Oocytes release oocyte-specific factors that act as signals to ovarian GCs. For example, oocytes secrete growth differentiation factor 9 (GDF9) outside of the cells, which then binds to receptors on the surface of GCs, activates the SMAD signaling pathway, and regulates expression of downstream genes as a ligand-activated transcription factor complex, thus involving the follicle selection process[8]. However, the key genes underlying the mechanism of follicle selection in hen ovarian follicles have not been fully elucidated.
BMP15, also known as GDF9B, is conserved across various species. In sheep, BMP15 is carried by the X chromosome and is also known as FecX, which mutations are associated with follicle development abnormalities[9]. And the variation affects litter size[10]. Similarly, BMP15 activates the SMAD1/5/8 pathway to promote GC proliferation in mice[11].
In recent years, transcriptome analysis has revealed and identified many genes relevant to follicular development and selection. Several differentially expressed genes (DEGs), and were identified in different stage follicles of Jilin Black chicken Lohmann Brown layer, including STMN4, FABP3, ROBO2, RSPO4, SLC6A15, and SLITRK3[12]. In the small yellow follicles and F6 follicles of Hy-Line Brown chickens, several DEGs related to follicle selection have been identified, including VLDLR1, WIF1, NGFR, AMH, BMP15, GDF6, and MMP13[1].
To ensure the continued ovulation of follicles in the late stage of egg production and identify potential genes related to follicle selection in the late-laying period, our study, focusing on 6−8 mm follicles from late-laying period Taihang chickens, leverages RNA-seq to identify potential genes associated with follicle selection. The findings on BMP15's role in follicle selection may provide novel strategies to enhance follicular recruitment in aging hens, thereby improving egg yield during the late laying period. Follicle selection is a pivotal process that determines the egg production rate, and BMP15-mediated regulation of GC differentiation offers a molecular target for optimizing this process.
-
We collected egg production records from 704 Taihang chickens, aged 20 to 43 weeks. All birds were reared in laying battery cages under standardized environmental conditions, including identical diet, lighting, temperature, humidity, and management practices. The number of eggs laid was recorded from 20 weeks of age to ensure consistency in data collection. Groups were divided into high egg-yielding (H) and low egg-yielding (L) based on egg production. During the late egg-producing period (43 weeks of age), three Taihang hens were randomly selected and obtained with H and L. Chickens were euthanized, and yellow follicles 6 to 8 mm in diameter were categorized as SYFs, immediately snap-frozen in liquid nitrogen then stored at −80 °C for RNA extraction.
RNA extraction, library construction, and sequencing
-
Total RNA was extracted from follicles using TRIzol (TaKaRa, Dalian, China) for each sample. RNA integrity was assessed using 1% agarose gel electrophoresis, where clear 28S and 18S rRNA bands were observed without smearing. Additionally, RNA quality was further evaluated using a NanoDrop-1000 spectrophotometer (Thermo Scientific, Wilmington, USA) and Agilent Bioanalyzer 2100 (Agilent Technologies, Santa Clara, Calif., USA). RNA samples with an integrity number greater than 8.0 and an optical density ratio of 260/280 nm between 1.8 and 2.0 were selected for further experiments. During library construction, mRNA was enriched from total RNA by poly-A oligo-attached magnetic beads. Double-stranded complementary DNAs were synthesized with random hexamer primers and purified with AMPure XP beads. Then the ends of the double strand were repaired, and an A-tailed link connector was added to the 3' end, followed by PCR amplification to construct a cDNA library. The insert size was determined using Agilent 2100, and the concentration of the library was quantified by quantitative PCR. The mRNA libraries were sequenced using an Illumina HiSeq 2500 at Allwegene Technologies. The quality of RNA-Seq reads was evaluated using Fast QC.
RNA-Seq data analysis
-
To obtain clean reads, we removed low-quality reads from raw reads and filter them. And follow up analyses were based on clean reads. Concurrently, the Q20, Q30, and GC content of the clean data were calculated. The clean reads were mapped to the chicken reference genome (GRCg6a) using the sequence alignment program HISAT 2.1.0[13].
Differential expression
-
We used HTSeq (ver. 0.6.1) software to count the reads mapped to each gene. The featurecounts software was utilized to calculate the read counts of mRNAs in each sample[14]. The TPM (Transcript per Kilobase per Million mapped reads) value was used to evaluate the expression level of mRNAs in each library. The edge R package was used to identify differentially expressed genes, applying a significance threshold of FDR < 0.05 and | log2 FoldChange | > 1 differential expression, to identify DEGs.
GO and KEGG analyses
-
To investigate the DEGs biological function, we conducted Gene Ontology (GO:
www.geneontology.org ) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG:www.kegg.jp ) pathway analyses. These annotations were used to explore the biological functions and pathways associated with the DEGs. A hypergeometric test was employed to ascertain the statistical significance of the enrichment of GO terms and KEGG pathways. A p-value < 0.05 was considered to indicate significant enrichment.Quantitative real-time PCR
-
To confirm the accuracy and repeatability of the RNA-Seq results, transcription levels of six selected genes in follicles were assessed by using quantitative real-time reverse transcriptase PCR (qRT-PCR). Total RNA was reverse transcribed into cDNA using a PrimeScriptTM RT Reagent Kit (TaKaRa, Dalian, China). Quantitative Real-Time PCR (qRT-PCR) was performed in triplicate using the SYBR Premix Ex Taq II kit (TaKaRa, Dalian, China) on a LightCycler 96 instrument (Roche, Indianapolis, IN, USA). Primers were designed to span exon-exon junctions, ensuring specificity for a single amplicon, as detailed in Table 1. The mRNA expression levels were normalized using the 2−ΔΔCᴛ method with β-actin as the internal control. All experiments were carried out in triplicate.
Table 1. Primer list.
Target Primer sequence (5'-3') Product length (bp) BMP15 F: ATGCTGGAGCTGTACCAACG 135 R: CACGTACCAGCGACCTGC BMPR1B F: CTAGGATTAGAGGGCTCGGAC 167 R: GGCCTTATGGTGAATGTTTCCTT FSHR F: CTGAGTTACACCGTGAGGTCT 242 R: TGGTGAGGACAAATCTCAGTTC CYP11A1 F: GGCGTGCTCCTCAAGACAG 125 R: ACAAAGTCCTGGCTCACCTG STAR F: AGAACTGAGCTCCACTGCAC 214 R: GCCTGGAGCTGAGCAGG SMOC1 F: TCCAAACCACTCTGTGCCTC 124 R: TTTGGCCAGCATCTTTGCAC ZP4 F: GTGCTGACTGCTTGGGATACT 122 R: CAGCTGGTGTAGGAAACGGA ZP2 F: ATTGGGTGCTCAGACAGCTC 150 R: CAAGTCAGACATGCGCGTT MATN3 F: AAGACTTGCTCAAGAGCCACA 194 R: GTGAGTAGAACGCTGGCTCA GDF-9 F: AGGGACCCGATTACAGGAGAC 252 R: CTCAACCCACTTGCGCTTTC β-actin F: CACGGTATTGTCACCAACTG 200 R: ACAGCCTGGATGGCTACATA GCs isolation and culture
-
GCs were isolated from 6-8 mm follicles of Taihang chickens with an average egg-production rate at 43 weeks of age, following a previously published method[15]. The cells were cultured in Dulbecco's modified Eagle's medium with high glucose (H-DMEM) (HyClone, USA), supplemented with 2.5% fetal bovine serum (FBS) (BI, Israel), and 1% penicillin-streptomycin (Solarbio, China).
The experimental design included six groups, each with three replicates, and each treatment was conducted in triplicate cultures. GCs were seeded in 6-well dishes and incubated for 12 h. Subsequently, the medium was refreshed with H-DMEM containing 0.1% bovine serum albumin (BSA) and 1% penicillin-streptomycin solution. Different treatments were then applied to the cells: 50 ng/ml Recombinant BMP15 Protein (R&D Systems, Minneapolis, MN), 50 ng/ml recombinant BMP15 protein combined with 100 ng/ml recombinant human Noggin (NOGGIN) (R&D Systems, Minneapolis, MN), or 50 ng/ml recombinant BMP15 protein combined with 25 ng/ml FSH (R&D Systems, Minneapolis, MN). After 24 h, the medium was replaced with fresh H-DMEM containing 0.1% BSA and 1% penicillin-streptomycin solution, and 50 ng/ml recombinant BMP15 protein was reapplied to the group treated with NOGGIN.
GCs were harvested for RNA extraction and subsequent qRT-PCR analysis. P4 levels in the culture medium were quantified using chicken progesterone (P4) enzyme-linked immunosorbent assay (ELISA) kit (Nanjing Jiancheng, Nanjing, China). The cAMP activity was measured using cAMP-GloTM Assay (Promega, USA).
Statistical analysis
-
The data are presented as the mean ± SEM. Data were analyzed using SPSS 24.0 software (IBM, Chicago, IL, USA). A t-test was used to determine significant differences between two groups (* p < 0.05; ** p < 0.01), and one-way ANOVA followed by a post hoc test-Least Significant Difference (LSD), was used to determine significant differences among three or more groups (different letters indicate significant differences, p < 0.05). Graphics were drawn using GraphPad Prism 8.0 software (San Diego, CA, USA).
-
An analysis of total egg-laying records was performed on 704 Taihang hens, aged 20 to 43 weeks. During the period from 20 to 38 weeks, the median egg production was 72 eggs, with the first and third quartiles being 55 and 81 eggs, respectively (Fig. 1a). From 39 to 43 weeks, the median was 36 eggs, with the first and third quartiles being 29 and 41 eggs, respectively (Fig. 1b). Individuals with a total of egg production from 20-38 weeks above 72 eggs, and those with egg production from 39−43 weeks below 29 eggs and above 41 eggs were classified as the low egg-yielding group (L) and high egg-yielding group (H), respectively. Finally, there were 13 hens in the L group and 74 in the H group. A significance analysis was conducted to validate the feasibility of egg production from 39−43 weeks between the H and L groups. The egg production showed the most significant differences (p < 0.01) (Table 2).
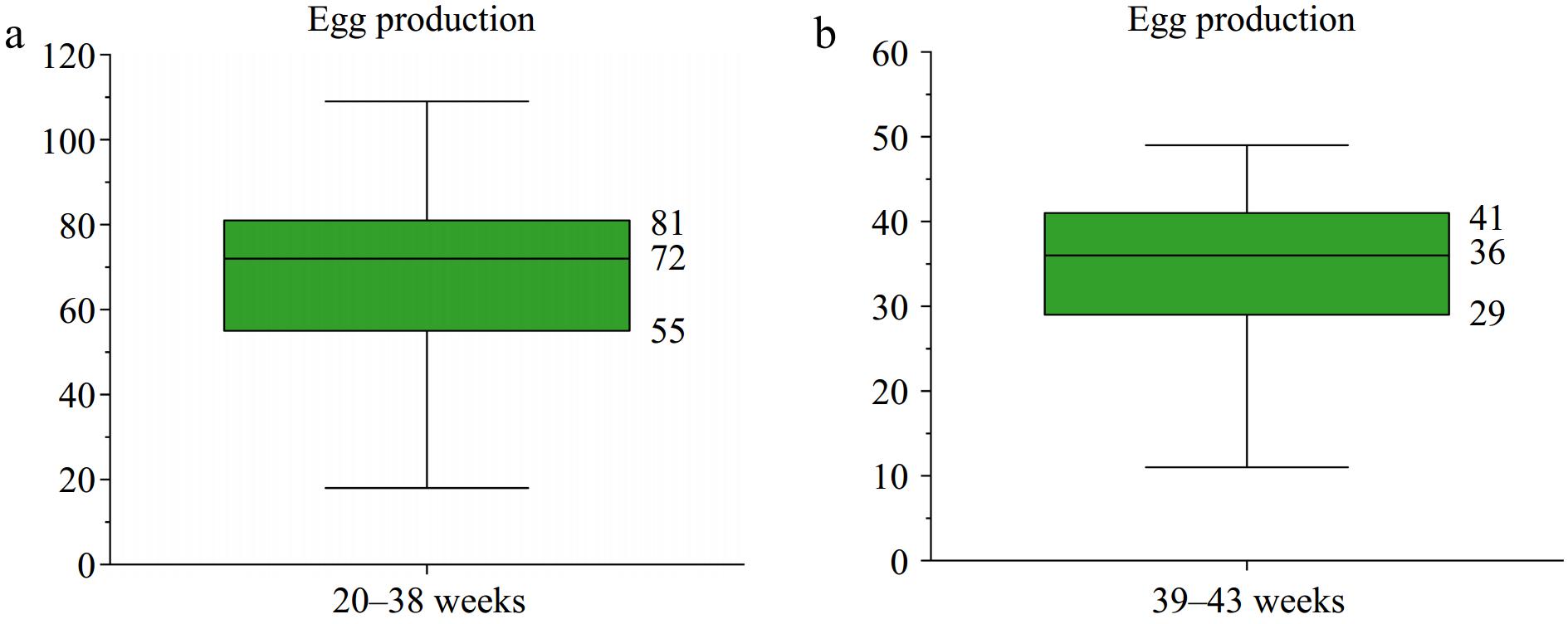
Figure 1.
Egg production of 704 Taihang chickens. (a) Egg production from 20 to 38 weeks. (b) Egg production from 39 to 43 weeks.
Table 2. Average egg production.
High egg-yielding (H) Low egg-yielding (L) 20−38 weeks 82.92 ± 8.76 83.69 ± 10.42 39−43 weeks 43.03 ± 2.00** 12.00 ± 3.71 The data are expressed as the mean ± SEM (* p < 0.05; ** p < 0.01) Sequencing results and quality control
-
We established libraries for both high and low egg-yielding groups using follicles of 6-8 mm from Taihang chickens at 43 weeks of age. A total of 62,748,991 and 92,837,832 raw reads were obtained from the H group and L group, respectively. After removing low-quality reads, we obtained 60,990,303 (H) and 905,616,94 (L) clean reads, which were subsequently used for analysis (Supplementary Table S1).
Differential expression analysis
-
A total of 99 DEGs were identified, characterized by significant changes in expression levels with |log2FoldChange| > 1 and FDR < 0.05. This included five upregulated and 94 downregulated genes (Supplementary Table S2). Among these genes, eight DEGs (Wnt6, BMP15, ZP4, MNR2, ZP2, WNT4, FOXL2, and GDF-9) were implicated in follicle development and selection (Table 3). Notably, the expression level of BMP15 mRNA was significantly downregulated with a log2 (fold change) of -8.10, suggesting a potentially key role in the selection process of chicken follicles. However, the precise function of BMP15 in this context remains to be fully elucidated. To confirm the RNA-seq findings, six genes were selected and quantified by qRT-PCR, which showed similar expression patterns as observed in the sequencing data (Fig. 2).
Table 3. The list of DEGs in follicle development and selection.
Gene name Description log2FC FDR Type Wnt6 Wnt family member 6 −6.0031 8.56E-03 Down MNR2 Homeodomain protein −11.8637 1.97E-02 Down BMP15 Bone morphogenetic protein 15 −8.1033 5.56E-03 Down ZP2 Zona pellucida glycoprotein 2 −8.7247 7.86E-03 Down ZP4 Zona pellucida glycoprotein 4 −8.745 7.86E-03 Down WNT4 Wnt family member 4 −5.5719 3.46E-03 Down FOXL2 Forkhead box L2 −1.8021 2.01E-02 Down GDF-9 Growth differentiation factor 9 −6.44 4.14E-02 Down 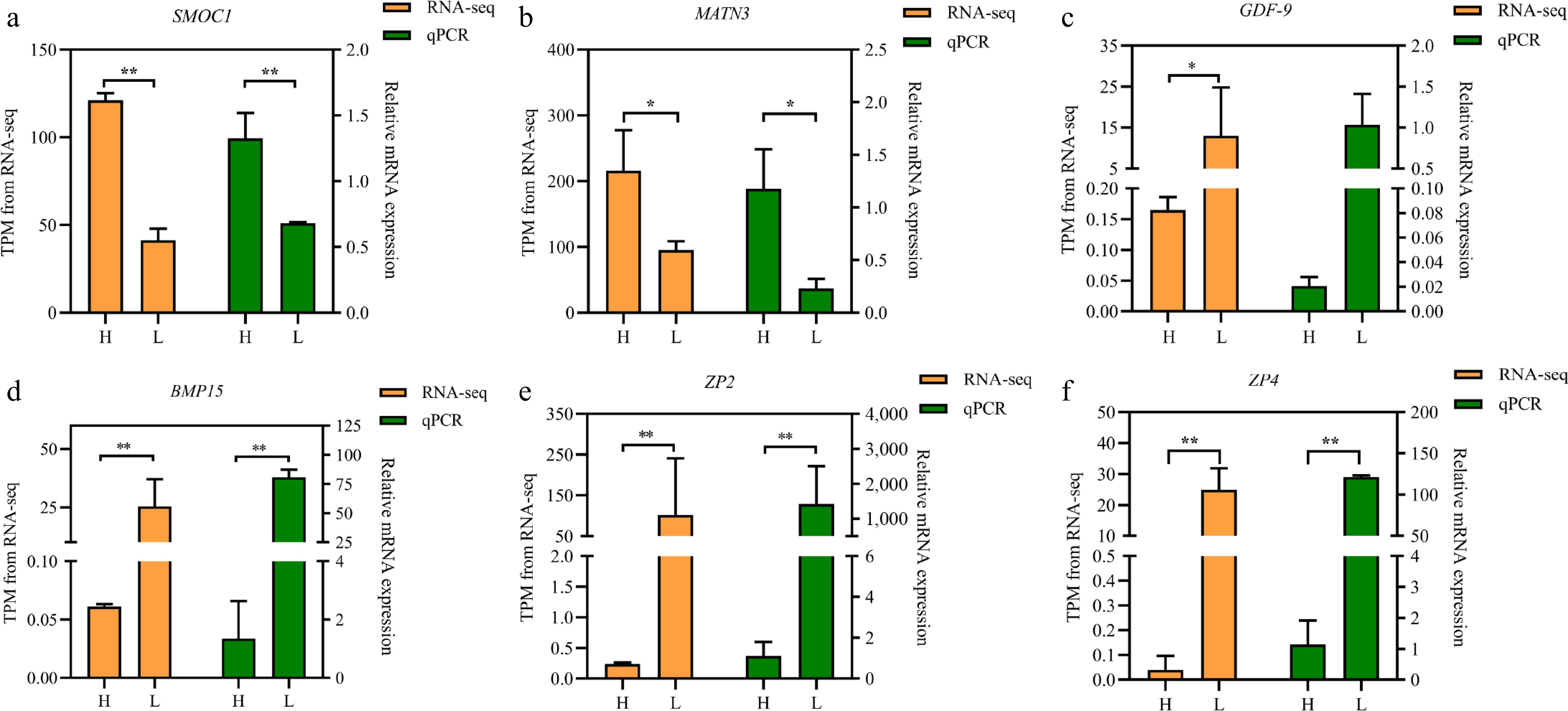
Figure 2.
Comparison of the gene expression of RNA-seq with qPCR of 6 randomly selected genes. (a) SMOC1, (b) MATN3, (c) GDF-9, (d) BMP15, (e) ZP2, and (f) ZP4. Note: The left and right axes represent the gene expression using TPM units by RNA-seq and the expression levels verified by qPCR, respectively. The yellow column indicates TPM value; the green column indicates qPCR. All experiments are performed in triplicate, and the data are expressed as the mean ± SEM (* p < 0.05; ** p < 0.01).
Functional analysis of differentially expressed genes
-
To identify the potential biological functions of DEGs in H and L groups, we conducted enrichment analysis using GO and KEGG pathway databases. DEGs were enriched in terms of related to reproductive process, cell proliferation, binding, transcription regulator activity, molecular transducer activity, transporter activity, and catalytic activity (Fig. 3, Supplementary Table S3).
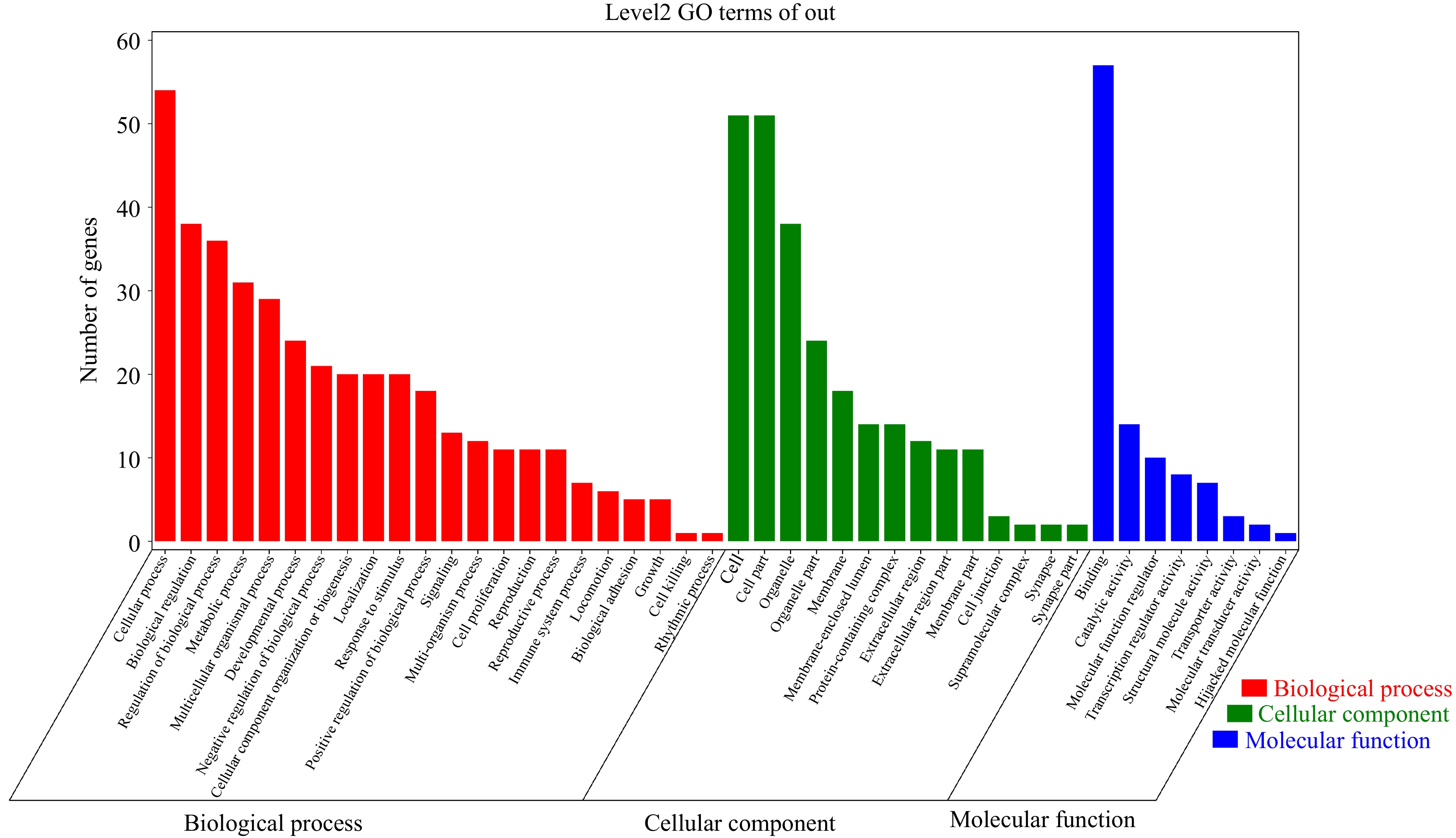
Figure 3.
GO terms enrichment analysis of DEGs.
The KEGG pathway analysis revealed several enriched terms, including Wnt signaling pathway (gga04310), mTOR signaling pathway (gga04150), cell cycle (gga04110), TGF-beta signaling pathway (gga04350), oocyte meiosis (gga04114), FoxO signaling pathway (gga04068), cytokine-cytokine receptor interaction (gga04060), regulation of actin cytoskeleton (gga04810), and the calcium signaling pathway (gga04020) (Fig. 4, Supplementary Table S4).
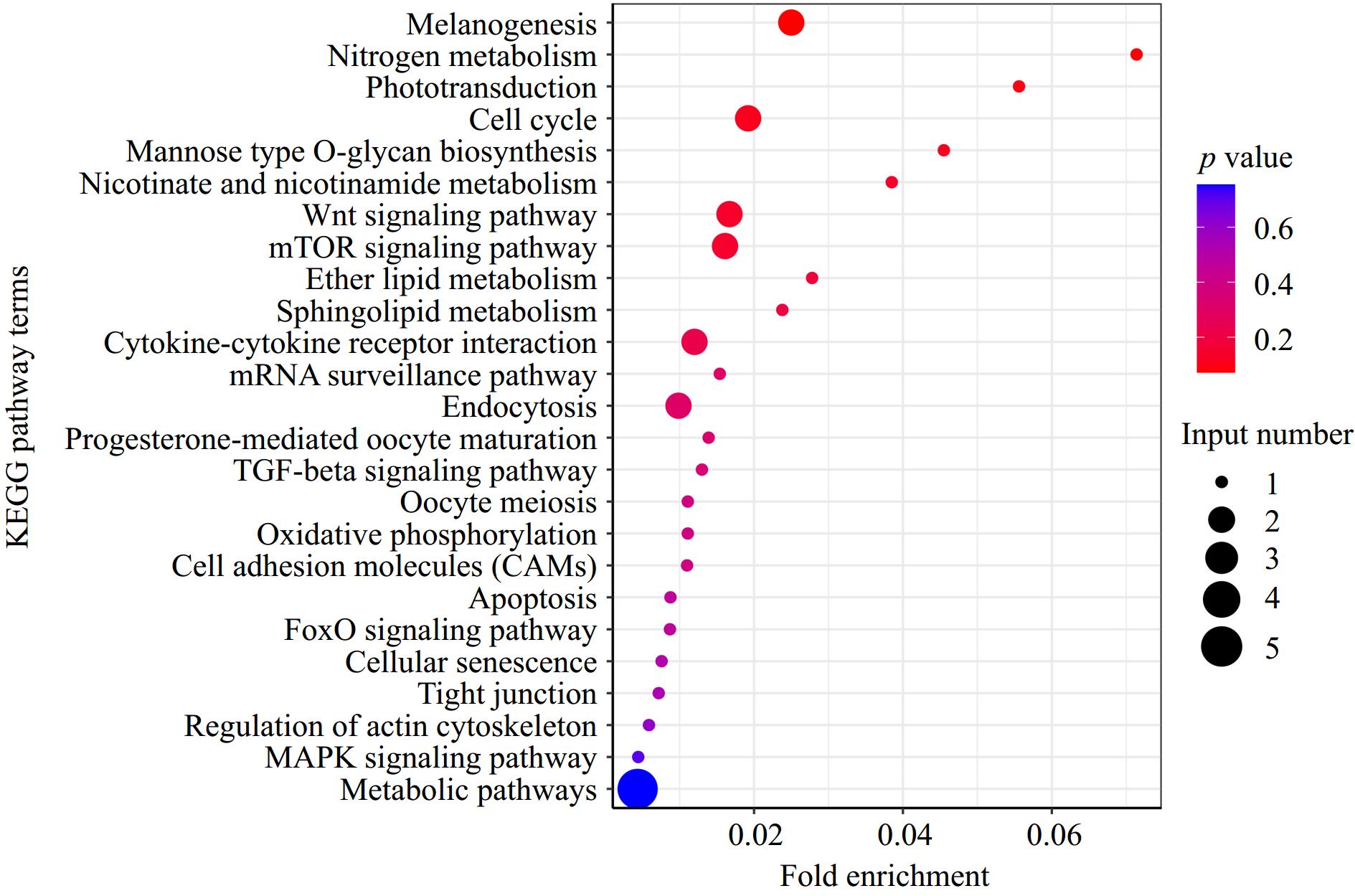
Figure 4.
KEGG pathway enrichment analysis of DEGs.
Function of BMP15 in chicken follicle selection
-
To explore the function of BMP15 in follicle selection, we added recombinant BMP15 protein to GCs. The expression levels of BMPR1B, a specific receptor for BMP15, increased by 22-fold (p < 0.001), confirming the successful addition of the protein (Fig. 5a). Additionally, the upregulation of SMAD1, a component of the SMAD1/5/8 signaling pathway, indicated activation of the pathway (Fig. 5b). The expression levels of genes that are involved in follicle development, such as FSHR, CYP11A1 and STAR, also increased. Furthermore, the results indicated an increase in cAMP activity and progesterone (P4) concentration, both of which are known to promote GC differentiation during follicle selection. There was a significant increase in cAMP activity and FSHR expression (Fig. 5c, d). These changes in gene expression and indices indicated that BMP15 was involved in GC differentiation and affected follicle selection. However, BMP15's influence on steroid hormone synthesis appeared to be inhibitory, as evidenced by the significant reduction in STAR and CYP11A1 expression (Fig. 5e, f). P4 concentration showed a decreasing trend, the change was not statistically significant (Fig. 5g).
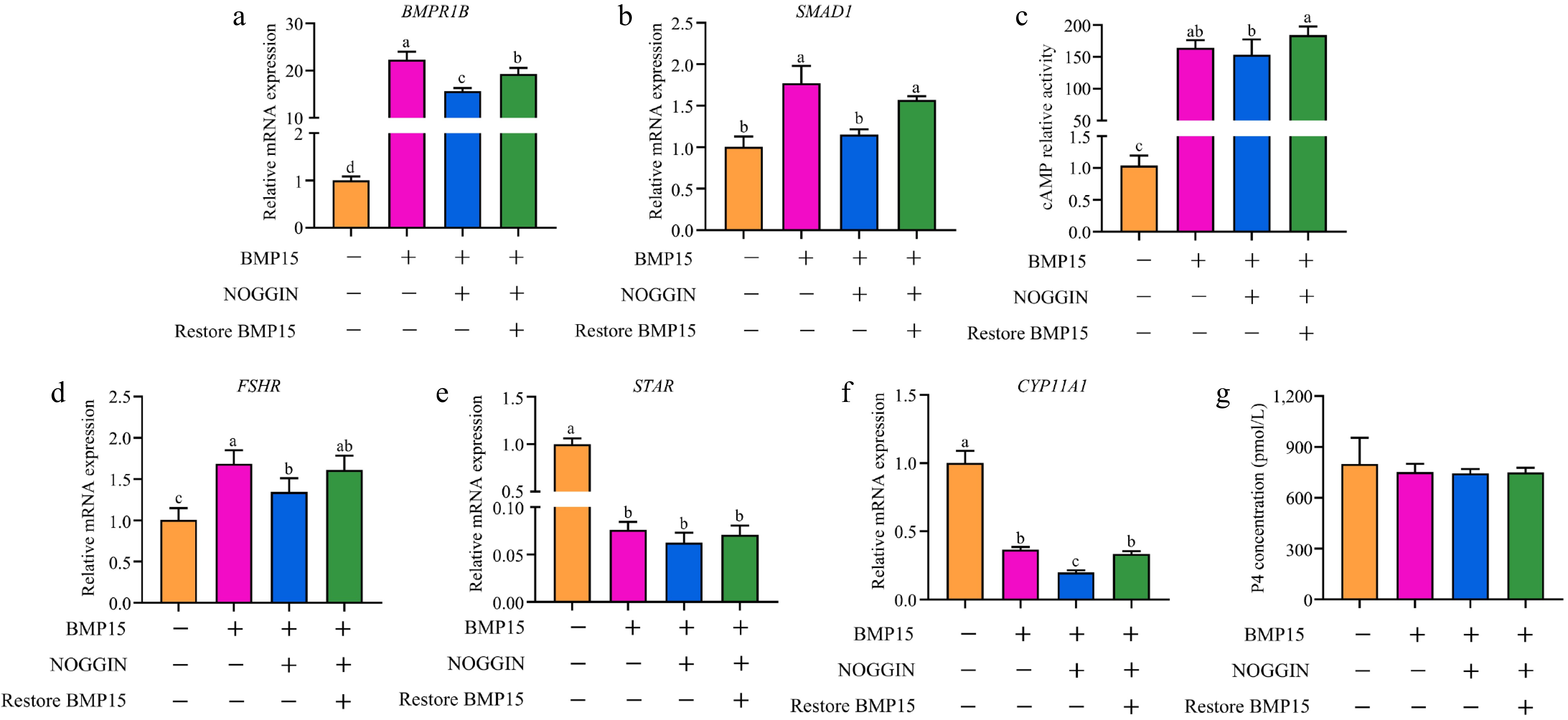
Figure 5.
Effects of recombinant BMP15 on gene expression in GCs. (a) BMPR1B, (b) SMAD1, (d) FSHR, (e) STAR, and (f) CYP11A1. (c) cAMP relative activity, and (g) P4 concentration. Note: All experiments were performed in triplicate, and the data are expressed as the mean SEM (different letters indicate significant difference, p < 0.05).
Subsequently, NOGGIN was added to the culture after 24 h of BMP15 treatment. The addition of NOGGIN resulted in a significant decrease in the expression of BMPR1B (Fig. 5a), indicating an effective disruption of the BMP15-BMPR1B binding. The expression of SMAD1 also decreased (Fig. 5b), suggesting that the SMAD1/5/8 signaling pathway was inhibited. Following NOGGIN addition, FSHR expression significantly decreased (Fig. 5d), and there was non-significant trend towards reduced cAMP activity and P4 concentration (Fig. 5c, g). The expression of STAR and CYP11A1 had a similar trend. These results showed that inhibiting BMP15 leads to a reduction in signaling pathway activity and key gene expression.
To further confirm the role of BMP15, we performed a recovery experiment. Following the disruption of BMP15-BMPR1B binding with NOGGIN, we added recombinant BMP15 protein. Notably, the expression level of BMPR1B was significantly increased compared to treatment with NOGGIN alone (Fig. 5a), indicating the effectiveness of the recovery experiment. Moreover, there was a notable increase in the levels of SMAD1, FSHR, STAR, CYP11A1, cAMP, and P4 upon BMP15 restoration (Fig. 5b−g). These results strongly suggest that BMP15 fosters GC differentiation, thereby regulating follicle selection.
Given FSH's pivotal role in follicle development, especially in selection, we explored its impact on BMP15-mediated processes. We treated GCs with both recombinant BMP15 and FSH, using BMP15-only treatment as a control. Interestingly, the expression of BMPR1B was reduced by half (Fig. 6a), indicating that FSH suppresses BMP15 and BMPR1B expression. While SMAD1 showed an upward trend, the change was not statistically significant (Fig. 6b). However, FSH significantly enhanced the expression of follicle development genes, such as FSHR, STAR, and CYP11A1 (Fig. 6d−f). Moreover, the activity of cAMP and the concentration of P4 were significantly improved (Fig. 6c, g). These findings indicated that FSH, despite inhibiting BMP15 expression, can still facilitate GC differentiation.
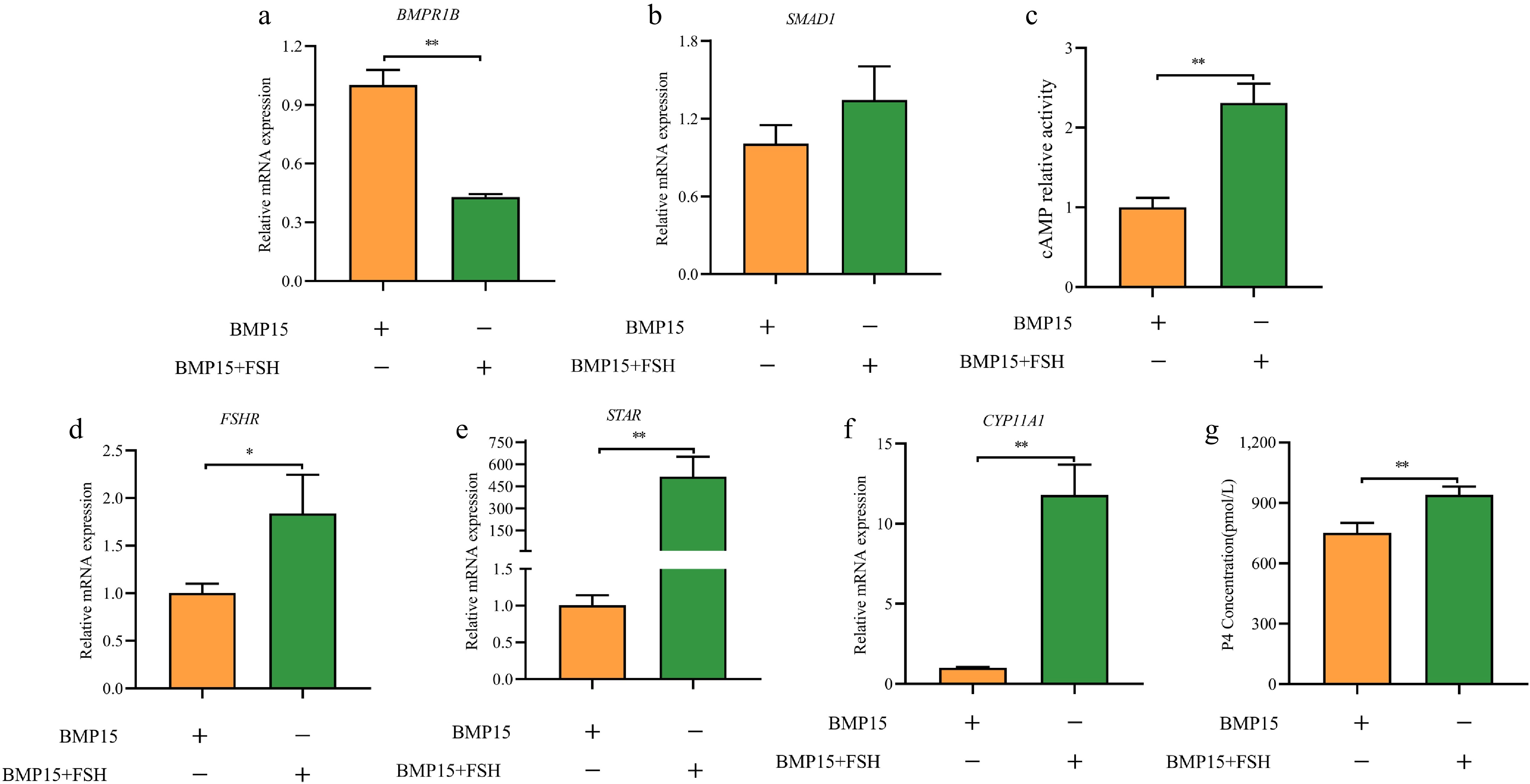
Figure 6.
FSH and BMP15 affected the expression of related genes. (a) BMPR1B, (b) SMAD1, (d) FSHR, (e) STAR, and (f) CYP11A1. (c) cAMP relative activity, and (g) P4 concentration. Note: All experiments were performed in triplicate, and the data are expressed as the mean SEM (* p < 0.05; ** p < 0.01).
-
In recent years, the egg production rate of commercial laying hens during the peak period has been close to the physiological limit. To enhance the utilization rate of these hens, it is essential to extend their egg-laying cycles and boost production rates in the later stages. Follicle selection is an important process that affects the egg production performance and fecundity of hens. It is directly related to the egg production of chickens, 6−8 mm follicles were the key period for determining follicle selection. To ensure continued ovulation of follicles in the late stage of egg production and to identify genes associated with follicle selection during this period, we collected 6-8 mm follicles of Taihang chicken with high- and low-yield during the late laying period for RNA-seq. The majority of DEGs were downregulated genes, suggesting that downregulated genes may play a significant role in follicle selection. Furthermore, Wnt6, MNR2, BMP15, ZP2, ZP4, WNT4, FOXL2, and GDF-9 have been reported to be involved in follicle development and selection[16−20]. Further enrichment and analysis of these DEGs revealed several pathways likely critical to the egg-laying performance of chickens. Among these, the CAMs have been identified as one of the most important pathways associated with high and low egg production[1]. The Wnt signaling pathway plays a crucial role in follicle selection[5]. Additionally, the calcium signaling pathway and cytokine-cytokine receptor interaction are involved in follicle development and follicle atresia[21]. The mTOR signaling pathway and FoxO signaling pathway regulate the cell cycle, proliferation, and apoptosis of chicken follicular GCs[22]. The TGF-β signaling pathway is involved in initiating follicle selection during the follicle selection period[23].
BMP15, a specific factor released by oocytes, belongs to the TGF-β family, and is specifically expressed in oocytes[24]. In mammals, BMP15 plays a crucial role in regulating GC differentiation, as well as participating in the regulation of the FSH signaling pathway[25,26], and follicle development[27]. Like most BMP ligands, BMP15 binds to its specific receptors, BMPR1B and BMPR2, leading to phosphorylation and activation of downstream signaling pathways[28]. Multiple lines of evidence indicate that BMP15 plays a role in regulating follicle development in various species. For instance, in sheep and cattle, BMP15 mutations are associated with altered ovulation rates and follicular development abnormalities[9, 29]. In mice, BMP15 and GDF9 synergistically regulate folliculogenesis and ovarian function[30]. Similarly, research in chickens has indicated that BMP15 influences ovarian reserve and egg production[31].
As a signal factor released by oocytes, BMP15 binds to the specific receptor BMPR1B and activates the SMAD1/5/8 signaling pathway[32]. In a previous study, the addition of recombinant BMP15 protein to mouse GCs activated the SMAD1/5/8 signaling pathway[33]. BMP15 plays an irreplaceable role in the regulation of follicular growth and development and granulosa cell growth and differentiation[34]. We observed that the expression levels of BMPR1B and SMAD1 were significantly increased when recombinant BMP15 protein was added to GCs, leading to the activation of the SMAD1 signaling pathway. The biological function of recombinant BMP15 protein has been previously demonstrated[35], while NOGGIN is known to be an antagonist of bone morphogenetic proteins (BMPs). It works by blocking epitopes on BMPs required for binding to both type I and type II receptors, thereby inhibiting BMP bioactivities[36]. To confirm the role of BMP15 in follicle selection, we disrupted the binding between BMP15 and BMPR1B by adding NOGGIN, which resulted in a significant reduction in BMPR1B expression levels and inhibited the SMAD1 signaling pathway. We then restored the addition of recombinant BMP15 protein, resulting in an increase in BMPR1B expression levels and re-activation of the SMAD1 signaling pathway. These results suggest that BMP15 plays a critical role in follicle selection by activating the SMAD1 signaling pathway via BMPR1B in GCs.
GC differentiation commences as follicles are selected to become hierarchical, with their capacity to synthesize and secrete P4 escalating alongside the gradual increase in FSHR and cAMP levels. The increased levels of FSHR and cAMP in GCs are considered indicative of follicle selection[3]. A marked increase in FSHR expression in GCs of small yellow follicles suggests that follicle selection may be imminent. In our study, we found that BMP15 can significantly increase the levels of FSHR and cAMP, thereby enhancing the sensitivity of GCs to FSH. It is possible that GCs are primed to await the FSH signal to exposure to BMP15. Our findings are corroborated by studies in other species, where BMP15 upregulated FSHR expression in GCs[37]. BMP15 is also known to promote FSHR levels through the SMAD signaling pathway, influencing follicle growth. Specifically, BMP15 binds to its receptor BMPR1B, initiating a signaling cascade that leads to the phosphorylation of SMAD1/5/8. Once phosphorylated, SMAD1 forms a complex with SMAD4 and is translocated into the nucleus, where it regulates the transcription of target genes, including FSHR. Previous studies in mammals have demonstrated that BMP15 modulates FSHR expression through SMAD-dependent transcriptional activation[11]. However, BMP15 may suppress the expression level of FSHR in mice, indicating that the role of BMP15 could vary among species. Additionally, the expression levels of STAR and CYP11A1 were significantly decreased after adding recombinant BMP15 protein, with the P4 concentration remaining unchanged. Previous studies have shown that BMP15 can reduce the levels of STAR, E2, and P4[38]. Two possible mechanisms may explain this effect. First, BMP15 binds BMPR1B and through the SMAD1/5/8 pathway may directly inhibit the transcription of steroidogenic genes. Previous studies have shown that TGF-β superfamily members, including BMPs, can negatively regulate steroidogenesis by interfering with SF-1 (Steroidogenic Factor-1), a critical activator of STAR and CYP11A1[39]. Alternatively, BMP15 may repress STAR and CYP11A1 through an indirect feedback mechanism. BMP15 enhances FSHR expression and increases cAMP levels, which are essential for follicle selection. However, excessive cAMP activation may also trigger a negative regulatory loop, preventing premature or excessive progesterone synthesis in pre-hierarchical follicles. This is consistent with findings in mammals, where BMP15 has been reported to fine-tune FSH responsiveness while keeping steroidogenesis in check[40]. When BMP15 was added alone, it had no influence on the synthesis of steroid hormones[26]. This may be the GCs of pre-hierarchical follicles are not yet differentiated and lack the capability to synthesize and secrete P4[3].
FSH, a glycoprotein secreted by the anterior pituitary gland, affects GCs and regulates follicular development[41]. Studies have shown that FSH promotes GC growth, inhibits their apoptosis[42], and enhances the synthesis and secretion of P4[43]. It has been observed that the responsiveness to FSH was suppressed in pre-hierarchical follicles, however, as pre-hierarchical follicles are selected into hierarchical follicles, some signals are activated, and responsiveness to FSH is initiated by increasing cAMP production in GCs[2]. To investigate the effects of FSH on BMP15 in GCs, we added both factors to the cells and compared the results to those obtained with BMP15 alone. Our analysis revealed a significant decrease in BMPR1B expression, suggesting that FSH may inhibit BMPR1B and BMP15 expression levels, indicating that FSH antagonizes BMP15 signaling while promoting steroidogenesis. This observation suggests that FSH functions as a stage-specific regulator of follicular development, facilitating the transition from BMP15-driven early follicle growth to FSH-dependent late-stage differentiation. Specifically, FSH may repress BMP15 expression through the cAMP-PKA pathway, which is activated during follicle maturation[44]. Increased cAMP levels stimulate steroidogenic enzymes (STAR, CYP11A1) while downregulating oocyte-derived BMP15, allowing granulosa cells to fully respond to FSH stimulation[45]. The interaction between BMP15 and FSH may serve as a developmental checkpoint, ensuring that pre-hierarchical follicles remain BMP15-dependent until selected for further growth, at which point FSH signaling takes precedence.
In our results of RNA-seq results, BMP15 expression was significantly higher in the high egg-yielding group compared to the low egg-yielding group. The high level of FSH in the bodies of high egg-yielding chickens may suppress BMP15 expression. While the SMAD1 level exhibited a slight upward trend, the effect was not statistically significant. Additionally, we also observed marked changes in the expression levels of FSHR, CYP11A1, STAR, P4, and cAMP. Notably, the expression levels of FSHR, STAR, and CYP11A1 were significantly increased, indicating that GCs began responding to FSH signaling and synthesizing P4. Furthermore, there was an upward trend in the concentration of P4 and cAMP activity. Based on these results, we suppose that BMP15 promotes follicle selection, enabling pre-hierarchical follicles to be chosen to enter the hierarchical follicles. Subsequently, GCs begin to respond to FSH, leading to GC differentiation and secretion of P4. As follicles progress to the hierarchical stage, the expression of BMP15 decreases. Consequently, the BMP15 and FSH pathways appear to act in concert during the development of the follicle from the pre-hierarchical to the hierarchical stage (Fig. 7).
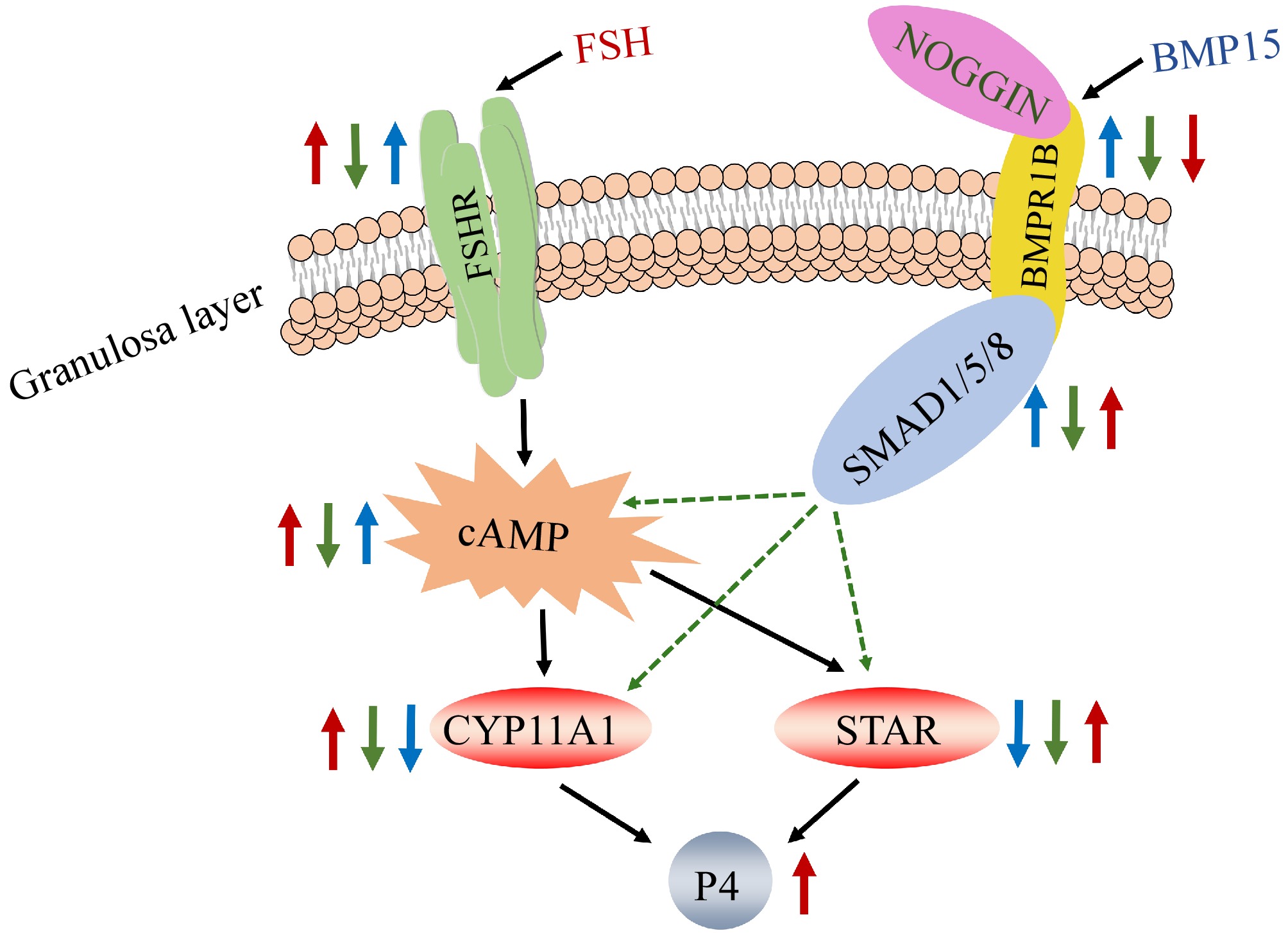
Figure 7.
Regulatory network model of BMP15 and FSH in chicken GCs. Note: The blue arrow shows that other genes expression levels after adding recombinant BMP15 protein; the green arrow shows that other genes expression levels after adding NOGGIN on the base of recombinant BMP15 protein added; the red arrow shows that other genes expression levels after adding FSH on the base of recombinant BMP15 protein added.
These insights have practical applications in poultry breeding and reproductive management. Given BMP15's role in enhancing FSH sensitivity and controlling steroidogenesis, it could serve as a genetic marker for selective breeding programs to improve egg production sustainability. Moreover, interventions aimed at modulating BMP15 expression may help maintain follicle recruitment and extend the productive laying cycle in hens.
-
Transcriptome analysis of 6−8 mm follicles from Taihang chickens with high- and low-yield revealed the role of BMP15 in chicken follicle selection. BMP15 was found to increase the levels of BMPR1B, FSHR, and cAMP, activating the SMAD1 signaling pathway, but decrease the expression of STAR and CYP11A1, and its expression was inhibited by FSH. Moreover, it was observed that from the pre-hierarchical to hierarchical follicles BMP15 interacts with FSH signaling to regulate follicle selection. This study demonstrated that BMP15 promotes follicle selection by enhancing FSHR expression and increasing cAMP levels. These findings on BMP15's role in follicle selection may provide novel strategies to enhance follicular recruitment in aging hens, thereby improving egg yield during the late laying period.
This work was supported by grants from Zhongyuan Young Top Talents for Scientific and Technological Innovation Project, National Natural Science Foundation of China (32341056 and 32102622), the Key Research Project of the Shennong Laboratory (SN01-2022-05).
-
All the animal experiments were approved by the Animal Care and Use Committee of Henan Agricultural University, identification number: 11-0085, approval date: 2019/3/3. The research followed the 'Replacement, Reduction, and Refinement' principles to minimize harm to animals. This article provides details on the housing conditions, care, and pain management for the animals, ensuring that the impact on the animals is minimized during the experiment.
-
The authors confirm contribution to the paper as follows: study conception and design: Zhang C, Kang X, Li W; data collection: Wang S, Tian K, Sun G, Wang K; analysis and interpretation of results: Zhang C, Tian Y, Gai Y; draft manuscript preparation: Zhang C, Shi J, Wang K, Li W. All authors reviewed the results and approved the final version of the manuscript.
-
All data in this study are included in the article content and the supplementary files. Transcriptome data were uploaded to the NCBI database sequence read archive, Accession no. PRJNA1080244 (www.ncbi.nlm.nih.gov/bioproject/PRJNA1080244).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Chenxi Zhang, Junlai Shi
- Supplementary Table S1 Sequencing output and quality evaluation statistics of 5 libraries.
- Supplementary Table S2 The list of DEGs in L and H groups.
- Supplementary Table S3 GO terms enrichment analysis of DEGs.
- Supplementary Table S4 KEGG pathway enrichment analysis of DEGs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang C, Shi J, Wang S, Tian K, Jia X, et al. 2025. Transcriptome for small yellow follicles reveals bone morphogenetic protein 15 involved in chicken follicle selection. Animal Advances 2: e016 doi: 10.48130/animadv-0025-0010 

Transcriptome for small yellow follicles reveals bone morphogenetic protein 15 involved in chicken follicle selection
- Received: 14 February 2025
- Revised: 12 March 2025
- Accepted: 17 March 2025
- Published online: 24 June 2025
Abstract: Sustaining egg production in late-laying chickens can extend their egg-laying cycle. Follicle selection is a pivotal process that determines the egg production rate, acting as the ultimate regulatory checkpoint to ensure only the most viable follicles progress to ovulation. This study aimed to identify and elucidate the role of a potential gene in follicle selection. Follicles measuring 6−8 mm were harvested from high egg-yielding (H) and low egg-yielding (L) Taihang chickens at 43 weeks of age for RNA sequencing to identify DEGs. The role of BMP15 in follicle selection was investigated by adding recombinant BMP15 protein to GCs. As a result, 99 DEGs were identified, and BMP15 was selected as the candidate gene for its role in follicle selection. The addition of recombinant BMP15 activated the SMAD1 signaling pathway, upregulated the expression of bone morphogenetic protein receptor type 1B (BMPR1B) and follicle-stimulating hormone receptor (FSHR), and increased cyclic adenosine monophosphate (cAMP) levels. However, BMP15 significantly repressed the expression of steroidogenic acute regulatory protein (STAR) and cytochrome P450 family 11 subfamily A member 1 (CYP11A1) and slightly decreased the progesterone (P4) levels. Furthermore, the co-administration of follicle-stimulating hormone (FSH) with recombinant BMP15 resulted in a significant decrease in BMPR1B expression and a marked increase in FSHR, STAR, and CYP11A1 expression, which led to a comparable change in P4 levels. These findings demonstrated that BMP15 is involved in follicle selection by enhancing FSHR expression and the cAMP level. BMP15 interacts with FSH to regulate follicle selection from the pre-hierarchical to hierarchical follicles.
-
Key words:
- Follicle selection /
- Small yellow follicle /
- RNA-seq /
- BMP15 /
- FSH