










-
As major coniferous timber species, larches (Larix spp.) hold substantial economic and ecological value due to their rapid growth and adaptability. However, their extended vegetative phase poses severe constraints for traditional breeding programs[1,2]. The plant life cycle progresses through distinct phases: dry seed, seed germination, juvenile and adult vegetative phase, followed by the reproductive phase, embryogenesis, seed maturation, and eventually seed set[3]. Thus, accelerating the life-cycle progression, especially the transition from the vegetative to the reproductive phases, is essential for improving genetic selection and promoting sustainable forestry.
In short-lived angiosperms such as Arabidopsis thaliana, phase transitions are precisely controlled by gene networks that integrate environmental and endogenous signals. Key regulators include the microRNA156 (miR156)-SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) and miRNA172-APETALA2 (AP2) modules[4,5]. miR172 represses AP2-like transcription factors to promote the vegetative phase transition, and reproductive phase transition, while stage-specific hormones further refine these processes[6,7]. While these pathways are well-studied in annual plants, their roles in conifers, which have long life cycles and distinct evolutionary histories, remain poorly understood. Understanding these mechanisms is significant for studying the genetic control of the development timing of coniferous trees.
The AP2/Ethylene Responsive Factor (AP2/ERF) transcription factor superfamily plays key roles in plant growth, phase transition, and stress responses[8−10]. This superfamily is classified into five major subfamilies—AP2, dehydration response element-binding protein (DREB), ERF, related to ABA-insensitive3/viviparous1 (RAV), and Soloist—based on the number and structure of their AP2 domains[10]. The A. thaliana AP2 subfamily includes AP2 and five AP2-like genes: SCHLAFMUTZE (SMZ), SCHNARCHZAPFEN (SNZ), TARGET OF EAT1 (TOE1), TOE2, and TOE3, all of which contain two AP2 domains[11]. These genes are post-transcriptionally regulated by miR172 and function as repressors of the vegetative phase, and reproductive phase transition[5,12−15]. The AP2 subfamily also includes AINTEGUMENTA (ANT) and PLETHORA (PLT), which are essential for floral organ identity and root growth, respectively[16−18]. AP2 also regulates other life-cycle events, such as embryo development and seed germination[19−21]. The DREB and ERF subfamilies contain genes with a single AP2 domain. DREB genes play critical roles in abiotic stress responses, such as drought, cold, and salinity, while ERF genes regulate hormone signaling, pathogen defense, and abiotic stress tolerance[10]. The RAV subfamily, which contains both an AP2 domain and a B3 domain, influences the reproductive phase transition as well as biotic and abiotic stress responses[22]. The Soloist subfamily genes also contain a single AP2 domain, but their sequences and gene structures strongly diverge from those of the DREB and ERF subfamilies[23].
In gymnosperms, functional studies of AP2 superfamily genes remain limited. Recently, Ginkgo biloba TOE1 is reported to enhance salt/drought tolerance, but its role in development is unknown[24]. In G. biloba and Gnetum gnemon, some AP2 subfamily genes are broadly expressed during the early stages of ovule development and later are specifically expressed in the nucellus and integument, suggesting their involvement in embryogenesis and seed formation[25]. Pinus thunbergii AP2-like 1/2 (PtAP2L1/2) are found to be expressed throughout cone development[26], while PaAP2L2 in Picea abies shows broad vegetative expression but remains functionally uncharacterized[27]. In Larix kaempferi (Japanese larch), AP2/ERF superfamily members such as LkAP2L2 and LkERF6 are found to be involved in branch formation, seed development, and stress responses[28,29]. Nevertheless, the functional roles of AP2/ERF superfamily genes in most gymnosperms—particularly euAP2 lineage homologs—in regulating key life-cycle progression, remain largely unexplored. This gap limits our understanding of the distinctive longevity and reproductive strategies in gymnosperms. Some gymnosperm AP2/ERF superfamily genes contain a miR172 target site[30], but their function in phase transition is unclear. Recent progress in conifer genomics[31−35], including the sequencing of the L. kaempferi genome, now enables the use of molecular approaches to study phase transitions, and identify targets for genetic improvement through genome-wide studies of L. kaempferi AP2/ERF superfamily genes.
Here, genome-wide identification of the AP2/ERF superfamily genes in L. kaempferi were integrated with spatiotemporal expression profiling across several life-cycle stages, including seed germination, seedling development, vegetative and reproductive growth, embryogenesis, and seed maturation, to investigate their roles in life-cycle progression. Using RNA-seq, qRT-PCR, the RUBY reporter system, and transgenic Arabidopsis, a developmental stage specifically expressed AP2 gene was characterized. The present findings will not only reveal an age-decreased euAP2 lineage, but also provide a candidate gene for promoting seed formation and germination in larch breeding.
-
The genome files of L. kaempferi were obtained from the Larix Genome Database (LarixGD,
www.Larixgd.cn/#/map )[34]. The Hidden Markov model (HMM) profiles of the AP2 domain (PF00847), and B3 domain (PF02362) were retrieved from the PFAM database (http://pfam.xfam.org ). Genes containing these domains were scanned in the L. kaempferi genome using TBtools-HMMER with an E-value threshold of 10−5[36].To confirm these candidate genes, their predicted protein sequences were analyzed for conserved structural domains in the NCBI CDD database (
www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ), with an E-value cutoff of < 10−5. Genes were classified into subfamilies (AP2, DREB, ERF, RAV, and Soloist), based on domain type and number[23].Protein physicochemical properties and subcellular localization prediction
-
The physicochemical properties, including aliphatic index, grand average of hydropathicity, instability index, molecular weight, and isoelectric point, were analyzed using TBtools software[36]. The potential subcellular localizations of these AP2/ERF superfamily proteins were predicted using WoLF PSORT (
https://wolfpsort.hgc.jp ).Multiple sequence alignment and phylogenetic analysis
-
To identify the Soloist subfamily genes, T-coffee scores of genes containing a single AP2 domain were obtained through multiple sequence alignment using MEGA12.0.9 software[37]. The genes with low scores were classified into the Soloist subfamily.
Atypical ERF and DREB genes in L. kaempferi were classified based on their evolutionary relationships. The protein sequences of typical ERF and DREB genes from A. thaliana were obtained from the A. thaliana information resource TAIR (
www.arabidopsis.org/ ). Firstly, protein sequences of A. thaliana and L. kaempferi genes were aligned and trimmed using trimAI software[38]. Next, these sequences were used to construct phylogenetic trees in MEGA12.0.9 software[37] using the maximum likelihood method (1,000 bootstraps), and the results were visualized in Evolview (https://evolgenius.info//evolview-v2 ).For phylogenetic analysis of AP2 subfamily genes in A. thaliana and L. kaempferi, a maximum likelihood tree was constructed with the same method as above. To predict miR172 target sites, the coding sequences of AP2 subfamily genes were aligned with A. thaliana AP2, and the results were visualized with JalView software[39].
Analysis of the expression patterns of AP2 subfamily genes using the age-related transcriptomes
-
To investigate the expression patterns of AP2 subfamily genes during L. kaempferi tree aging, transcriptome data were obtained from the NCBI sequence read archive under BioProject accession: PRJNA234461. Branches from the upper crowns collected from dormant trees at ages 1, 4, 8, 12, 20, and 50 years[40], and the uppermost main stems collected from active trees at ages 1, 2, 5, 10, 25, and 50 years were used for transcriptome sequencing[41]. After the removal of buds or needles, the left stems from at least three trees from each category were pooled and used for RNA extraction. These clean reads were mapped to the L. kaempferi genome[34], and gene expression levels were calculated as transcripts per million (TPM). Heatmaps were generated in TBtools software, using the normalized TPM (log2) values.
Plant materials used for qRT-PCR analysis
Experiment I: expression patterns of LkTOE1-2 during L. kaempferi tree aging
-
Samples from 0.58-, 2.58-, 4.5-, 5.58-, 8.58-, 10.50-, and 12.50-year-old dormant L. kaempferi trees were collected on 11 November 2020, while samples from 1.25-, 3.17-, 5.17-, 7.25-, 9.17-, 11.17-, and 13.17-year-old active L. kaempferi trees were collected on 4 July 2019. These samples were collected in our previous study[42]. The exact growth time was calculated based on afforestation records from the Dagujia seed orchard (42°22' N, 124°51' E), Liaoning Province, Northeast China. For 0.58-year-old trees, the main stem was sampled, while for all other age groups, lateral branches from the uppermost main stem, formed during the current year, were used. Buds and needles were removed before pooling samples from at least three trees per age group. Samples were then frozen in liquid nitrogen and stored at –80 °C until RNA extraction.
Experiment II and III: expression patterns of LkTOE1-2 in L. kaempferi seeds and seedlings
-
Mature seeds were collected from 38-year-old trees on 22 August 2020. Endosperm and embryo samples were taken from ten mature seeds after two days of water soaking. Ten 22-day-old seedlings were sampled on 13 May 2021, while ten 54-day-old, and six 74-day-old seedlings were sampled on 14 June and 4 July 2021, respectively. To analyze the expression pattern of LkTOE1-2 in different organs, roots, hypocotyls, stems, and needles were collected from 15 74-day-old seedlings. All seedlings were sown on 22 April 2021. Before sowing, seeds were soaked in water until the embryonic root emerged through the seed coat. These samples were collected in our previous study[42].
Experiment IV: expression patterns of LkTOE1-2 during L. kaempferi seed formation
-
Pollen grains were collected from 20 microsporophylls of 13-year-old trees on 13 April 2020. Immature embryos, endosperms, and seed coats were collected from ten immature seed cones on 30 June and 27 July 2022. Mature embryos, endosperms, and seed coats were collected from ten mature seed cones on 22 August 2020. These samples were collected in our previous study[42].
Experiment V: expression patterns of LkTOE1-2 during L. kaempferi somatic embryogenesis
-
Callus was induced from immature embryos of L. kaempferi with the same medium as described in a previous study[43]. Other mediums used in somatic embryogenesis were according to the method of Song et al.[44]. Callus proliferation was maintained on proliferation medium (BM basal medium supplemented with 0.05 mg/L 6-BA, 0.05 mg/L KT, 0.50 mg/L NAA, 1 g/L glutamine, 0.50 g/L acid hydrolyzed casein, 25 g/L sucrose, 4 g/L agar, pH 5.75–5.85), under dark conditions, with subculturing every two weeks. Somatic embryos were induced from callus on maturation medium (BM basal medium with 15.86 mg/L ABA, 0.20 mg/L IBA, 10 g/L activated carbon, 68.40 g/L sucrose, 4 g/L agar, pH 5.75–5.85) for six weeks in the dark. Somatic seedlings were obtained by putting somatic embryos on germination medium (WPM basal medium containing 3 mg/L VB1, 2 g/L activated carbon, 20 g/L sucrose, 4 g/L agar, pH 5.75–5.85). The germination process consisted of a two-week dark culture followed by a one-month light culture.
Experiment VI: response of LkTOE1-2 to temperature treatment and water soaking
-
Seed cones were collected on 22 August 2020 and kept at room temperature for seed extraction. Group 1 was collected on 18 September 2020 from seeds that had not undergone 4 °C treatment, but were exposed to room temperature and water soaking. Groups 2–5 were collected from seeds stored at 4 °C for over a year. Group 2 was collected from seeds with no room temperature exposure or water soaking. Group 3 was collected from seeds with water soaking for two days without room temperature exposure. Group 4 was collected from seeds kept at room temperature for two weeks without water soaking. Group 5 was collected from seeds kept at room temperature for two weeks and with water soaking for two days. During water soaking, seeds were maintained at room temperature. For sampling, seed coats were removed from ten seeds and the remainder were collected.
RNA extraction and qRT-PCR
-
Total RNA was extracted using the EasyPure RNA Kit (TransGen Biotech, ER101-01, Beijing, China) according to the manufacturer's instructions. A 2.5 µg aliquot of total RNA was reverse-transcribed into cDNA using the TransScript II One-step gDNA Removal and cDNA Synthesis SuperMix Kit (TransGen Biotech, Beijing, China), and then diluted for sequence cloning and qRT-PCR.
LkTOE1-2 (also known as LaAP2-1, GenBank accession number: MN790757) was cloned in our previous work[45]. The primers 5'-GCGGACCAACAACTCCAGTA-3' and 5'-GTTGCCATATGCAAGCTCGG-3' were designed to amplify LkTOE1-2, 5'-GACTGTACCGTTGGTCGTG-3' and 5'-CCTCCAGCAGAGCTTCAT-3' for L. kaempferi translation elongation factor-1 alpha 1 (LkEF1A1, GenBank Accession No.: JX157845), 5'-TTCGGCTTTGAAGGAGGGTC-3' and 5'-TTGGACGCAGTCCCCATAAG-3' for L. kaempferi zinc finger protein (LkZFP, GenBank Accession No.: MZ965074), 5'-TGGCGTCCAAAAGGATTCTCA-3' and 5'-TCCCATGATTGTAGCTTGCCA-3' for L. kaempferi ubiquitin-conjugating enzyme E2 28 (LaUBC1, GenBank Accession No.: ON887160)[42]. LkEF1A1 was used in all experiment, LkZFP was used in experiments I, and LkUBC1 was used in experiments II–IV, and VI (Supplementary Table S1). △Ct value (CtLkEF1A − CtLkTOE1-2) was used for relative quantification analysis, where Ct represents the threshold cycle. Transcript levels shown in all figures are presented based on these values. QRT-PCR was performed with three or four technical replicates, and data were shown as mean ± SD. Statistical significance was assessed by one-way ANOVA followed by Tukey's multiple comparisons test.
Expression patterns of LkTOE1-2 during seed formation detected by RNA-seq
-
Immature embryos, endosperms, and seed coats were collected from ten immature seed cones on 9 July 2024. Mature seeds were collected from ten mature seed cones on 22 August 2020. All samples were frozen in liquid nitrogen, and then stored at −80 °C for RNA isolation. Transcriptome sequencing was performed on the Illumina HiSeqTM 2500 platform (Illumina, CA, USA) as described by Xiang et al.[46]. The raw reads have been uploaded to the China National Center for Bioinformation under Accession No.: PRJCA030694. Gene expression levels were calculated as above, and presented as means, with error bars representing the standard deviations (n = 3).
Promoter cloning
-
The genomic DNA from L. kaempferi seedlings was extracted with the Plant Genomic DNA Kit (Tiangen Biotech, DP305-03, Beijing, China), according to the manufacturer's protocol. The sequences 2,000 bp upstream of LkTOE1-2 ATG initiation codon were amplified using primers 5'-GTTAGCTCCGCGTTAGTTTTGAC-3', and 5'-CTTCGCTGCCGGAAGGCCAAGG-3'. PCR products were purified using a gel extraction kit (Tiangen, DP209-03, Beijing, China), and subsequently sequenced.
Construction of proLkTOE1-2::RUBY vector and L. kaempferi transformation
-
The proDR5::RUBY vector was provided by Yubing He from the Institute of Crop Sciences, Chinese Academy of Agricultural Sciences. The promoter sequences of LkTOE1-2 (proLkTOE1-2), and CaMV35S were separately subcloned into proDR5::RUBY vector via the HindIII cleavage site, replacing the DR5 promoter[47]. This resulted in the proLkTOE1-2::RUBY and pro35S::RUBY vectors, respectively, with proMV35S::RUBY serving as a control. The primers 5'-gactgaccacccggggatccGTTAGCTCCGCGTTAGTTTTGAC-3', and 5'-gcgagggtcgcatgatccatCTTCGCTGCCGGAAGGCCAAGG-3' were used for proLkTOE1-2. The primers 5'-gactgaccacccggggatccTGAGACTTTTCAACAAAGGG-3' and 5'-gcgctgaagcttggctgcagTGTTCTCTCCAAATGAAATG-3' were used for pro35S. The recombinant vectors were first introduced into Agrobacterium tumefaciens strain GV3101, and subsequently transferred into L. kaempferi callus using a previously described method[43].
Construction of pro35S::LkTOE1-2 vector and A. thaliana transformation
-
The coding sequence of LkTOE1-2 was subcloned into the plant expression vector pBI121 via the BamHI cleavage site, and it was downstream of the 35S promoter, resulting in the pro35S::LkTOE1-2 vector. The primers 5'-tctagaaagcttCTGCAGATGATGACCAGAGATACTTCTCG-3', and 5'-ggtaccggatccACTAGTCGTGTCCCTTTGGACGCTTG-3' were used. The empty vector was used as a control. The recombinant vector was first introduced into A. tumefaciens strain GV3101, and subsequently transferred into A. thaliana ecotype Col-0 with the floral-dip method[48]. T1 transformants were selected on 1/2 Murashige and Skoog (MS) culture medium supplemented with 50 mg/L hygromycin. T2 transgenic plants were validated by PCR, RT-PCR, and qRT-PCR before phenotypic analysis (Supplementary Fig. S1). Plant growth conditions and statistical standards were the same as reported in our previous study[49].
-
In total, 374 genes were identified in the L. kaempferi genome, each containing at least one AP2 domain (Supplementary Table S2, Supplemental Figs S2, S3). These genes were assigned to five subfamilies, including eight AP2, 163 DREB, 158 ERF, 12 RAV, one Soloist, and 32 other subfamily genes (Table 1). Compared with A. thaliana (224 members), and Oryza sativa (178 members), the AP2 subfamily in L. kaempferi is seven times fewer than in A. thaliana (56), and four times fewer than in O. sativa (32) (Table 1). The DREB subfamily in L. kaempferi expands significantly, because L. kaempferi has more members (163) than A. thaliana (eight), and O. sativa (five) (Table 1). This expansion may be linked to the complex genome evolution of conifers, including frequent whole-genome duplication events and adaptations to abiotic stresses such as cold, drought, and salinity[31,33].
Table 1. Summary of the AP2/ERF superfamily in Arabidopsis thaliana, Oryza sativa and Larix kaempferi.
Classification Arabidopsis thaliana Oryza sativa Larix kaempferi AP2 subfamily 56 32 8 DREB subfamily 8 5 163 ERF subfamily 134 127 158 Other subfamily no no 32 RAV subfamily 26 14 12 Soloist subfamily Uncharacterized Uncharacterized 1 Total 224 178 374 The data for A. thaliana and O. sativa AP2/ERF superfamily was from Choudhury[63]. Among the eight AP2 subfamily members, LkAP2 and LkTOE1-1/2/3/4 belonged to the euAP2 lineage, and LkAIL1, LkSMOS1 and LkWRI1 belonged to ANT lineage (Fig. 1a). The miR172 complementary sequences were present in LkTOE1-1/2/3/4 (Fig. 1b), suggesting their potential post-transcriptional regulation by miR172. LkWRI1 and LkAP2 showed increased expression with age in the active stage, whereas LkTOE1-2 and LkTOE1-3 displayed high expression in one-year-old seedlings followed by rapid decline in both the active and dormant stages (Fig. 1b). The 'early high, later silent' pattern of LkTOE1-2/3 is similar to TOE homologs in Arabidopsis[12], maize[50], and Cardamine flexuosa[51], Pinus elliottii × P. caribaea[52] and other species[53−55], suggesting a conserved regulatory role during early seedling development. Given that LkTOE1-3 (also known as LkAP2L2) has been functionally characterized previously[28], LkTOE1-2 was selected for further investigation.
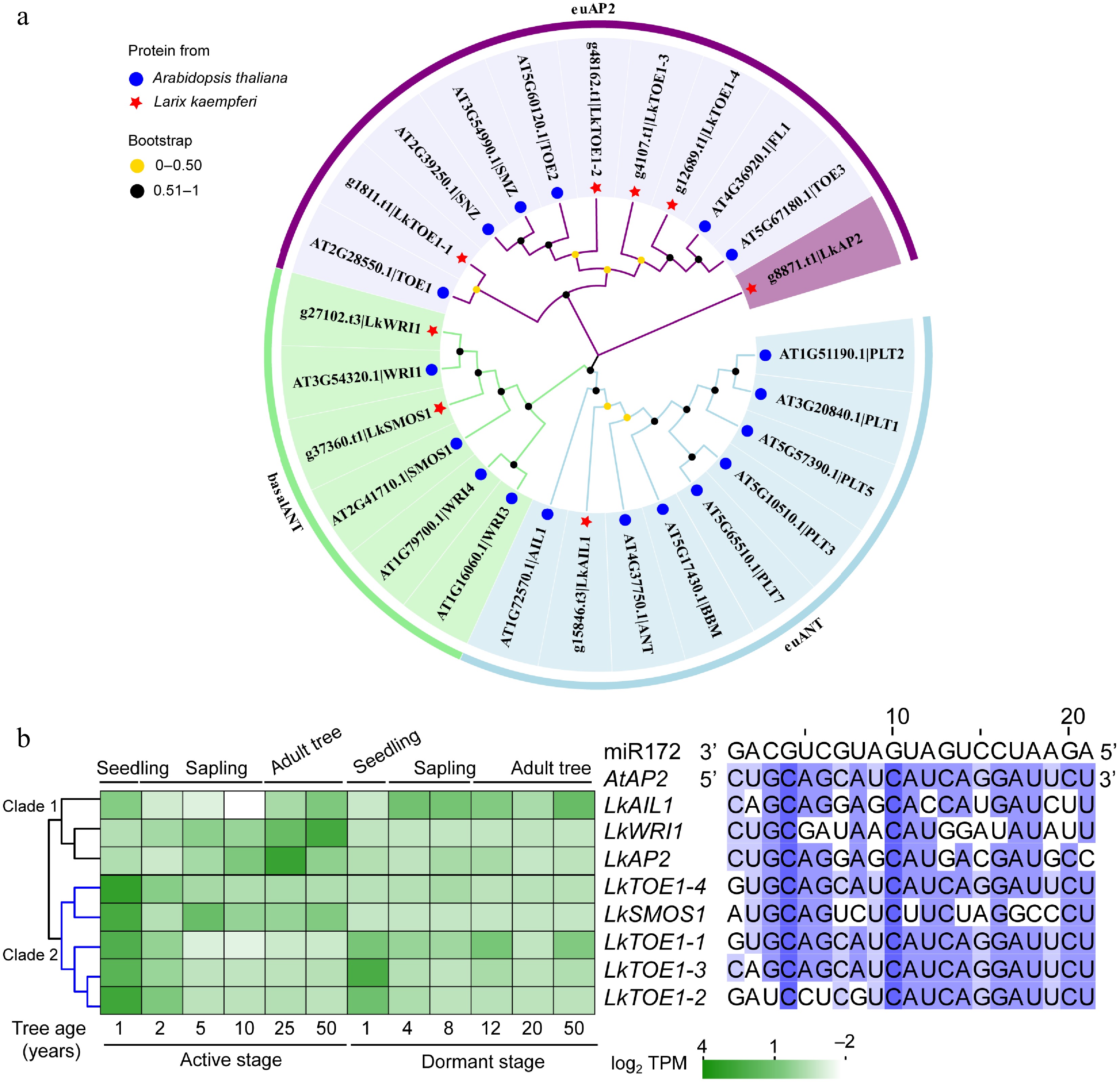
Figure 1.
Phylogeny, sequence, and expression analysis of AP2 subfamily genes in Larix kaempferi. (a) Phylogenetic tree of AP2 subfamily proteins from L. kaempferi and Arabidopsis thaliana. (b) Sequence alignment of conserved miR172 target sites in A. thaliana and L. kaempferi AP2 with A. thaliana miR172 as a reference and expression patterns of AP2 subfamily genes during L. kaempferi tree aging detected by RNA-seq. Heatmaps were shown with the average normalized TPM (log2) values (–2 to 4).
LkTOE1-2 transcription is regulated during seed formation and germination
-
LkTOE1-2 expression level was high in the active and dormant seedlings (Fig. 2a), indicating that it plays roles in early seedling development. Compared with seeds, LkTOE1-2 expression level was elevated in seedlings, particularly in stems and needles (Fig. 2b, c). During seed development from June to August, LkTOE1-2 was strongly expressed in embryos but weakly expressed in endosperms and seed coats (Fig. 2d, e). These findings suggested that LkTOE1-2 transcription is induced during germination and is regulated during seed formation. These similar expression patterns of LkTOE1-2 homologs in zygotic embryogenesis are also found in G. biloba and G. gnemon[25], suggesting that these AP2 subfamily genes function in zygotic embryogenesis.
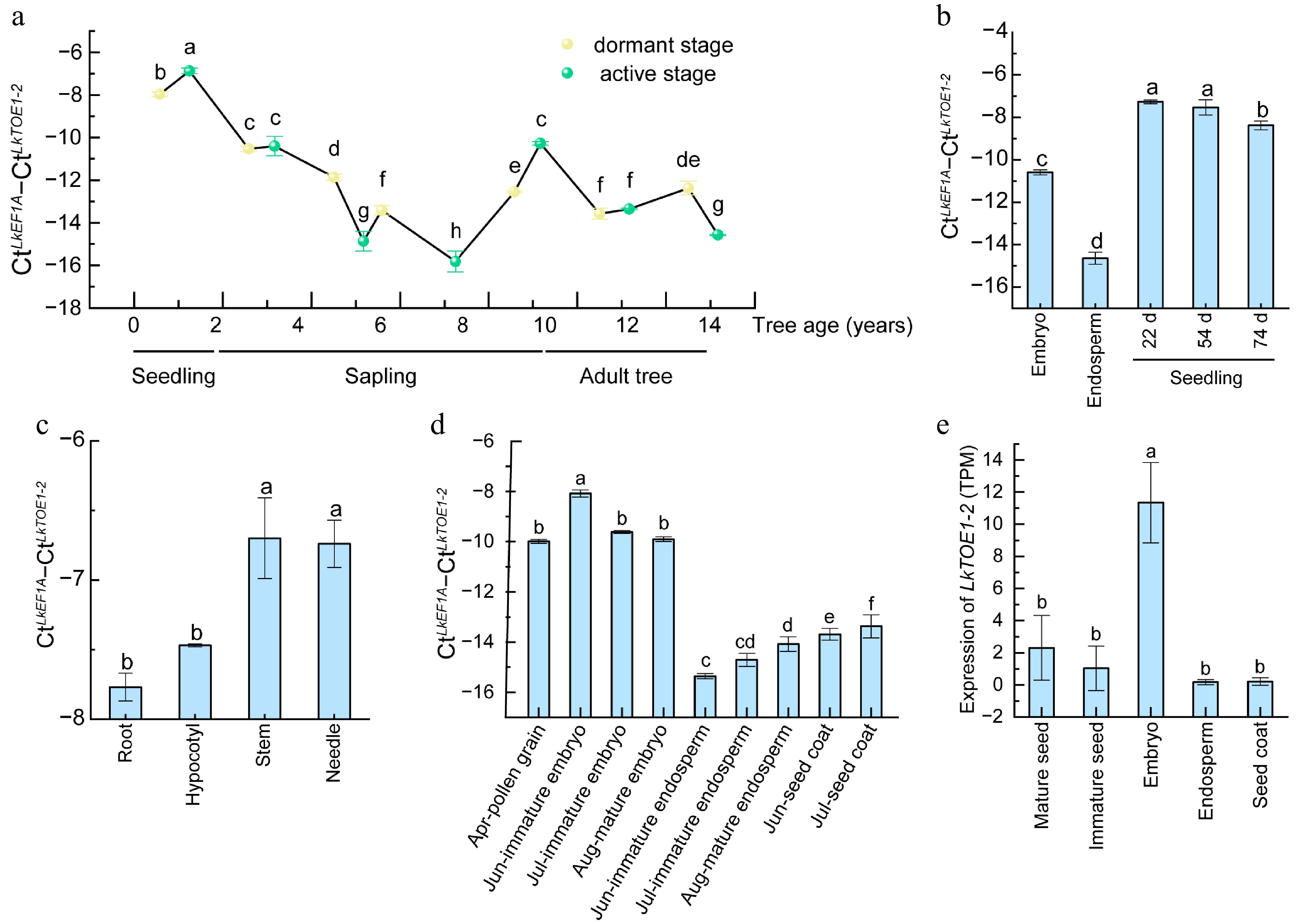
Figure 2.
Expression patterns of LkTOE1-2 during tree aging and seed formation in Larix kaempferi. (a) Expression patterns of LkTOE1-2 during L. kaempferi tree aging detected by qRT-PCR. (b) Expression patterns of LkTOE1-2 in seeds and seedlings. (c) Expression patterns of LkTOE1-2 in different organs of seedlings. Expression patterns of LkTOE1-2 during L. kaempferi seed formation detected by (d) qRT-PCR, and (e) RNA-seq. Different letters on the bars indicate significant differences as determined by one-way ANOVA followed by Tukey's multiple comparisons test, p < 0.05.
LkTOE1-2 transcription was induced during L. kaempferi somatic embryo formation and germination
-
During somatic embryogenesis, LkTOE1-2 expression level was 1.54-fold higher in somatic embryos and 4.73-fold higher in somatic seedlings compared with embryogenic calli (Fig. 3a), showing that LkTOE1-2 transcription is induced during L. kaempferi somatic embryo formation and germination. Using the RUBY reporter system[47], enhanced promoter activity of LkTOE1-2 was observed during embryo maturation. Wild-type calli appeared white, calli expressing pro35S::RUBY were red, and calli expressing proTOE1-2::RUBY were also white (Fig. 3b). After a maturation culture, light yellow somatic embryos formed from white wild-type calli, and red somatic embryos formed from red transgenic calli expressing pro35S::RUBY and white calli expressing proTOE1-2::RUBY (Fig. 3b). These visible observations showed that LkTOE1-2 transcription is induced during somatic embryo formation. Unfortunately, these red transgenic somatic embryos did not develop into somatic seedlings. L. × marschlinsii AP2L1 and L. decidua BABYBOOM (BBM) also have this similar expression pattern during somatic embryogenesis[56,57], suggesting that these AP2 subfamily genes function in somatic embryogenesis. Altogether, AP2 subfamily genes function in both zygotic and somatic embryogenesis in gymnosperms, and are similar to Arabidopsis BBM[58−60].
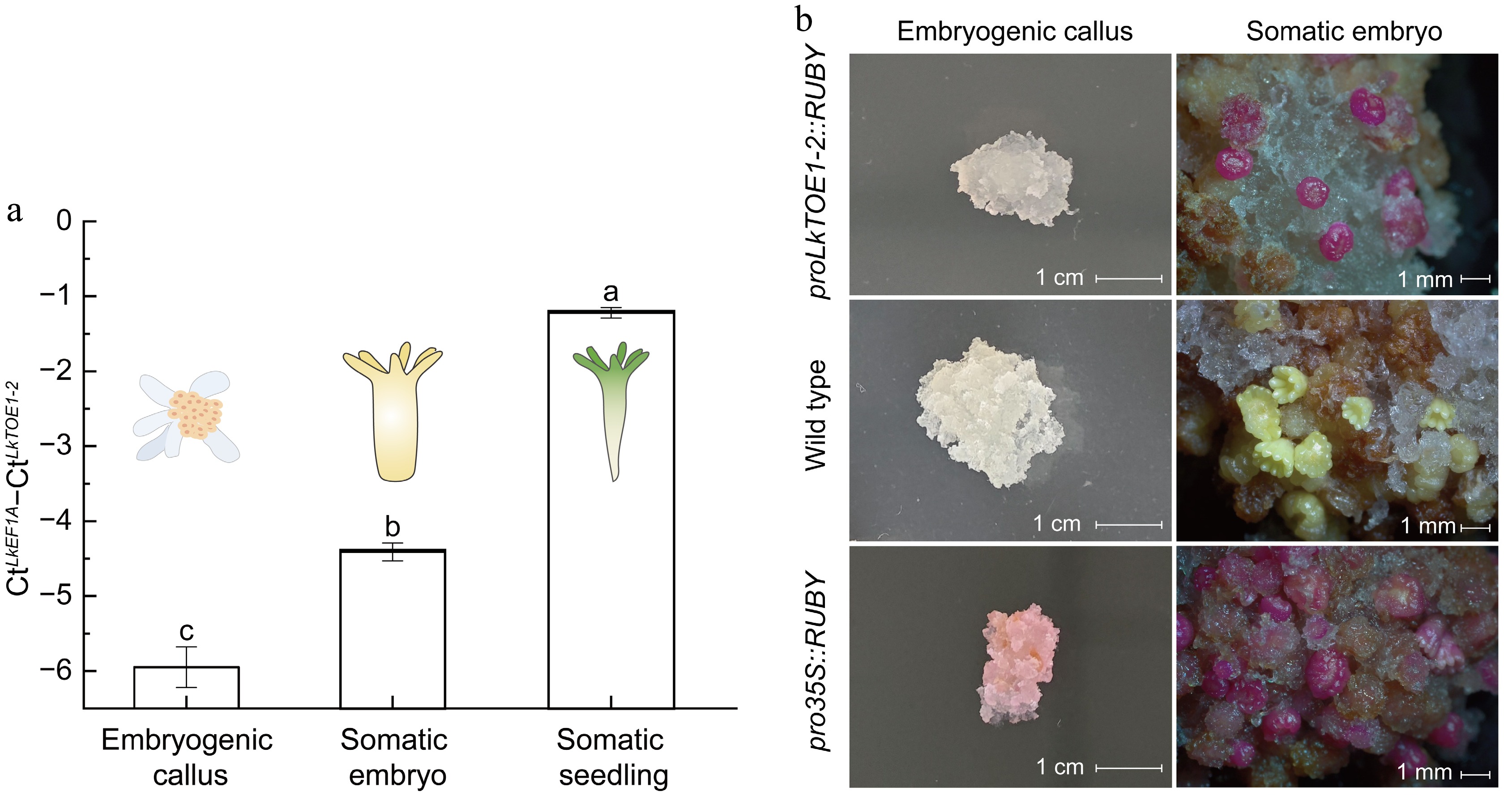
Figure 3.
Expression patterns and promoter activity of LkTOE1-2 during Larix kaempferi somatic embryogenesis. (a) Expression patterns of LkTOE1-2 in embryogenetic calli, somatic embryos, and somatic seedlings detected by qRT-PCR. Different letters on the bars indicate significant differences as determined by one-way ANOVA followed by Tukey's multiple comparisons test, p < 0.05. (b) Promoter activity of LkTOE1-2 in embryogenetic calli and somatic embryos detected with the RUBY reporter system.
LkTOE1-2 transcription was induced by low temperature and following water soaking
-
To investigate the regulatory mechanism of LkTOE1-2 during seed germination, the impacts of two key factors essential for seed germination—low temperature and water soaking—on LkTOE1-2 expression were examined. LkTOE1-2 expression level was the lowest in group 1 that had not undergone 4 °C treatment (Fig. 4). It was high in groups 3 and 5 that had been treated with 4 °C and water soaking, and were low in groups 2 and 4 that had been treated with 4 °C, but without water soaking (Fig. 4). Compared with group 1, it was high in group 5, indicating that 4 °C treatment increased LkTOE1-2 expression. Compared with group 2, it was high in group 3, and compared with group 4, it was high in group 5, indicating that water soaking increased LkTOE1-2 expression (Fig. 4). Notably, compared with group 2 and 3, it did not increase in group 4 and 5, respectively, indicating that without water soaking, only room temperature treatment could not increase LkTOE1-2 expression (Fig. 4). These data further showed that low temperature and the following water soaking induce LkTOE1-2 transcription.
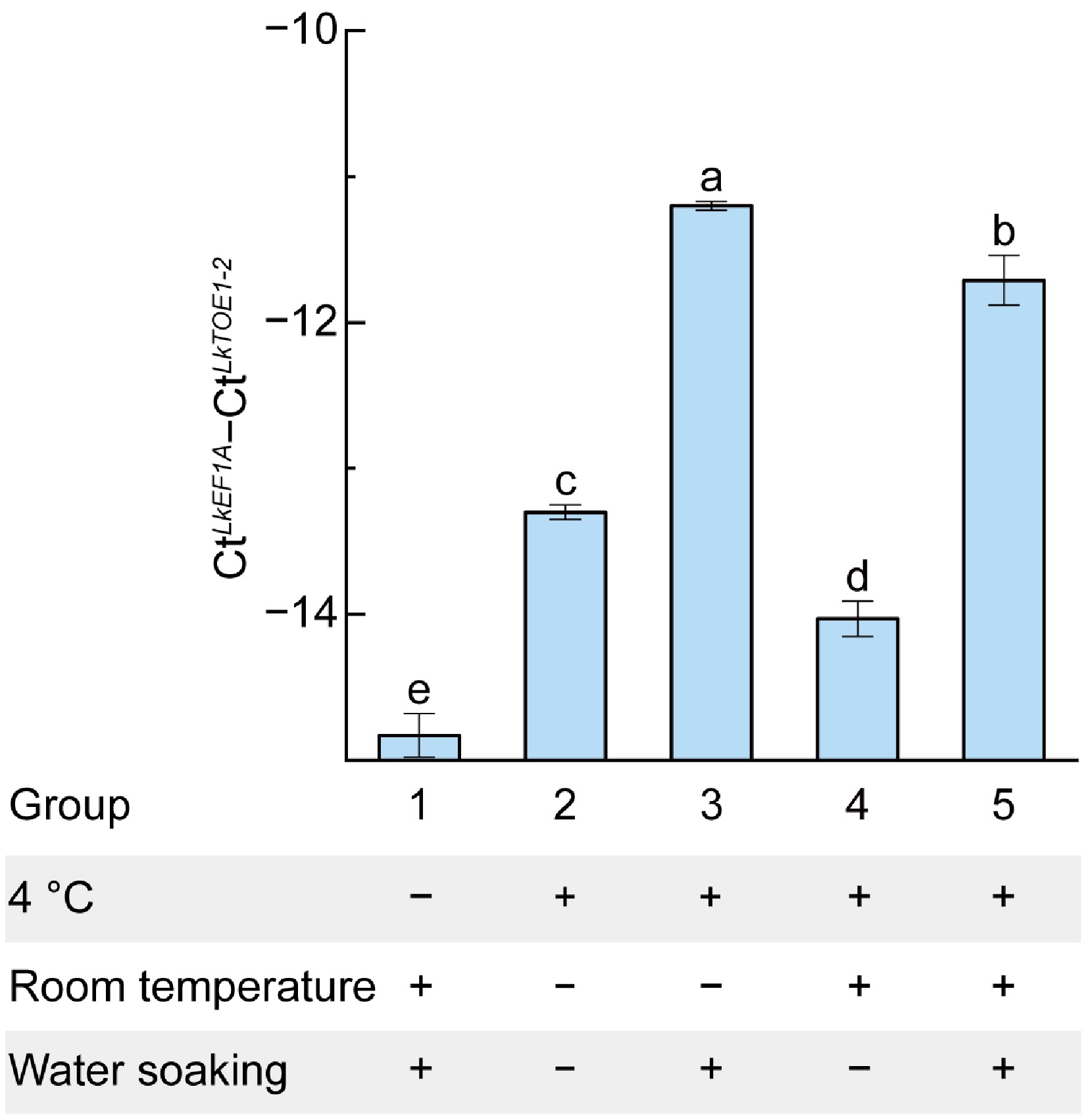
Figure 4.
Responses of LkTOE1-2 expression to temperature and water soaking detected by qRT-PCR. Different letters on the bars indicate significant differences as determined by one-way ANOVA followed by Tukey's multiple comparisons test, p < 0.05.
LkTOE1-2 over-expression promoted the germination, bolting, and the first flowering in A. thaliana
-
To test the function of LkTOE1-2, LkTOE1-2 was over-expressed in A. thaliana. After measuring the germination of transgenic seeds, it was found that LkTOE1-2 over-expression promoted the germination of A. thaliana seeds, as it took A. thaliana seeds over-expressing LkTOE1-2 2.02 d to germinate, while it took A. thaliana seeds expressing empty vector 2.65 d (Fig. 5a, b); it was ~0.63 d (23.77%) earlier (p < 0.001, Student' t-test). These data showed that LkTOE1-2 possesses able to enhance seed germination.
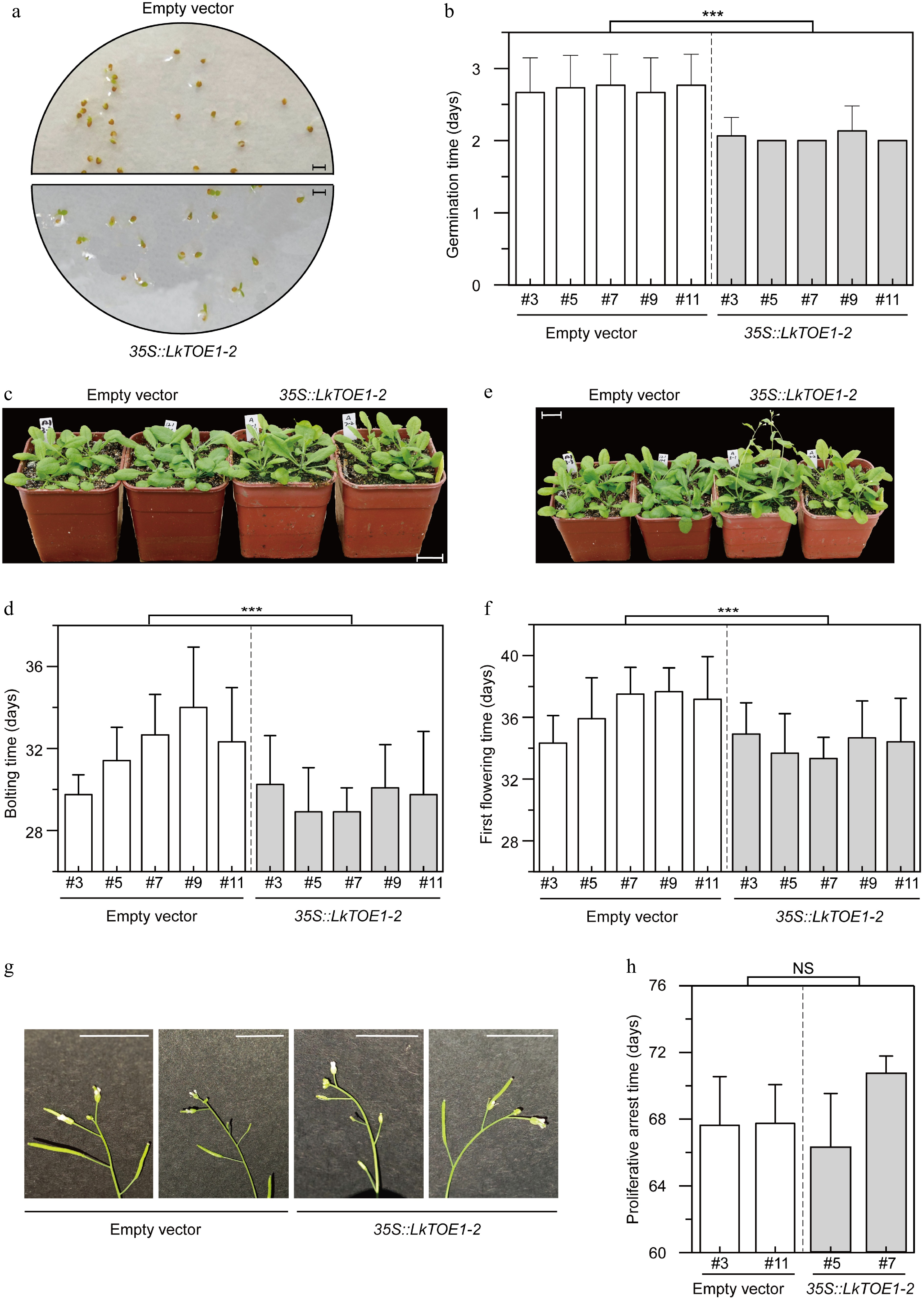
Figure 5.
Phenotypes of transgenic Arabidopsis thaliana over-expressing LkTOE1-2. (a) Germination of transgenic A. thaliana. Scale bar = 3 mm. The pictures were taken on the second day after 4 °C treatment. (b) Germination time of transgenic A. thaliana. n = 30. Germination was counted when two cotyledons were visible. (c) Bolting of transgenic A. thaliana. Scale bar = 2 cm. The pictures were taken on the 29th day after 4 °C treatment. (d) Bolting time of transgenic A. thaliana. Twelve plants were used in each line. Bolting was counted when inflorescence axis was visible, and bolting time was recorded from 4 °C treatment. n = 12. (e) Flowering of transgenic A. thaliana. Scale bar = 2 cm. The pictures were taken on the 31th day after 4 °C treatment. (f) First flowering time of transgenic A. thaliana over-expressing LkTOE1-2. First flowering time was recorded from 4 °C treatment. n = 12. (g) Proliferative arrest of the inflorescence axis in transgenic A. thaliana. Scale bar = 1 cm. The pictures were taken on the 65th day after 4 °C treatment. (h) Proliferative arrest time of the inflorescence axis in transgenic A. thaliana. Proliferative arrest was counted when a spherical structure is formed in the apex of the inflorescence axis, and proliferative arrest time was recorded from 4 °C treatment. n = 8. Transgenic A. thaliana expressing the empty vector was used as a control. *** p < 0.001, NS p ≥ 0.05, Student's t-test.
To test the function of LkTOE1-2 in the reproductive phase transition, the bolting time of transgenic A. thaliana was counted, and it was found that LkTOE1-2 over-expression resulted in the early reproductive phase transition, because it took A. thaliana over-expressing LkTOE1-2 29.58 ± 2.26 d to bolt, while it took A. thaliana expressing empty vector 31.78 ± 2.42 d (Fig. 5c, d); it was ~2.20 d (6.93%) earlier (p < 0.001, Student' t-test).
It was also found that the first flowering time was promoted by LkTOE1-2 over-expression, because it took A. thaliana over-expressing LkTOE1-2 34.17 ± 2.30 d and A. thaliana expressing empty vector 36.32 ± 2.54 d, respectively, to form the first flower (Fig. 5e, f); it was ~2.15 d (5.92%) earlier (p < 0.001, Student' t-test). The timing of proliferative arrest was not changed by LkTOE1-2 over-expression (Fig. 5g, h, p = 0.38, Student' t-test).
Based on these data, it was concluded that the function of LkTOE1-2 in bolting and flowering of A. thaliana was opposite to that of AtAP2[5,12−15]. The promotion of bolting (6.93%) and flowering (5.92%) was weaker than that of seed germination (23.77%), and it was speculated that this promotion might be affected by the early seed germination. Almost the same time was promoted in bolting and the first flowering, suggesting that the duration between bolting to the first flowering was not influenced by LkTOE1-2 over-expression.
The functions of some gymnosperm homologs of LkTOE1-2 in flowering have been revealed. In P. abies, PaAP2L1 does not cause detectable changes in plant morphology or flowering time, whereas PaAP2L2 delays flowering by an average of 4 d (19.0%) and increases the number of stamens and carpels in some flowers[61]. In L. kaempferi, LkAP2L1 delays flowering by an average of 8.5 d (34.4%), and increases organ size, but has no effect on floral organ identity[62]; LkAP2L2 increases shoot branching, although its effect on flowering time and floral organ identity is not prominent[28]. Collectively, these studies indicate that the functions of AP2 subfamily genes in flowering are different, and the mechanism underlying this functional diversification needs further study.
In addition, the lifetime was not affected by LkTOE1-2 because the proliferative arrest time is not changed in A. thaliana over-expressing LkTOE1-2. These results imply a potential role of LkTOE1-2 for shortening the juvenile phase of larch via molecular breeding.
-
This study provides the first comprehensive genome-wide identification and characterization of the AP2/ERF superfamily in L. kaempferi. Age-decreased expression patterns in the euAP2 lineage genes (LkTOE1-1/2/3/4) were revealed. Functional dissection confirmed that LkTOE1-2 expression is developmentally regulated during seed formation, somatic embryogenesis, and seed germination. Its promoter activity, validated by the RUBY reporter system, is enhanced during somatic embryo maturation. Over-expression of LkTOE1-2 in Arabidopsis accelerates seed germination, bolting time, and flowering. Collectively, these findings highlight not only divergent regulatory pathways in L. kaempferi compared with angiosperms but also provide LkTOE1-2 as a candidate gene for molecular breeding strategies aimed at shortening the juvenile phase and enhancing seed formation and germination in larch.
The authors thank Yubing He from the Institute of Crop Sciences, Chinese Academy of Agricultural Sciences for providing the proDR5::RUBY vector, and Dong-Xia Cheng and Tang-Quan Liao from the Chinese Academy of Forestry for help in RNA-seq analysis. This work was supported by Biological Breeding-National Science and Technology Major Projects (2022ZD0401602), and the National Natural Science Foundation of China (32271904).
-
The authors confirm their contributions to the paper as follows: carried out the study: Ye ZL, Li XY, Nong ML; analyzed the data, and wrote the manuscript: Ye ZL, Li XY; jointly supervised the study: Sun XM, Li W; conceived and designed the study: Li W; analyzed the data and revised the manuscript: Li W. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in the Supplemental Tables.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Zha-Long Ye, Xiang-Yi Li
- Supplementary Table S1 Expression levels of LkTOE1-2 assayed by qRT-PCR.
- Supplementary Table S2 Classification of AP2/ERF superfamily genes in Larix kaempferi.
- Supplementary Fig. S1 Verification of transgenic Arabidopsis thaliana.
- Supplementary Fig. S2 Multiple sequence alignment between AP2 proteins of Arabidopsis thaliana and Larix kaempferi, both containing a single AP2 domain, based on their conserved amino acid sequences.
- Supplementary Fig. S3 Phylogenetic tree of AP2 proteins from Arabidopsis thaliana and Larix kaempferi, both containing a single AP2 domain.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ye ZL, Li XY, Nong ML, Sun XM, Li W. 2025. The age-related expression patterns of Larix kaempferi AP2 subfamily genes and functional dissection of LkTOE1-2 in seed formation and germination. Forestry Research 5: e027 doi: 10.48130/forres-0025-0028 

The age-related expression patterns of Larix kaempferi AP2 subfamily genes and functional dissection of LkTOE1-2 in seed formation and germination
- Received: 01 July 2025
- Revised: 20 October 2025
- Accepted: 04 November 2025
- Published online: 19 November 2025
Abstract: Conifers pose challenges for breeding programs due to their extended vegetative phases. Despite the critical role of APETALA2 (AP2) in regulating phase transitions, the AP2/ERF superfamily remains largely unexplored in gymnosperms. Here, the first genome-wide analysis of the AP2/ERF superfamily in a conifer, Larix kaempferi (Japanese larch) is presented, and 374 members were identified. Among all eight paralogs, four euAP2 lineage genes, TARGET OF EATs (TOEs), exhibit age-decreased expression patterns. Functional characterization of LkTOE1-2 demonstrates its involvement in somatic embryogenesis and seed germination. Importantly, the RUBY reporter system confirmed an enhanced promoter activity in somatic embryo maturation. Over-expression of LkTOE1-2 in Arabidopsis accelerates seed germination by 23.77%, bolting by 6.93%, and flowering by 5.92%. This study provides not only an expanded genomic resource for gymnosperms but also a candidate gene (LkTOE1-2) to shorten the juvenile phase of larch via molecular breeding.
-
Key words:
- Aging /
- APETALA2 /
- Gymnosperms /
- Phase transition /
- RUBY reporter system /
- Somatic embryogenesis