










-
Soil salinity is a major abiotic stress that severely disrupts plant growth by impairing root development, inducing oxidative damage, and destabilizing hormonal homeostasis[1]. To mitigate salt toxicity, plants deploy a variety of strategies, including architectural remodeling of roots, accumulation of osmoregulatory metabolites, and activation of antioxidant systems[2]. As the primary organ sensing salinity, the root perceives salt stress and responds by promoting downward growth to access deeper soil layers, thereby enhancing water and nutrient acquisition, while simultaneously limiting salt influx[3]. Flavonoids function as non-enzymatic antioxidants that are pivotal for scavenging reactive oxygen species (ROS), and maintaining redox equilibrium under stress[4,5].
Plant growth and stress responses are tightly regulated by phytohormones, which coordinate physiological adaptations to environmental cues. Under salt stress, hormonal networks, especially the interplay between auxin-mediated growth regulation and abscisic acid (ABA)-driven stress responses, are crucial for maintaining cellular homeostasis[6]. Auxin, a key regulator of plant development, mediates root architectural adjustments essential for nutrient acquisition in saline soils[7]. Meanwhile, ABA acts as a central hub for stress tolerance, orchestrating stomatal closure to curtail water loss and simultaneously driving root elongation to enhance water acquisition[8,9]. The crosstalk between antioxidant pathways and hormonal signaling is critical for balancing the inherent trade-off between growth and stress defense in plants facing salinity.
Plants coordinate biochemical reactions and metabolic pathways through a complex network of key genes. NAC transcription factors (TFs) function as master regulators that integrate these adaptive responses, enhancing stress resilience by directly activating antioxidant biosynthesis genes and bridging with hormonal pathways[10]. For example, NAC TFs enhance ABA signaling to improve stomatal closure and ROS detoxification, while simultaneously facilitating auxin-driven root plasticity in species such as Vitis vinifera and poplar[11]. In addition, the Arabidopsis NAC gene SMB regulates AUX1 expression, thereby activating auxin signaling and root halophytic responses[12]. However, the mechanisms by which NAC proteins coordinate flavonoid-antioxidant systems with auxin-ABA crosstalk to balance salt adaptation in economically important tree species remain unclear.
Ginkgo biloba, a medicinal tree highly valued for its flavonoid content, exhibits severe growth suppression under saline conditions. Although NAC TFs such as GbNAC2 have been previously identified as salt-responsive genes in ginkgo[13], their functional mechanisms in integrating antioxidant and hormonal pathways remain unexplored. In the present study, GbNAC2 was identified as a critical salinity-tolerance gene, revealing its ability to enhance salt resistance through multiple synergistic strategies. Specifically, GbNAC2 promotes flavonoid accumulation to strengthen antioxidant defenses, modulates auxin signaling to optimize root architecture for stress adaptation, and activates ABA-mediated pathways to amplify salt resilience. These findings provide critical insights into the molecular mechanisms underlying salt tolerance in ginkgo, serving as a valuable genetic resource for the development of salt-tolerant ginkgo through biotechnological breeding.
-
Surface-sterilization of Ginkgo seeds (Fozhi cultivar) was performed using 15% sodium hypochlorite for 15 min, followed by three rinses with sterile water. Subsequently, the seeds were placed in an incubator set at 25 °C for a 10-d germination period. Seeds with uniform germination were chosen and transplanted into pots containing nutrient-dense soil. The seedlings were cultivated in a growth cabinet at 25 °C, under a photoperiod of 16 h of light and 8 h of darkness. Ginkgo seedlings cultivated for 2 months, with good growth status and similar plant height and stem diameter, were used for various experiments. The sampling standard for leaves was the middle part of each plant.
For salt treatment, seedlings were treated with 200 mM NaCl as described in previous research[13]. For ABA treatment, leaves were sprayed with a 70 μM ABA solution. For combined NaCl and ABA treatment, seedlings were first sprayed with 20 μM ABA, followed by 400 mM NaCl treatment the next day. After 3 d of NaCl treatment, seedlings' phenotypes were observed, and leaf samples were collected for further analysis.
Phylogenetic analysis
-
Homologous protein sequences of the GbNAC2 and GbAREB3 from other species were obtained from the National Center for Biotechnology Information (NCBI) database. The protein sequences were aligned using ClustalW, and a neighbor-joining (NJ) phylogenetic tree was constructed using MEGA software[14]. The corresponding accession numbers (IDs) of genes are provided in Supplementary Table S1.
Expression analysis
-
Total RNA was extracted from various ginkgo tissues (root, stem, leaf, embryo, ovule, and stamen), calli, and transgenic plant leaves utilizing the RNA Extraction Kit (Nanjing, Vazyme). Samples were pulverized into a fine powder in liquid nitrogen, and 0.1 g of this powder was subjected to RNA extraction. The RNA was validated for concentration using NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA). Quantitative real-time PCR (qRT-PCR) analysis was conducted with SYBR qPCR Master Mix (Nanjing, Vazyme), with G. biloba GAPDH serving as the reference gene for normalizing relative expression levels. Relative gene expression was computed using the 2−ΔΔCᴛ method[15]. The primer sequences used are provided in Supplementary Table S2.
Subcellular localization
-
The coding sequence (CDS) of GbNAC2 was inserted into the pACT2 vector to generate a GbNAC2-green fluorescent protein (GFP) fusion construct (35S::GbNAC2-GFP). The plasmid was transformed into competent cells of the Agrobacterium tumefaciens (A. tumefaciens) strain GV3101. Subsequently, 1-month-old tobacco plants with healthy growth were selected for transient transformation via infiltration with Agrobacterium harboring 35S::GbNAC2-GFP, following the method described by Liu et al.[16]. Leaf samples were collected after 48–72 h of dark incubation, and GbNAC2 localization was determined by observing GFP fluorescence using a laser confocal microscope. The primer sequences used are provided in Supplementary Table S2.
Heterologous overexpression in Arabidopsis and poplar
-
To obtain stable transgenic Arabidopsis plants, 1-month-old wild-type (WT) Arabidopsis was transformed using the floral dip method as previously described[17]. The Arabidopsis transformation was conducted using the GV3101 strain of A. tumefaciens. For the transformation of '84K' poplar, an Agrobacterium-mediated leaf transformation method was used as described by Wen et al.[18]. The selected plants were then cultivated for approximately 4 weeks, after which genomic DNA and RNA were extracted for verification of successful transformation.
Transient overexpression of GbNAC2 in ginkgo calli
-
To analyze the downstream genes regulated by GbNAC2, the CDS of GbNAC2 was constructed into the BamHI restriction site of the pRI101-AN vector, yielding the overexpression vector GbNAC2-pRI. Agrobacterium-mediated transformation was used to introduce the recombinant plasmid into ginkgo calli, following the procedures outlined in a previous study[19]. The primer sequences used are provided in Supplementary Table S2.
Determination of physiological indexes
-
The detection of flavonoids was performed according to the previous method by Xu et al.[13] using a flavonoid detection kit (Cat#BC1330; Suzhou Conmin Biotechnology Co., Ltd.). Samples were dried to constant weight and extracted using the ultrasonic extraction method (300 Hz, 60 °C, 30 min). After extraction, the mixture was centrifuged at 12,000 rpm at 25 °C for 10 min, and the absorbance of the resulting supernatant was determined at 470 nm via spectrophotometry.
The determination of the contents of hydrogen peroxide (H2O2) and malondialdehyde (MDA), as well as the determination of the activities of catalase (CAT) and superoxide dismutase (SOD), were all performed according to previous methods[20]. Additionally, the ultraviolet spectrophotometry was used in accordance with the manufacturer's instructions (Suzhou Conmin Biotechnology Co., Ltd.). Specifically, CAT (Cat#BC0200), SOD (Cat#BC01700), H2O2 (Cat#BC3590), and MDA (Cat#BC0020) content assay kits were used respectively. CAT activity was determined at 240 nm, with results expressed as U·g−1; SOD activity was assayed at 560 nm, also quantified as U·g−1; H2O2 content was measured at 415 nm, with the unit of μmol·g−1; MDA concentration was determined at dual wavelengths of 532 and 600 nm, and expressed as nmol·g−1.
Germination experiment of Arabidopsis seeds
-
Seeds of WT Arabidopsis and GbNAC2 transgenic Arabidopsis were surface-sterilized with 15% sodium hypochlorite, and 70% ethanol, respectively. Subsequently, the seeds were evenly sown on 1/2 MS solid medium, 1/2 MS solid medium supplemented with 100 mM NaCl, and 1/2 MS solid medium supplemented with 200 mM NaCl. After stratification at 4 °C for 3 d, the seeds were transferred to a growth chamber at 26 °C. Then, the germination rate was assessed after 7 d.
Transcriptome sequencing and DEGs analysis
-
RNA sequencing was performed using ginkgo callus samples with empty vector (WT), and GbNAC2-overexpression (GbNAC2-OE), with three biological replicates set for each treatment. After the samples were ground into fine powder in liquid nitrogen, total RNA was extracted using the RNAprep Pure Plus Kit (TIANGEN, Beijing, China) according to the operating protocol provided by the manufacturer. RNA integrity was evaluated using the RNA Nano 6000 Assay Kit matched with the Bioanalyzer 2100 System (Agilent Technologies, CA, USA). A total of six libraries were constructed in the experiment, and subsequent sequencing was completed using the Illumina NovaSeq sequencing platform at Novogene (Beijing, China).
Quality control was performed on the raw data to obtain clean reads through the following specific steps: removing adapter sequences from the reads, discarding reads with more than 10% unknown bases (N), and filtering out reads with a sequencing quality score lower than Q20. Filtered clean reads were aligned to the ginkgo reference genome[21]. Gene expression levels were quantified as fragments per kilobase of transcript per million mapped reads (FPKM). Differentially expressed genes (DEGs) between the WT and GbNAC2-OE groups were identified using the DESeq2 package in R (version 1.16.1), based on p-value < 0.05, and fold change ≥ 2. Functional enrichment analysis of the DEGs was conducted according to the KEGG database (
www.genome.jp/kegg )[22].Transcriptional activation analysis of GbNAC2 protein
-
The GbNAC2 CDS was constructed into the BamHI restriction sites of the pGBKT7 (BD) vector following the method described by Hou et al.[23]. The BD-GbNAC2 plasmid was co-transformed with the empty vector pGADT7 (AD) into the Y2H Gold yeast strain. Following transformation, the yeast cells were spread onto SD/-Leu/-Trp selective medium (DDO), and incubated at 30 °C for 2–4 d. Subsequently, the yeast strains were transferred into the SD/-Leu/-Trp/-His/-Ade/5-bromo-4-chloro-3-indolyl-α-d-galactopyranoside (X-α-Gal) selective medium (QDO). The plates were incubated again at 30 °C for 2–4 d, during which the development of the colonies and any color changes were monitored. The appearance of blue yeast strains indicated the presence of transcriptional activation activity. The primer sequences used are provided in Supplementary Table S2.
Yeast one-hybrid (Y1H) assay
-
Previous studies have described the methods for performing Y1H[24]. The promoter sequence (2000 bp) of GbAREB3 and the GbNAC2 CDS were respectively inserted into the pLacZi-2μ and pb42AD vectors. The constructs were co-transformed into competent yeast cells (EGY48), which were then plated on selection medium (SD/-Ura/-Trp), and incubated at 28 °C for ~3 d. Yeast monoclonal colonies from the selection media were transferred to a color development medium containing 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-β-gal), and Buffer (BU) salt and incubated in a 28 °C incubator for ~3 d. If the target protein binds to DNA, the colonies will exhibit a blue color. The primer sequences used are provided in Supplementary Table S2.
Statistical analysis
-
All experiments were performed with at least three biological replicates. Asterisks indicate significant differences by two-sided Student's t-test (* p < 0.05, ** p < 0.01). Different letters indicate significant differences by one-way analysis of variance (ANOVA, p < 0.05). Error bars represent the mean ± standard deviation (SD).
-
A previous study has shown that NAC genes in ginkgo exhibit significant responses to salt stress[13]. In this study, four NAC genes positively responsive to salt stress were identified from RNA-seq data of salt-treated ginkgo leaves (Fig. 1a). To determine which NAC genes participate in the salt stress response in Ginkgo, seedlings were treated with 200 mM NaCl. The expression patterns of the four salt-induced NAC genes were analyzed by qRT-PCR within the first 24 h following treatment. Among them, Gb_41540 showed a rapid induction within 3 h and maintained elevated expression levels in the leaves for up to 24 h (Fig. 1b).
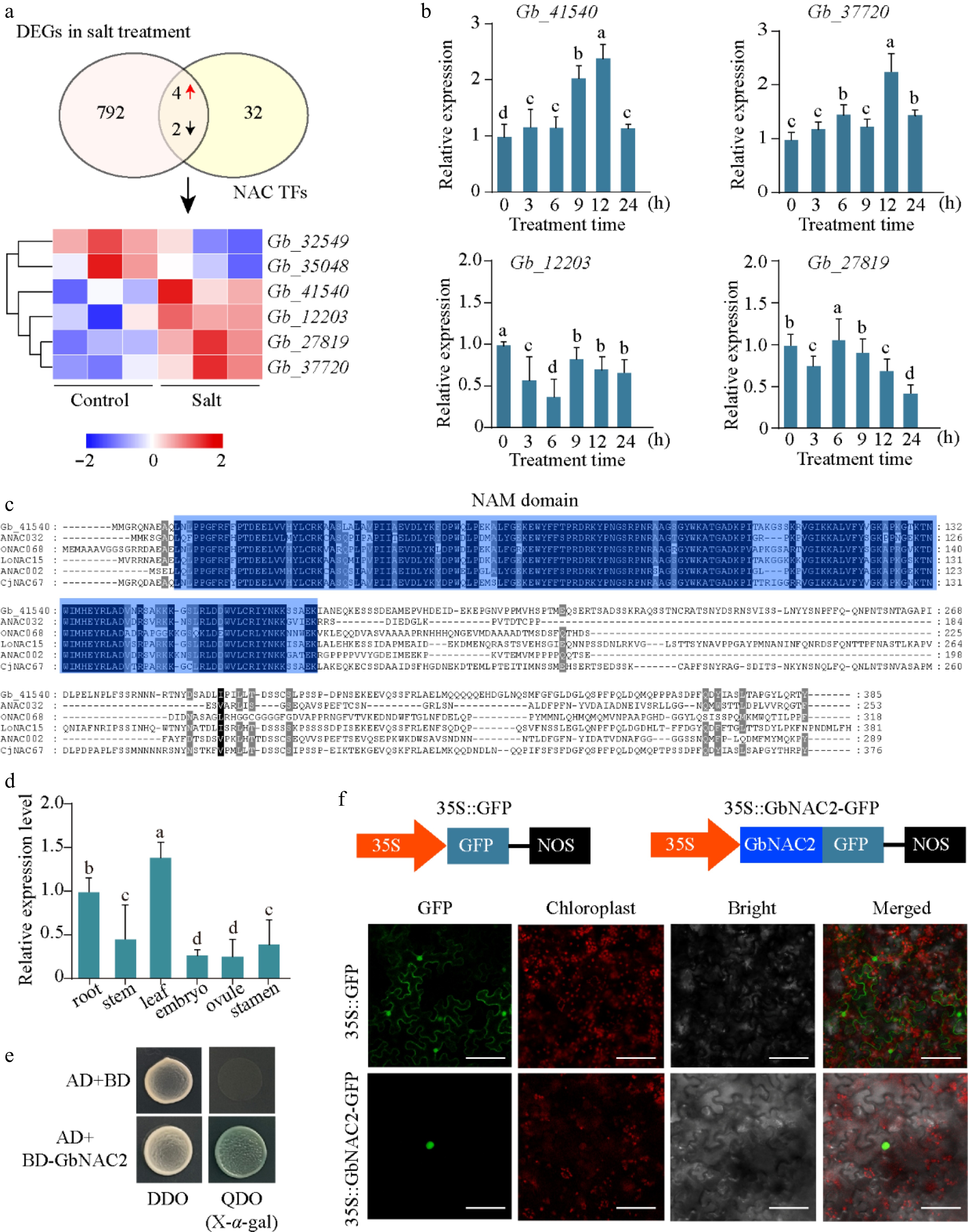
Figure 1.
Identification of key NAC TFs. (a) Overlap analysis of NAC genes in G. biloba and DEGs under salt stress transcriptome. The FPKM value of six key NACs following salt treatment are depicted in a heatmap (red represents upregulated, and blue represents downregulated). (b) The relative expression of four NAC genes at different time points following salt treatment was quantified using qPCR. (c) Sequence analysis of GbNAC2 and other NAC TFs from different plant species. (d) The spatial-temporal expression of GbNAC2 in different issues in ginkgo. (e) Analysis of transcriptional activation activity of GbNAC2. (f) Subcellular localization of the GbNAC2 protein. Scale bar = 10 μm. Different letters indicate significant differences (one-way analysis of variance, p < 0.05).
Gb_41540 contains an open reading frame of 1,158 bp and is predicted to encode a 385-amino-acid protein featuring a conserved NAM domain typical of the NAC transcription factor family (Fig. 1c). Phylogenetic analysis showed that the Gb_41540 protein was clustered together with NAC002 from A. thaliana, and Gb_41540 was referred to as GbNAC2 (Supplementary Fig. S1). Tissue-specific expression analysis revealed that the highest GbNAC2 expression in leaves and roots (Fig. 1d). To test the transcriptional activation activity of the GbNAC2 protein, the full-length GbNAC2 was fused to the GAL4 DNA-binding domain of the pGBKT7 (BD) vector and transformed into yeast. The yeast's growth and blue color on quadruple-dropout medium with X-α-gal indicated GbNAC2 has transcriptional activation (Fig. 1e). Additionally, the transient transformation of tobacco confirmed the nuclear localization of GbNAC2 (Fig. 1f).
Overexpressing GbNAC2 promotes root development
-
To explore the functional role of GbNAC2, GbNAC2 overexpression Arabidopsis transgenic lines (GbNAC2-OE) were generated (Fig. 2a, d). The GbNAC2-OE Arabidopsis displayed accelerated growth and earlier flowering phenotype compared to the WT (Fig. 2e). Additionally, transgenic Arabidopsis roots were 1.6–2.0-fold longer than those of the WT plants (Fig. 2f, h). GbNAC2-OE transgenic poplar lines were also generated (Fig. 2c, d). In consistent with GbNAC2-OE Arabidopsis lines, compared to the WT, the number of lateral roots in GbNAC2-OE poplar plants was significantly increased, and root length was approximately 1.6- to 2.4-fold longer than in WT plants (Fig. 2g, i). Furthermore, correlation analysis results displayed a strong association between the expression level of GbNAC2 and root length in transgenic plants (Fig. 2j, k).
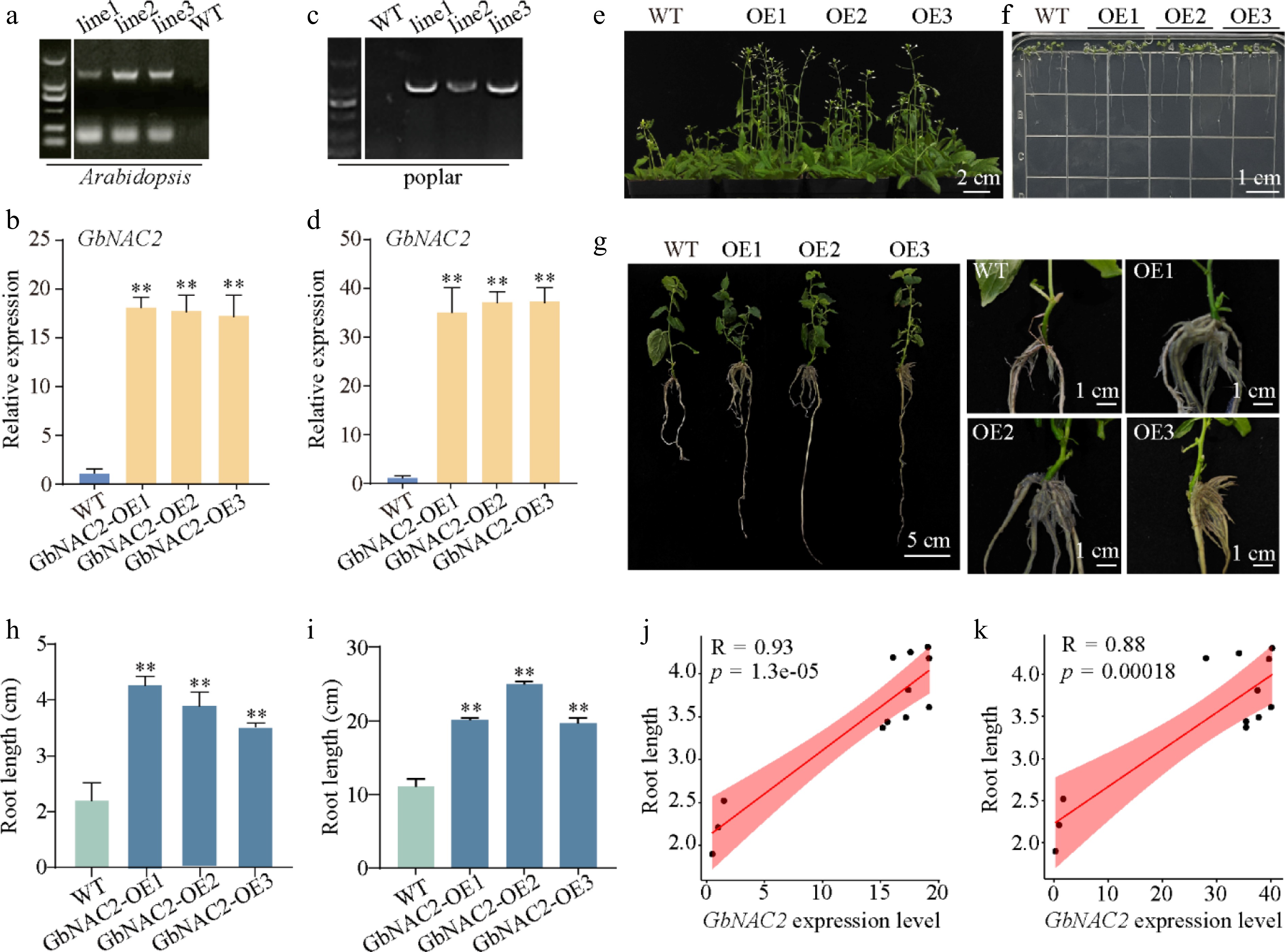
Figure 2.
Phenotypic analysis of overexpressed GbNAC2 in Arabidopsis and poplar. The (a), (c) positive detection and (b), (d) GbNAC2 expression level in transgenic plants. Ectopic expression of GbNAC2 significantly promotes the (e) flowering, and (f) root development of WT Arabidopsis. (g) The ectopic expression of GbNAC2 markedly enhances the development of primary and lateral roots in poplar. A statistical analysis of the root length of (h) transgenic Arabidopsis, and (i) transgenic poplar was conducted. The relationship between GbNAC2 expression and root length in (j) transgenic Arabidopsis, and (k) poplar. ** p < 0.01.
GbNAC2 enhances salt tolerance by inhibiting ROS accumulation
-
To assess the role of GbNAC2 in salt stress tolerance, WT plants showed pronounced leaf wilting and bleaching under salt stress, whereas GbNAC2-OE lines exhibited much milder chlorosis (Fig. 3a). The MDA content, which is a key parameter of stress tolerance, within these plants was further examined[25]. After salt treatments, the accumulation of MDA in WT plants was significantly higher than that in the GbNAC2-OE plants (Fig. 3b). Additionally, salt stress also increased the content of H2O2 in all plants, but the increase rate was less in GbNAC2-OE lines compared to WT (Fig. 3c). Antioxidant enzyme activities were also measured, finding that both CAT and SOD activities were significantly higher in the GbNAC2-OE lines than in WT plants (Fig. 3d, e). Germination assays showed no difference between WT and transgenic Arabidopsis under normal conditions, but under salt stress, transgenic seeds displayed significantly higher germination rates (Supplementary Fig. S2).
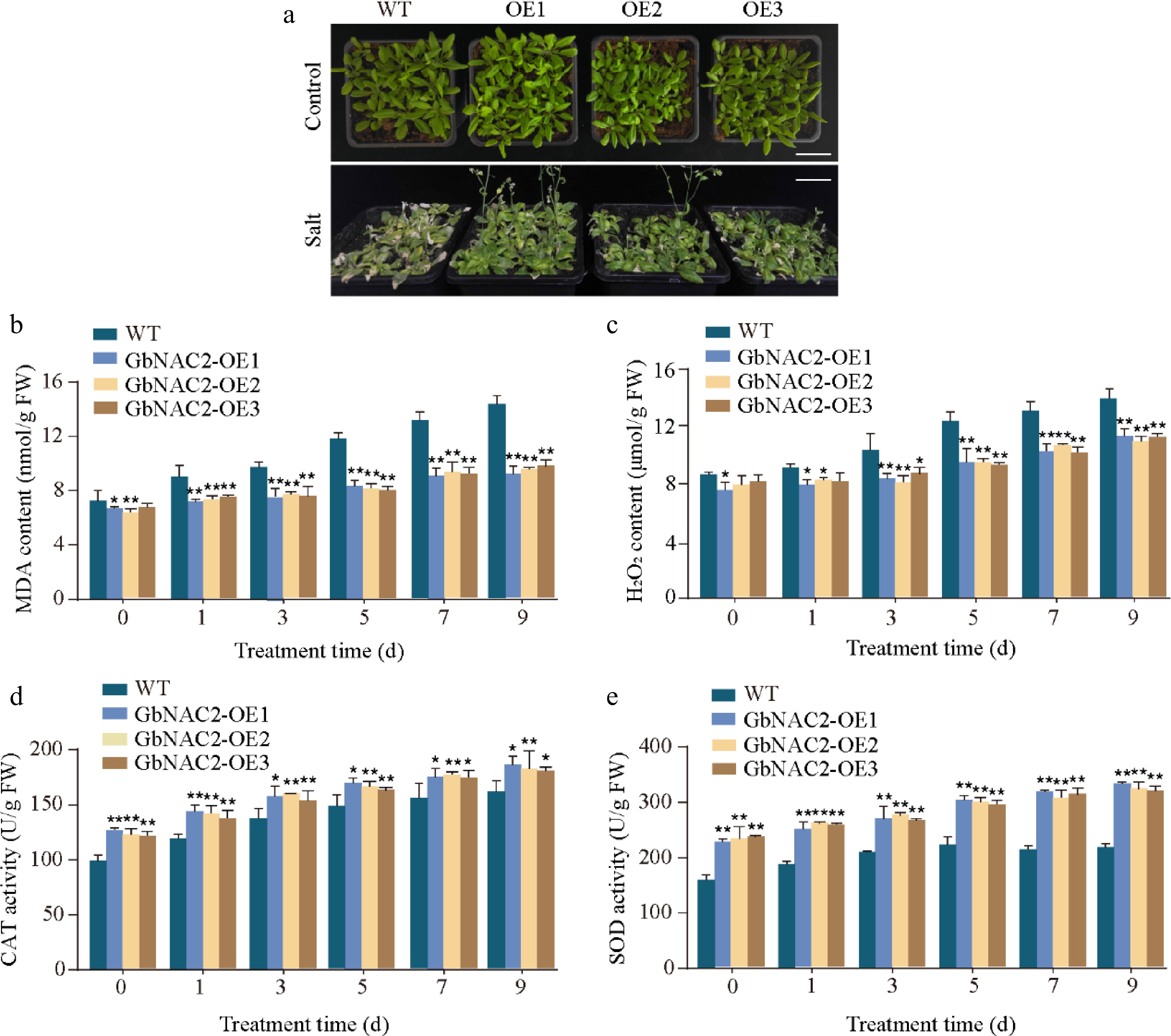
Figure 3.
Overexpression of GbNAC2 enhances the salt tolerance in Arabidopsis. (a) Phenotypic appearance of the plants after 9 d of salt treatment. (b) Levels of MDA in leaves. (c) Levels of H2O2 in leaves. (d) Activities of CAT in leaves. (e) Activities of SOD in leaves. * p < 0.05, ** p < 0.01.
Similar results were observed in GbNAC2-OE poplar. WT poplars showed noticeable leaf yellowing after 5 d of salt treatment, with most leaves wilting and dying by day 9. In contrast, GbNAC2-OE lines maintained healthy growth throughout the treatment period (Fig. 4a). Physiological analyses under salt stress revealed that GbNAC2-OE poplars consistently had lower levels of H2O2 and MDA, along with higher CAT and SOD activities compared to WT plants (Fig. 4b, e). Together, these findings suggest that GbNAC2 significantly enhances salt tolerance by reducing oxidative stress and boosting antioxidant enzyme activity, leading to improved growth and survival under salt stress conditions.
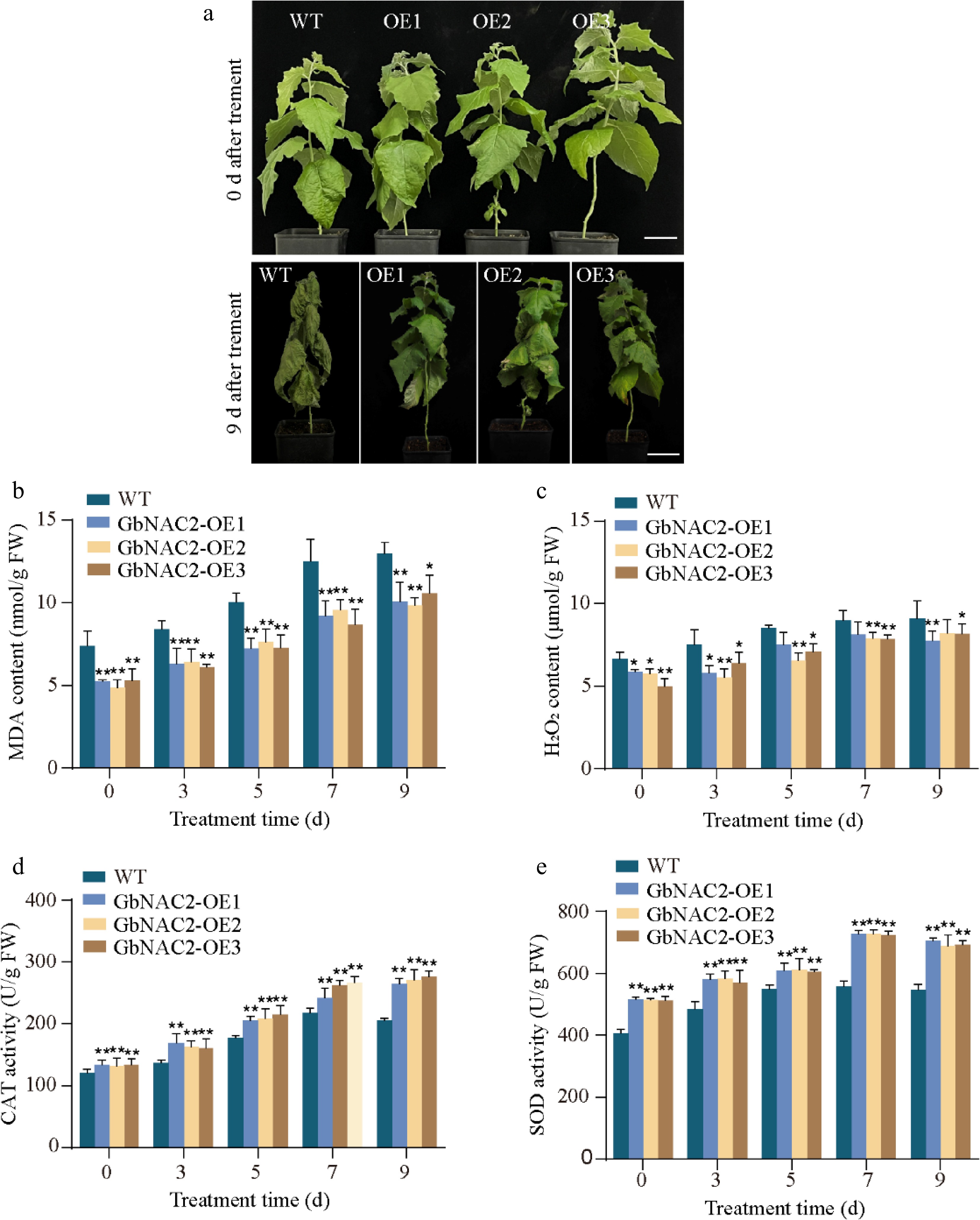
Figure 4.
Overexpression of GbNAC2 enhances the salt tolerance in poplar. (a) Phenotypic appearance of the plants under salt treatment. (b) Levels of MDA in leaves. (c) Levels of H2O2 in leaves. (d) Activities of CAT in leaves. (e) Activities of SOD in leaves. * p < 0.05, ** p < 0.01.
GbNAC2 promotes flavonoid biosynthesis
-
The flavonoid content in GbNAC2-OE lines after salt treatment were measured. Prior to treatment, the flavonoid content in transgenic Arabidopsis and poplar were 1.2- and 1.3-fold that of the WT, respectively. During salt stress, flavonoid content in WT plants initially increased but then declined, whereas in GbNAC2-OE Arabidopsis, flavonoid levels steadily rose throughout the treatment. After 9 d of salt exposure, flavonoid content in the transgenic Arabidopsis was about 2.1 times higher than in WT plants (Fig. 5a). A similar pattern was observed in poplar, with flavonoid levels in GbNAC2-OE lines reaching approximately 1.4-fold that of WT after 9 d (Fig. 5b).
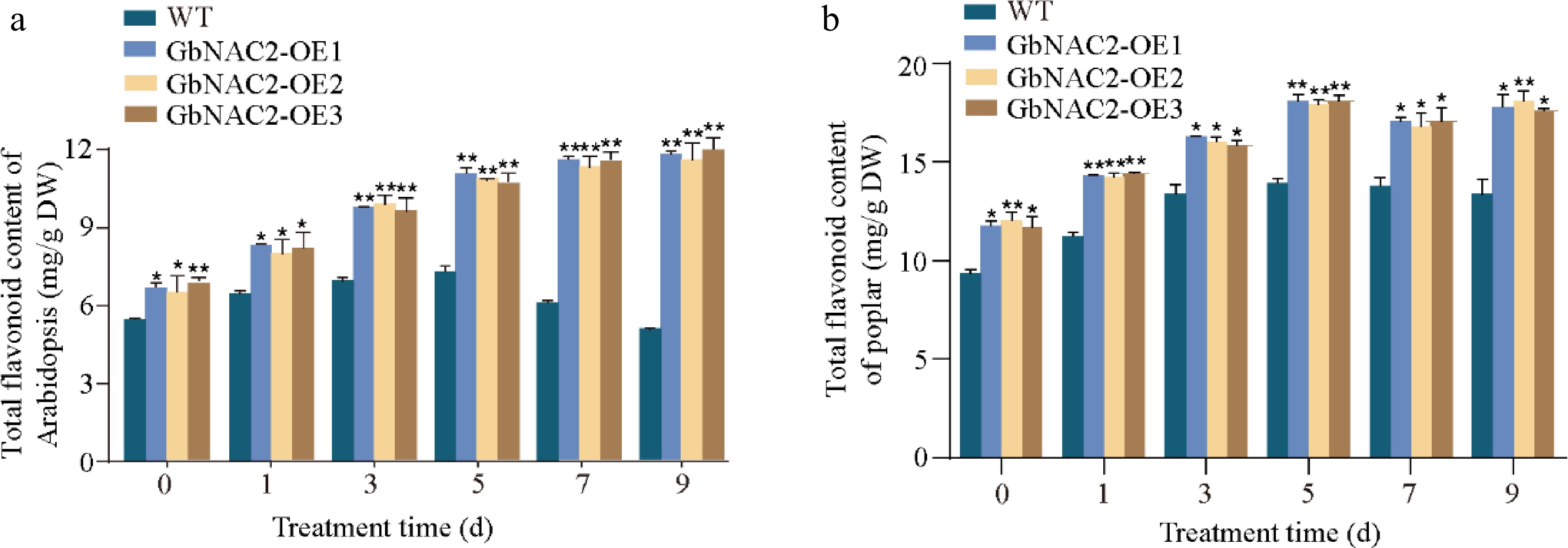
Figure 5.
Determination of flavonoid content at different time points following salt treatment in transgenic (a) Arabidopsis, and (b) transgenic poplar. * p < 0.05, ** p < 0.01.
To elucidate the molecular mechanisms how GbNAC2-mediated salt response, GbNAC2 was overexpressed in ginkgo calli, followed by transcriptomic analysis. The expression level of GbNAC2 in the calli was significantly higher compared to non-transformed samples, suggesting successful overexpression of GbNAC2 in calli (Fig. 6a). A total of 2,043 differentially expressed genes (DEGs) were identified between the GbNAC2-OE calli and control, with 1,059 genes upregulated, and 984 genes downregulated (Supplementary Fig. S3). KEGG enrichment analysis demonstrated significant enrichment of these DEGs in phenylpropanoid biosynthesis, flavonoid biosynthesis, and plant hormone signal transduction pathways (Fig. 6b). Given that the overexpression of GbNAC2 significantly promotes the synthesis of flavonoids, key genes related to the flavonoid biosynthesis pathway were focused on. Critical structural genes involved in the flavonoid biosynthesis pathway exhibited coordinated upregulation, including cinnamate 4-hydroxylase (C4H), chalcone synthase (CHS), flavanone 3-hydroxylase (F3H), flavonoid 3′,5′-hydroxylase (F3′5′H), and dihydroflavonol 4-reductase (DFR) (Fig. 6c). These transcriptional changes were validated by qRT-PCR, which confirmed consistent upregulation patterns (Fig. 6d). These results suggest that GbNAC2 functions as a regulatory hub to amplify flavonoid biosynthesis by modulating the expression of key flavonoid biosynthesis pathway genes under salt stress.

Figure 6.
Transcriptome analysis reveals downstream signaling pathways regulated by GbNAC2. (a) Expression of GbNAC2 in GbNAC2-OE ginkgo calli. (b) Enrichment analysis of DEGs in the KEGG pathways. (c) A heatmap is presented to illustrate the transcriptional changes of genes in the flavonoid pathway between WT and transgenic calli (red represents upregulated, and blue represents downregulated). (d) qPCR analysis of key structural genes. ** p < 0.01.
Identification of genes involved in hormone signaling pathways
-
Systematic analysis of hormone-related DEGs in GbNAC2-OE calli was performed to identify the hormone signaling networks influenced by GbNAC2. Intriguingly, 12 DEGs were significantly enriched in the auxin signaling pathway, including five SAUR (SMALL AUXIN UP-REGULATED RNA) genes—a class of early auxin-responsive regulators implicated in cell expansion and root development[26]. Notably, all differentially expressed SAUR genes exhibited pronounced upregulation in transgenic calli, with qRT-PCR confirming the elevated expression of five representative SAUR paralogs (Supplementary Fig. S4).
ABA signaling has been widely documented to play a crucial role in regulating plant salt tolerance[27]. To explore ABA's role in ginkgo salt response, GbNAC2 transcription was analyzed in ABA-treated seedlings. The results showed that exogenous ABA significantly induced the GbNAC2 expression, with rapid upregulation within 1 h, and reaching a peak at 12 h (Fig. 7a). Ginkgo seedlings were then treated with 400 mM NaCl alone or in combination with 20 μM ABA. Severe wilting phenotypes in NaCl-treated plants were found whereas the ABA-treated plants maintained good growth status after salt treatment, with only minor leaf wilting (Fig. 7b). After 3 d of salt treatment, the seedlings co-treated with exogenous ABA and NaCl exhibited significantly lower levels of MDA and H2O2 compared to those treated with NaCl alone (Fig. 7c, d). Meanwhile, the expression level of GbNAC2 increased by more than 2-fold after salt treatment; under the co-treatment of ABA and salt stress, its expression level further increased by more than 4-fold (Fig. 7e). In the GbNAC2-OE ginkgo calli, seven genes related to the ABA signaling pathway were identified, among which four genes were up-regulated (Fig. 7f). Notably, the genes Gb_35096 and Gb_16239 exhibited positive responses to salt stress (Fig. 7g).
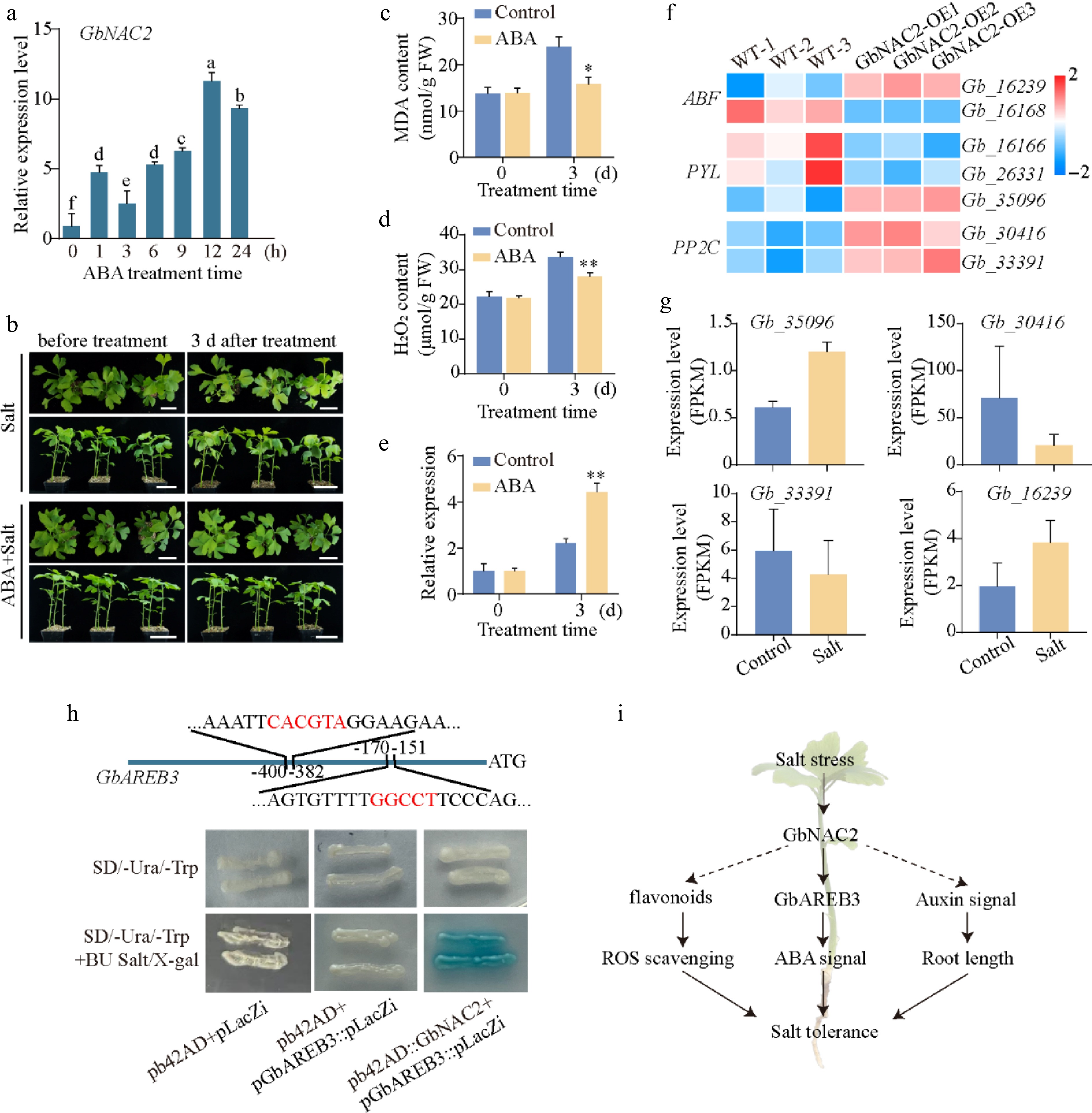
Figure 7.
GbNAC2 participates in ABA signaling by interacting with GbAREB3 promoters. (a) The expression profile of GbNAC2 within 24 h post ABA treatment. Different letters indicate significant differences (one-way analysis of variance, p < 0.05). (b) Phenotypic changes in ginkgo seedlings before and after NaCl and ABA + NaCl treatment. Scale bar = 4 cm. The content of (c) MDA, (d) and H2O2 before and after NaCl and ABA + NaCl treatment. (e) The expression of GbNAC2 before and after treatment. * p < 0.05, ** p < 0.01. (f) Heatmap of significantly differentially expressed ABA pathway-related genes following overexpression of GbNAC2. (g) Expression analysis of four ABA pathway-related genes under salt stress. (h) Verification of the interaction between GbNAC2 and the GbAREB3 promoter through Y1H assay. (i) Schematic diagram illustrating the promotion of salt stress tolerance in ginkgo by GbNAC2.
GbNAC2 activates GbAREB3 expression by directly binding to its promoter
-
Among those candidates, two NAC family protein-binding SNBE elements were identified on the GbAREB3 promoter, located at 151–170 and 382–400 bp, respectively. Through evolutionary analysis with Arabidopsis, Gb_16239 is most closely related to AtAREB3. Consequently, Gb_16239 has been designated as GbAREB3 (Supplementary Fig. S5). It is hypothesized that GbAREB3 is regulated upstream by NAC factors. To investigate the binding of GbNAC2 to the GbAREB3 promoter, the Y1H assay was utilized to confirm that GbNAC2 can directly bind to the GbAREB3 promoter. The results showed that yeast cells containing both GbNAC2 and pGbAREB3 exhibited normal growth and turned blue on a selective medium containing BU salt and X-gal. In contrast, yeast cells containing only pGbAREB3 were able to grow but did not turn blue on the same selective medium (Fig. 7h). These results suggest that GbNAC2 can directly bind to the GbAREB3 promoter to activate its expression.
-
G. biloba leaves are rich in flavonoids, which serve as essential raw materials for the pharmaceutical and nutraceutical industries[28]. Salt stress poses a significant challenge to plant growth, severely impacting ginkgo trees. However, the molecular mechanisms underlying ginkgo's tolerance to salt stress are still largely unexplored. NAC TFs are one of the largest gene families in plants and have been widely reported as pivotal in plant responses to salinity[29]. In this study, qRT-PCR results indicated that GbNAC2 responds rapidly to salt treatments with a significant increase in transcriptional levels, suggesting its potential role as a key regulator of ginkgo's salt stress tolerance. Notably, GbNAC2 is predominantly expressed in leaves and roots. To further investigate its function, GbNAC2 was overexpressed in both Arabidopsis and poplar. Compared to control plants, the transgenic plants exhibited significantly enhanced growth under salt stress, demonstrating improved salt tolerance. These data validate the vital role of GbNAC2 in enhancing ginkgo's salt stress resilience.
In the physiological regulatory network of plants adapting to the environment, the trade-off between growth and stress resistance is a universally existing core strategy. Due to the limited availability of resources such as carbon sources and energy in plants, the enhancement of resistance to external stresses is often accompanied by reduced growth[30]. This phenomenon has been repeatedly verified in studies on the regulation of plant stress resistance by NAC transcription factors. For instance, in Populus euphratica, PeNAC036-overexpressing plants exhibit significantly inhibited growth and enhanced salt tolerance[31]. In Populus euphratica, the overexpression of the PeNAC122 inhibits plant growth while significantly enhancing its resistance to osmotic stress[32]. These examples demonstrate the typical trade-off relationship of stress resistance enhancement coupled with growth inhibition. Nevertheless, some studies have found that certain NAC genes can break through this trade-off constraint. For example, overexpression of BpNAC2 from Betula platyphylla not only promotes plant growth but also enhances adaptability to salt stress[33]. PtNAC3 from Pinus tabuliformis not only enhances the resistance of transgenic plants to multiple abiotic stresses, but also promotes the seed yield of plants under stress conditions[34].
In this study, it was found that overexpression of GbNAC2 promoted the growth and salt resistance of transgenic plants. Meanwhile, the root length of transgenic plants was increased by more than 1.6-fold compared with the control. Transcriptome analysis revealed that the expression level of the auxin-responsive gene SAUR was upregulated by more than 2-fold in GbNAC2-overexpressing plants relative to the control. As downstream effectors of the auxin signal, SAUR family genes can promote the elongation of cells in the root elongation zone by activating plasma membrane H+-ATPase[35]. Considering that roots serve as the primary organ for plants to absorb water and perceive stress signals, the increase in root length can expand the water absorption range and reduce the accumulation of salt ions in the root epidermis[36]. Therefore, it is proposed that GbNAC2 may regulate the transcriptional expression of SAUR to initiate auxin-mediated root development, thereby enhancing salt stress tolerance. However, the precise regulatory mechanism between GbNAC2 and SAUR requires further clarification.
Salt stress triggers ionic and osmotic imbalances, which can lead to secondary stresses, particularly oxidative stress caused by ROS. Consequently, maintaining homeostatic levels of ROS through scavenging pathways is crucial for protecting plants from oxidative damage[37]. Plants predominantly utilize protective strategies to scavenge ROS: enzymatic and non-enzymatic antioxidants[38]. SOD and CAT are two crucial enzymes within the enzymatic detoxification system[39]. In this study, measurements of H2O2 content revealed that overexpression of GbNAC2 suppresses ROS accumulation induced by salt treatments. The levels of SOD and CAT were significantly higher in the GbNAC2-OE lines than in the WT plants. These findings indicate that GbNAC2 may regulate ginkgo salt tolerance through ROS scavenging.
In addition to the enzymatic metabolic system, numerous metabolites function as non-enzymatic antioxidants[40]. Flavonoids are particularly important in this context and have been shown to possess antioxidant properties[41]. Extensive research has highlighted the significant role of flavonoids in mitigating the adverse impacts of salt stress by increasing the levels of antioxidant enzymes and reducing ROS accumulation[42,43]. Gao et al.[44] found that salt stress induces the expression of EbbHLH80, and overexpression of EbbHLH80 enhances salt tolerance, accompanied by elevated flavonoid accumulation and reduced ROS levels compared to WT plants. Moreover, the IAA17.1/HSFA5a module modulates poplar root adaptation to salt stress by regulating flavonol biosynthesis and controlling ROS accumulation[45]. The present findings demonstrated that overexpression of GbNAC2 facilitates the accumulation of flavonoids and activates genes associated with flavonoid biosynthesis, including CHS, F3H, C4H, and DFR. Notably, GbNAC2-OE plants exhibited an increasing trend in flavonoid accumulation after salt treatments. These results suggest that GbNAC2 may enhance ginkgo's salt tolerance by regulating flavonoid biosynthesis to maintain ROS homeostasis. However, whether GbNAC2 directly regulates these flavonoid biosynthesis genes requires further investigation.
Plant hormones play a critical role in regulating plant salt tolerance, with ABA being a key hormone in modulating the plant's response to salt stress. Under high salinity, ABA mediates stomatal closure, thereby preventing water loss due to osmotic stress[8,46]. Exogenous ABA treatment has been shown to enhance the salt tolerance of rice seedlings[47], while the rice SAE1 gene influences salt stress tolerance by inhibiting ABI5-mediated ABA signaling[48]. Conversely, PalWRKY77 in poplar mitigates ABA-mediated growth inhibition and stomatal closure by suppressing the expression of ABA-responsive genes, thereby reducing salt tolerance[49]. In this study, after three days of salt treatment, ginkgo seedlings exhibited a wilting phenotype, and concurrently, the expression level of GbNAC2 was significantly upregulated. Furthermore, it was observed that the application of exogenous ABA could significantly alleviate the damage caused by high salt stress to ginkgo seedlings, with the expression level of GbNAC2 increasing by more than 4-fold. These results indicate that elevated GbNAC2 expression contributes to alleviating the damage caused by salt stress in ginkgo. A key GbAREB3 gene was also identified in the ABA signaling pathway downstream of GbNAC2. GbAREB3 belongs to the ABF family of transcription factors, which are widely reported to be involved in the plant stress response[50]. It was found that the GbAREB3 transcription level is regulated by salt stress, and overexpression of GbNAC2 promotes the GbAREB3 expression. Y1H validation demonstrated that GbNAC2 can directly bind to the GbAREB3 promoter to regulate its expression. Therefore, GbNAC2 enhances ginkgo's salt stress tolerance by regulating GbAREB3 expression to activate the ABA signaling pathway.
-
In conclusion, the present study elucidates the dual role of GbNAC2 in enhancing salt tolerance in ginkgo through the integrated regulation of flavonoid biosynthesis and auxin-ABA crosstalk (Fig. 7i). As a central transcriptional regulator, GbNAC2 promotes stress-resilient root architecture remodeling and flavonoid-driven antioxidant capacity by activating auxin signaling and flavonoid metabolic genes. Simultaneously, it amplifies ABA-mediated detoxification through direct binding to the GbAREB3 promoter. This unique coordination of developmental plasticity (via auxin), and stress-responsive metabolism (via flavonoids/ABA signaling) effectively resolves the growth-defense trade-off under salinity stress. Together, these results demonstrate that GbNAC2 serves as a key salt-tolerance gene in ginkgo, contributing to salinity adaptation through multiple avenues. Its multifunctional role makes GbNAC2 a promising target for biotechnological strategies aimed at improving salt tolerance in ginkgo through molecular breeding.
This project was supported by the Jiangsu Provincial Key Research and Development Program (modern agriculture) (Grant No. BE2021367) and the National Natural Science Foundation of China (Grant Nos 32501718 and 32171838).
-
The authors confirm their contributions to the paper as follows: study conception and design: Wang L, Wang Q, Lu J, Tang H, Li W; data collection: Lu J, Tang H, Jiang Y, Zou H; analysis and interpretation of results: Lu J, Tang H, Wang Q; draft manuscript preparation: Lu J, Wang L, Wang Z. All authors reviewed the results and approved the final version of the manuscript.
-
The raw data of the transcriptome in this study are stored in the National Genomics Data Center's Genome Sequence Archive, with Accession No. CRA016156.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jinkai Lu, Han Tang
- Supplementary Table S1 Names and corresponding accession numbers (IDs) of genes used in this work.
- Supplementary Table S2 Oligonucleotide sequences of the primers used in this work.
- Supplementary Fig. S1 Phylogenetic relationships of G. biloba NAC2 and NAC family proteins in Arabidopsis. Accession numbers are listed in Table S1.
- Supplementary Fig. S2 The Effect of GbNAC2 overexpression on seed germination in transgenic Arabidopsis.
- Supplementary Fig. S3 DEGs analysis in GbNAC2 overexpression calli and WT calli.
- Supplementary Fig. S4 Analysis of genes related to Auxin pathway.
- Supplementary Fig. S5 Phylogenetic relationships of G. biloba AREB3 and ABF family proteins in Arabidopsis.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lu J, Tang H, Li W, Jiang Y, Zou H, et al. 2025. Dual function of GbNAC2 in flavonoid metabolism and hormonal pathways enhances salt tolerance in Ginkgo biloba. Forestry Research 5: e028 doi: 10.48130/forres-0025-0027 

Dual function of GbNAC2 in flavonoid metabolism and hormonal pathways enhances salt tolerance in Ginkgo biloba
- Received: 15 May 2025
- Revised: 28 September 2025
- Accepted: 23 October 2025
- Published online: 20 November 2025
Abstract: NAC transcription factors are central regulators of plant salt tolerance, yet their specific roles in ginkgo salt response remain unclear. Here, GbNAC2 was identified as a salinity-induced transcriptional activator in ginkgo, orchestrating two key adaptive responses. GbNAC2 overexpression significantly improved salt tolerance in transgenic plants, accompanied by over 60% increase in root length, and more than 20% increase in flavonoid content compared to wild type (WT). Transcriptome analysis of GbNAC2-overexpressing ginkgo calli revealed that genes related to auxin biosynthesis, and those involved in the flavonoid synthesis pathway, were significantly upregulated in transgenic calli. Mechanistically, GbNAC2 directly binds the GbAREB3 promoter to enhance ABA signaling, and exogenous ABA treatment further enhances salt resilience. The present findings unveil a unique crosstalk mediated by GbNAC2 between flavonoid-antioxidant systems and auxin-ABA hormonal networks, effectively resolving the growth-defense trade-off under salinity in ginkgo.
-
Key words:
- Ginkgo biloba /
- Salt stress /
- NAC transcription factor /
- Flavonoid /
- ABA