









-
Potatoes, the fourth largest staple crop globally after rice, wheat, and maize, are the tubers of the plant Solanum tuberosum L. Given the rich nutritional and commodity value of potatoes, it is an important food resource around the world[1]. Since 1993, China has been the world's largest potato producer, contributing to about 22% of global production, followed by India, Ukraine, and the United States. In addition to being eaten fresh, potato has been widely used in the production of traditional staple food products such as buns and noodles, thus increasing its position in the national dietary structure[2]. With the increasing demand of potato food staples, the planting area of potatoes in China has increased, and the processing industry has been gradually developing and expanding[2]. Recently, the taste requirements for potato and its food staples have also gradually improved due to the changing demands of consumers. Therefore, potato quality has become an important indicator of its competitiveness in the marketplace.
Flavor is a key quality indicator in evaluating potatoes and their products. The flavor of fruits and vegetables, including potatoes, are mainly composed of volatile aromatic substances (e.g., esters, alcohols, aldehydes, ketones, and sulfur-containing compounds), which depend on precursors such as fatty acids, amino acids, and carbohydrates, and are produced through complex biosynthetic pathways[2,3]. Potato flavor quality is determined by a combination of physical and sensory attributes, which in turn are influenced by the structure and chemical composition of the potato. Studies have shown that the flavor precursors of raw potato are mainly composed of sugars, amino acids, ribonucleic acids, and lipids, which form a variety of flavor compounds through the Maillard reaction during cooking[4]. The factors affecting flavor are complex and include pre-harvest varieties, and climate as well as mechanical and chemical treatments during processing[5]. Freshness techniques and suitable packaging materials are often used to control flavor changes, but improper handling may lead to flavor deterioration.
Potatoes are rich in carbohydrates, vitamins, proteins, and several essential amino acids. These nutritional properties indicate that the rich nutritional value of potato is capable of meeting the body's daily requirement for most vitamins and minerals[6,7]. The carbohydrate (mainly starch) content of potato, which is the main source of energy, can reach 18% of its fresh weight. Moreover, potatoes are abundant in various polyphenols such as phenolic acids, flavonoids, and anthocyanins[8]. Previous studies have reported that the health benefits of potato include antioxidant, antidiabetic, hepatoprotective. and antibacterial properties[9].
The flavor and composition of potatoes are affected by a variety of factors, including genotype, environmental conditions during growth, post-harvest storage conditions, cooking, and processing[10]. Variety is a key factor influencing potato quality, but there is relatively limited scientific information on how to systematically screen and evaluate high-quality flavorful potato varieties[11]. Zhang et al., demonstrated that different potato varieties from the same area contained large differences in their volatile compounds and metabolites[12]. Several studies have reported on the effects of environmental growth conditions on potato composition[13,14]. A previous study demonstrated that potatoes from three areas in Greece showed large differences in physicochemical attributes, minerals, and sensory attributes among different areas[15]. However, the detailed influences of geographical factors on the flavor and metabolites of potatoes in China remains unknown. To achieve this, Qingshu 9 potatoes from five provinces (Inner Mongolia, Shanxi, Gansu, Hubei, and Yunnan) in China were collected. The potato variety Qingshu 9 has been reported to be highly adaptable to diverse geographic regions of China, and results in wide planting in the country[16]. Afterward, the basic nutrients, volatile compounds, and metabolites were identified to study the variations in potato compositions. Moreover, correlation analysis was used to identify the relationships between volatile and metabolites with geographical factors. Together, these results will provide a novel insight into the changes in potato compositions in different areas, and provide potential applications of potato products in the food industry in the future.
-
All chemicals used in this study were HPLC grade reagents suitable for volatilomics and metabolomics analysis. Folin-phenol (F6010), amylopectin (A6150), amylose (A6160), sodium carbonate (S7795), and gallic acid (G0200) were purchased from Shanghai Yuanye Biotechnology Co., Ltd. (Shanghai, China). Iodine were procured from Macklin (Shanghai, China). Other chemicals used in this study were obtained from Beijing Tong Guang Fine Chemical Co., Ltd (Beijing, China), and Xilong Scientific Co., Ltd. (Shanghai, China). The potatoes (Solanum tuberosum L. Qingshu 9) were respectively collected from local farms in Inner Mongolia (M9), Shanxi (J9), Gansu (G9), Hubei (E9), and Yunnan (D9) provinces. After washing, the potatoes were treated with liquid nitrogen for 2-3 mins and ground using a blender. The ground samples were then passed through a 50-mesh sieve, and the resulting powder was stored at −80 °C for subsequent analysis.
Color profile and basic nutrients
-
The color profile of Qingshu 9 potatoes was evaluated using a color spectrophotometer (CR-400, Minolta Co. Ltd, Osaka, Japan). Fresh potato samples were placed on a flat dish, and reflectance readings were recorded using a 90° standard observer and illuminants. The measurements recorded values for lightness (L*), the green-red axis (a*), and the blue-yellow axis (b*) on the Hunter scale.
The identifications of basic nutrients of potato samples followed the protocols reported by a previous study[12]. Polyphenol content was identified by Folin-Ciocalteu assay[17]. First, 1 g freeze-dried potato powder sample was dissolved in 20 mL of acetone with 80% concentration, and gently shaken for 10 min. Second, 0.5 mL sample solution was combined with 0.5 mL of Folin phenol reagent, 3.0 mL of 10 g/100 mL sodium carbonate solution was added to the sample solution. The total volume of the mixture was adjusted to 5.0 mL, and reacted in the absence of light for 30 min. Finally, the absorbance was measured at 760 nm, and the total polyphenol content calculated in potatoes based on the standard curve.
To measure starch content, 100 mg potato powder was mixed with 1 mL of ethanol and 9 mL of 1 M potassium hydroxide solution[16]. The sample solution was heated for 10 min at 95oC. After cooling, the final volume was adjusted to 100 mL. Next, 2 mL of the prepared mixture was dissolved in 6 mL of potassium hydroxide solution, 2 mL of acetic acid, and 1 mL of 0.02 N iodine solution (mixture of I : KI = 1:2). The final mixture reacted for 30 min at room temperature, and the absorbance of mixtures was measured at 427, 535, 630, and 757 nm, respectively. Standard curves were prepared using amylose and amylopectin solutions with concentrations ranging from 0.0 to 36.0 μg/mL[16], and the amylose and amylopectin concentrations calculated in potato samples.
Sample preparation and GC-MS analysis
-
The identification of volatiles in potatoes was performed using solid-phase microextraction coupled with gas chromatography and mass spectrometry (SPME-GC/MS) (7890-5985C, Agilent, USA) following the method of Blanda et al.[18]. Each potato sample was analyzed in triplicate. Three grams of fresh potato was combined with 0.3 g NaCl in a 20 mL headspace vial. A semiquantitative internal standard method was employed using 0.5 μL 0.816 μg/μL 2-Methyl-3-heptanone. The headspace vial was with a silicon cap and heated at 50 °C for 30 min. The divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS, 50/30 μm, coating 2 cm) manual holder (Supelco Ltd., Bellefonte, PA, USA) was used to extract at 50 °C for 40 min. The chromatographic separation was achieved using a DB-WAX column (30 m × 250 μm × 0.25 μm). The temperature program was set as follows: an initial hold at 40 °C for 3 min, increasing from 40 to 200 °C at a rate of 5 °C/min, followed by another ramp from 200 to 230 °C at the same rate, with a final hold at 230 °C for 5 min. Qualitative identification of volatile compounds was carried out using the NIST library and retention indices (RI), and the volatiles with RI values above 70% were considered. The quantification analysis for identified volatile compounds was calculated as follows:
$ C=\frac{S}{{S}_{i}}\times \frac{{C}_{i}}{m} $ where, C is the concentration of identified compounds (μg/g); Ci is the concentration for the internal standard; S is the peak area for identified compounds; Si is the peak area for the internal standard; m is the sample weight (g).
Sample preparation for metabolomics
-
The preparation of potato samples for metabolomics analysis was conducted following the method described in a previous study[19]. Potato samples were first freeze-dried, and the dried samples were ground using a high-speed blender (Midea MJ-PB80P2-C1, China). Next, 1 g of the dried potato powder was added to 10 mL of cooling 70% (v/v) methanol solution and subjected to ultrasonic treatment for 30 min. The mixture was then stored at −20 °C for 10 min and centrifuged at 10,000 g for 20 min. All the potato samples were mixed to use as a quality control (QC) sample, which is used to evaluate the accuracy of untargeted metabolomics data[20]. The supernatant of mixture was adjusted to a final volume of 10 mL and passed through a 0.22 µm membrane filter for subsequent analysis.
UPLC-QTOF-MS-based metabolomics analysis
-
Sample separation was conducted using a UPLC system (ExuinLC AB SCIEX, Massachusetts, USA) using an ACQUITY UHPLC HSS T3 column (1.8 µm × 2.1 µm × 100 mm, Waters, Massachusetts, USA) with a flow rate at 0.30 mL/min at 40 oC. Mobile phase A consisted of 0.1% formic acid in water and phase B was 0.1% formic acid in acetonitrile, using the following elution conditions: 95%−70% A at 0.00–11.50 min, 70%–0% A at 11.50–11.51 min, 0% A at 11.51–15.00 min, 0–95% A at 15.00–15.01 min, and 95% A at 15.01–18.00 min. The injected volume for sampling and QC was set at 10 μL. Mass spectrometry analysis was performed using QTOF-MS (6600 SCIEX, Massachusetts, USA) in both positive and negative ionization modes. The full mass spectrometric scan range was 75–1,125 m/z, with a resolution of 70,000. The main precursor ion scan covered a range of 100–1,000 Da, while the product ion scan spanned 50–1,000 Da. The QTOF-MS was set as follows: source temperature: 500 °C; ion source gas 1 and gas 2: 50 °C; curtain gas: 25 °C; ion spray voltage floating: 5.5 kV; declustering potential: 60 V; collision energy: 10 eV.
Data analysis
-
Metabolomics raw data were analyzed using MS-DIAL software (v 5.1.23) for metabolite identification by matching the public and local databases. The mass tolerance for peak extraction was set at 15 ppm, and the retention time tolerance was 0.1 min. Metabolites were included in the analysis only if their detection rate (DR) exceeded 60% across all samples. For further analysis, metabolites were selected based on having an MS/MS spectrum and similarity score > 0.95.
Statistical analysis
-
Statistical analysis was performed using SPSS v 26.0. A paired t-test was conducted to compare metabolites within paired samples, with a significance level of p < 0.05. A one-way ANOVA was used to compare metabolites among different Qingshu 9 potatoes, with significance set at p < 0.05. For multiple comparisons between Qingshu 9 from different areas, Duncan's test was used as a post hoc analysis. Partial least squares discriminant analysis (PLS-DA) and Pearson rank correlation were carried out using R (R Core Team, 2016). KEGG pathway analysis of differentially expressed metabolites in potatoes was performed using MetaboAnalyst 6.0 (
www.metaboanalyst.ca ). -
The geographical location and appearance of Qingshu 9 potatoes from different areas of China is shown in Fig. 1a. It is clear that the appearances of these potatoes were very similar. This is also confirmed by the color profile of the potato surface from different areas of China, which indicated that there were no significant differences in the L* value (Fig. 1b). a* and b* were significantly different among potatoes in different areas. For example, the potatoes from Gansu and Inner Mongolia were redder with higher values of a*. The potatoes from Shanxi were more yellow, and the potatoes in Gansu were more blue. The analysis of basic nutrients including polyphenol, amylose, and amylopectin contents are shown in Fig. 1c, d. The results showed that the total polyphenol contents were highest in potatoes in Inner Mongolia (30.13 ± 1.08 mg/100 g) followed by these in Hubei (28.22 ± 0.53 mg). As for the starch content, the results indicated that the potatoes in Inner Mongolia and Shanxi were abundant in both amylose and amylopectin content, while the potatoes in Yunnan were lowest in starch content. This reveals that the geographical factors significantly altered the basic nutrients of potatoes even with the same variant, which was in line with a previous study[21]. The different content of potato starch in Qingshu 9 from different areas was related to the changes in their applications in the food industry[22]. Qingshu 9 showed great performance in fresh eating and food processing, while it may have different situations for potatoes planted in different areas. Overall, the appearances and basic nutrients of Qingshu 9 from different areas of China were significantly different.
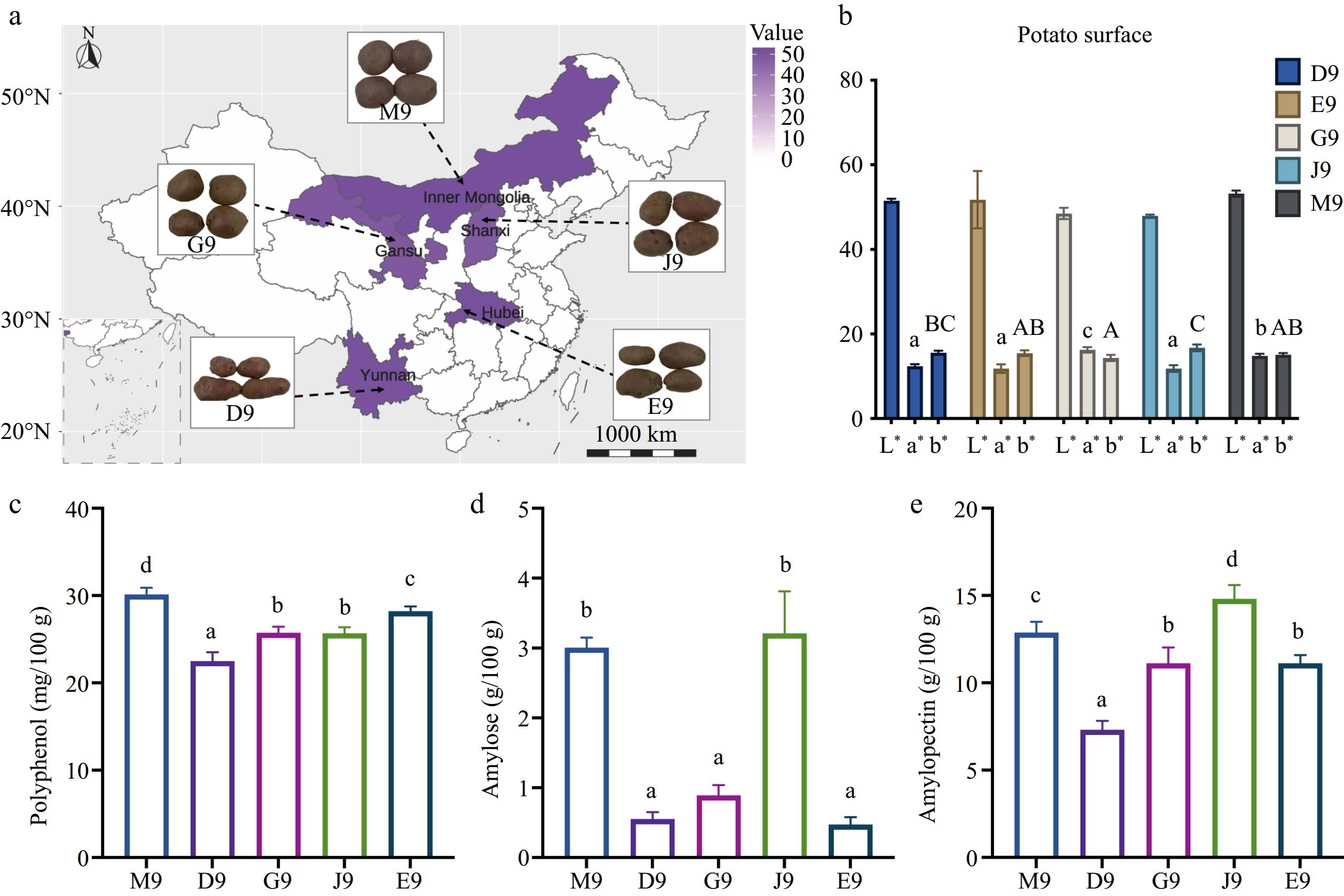
Figure 1.
(a) Geographical locations for cultivation of potato plants in China, the color means the different appearances of the potato surface, the darker the purple means the darker the surface color. (b) Color profile, lowercase letters indicate significant differences in the green-red axis (a*), capital letters indicate significant differences in the blue-yellow axis (b*) by Duncan test (p < 0.05). (c) Total polyphenol content (mg/100 g) in potato. (d) Amylose content (g/100 g). (e) Amylopectin content (g/100 g), lowercase letters indicate significant differences (p < 0.05) of polyphenol, amylose, and amylopectin content among different potatoes.
Volatile compounds in potatoes from different areas
-
In addition to the basic nutrients, the volatile compounds in potatoes from different areas of China were analyzed, as shown in Fig. 2. In total, 35 volatile compounds were identified. The dominant volatile compounds were 2-(E)-Octenal, 2-pentyl-Furan, (E,Z)-2,4-Decadienal, and n-Hexane, which is in line with previous studies[23]. Of these, 2-(E)-Octenal has been reported as the classic potato flavor with the description of baked potato, bakery, boiled, and chipped[24]. A previous study reported that the formation of 2-pentylfuran mainly resulted from extensive degradation reactions in the presence of methionine[25]. Moreover, Fig. 2a shows that the contents of volatile compounds were different in potatoes from different areas of China. Of these, D9 and J9 contained higher amounts of volatile compounds with higher concentrations such as 2-(E)-Octenal, 2-(E)-Nonenal in J9, and methyl-Cyclopentane, 2-methyl-Benzaldehyde in D9.
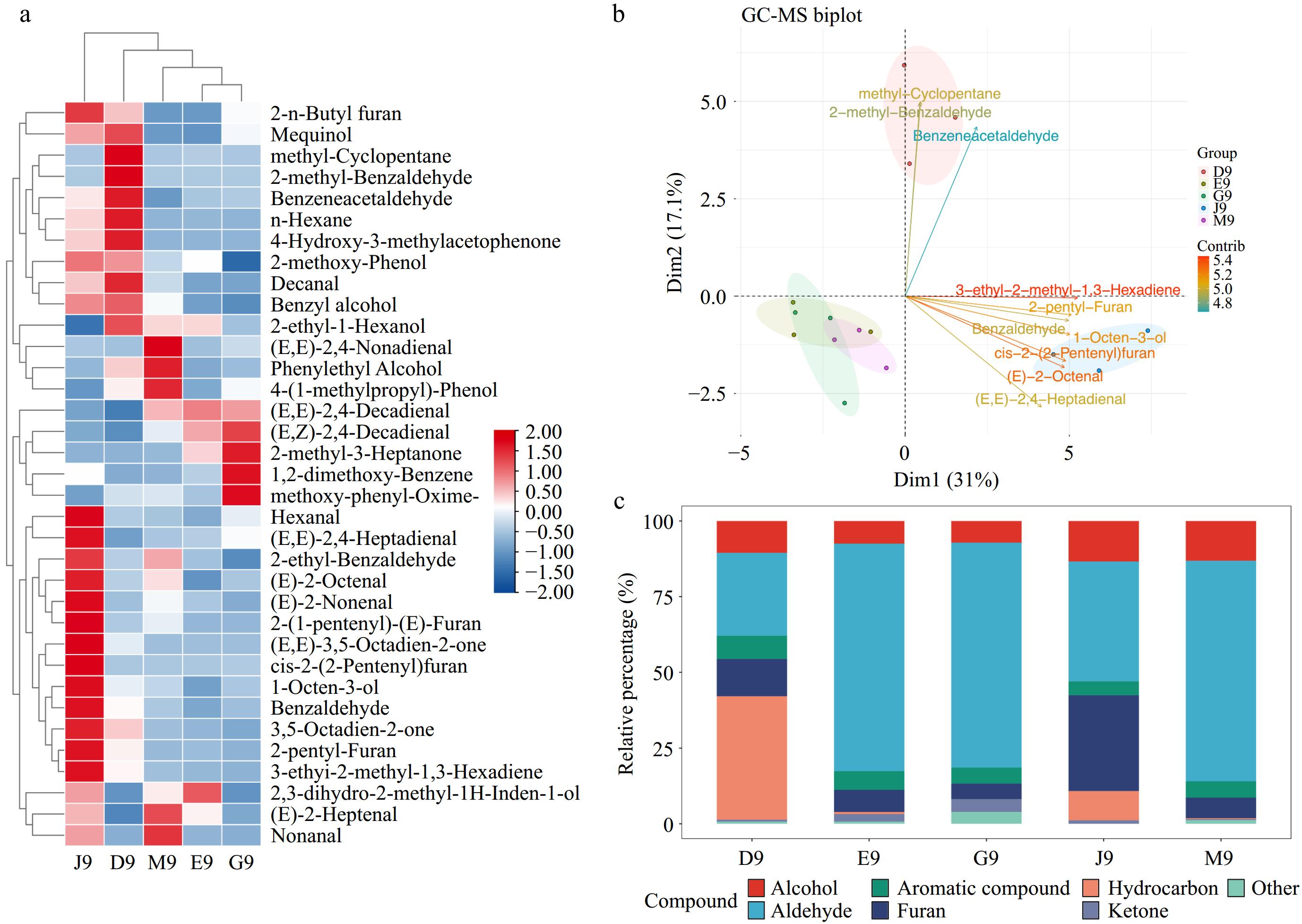
Figure 2.
The analysis of volatile compounds in the potatoes from different areas of China. (a) Heatmap of all volatile compounds. (b) Biplot analysis. (c) Stacked bar chart of volatile compound categories.
To understand the volatile compound differences, PCA analysis was performed, as shown in Fig. 2b. The results showed that PC1 (31.0%) well distinguished J9 and PC2 (17.1%) well distinguished D9 from Qingshu 9 from other areas. However, potatoes from Hubei (E9), Gansu (G9), and Inner Mongolia (M9) were not well distinguished. The loading plot shows that the dominant discriminators for J9 were 3-ethyl-2-methyl-1, 3-Hexadiene, 2-pentyl-Furan, cis-2-(2-Pentenyl) furan, and 2-(E)-Octenal. Of these, 2-(E)-Octenal has been reported as baked, boiled, and fatty flavors in potatoes[26]. Methyl-cyclopentane, 2-methyl-benzaldehyde, and benzeneacetaldehyde were the main discriminators for D9 to be different from other potatoes. Previous studies reported that methyl-cyclopentane and benzeneacetaldehyde have been identified as volatile compounds of baked potatoes[27,28]. This reveals that J9 and D9 may have stronger potato characteristic flavors. Figure 2c shows the stacked bar chart of the relative percentage of volatile compound categories. The results showed that E9, G9, and M9 contained higher amounts of aldehyde compounds, which was similar to a previous study on aldehyde as the dominant volatile compound in potatoes[23]. In contrast, D9 contained a higher amount of hydrocarbon and J9 contained higher proportions of furan. A previous study reported that the various hydrocarbon volatile compounds in potato were produced by fungi in soil[29]. This is also reasonable because of the higher fungi diversity in Yunnan (D9) province[30]. Furthermore, the formation of furan volatile compounds was related to the lipid degradation in potatoes[31]. Overall, the large differences of volatile compounds for Qingshu 9 from different areas of China were observed, which may result in the different performance in their applications in the food industry.
Metabolomics analysis of potatoes from different areas of China
-
To investigate the metabolite compositions among Qingshu 9 from different areas of China, non-targeted metabolomics analysis was performed (Fig. 3). The metabolomics raw data was filtered by MS/MS spectrum and MS/MS similarity. After correction, there were 361 metabolites in positive mode and 154 metabolites in negative mode. We combined the metabolites in both positive and negative mode and there were 369 metabolites identified. PCA analysis was used to provide an overview of all metabolites among potatoes from different areas (Fig. 3a). The results showed that the metabolites in QC samples were located in the center of the PCA axis, which revealed the accuracy of the data. Furthermore, PC1 and PC2 (31.9% and 29.4%, respectively) well distinguished the metabolites in E9, M9, and J9, while G9 and D9 were not distinguished. The overview of all metabolites were shown as a heatmap in Fig. 3b. The results were consistent with the PCA analysis, where M9 and E9 were clustered together. The main metabolite categories in potatoes included phenolics, amino acids, organic acids, esters, and saccharides (Fig. 3c). KEGG pathway analysis was performed to identify the biological functions of these metabolites in potatoes. Of these, most of the metabolites were involved in purine metabolism, alanine, aspartate, and glutamate metabolism, valine, leucine, and isoleucine biosynthesis, glucosinolate biosynthesis, and arginine biosynthesis. Purine metabolism plays a critical role in the energy-related aspects of metabolism as well as in a number of essential coenzymes and signaling molecules in potato[32]. Overall, the overview of metabolite composition in potatoes from different areas of China was studied, and larger differences in metabolites may explain the changes in basic nutrients and volatile compounds.
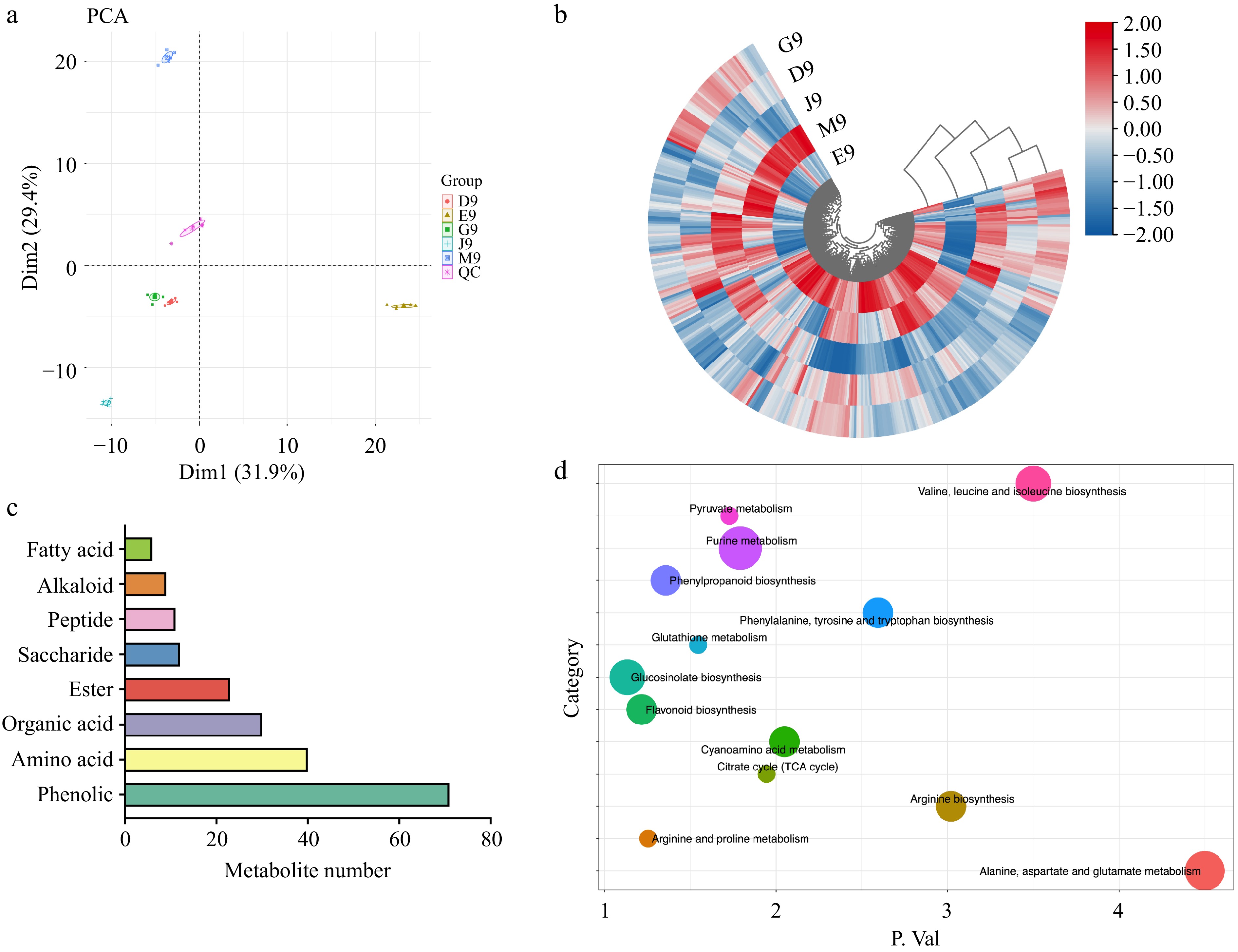
Figure 3.
The analysis of metabolites in the potatoes from different areas of China. (a) PCA analysis. (b) Heatmap of all identified metabolites. (c) Numbers of dominant metabolite categories. (d) KEGG pathways of metabolites.
Different expressed metabolite in potatoes from different areas
-
PLS-DA was used to identify the variations of metabolite composition among Qingshu 9 from different areas of China (Fig. 4). The results showed R2X = 0.886, R2Y = 0.996, and Q2 = 0.993 for the PLS-DA model, which revealed the excellent fitness and predictability of the model (Fig. 4a). The loading plot was used to identify the characteristic metabolites in Qingshu 9 from different areas of China: linolenic acid and adenine in G9 and D9; 4-benzyl-7-hydroxy-1-hydroxymethyl-tetrahydro-pyrrolizinium in E9; 3-octahydrodibenzo hexaoxacyclooctadecine in J9; 2'-Deoxyinosine 5'-monophosphate, Trigonelline, ferulic acid in M9 (Fig. 4b). A previous study reported that linolenic acid is a fatty acid and most volatile compounds were formed from linoleic acid degradation[25]. Moreover, ferulic acid has been reported as the predominant bound phenolic acid in potato peel[33], which may reveal the differences in appearance of M9 compared to others.
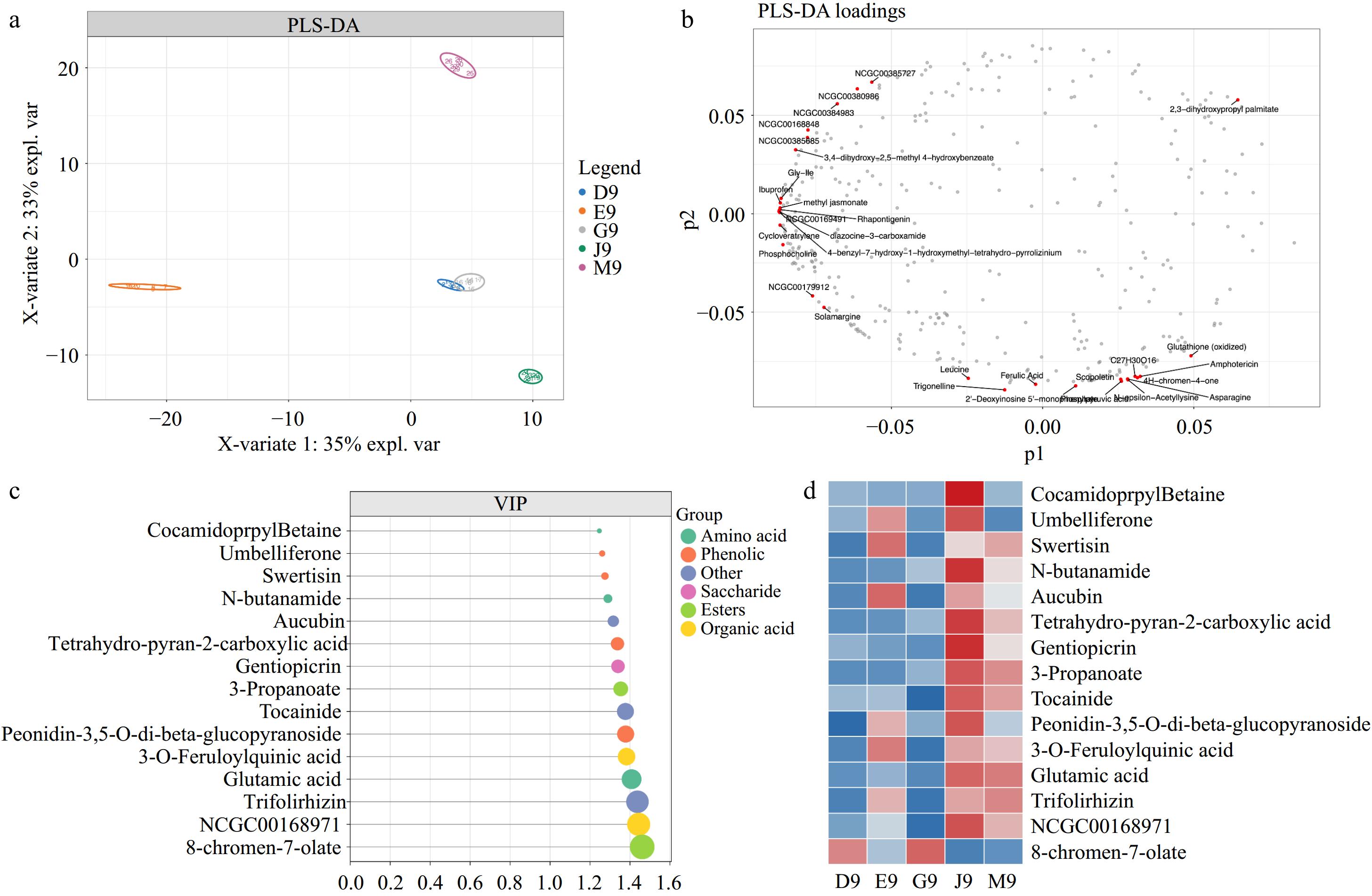
Figure 4.
(a) PLS-DA score plot. (b) PLS-DA loading plot. (c) Metabolites with the top 15 highest VIP values. (d) Heatmap analysis of the intensity of the top 15 VIP metabolites.
The top 15 metabolites with the highest variable importance in projection (VIP) values are shown in Fig. 4c, and the intensity of the top 15 the VIP value metabolites are shown as a heatmap in Fig. 4d. The results showed that the metabolite with the highest VIP value was 8-chromen-7-olate followed by trifolirhizin, glutamic acid, and 3-O-feruloylquinic acid. Trifolirhizin, a pterocarpan flavonoid, was isolated from the roots of Sophora flavescens, which has been reported to have potential anti-inflammatory and anti-cancer activities[34]. Furthermore, these metabolites with higher VIP values were classified as phenolic, organic acids, and others. The heatmap in Fig. 4d showed that most of the metabolites with higher VIP values were abundant in J9 and M9. Moreover, 8-chromen-7-olate was found to be abundant in D9 and G9, and E9 contained higher amounts of 3-O-feruloylquinic acid. Glutamic acid has been reported to prevent the browning of fresh potatoes by repressing the activity of polyphenol oxidase[35]. The higher contents of glutamic acid in J9 and M9 may reveal better applications of these in fresh potatoes (Fig. 4d). Furthermore, 3-O-feruloylquinic acid, as the subgroup of chlorogenic acid, showed various biological functions such as antioxidant and increases the tolerance of glucose to humans[36].
The differently expressed metabolites in Qingshu 9 from different areas of China were analyzed, as shown in Fig. 5. The results showed that potatoes from different areas of China contained larger differences in their metabolites such as J9 vs E9, M9 vs E9, and M9 vs J9 (Fig. 5a). These differently expressed metabolites reveal potential functional changes, leading to the variations of nutritional components in Qingshu 9 potatoes from different areas of China. Accordingly, the KEGG analysis of differently expressed metabolites was applied, as shown in Fig. 5b and c. The results showed that most of the different expressed metabolites were involved in phenylalanine, tyrosine, and tryptophan biosynthesis, flavone and flavonol biosynthesis, purine metabolism, and phenylpropanoid biosynthesis. Moreover, the network analysis for the differently expressed metabolites between Qingshu 9 from different areas of China was performed to identify the dominant metabolites and the top three networks are shown in Fig. 5d. The size and color of the nodes was used to present the degree and betweenness of these differently expressed metabolites. Their networks showed several dominant differently expressed metabolites including isoquercitrin, indoline, 2,3-dihydroxypropyl palmitate, 3-methylcrotonylglycine, p-coumaric acid, liensinine, and piperidine. Isoquercitrin is a powerful antioxidant and could prevent the oxidative-induced damage in plants[37], which may reveal the stress-resistant ability to soil environments for the first network. Moreover, 3-methylcrotonylglycine in the second network is an amino acid metabolite, while the detailed functions of it in potato are unknown. p-coumaric acid in the network has been reported to inhibit the browning of potatoes[38]. Overall, the differently expressed metabolites in Qingshu 9 from different areas of China were observed, and these metabolites were involved in various pathways and biological functions. This may indicate the potential large differences in their applications in the food industry.
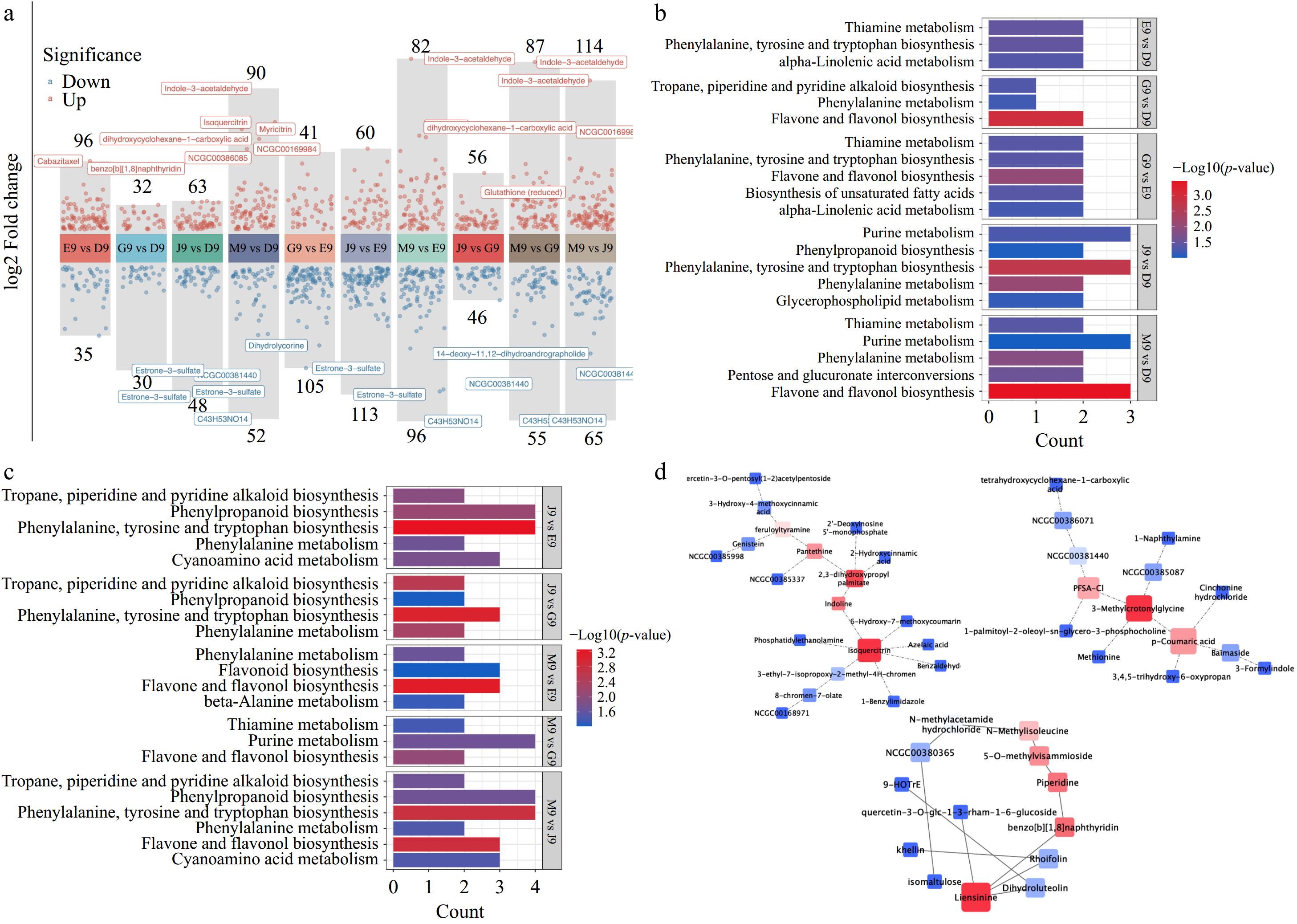
Figure 5.
(a) Volcano plots of metabolites in potatoes from different area of China. (b), (c) KEGG analysis of differently expressed metabolites. (d) Metabolite network analysis of differently expressed metabolites.
Correlation analysis of volatile and metabolic compounds
-
The above sections mention large differences in volatile and metabolite compounds in Qingshu 9 from different areas of China. To better understand the regulation of volatile compounds in Qingshu 9 from different areas of China, the conjoint analysis of metabolomics and GC-MS was applied, as shown in Fig. 6. The results from GC-MS showed large differences in volatile compounds in D9 and J9 with G9, E9, and M9. Accordingly, the common different expressed metabolites between D9 vs G9, D9 vs E9, D9 vs M9, and J9 vs G9, J9 vs E9, J9 vs M9 were used for the KEGG pathways shown as D9 vs E9M9G9, J9 vs E9G9M9, and J9 vs D9 (Fig. 6a). The results showed that three comparison groups showed the main involved KEGG pathways were phenylalanine, tyrosine, tryptophan biosynthesis, and phenylpropanoid biosynthesis. To further verify the relationships between these KEGG pathways and volatile compounds, the Pearson correlation analysis was performed in Fig. 6b. The results showed that the main discriminators from PCA and the volatile categories were negatively correlated with three KEGG pathways which were phenylalanine, tyrosine, and tryptophan biosynthesis, phenylpropanoid biosynthesis, and tropane, piperidine, and pyridine alkaloid biosynthesis. These KEGG pathways were involved in the production of numerous secondary metabolites that help plants defend against environmental factors. For example, a previous study reported that phenylpropanoid biosynthesis produced an enormous array of secondary metabolites based on shikimate and several intermediates as the core unit in phenylalanine, tyrosine, and tryptophan biosynthesis, and thus generated phytoalexins, volatiles, flavonoids, and anthocyanins[39]. Furthermore, the intensities of key metabolites in these KEGG pathways are shown in Fig. 6c. The results showed that these metabolites were significantly different among Qingshu 9 in different areas of China. For example, L-tyrosine, L-phenylalanine, and phosphatidylethanolamine contents were significantly lower in J9, and D9 contained higher contents of phosphatidylethanolamine (Fig. 6d). This is also in line with the boxplots for the key metabolites, which showed that most of these metabolites were lower expressed in D9 and J9.
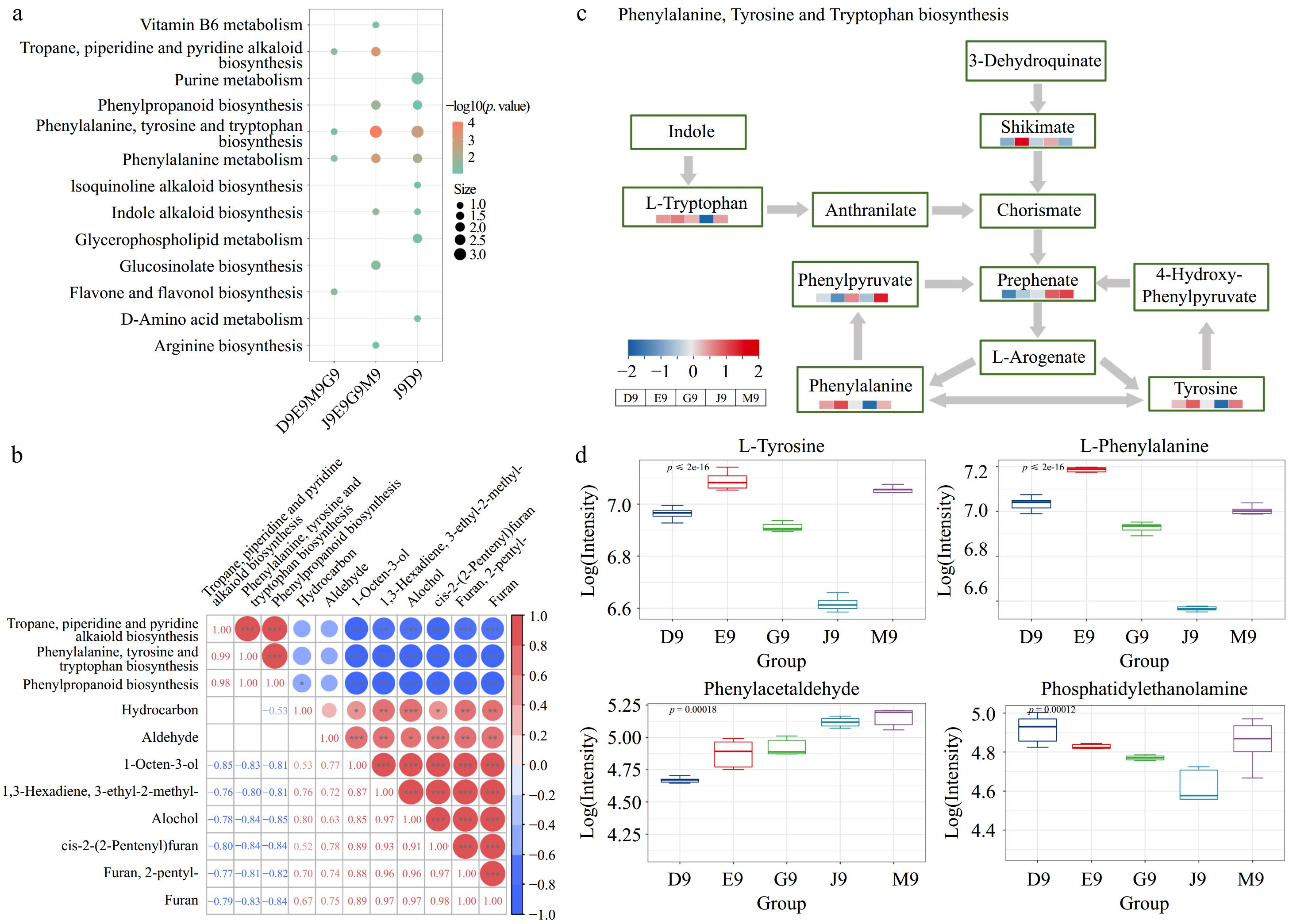
Figure 6.
Correlation analysis of volatile compounds and metabolites in potatoes from different areas of China. (a) Bubble plot of enriched KEGG pathways. (b) Pearson correlation analysis of volatile compounds and identified KEGG pathways. (c) KEGG analysis of identified metabolites in phenylalanine, tyrosine, and tryptophan biosynthesis. (d) Boxplots of several dominant metabolites.
The KEGG pathway map of phenylalanine, tyrosine, and tryptophan biosynthesis showed that several metabolites were differently expressed in potatoes from different areas of China. Of these, three aromatic amino acids, which were tyrosine, tryptophan, and phenylalanine, showed similar trends in Qingshu 9 from different areas of China. The results showed that three aromatic amino acids were lowest expressed in J9, but higher expressed in Qingshu 9 from four other areas especially in E9. A previous study reported that aromatic amino acids were involved in the synthesis of proteins and a variety of secondary metabolites, and a subset of them were involved in the synthesis of pigment compounds, plant hormones, and biological polymers[40]. Of these secondary metabolites, some of them were volatile compounds such as phenylpropanoids, benzenoids, phenylpropenes, and methylbenzoate, which were involved in plant reproduction and defense response[41]. Furthermore, shikimate plays a critical role in the synthesis of aromatic amino acids[41], which was highly expressed in E9 and J9, but lower in other areas. A previous study mentioned that phenylpyruvate could serve as a precursor for the synthesis of several volatile compounds, including phenylacetaldehyde, 2-phenylethanol, and 2-phenylethyl β-D-glucopyranoside[42], which was higher expressed in G9 and M9. This reveals that the large differences in aromatic amino acid biosynthesis and metabolism resulted in the variations of volatile compounds in Qingshu 9 from different areas of China. Overall, the combination analysis between volatile and metabolite compounds showed that the large differences in the shikimate pathway-induced aromatic amino acid synthesis resulting in the variations of volatile compounds in Qingshu 9 from different areas of China. The differences in aromatic amino acid synthesis may be caused by the production of secondary metabolites against the environment.
Influences of geographical factors on volatile and metabolite compounds
-
Based on these results, it is clear that volatile and metabolite compounds were significantly different in the same potato variety Qingshu 9 from different areas of China. Pearson correlation analysis between volatile and metabolites categories was conducted, as shown in Fig. 7. Strong correlations between metabolites and volatile compounds were found. For example, the contents of saccharide and peptides were positively correlated with most of the volatile compounds including aldehydes, hydrocarbons, and alcohol compounds. Moreover, aldehyde compounds were positively correlated with the contents of alkaloid (R2 = 0.64) and phenols (R2 = 0.90). A previous study reported that the alkaloid biosynthesis in plants typically requires an aldehyde precursor[43]. In contrast, amino acid contents were found to be negatively correlated with alcohol (R2 = −0.89), furan (R2 = −0.87), aromatic compounds (R2 = −0.81), ketones (R2 = −0.84), and slightly correlated with hydrocarbons (R2 = −0.63). This may because amino acids are precursors of aroma volatiles in biological systems[44].
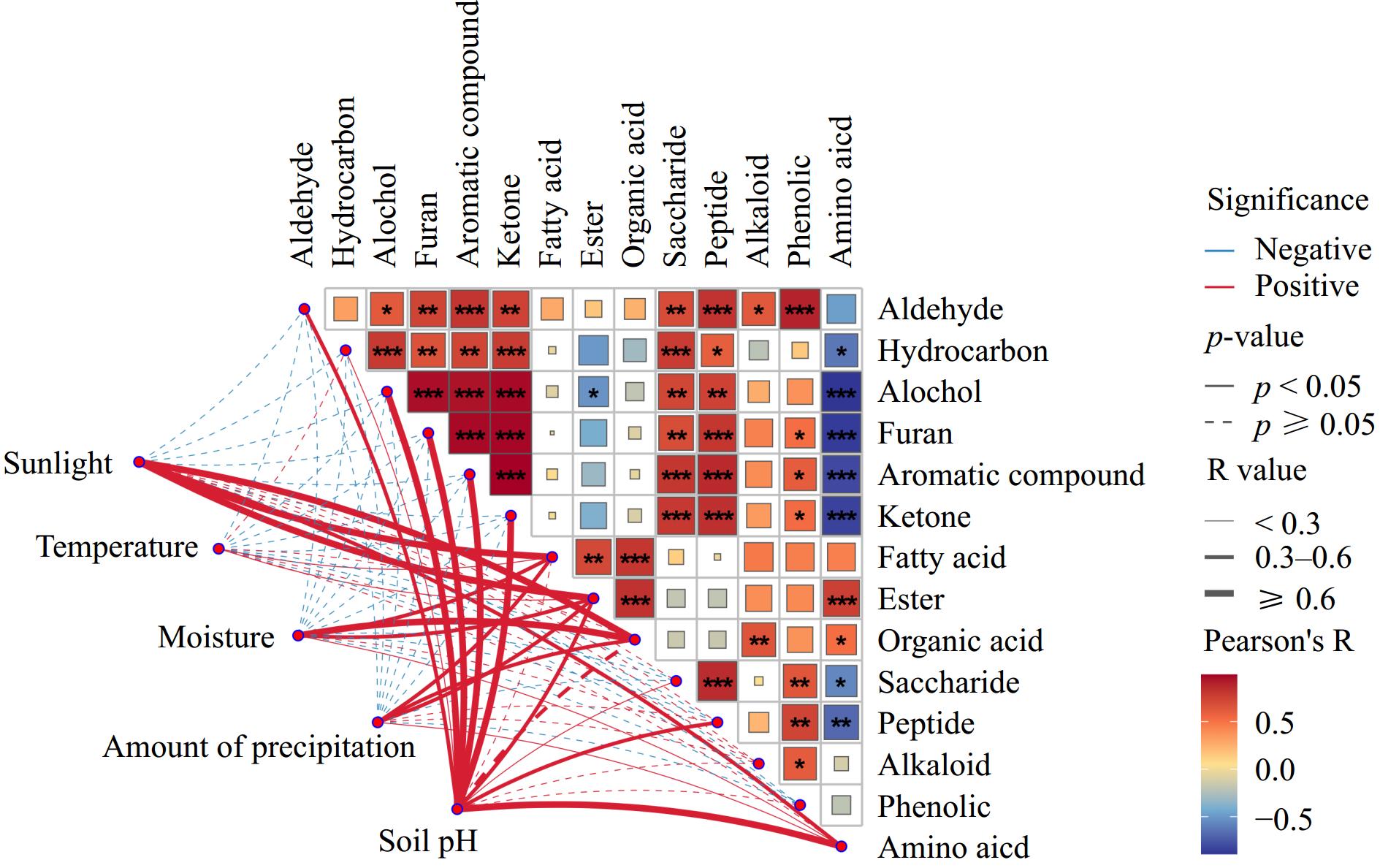
Figure 7.
Correlation analysis between geographical and environmental factors with volatile and metabolite compounds.
To study the influences of geographical factors on potato, the geographical information including sunlight, temperature, moisture, and soil pH was collected from local government websites (Supplementary Table S1). Pearson analysis between geographical information with volatile and metabolite categories was performed (Fig. 7). The results showed that soil pH plays a critical role in the formation of potato volatile and metabolite compounds. This aligns with previous studies which demonstrated that potato quality was influenced by the interaction of soil nutrients.[45] For example, soil pH was found to be most strongly positive correlated with the intensity of metabolite such as amino acids and peptides, volatile compounds in particle including aldehydes, hydrocarbons, alcohols, furan, and aromatic compounds. Moreover, the average time of sunlight, moisture, and amount of precipitation have been found to be related to the intensities of fatty acids, organic acids, and esters. Sunlight was also positively correlated with the intensity of amino acids. However, the average temperature was found to be only slightly correlated with fatty acids and esters. A previous study reported that environmental conditions such as temperature, moisture, and especially soil pH, played an important role in volatile organic compound emissions in maize cropping systems[46]. Numerous studies reported that soil and plant-associated microorganisms produced a group of largely secondary metabolites including volatile compounds[47]. This indicates that the influences of soil pH on metabolites and volatile compounds were related to the soil bacterial content. A previous study reported that the soil bacterial abundance significantly increased the soil pH[48], and the higher soil pH increased the abundance of metabolite and volatile compounds in potatoes. For example, the soil pH for J9 (9.75), G9 (7.0), and D9 (6.27) was higher than that in M9 (5.7), and E9 (5.0) (Supplementary Table S1), and higher abundances of metabolites and volatile compounds in Qingshu 9 from these provinces were observed (Figs 2 and 3).
The influence of sunlight on potato composition may be caused by photosynthesis and affect potato tubers. For example, Sasaki et al. demonstrated that fatty acid synthesis in chloroplasts was regulated by light and catalyzed by acetyl-CoA carboxylase[49]. The moisture and amount of precipitation in different areas of China mainly affected the soil conditions[50], which may result in the changes in fatty acids, organic acids, and esters in Qingshu 9 in response to environmental stresses. For example, fatty acid abundance as general defenders play critical roles against abiotic stresses to avoid or abide the adverse effects[51]. Overall, it is clear that geographical factors such as soil pH and moisture play critical roles in the formation of potato composition and metabolite compounds in Qingshu 9. This may be caused by the stress resistance such as soil microbiome, cold, and salt, thus producing a variety of secondary metabolites.
-
In conclusion, this study analyzed the volatile compounds and metabolites in Qingshu 9 potatoes from five areas of China and discussed the correlations with geographical factors. The results indicated the large differences in nutritional components between Qingshu 9 from different areas of China, and the shikimate pathway showed critical roles in aromatic amino acid synthesis resulting in the variations of volatile compounds in Qingshu 9 from different areas of China. Geographical factors, particularly soil pH, moisture, and sunlight, were found to affect the formation of volatile and metabolite compounds in potatoes. Future studies should focus on the relationship between soil microbiomes and potato composition and further verify the results of this study. These results will contribute to the production of potato products in the food industry in different areas of China.
This work was supported by The Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-IVFCAAS)
-
The authors confirm contribution to the paper as follows: study conception and design: Zhang W, Deng J; experimental processing: Li L, Zhao Y, Wang W; data collection: Xu Z, Wang X; analysis and interpretation of results: Zhang W, Huang T, Wang Q; draft manuscript preparation: Zhang W, Deng J, Yang H. All authors contributed to the discussion, reviewed the results, and approved the final version of the manuscript.
-
The data will be made available upon reasonable request from the corresponding author.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The geographical information of Qingshu 9 from different areas in China.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang W, Li L, Zhao Y, Huang T, Wang Q, et al. 2025. The variations of volatile and metabolite compounds in potato from different areas of China: the critical role of geographical factors. Food Innovation and Advances 4(4): 461−470 doi: 10.48130/fia-0025-0041 

The variations of volatile and metabolite compounds in potato from different areas of China: the critical role of geographical factors
- Received: 28 March 2025
- Revised: 16 June 2025
- Accepted: 18 June 2025
- Published online: 29 October 2025
Abstract: Potato is an important food resource around the world. Potato cultivar Qingshu 9 is widely planted in China. To study the influence of geographical factors on potato composition, Qingshu 9 was planted in five areas of China and the resulting potatoes were collected and analysed. The results showed large differences in basic nutrients, volatile compounds, and metabolites between Qingshu 9 from different areas. The characteristic volatile and metabolite compounds in different areas were identified, such as 2-pentyl-Furan in J9 from Shanxi; methyl-cyclopentane in D9 from Yunnan; and ferulic acid in M9 from Inner Mongolia. The differences in volatile compounds were correlated with the shikimate pathway-induced aromatic amino acid synthesis. Geographical factors, such as soil pH, sunlight, and moisture, were significantly correlated with the volatile and metabolite compounds in Qingshu 9. Together, these results highlight the significant influence of cultivation region and geographical factors on the metabolic composition and flavor profiles of potatoes, providing reference for the optimization of regional farming practices and demand-oriented potato selection in the food industry.
-
Key words:
- Potato /
- Qingshu 9 /
- Volatile compounds /
- Metabolite /
- Geographical factors