









-
MicroRNAs (miRNAs), as non-coding RNAs with a length of 21 to 24 nucleotides (21−24 nts), recognized the post-transcriptional target genes with high complementarity, which could lead to target gene degradation or inhibition of translation and ultimately inhibiting the target gene expression[1]. MiRNAs were first discovered in Caenorhabditis elegans and produced by the RNase III-type enzyme Dicer from endogenous transcripts containing local hairpin structures[2]. In most ancient plants, miRNA sequences were highly conserved, and this conservation was their defining characteristic[3].
At present, many studies have reported the role of miRNAs in genes expression and translation regulation[4−7]. MiRNAs in plants were involved in many biological processes, including plant growth, biological and abiotic stress, and fruit development[8−11]. MiR160 down-regulated the auxin response factors (ARFs) to control the differentiation and growth of root[12], miR172 promoted flower opening by inhibiting Apetala2 (AP2) expression[13], miR169 enhanced plant drought resistance by affecting nuclear factor Y subunit A 5 (NFYA 5) expression at the translational level[14], miR393 regulated some genes from the plant hormone response pathway to enhance plant salt tolerance[13]. MiRNAs are also involved in the interaction between plants and climate, insects, pathogens, and the environment[15−17].
MiRNA-mediated regulation of fruit quality has been investigated. MiRNA172 was a key factor controlling apple fruit size[18]. miR156 targeted SPL9 protein has been shown to regulate flavonoid biosynthesis in Arabidopsis[19]. miR159a, miR396a, and miR171b were reported to be associated with carotenoid biosynthesis in cassava (Manihot esculenta Crantz)[20]. miR167, miR845, miR319, and miR166 regulated the flavonoid accumulation in tea plants, while miR171b and miR166d regulated terpenoid accumulation[21]; miR164 was associated with fruit ripening and expressed higher with sweet orange ripening[22]. In addition, some novel miRNAs regulating ripening-related processes have been identified in banana fruit[23].
Apple (Malus domestica Borkh.) is a widely cultivated fruit tree worldwide. Fruit flavor has been an important factor influencing market value and consumer preference[24]. At present, the main cultivated apple fruit aroma has been identified, and the aroma synthesis pathways have been preliminarily explored[24−28]. Fatty acid, amino acid, and carotenoid metabolism are important pathways for the production of apple aroma[29]. However, the molecular mechanisms involved in miRNA controlling these metabolic pathways are still unclear. At present, miRNA-mediated regulation of plant volatile organic compounds (VOCs) has only been reported in 'Nanguo' pear and tea plants[30,31]. To date, the function of miRNAs in apple aroma production is unknown.
Our previous study clearly defined the aroma characteristics of 'Binzi' (Malus domestica)[32]. The 'Binzi' — an ancient cultivar in China, had a special aroma which was only produced after five days of storage. The VOCs in 'Binzi' fruit were mainly concentrated in the peel. The critical aroma compounds in 'Binzi' peels have been identified as ethyl butyrate, ethyl caproate, ethyl 2-methylbutyrate, hexyl alcohol, 6-methyl-5-hepten-2-one, etc, which were from fatty acid metabolism, amino acid metabolism, and carotenoid metabolism (Table 1). Of these, ethyl butyrate imparted a unique pineapple flavor to 'Binzi' apple, distinguishing it from 'Starking' and 'Golden Delicious'[32]. Furthermore, some previous studies also didn't detect the ethyl butyrate in mature apples[24−28].
Table 1. Differential VOCs in 'Binzi' peels during storage (μg/kg)[32].
Compound Metabolism pathway Post-harvest storage period 0 d 5 d 10 d Ethyl butyrate Fatty acid metabolism – 3,439.60 1,727.43 Ethyl caproate Fatty acid metabolism – 1,167.97 782.60 Ethyl 2-methylbutyrate Amino acid metabolism – 1,546.96 383.70 Hexanal Fatty acid metabolism 93.26 171.20 454.31 Ethanol Fatty acid metabolism – 233.95 973.26 Hexyl alcohol Fatty acid metabolism 379.26 1,486.10 1,772.29 (E)-2-Hexen-1-ol Fatty acid metabolism 523.72 – – 6-Methyl-5-hepten-2-one Carotenoid metabolism 228.80 3,674.34 973.51 – not detected. In the present study, to understand the role of miRNAs on fruit aroma production during storage, six miRNA libraries (on the day of harvest with low aroma and on the 5th day of post-harvest storage with high aroma) from 'Binzi' peels were constructed and sequenced. We identified differentially expressed miRNAs (DEMs) that might be associated with aroma synthesis in comparison to low-aroma samples, predicted their target genes, and constructed a miRNA-mRNA regulatory network. We further verified the negative correlation in expression levels between miRNA and mRNA by qRT-PCR. Our results provisionally revealed the role of miRNA in aroma regulation.
-
The samples used in this study were consistent with those from our previous study[32]. The peel samples of 'Binzi' on the day of harvest were grouped as BZP01, and the peel samples on the 5th day of post-harvest storage were grouped as BZP02. Each group contained three biological replicates (namely BZP01a1, BZP01a2, BZP01a3, BZP02a1, BZP02a2, and BZP02a3). miRNAs from six 'Binzi' fruit samples were sequenced.
RNA extraction and sequencing
-
Total RNA was extracted from 'Binzi' peels and sent to Biomarker (Beijing, China) for small RNA sequencing. The detection of RNA samples was conducted using the Agilent 2100 bioanalyzer to assess RNA integrity and other parameters, ensuring the use of qualified samples for sequencing.
Library preparation
-
Following successful sample qualification, 2.5 ng of RNA was used as the starting material. The volume was adjusted to 6 μL with water, and library preparation was performed using the Small RNA Sample Prep Kit. Due to the phosphate group at the 5' end and hydroxyl group at the 3' end of small RNAs, T4 RNA Ligase 1 and T4 RNA Ligase 2 (truncated) were employed to ligate adapters to the 3' and 5' ends of the small RNAs, respectively. Subsequently, reverse transcription was performed to synthesize cDNA, followed by PCR amplification. Target fragments were selected via agarose gel electrophoresis, and the excised gel bands were purified to obtain the final small RNA library.
Data preprocessing and quality control
-
For each sample, the following quality control steps were performed: removal of low-quality reads, elimination of reads containing ≥ 10% unknown bases (N), exclusion of reads lacking the 3' adapter sequence, trimming of the 3' adapter, and filtration of sequences shorter than 18 nucleotides or longer than 30 nucleotides.
miRNA annotation and identification
-
The clean reads were aligned against the Silva database, GtRNAdb database, Rfam database, and Repbase database using Bowtie software to filter out non-coding RNAs (ncRNAs) such as ribosomal RNA (rRNA), transfer RNA (tRNA), small nuclear RNA (snRNA), small nucleolar RNA (snoRNA), and repetitive sequences, thereby obtaining unannotated reads containing miRNAs. Subsequently, the unannotated reads were mapped to the reference genome (Malus × domestica GDDH13 v1.1) using Bowtie software to acquire positional information on the genome, referred to as mapped reads. The mapped reads were analyzed using the miRDeep2 software package to identify known miRNA expression by aligning them against precursor sequences of known miRNAs in the miRBase database. For novel miRNA prediction, candidate precursor sequences were derived from the genomic loci of the mapped reads. Subsequently, a Bayesian model-based scoring system was applied, integrating the distribution patterns of reads across precursor sequences (characterized by mature, star, and loop regions reflecting miRNA biogenesis) and the structural energy characteristics of precursors (calculated using RNAfold and Randfold tools), to achieve high-confidence novel miRNA identification.
miRNA quantification
-
The expression of miRNAs was calculated using the TPM algorithm and normalised. The TPM normalization formula is expressed as:
TPM = Readcount × 1,000,000 – Mapped Reads.
In the formula, Readcount denotes the number of reads mapped to a specific miRNA; Mapped Reads represents the total number of reads aligned to all miRNAs in the sample.
Differentially expressed miRNA (DEM) analysis
-
Differential expression analysis between the two groups was performed by DESeq R package (1.10.1). The miRNAs with |log2(FC)| ≥ 1 and p-value ≤ 0.05 were assigned as differential expressed miRNAs.
Prediction of miRNA target genes
-
Target gene prediction was performed using the TargetFinder software based on sequence information from both known miRNAs and novel miRNAs, along with their corresponding species-specific genomic sequences. Subsequently, the predicted target gene sequences were aligned against the NR, Swiss-Prot, GO, COG, KEGG, KOG, and Pfam databases using BLAST to obtain functional annotations of the target genes.
qRT-qPCR validation
-
Two miRNAs and four target genes associated with aroma metabolic pathways were selected. Sequence-specific primers for real-time quantitative PCR (qRT-PCR) were designed. Reverse transcription was performed using the RNA provided by the company to generate cDNA templates. Subsequently, qRT-PCR was conducted to validate the expression levels of the selected miRNAs and their candidate target genes.
The reverse transcription of mRNA was performed using PrimeScript First Strand cDNA Synthesis Kit (TaKaRa, Japan). The expression level of the target genes came from RNA-seq. The reaction system consisted of 2 μL of total RNA, 4 μL of 5 × TURE Reaction Mix, 1.5 μL of oligo (dT), with RNase Free ddH2O added to a total volume of 20 μL. The reaction conditions were incubation at 42 °C for 30 min, followed by incubation at 85 °C for 5 min.
The reverse transcription of miRNA was performed using an miRNA First Strand cDNA Synthesis Kit (by tailing A) (Vazyme, China). The total RNA was used as a template, and Poly A polymerase (PAP) was added to its 3' end, followed by the reverse transcription reaction using universal reverse transcription primers. The reaction system included 1 μL of total RNA, 2 μL of HiScript miRNA Enzyme Mix, 10 μL of 2×miRNA RT Mix, with RNase Free ddH2O added to a total volume of 20 μL. The reaction conditions were incubation at 37 °C for 60 min, and 85 °C for 5 min.
The qRT-PCR was performed using SYBR® Green Supermix (Vazyme, China) in a Light Cycler 96 quantitative PCR detection system (Roche, Switzerland). The detailed reaction system and method were as described in our previous study[33]. The reaction mixture (10 μL total volume) contained: 1 μL cDNA, 3 μL ddH2O, 0.5 μL forward/reverse primers (each), and 5 μL 2 × SYBR® Green Supermix. Thermal cycling conditions were as follows: initial denaturation at 95 °C for 3 min; 40 cycles of 95 °C for 10 s (denaturation) and 60 °C for 30 s (annealing/extension); followed by a melt curve analysis from 60 to 95 °C with an increment of 1 °C per cycle. Reactions were held at 4 °C for storage. All qRT-PCR assays were performed in three technical replicates, and MdActin was used as the internal reference gene. The primer sequences of miRNA and mRNA-specific primers used are listed in Supplementary Table S1. The reaction system was performed according to the manufacturers' instructions. The relative expression levels were calculated with the 2−ΔΔCᴛ method[34].
-
We filtered the small RNA sequence data from six 'Binzi' peel samples, and a total of 79,202,746 clean reads were ultimately obtained. The alignment efficiency of clean reads ranged from 45.96% to 52.57% (Supplementary Table S2).
A total of 472 miRNAs, including 183 known miRNAs and 289 novel miRNAs, were identified. The lengths of these miRNAs were mainly 21 and 24 nt, respectively (Fig. 1a–f). In the BZP01 group, 452, 435, and 436 miRNAs were detected across three replicates, while in the BZP02 group, 450, 444, and 441 miRNAs were identified respectively. By analyzing miRNAs common across all three replicates, a total of 438 core miRNAs were identified, with 433 miRNAs detected in BZP01, and 428 miRNAs in BZP02 (Fig. 1g–h).

Figure 1.
(a)−(f) The length distribution of miRNAs in six samples. Three repeated samples venn plots of (g) BZP01, and (h) BZP02. The (i) first bias preference, and (j) each point base bias.
Dicer and DCL enzymes exhibited a stronger bias towards the 5' first base U when recognizing and cleaving precursor miRNAs. By analysing the base preference of miRNA, typical miRNA base ratios can be obtained. The first base preference of miRNAs is shown in Fig. 1i. First base preference of miRNA had a stronger preference for U. Fig. 1j shows the base preference on each site and pointed out that the tenth base preferred for A. The preferences of base complied with the rules of miRNA, which indicated the reliable results in our study.
miRNAs expression analysis
-
The miRNAs TPM expression data in six samples were shown in Supplementary Table S3. A total of 423 core miRNAs were shared between BZP01 and BZP02, with 15 miRNAs uniquely detected at specific storage periods (Fig. 2a). Compared to BZP01, 199 miRNAs were down-regulated and 239 up-regulated in BZP02 (Fig. 2b). Hierarchical clustering of TPM expression levels for these miRNAs revealed that the six sequencing samples segregated into two distinct clusters, with triplicates from the same storage period grouping together, suggesting stage-specific regulatory roles of miRNAs (Fig. 2c). According to |log2(FC)| ≥ 1 and p-value ≤ 0.05, 14 miRNAs were differentially expressed in BZP02 compared with BZP01, among which nine DEMs were significantly up-regulated and five DEMs were significantly down-regulated in BZP02 compared to BZP01 (Fig. 2d). Ultimately, 29 DEMs (15 miRNAs uniquely detected at specific storage periods and 14 miRNAs differentially expressed in different storage periods) were predicted to potentially regulate aroma compound biosynthesis in 'Binzi' apple peels.

Figure 2.
The expression levels of miRNAs in BZP01 and BZP02. (a) Venn plots of BZP01 and BZP02. (b) The number of upregulated and downregulated miRNAs. (c) Expression heatmap of 438 miRNAs between BZP01 and BZP02. (d) Volcano plot of BZP01 vs BZP02.
Analysis of target genes of DEMs
-
To understand the functions of miRNAs from six 'Binzi' samples, 604 genes targeted by 29 DEMs were predicted. GO and KEGG enrichment analysis are shown in Fig. 3. GO enrichment analysis showed that 604 targeted genes were classified into three categories (containing 43 terms) (Fig. 3a). In biological processes (BPs), the major terms were cellular process (144), metabolic process (137), and single-organism process (113); in molecular functions (MFs), the major terms were catalytic activity (126) and binding (126); the major terms of cellular components (CCs) were cell (78), cell part (78), and organelle (56).

Figure 3.
GO and KEGG enrichment analysis of 604 genes targeted by 29miRNAs. (a) GO enrichment analysis. (b) KEGG pathway annotation. (c) The top 20 KEGG pathway enrichment.
KEGG pathways showed that 604 targeted genes were classified into five categories (included 17 terms) (Fig. 3b). The major terms were global and overview maps (67), biosynthesis of other secondary metabolites (16), carbohydrate metabolism (15), and amino acid metabolism (15). Furthermore, metabolic pathways and biosynthesis of secondary metabolites were the most significantly enriched categories, harboring 66 and 44 targeted genes, respectively. Key pathways related to aroma metabolism—fatty acid metabolism (6), unsaturated fatty acid biosynthesis (2), and carotenoid biosynthesis (3)—ranked among the top 20 enriched KEGG pathways. These five pathways (highlighted in red boxes, Fig. 3c) were probably functionally associated with physiological changes and VOC biosynthesis during the storage of 'Binzi' apples.
We further analysed the relationship between mRNAs and miRNAs within the five aforementioned metabolic pathways (highlighted in red boxes, Fig. 3c). A total of 67 mRNAs across these pathways were targeted by 27 distinct miRNAs (Fig. 4). mdm-miR159c, novel_Chr10_6452, and mdm-miR408c exhibited the highest number of targeted genes, regulating 18, 12, and eight mRNAs, respectively. However, only five targeted genes were functionally annotated to fatty acid metabolism and unsaturated fatty acid biosynthesis, all exclusively targeted by mdm-miR408c. In contrast, the targets of mdm-miR159c and novel_Chr10_6452 were primarily associated with metabolic pathways and biosynthesis of secondary metabolites. These results suggest that mdm-miR408c may act as a critical miRNA regulating aroma biosynthesis in 'Binzi' peels, while mdm-miR159c and novel_Chr10_6452 likely modulate physiological changes during fruit storage.
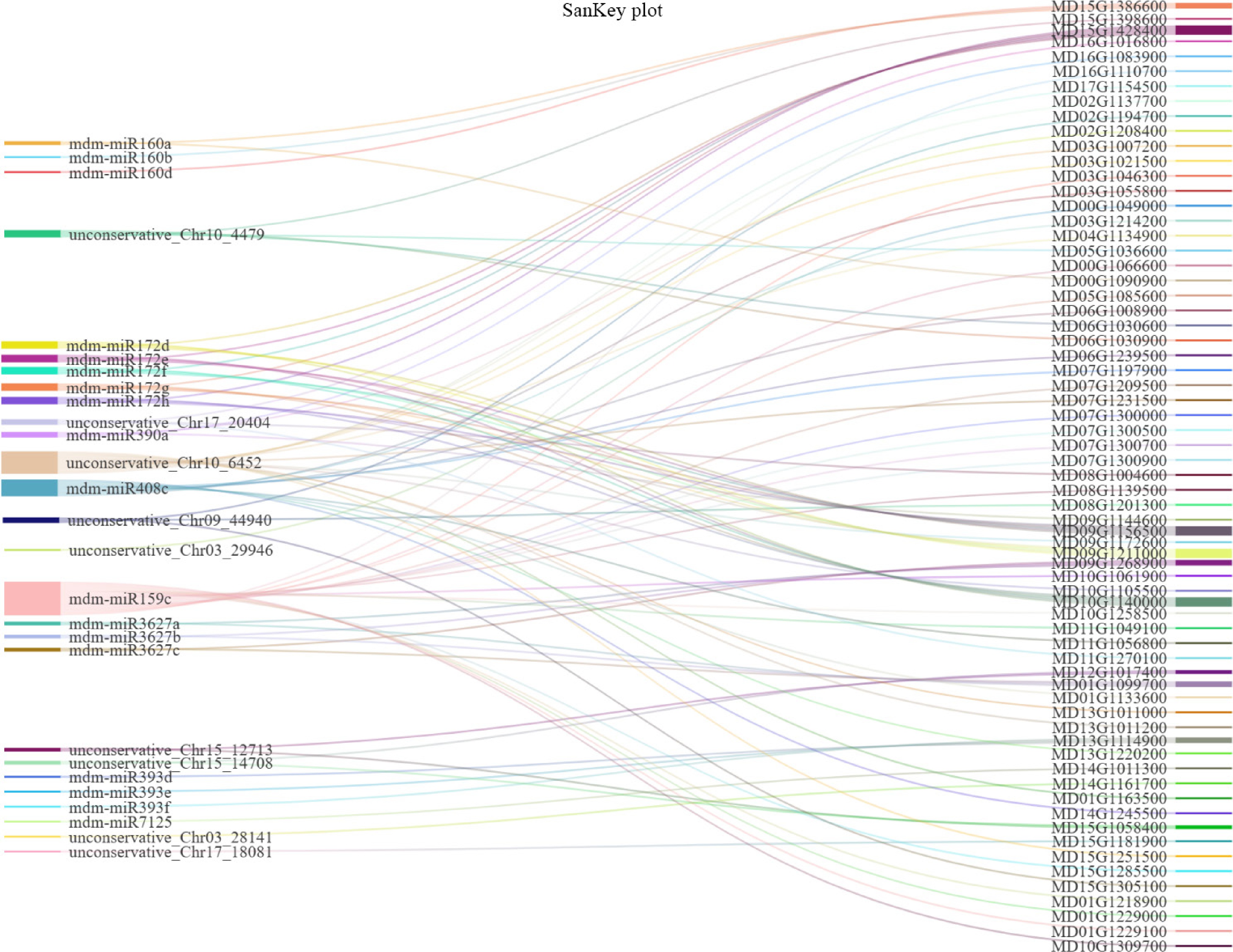
Figure 4.
Sankey diagram of 67 mRNAs and 27 miRNAs. The mRNAs derived from five pathways (highlighted in red boxes, Fig. 3c) associated with physiological changes and VOCs biosynthesis of 'Binzi' peels.
Regulatory networks of miRNAs involved in 'Binzi' aroma biosynthesis
-
To study the biosynthesis of volatile organic compounds during the post-harvest storage of 'Binzi' fruit at the post-transcriptional level, a regulatory network of mRNAs and miRNAs was constructed (Fig. 5 & Table 2). Based on the characteristics of VOCs of 'Binzi' peels, seven metabolic pathways involved in aroma synthesis were screened from the 72 KEGG-enriched pathways, including four biosynthetic pathways (biosynthesis of unsaturated fatty acids (ko01040), fatty acids (ko00061), amino acids (ko01230), and carotenoid (ko00906)), two metabolism pathways (metabolism of fatty acid (ko01212) and alpha-linolenic acid (ko00592)), and one fatty acid degradation (ko00071). A total of six miRNAs (mdm-miR408c, mdm-miR159c, mdm-miR390a, mdm-miR7125, novel_Chr03_29946, and novel_Chr17_18081) were found to be involved in these seven pathways, and their relationship network with target genes was shown in Fig. 5. The six miRNAs collectively targeted 274 mRNAs and their interaction network with the target genes is illustrated in Fig. 5. However, only 10 target genes were functionally linked to the seven aroma-related metabolic pathways. Pathway annotations for both miRNAs and their target genes are summarized in Table 2. Based on the pathway annotation of the target genes, the regulatory networks were categorized into three modules: fatty acid metabolism, amino acid biosynthesis, and carotenoid biosynthesis.
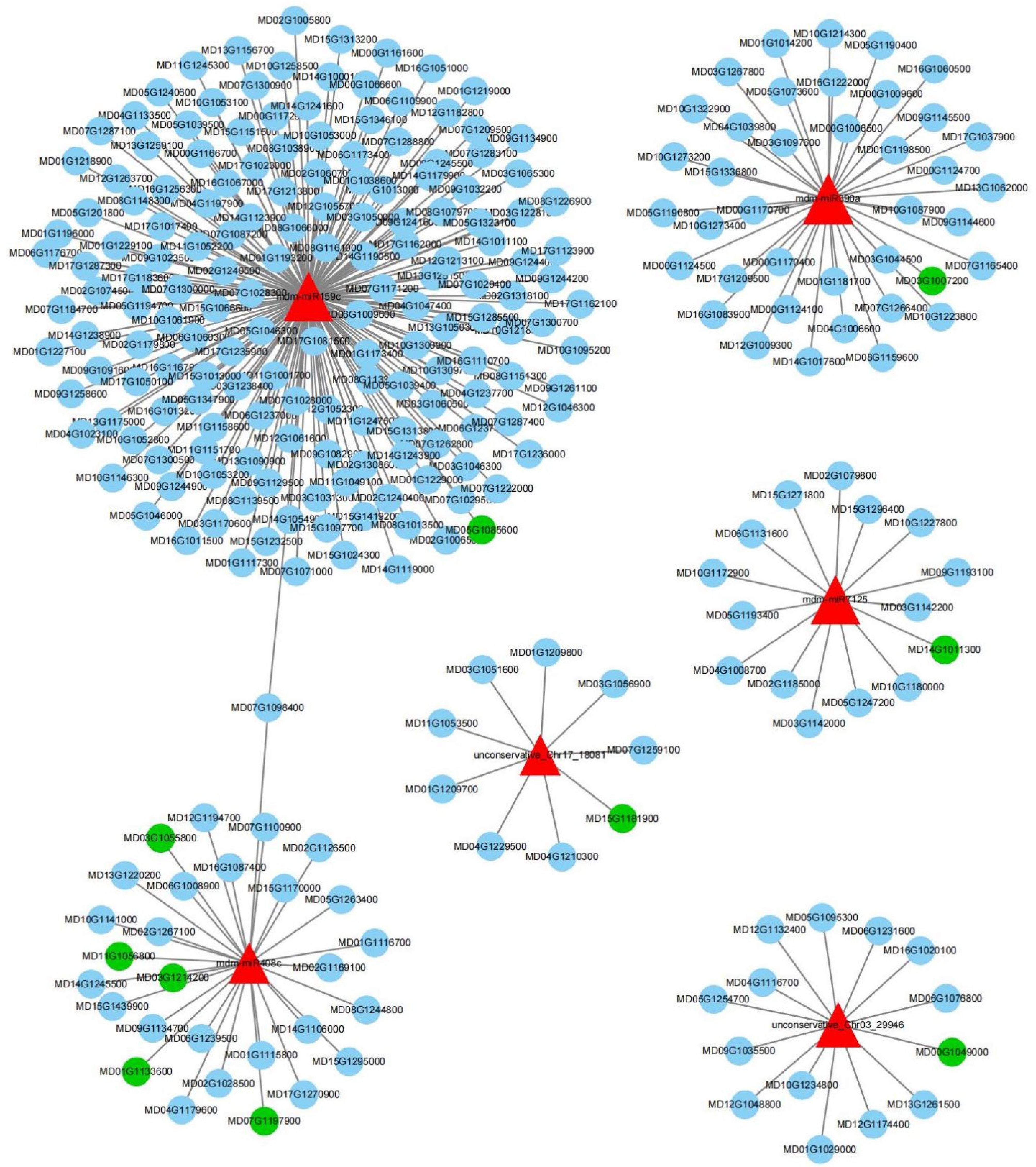
Figure 5.
Network of miRNA and mRNA involved in volatile organic components synthesis. The red triangle represents miRNAs, the blue circles represent targeted genes, and the green circles represent genes related to the pathways of aroma production.
Table 2. Pathways detail of miRNAs and target genes involved in volatile organic component synthesis.
Pathway Pathway ID miRNAs Target genes Target genes name K_id K_id entry Fatty acid metabolism ko01212 mdm-miR408c MD03G1214200 3-Oxoacyl-[acyl-carrier protein] reductase (fabG) K00059 [EC:1.1.1.100] mdm-miR408c MD03G1055800 [acyl-carrier-protein]
S-malonyltransferase (fabD)K00645 [EC:2.3.1.39] mdm-miR408c MD11G1056800 [acyl-carrier-protein]
S-malonyltransferase (fabD)K00645 [EC:2.3.1.39] mdm-miR408c MD07G1197900 Very-long-chain enoyl-CoA reductase (TER) K10258 [EC:1.3.1.93] mdm-miR408c MD01G1133600 Very-long-chain enoyl-CoA reductase (TER) K10258 [EC:1.3.1.93] novel_Chr17_18081 MD15G1181900 Enoyl-CoA hydratase/
3-hydroxyacyl-CoA dehydrogenase (MFP2)K10527 [EC:4.2.1.17, 1.1.1.35, 1.1.1.211] Biosynthesis of unsaturated fatty acids ko01040 mdm-miR408c MD07G1197900 Very-long-chain enoyl-CoA reductase (TER) K10258 [EC:1.3.1.93] mdm-miR408c MD01G1133600 Very-long-chain enoyl-CoA reductase (TER) K10258 [EC:1.3.1.93] Fatty acid biosynthesis ko00061 mdm-miR408c MD03G1214200 3-Oxoacyl-[acyl-carrier protein] reductase (fabG) K00059 [EC:1.1.1.100] mdm-miR408c MD03G1055800 [acyl-carrier-protein]
S-malonyltransferase (fabD)K00645 [EC:2.3.1.39] mdm-miR408c MD11G1056800 [acyl-carrier-protein]
S-malonyltransferase (fabD)K00645 [[EC:2.3.1.39] Fatty acid degradation ko00071 novel_Chr17_18081 MD15G1181900 Enoyl-CoA hydratase/
3-hydroxyacyl-CoA dehydrogenase (MFP2)K10527 [EC:4.2.1.17 1.1.1.35 1.1.1.211] alpha-Linolenic acid metabolism ko00592 novel_Chr17_18081 MD15G1181900 Enoyl-CoA hydratase/
3-hydroxyacyl-CoA dehydrogenase (MFP2)K10527 [EC:4.2.1.17 1.1.1.35 1.1.1.211] Biosynthesis of amino acids ko01230 mdm-miR7125 MD14G1011300 Cysteine synthase (cysK) K01738 [EC:2.5.1.47] Carotenoid biosynthesis ko00906 mdm-miR390a MD03G1007200 15-Cis-phytoene synthase (crtB) K02291 [EC:2.5.1.32] novel_Chr03_29946 MD00G1049000 Lycopene beta-cyclase (lcyB) K06443 [EC:5.5.1.19] mdm-miR159c MD05G1085600 Abscisate beta-glucosyltransferase (AOG) K14595 [EC:2.4.1.263] Within the fatty acid metabolism regulatory network, mdm-miR408c and novel_Chr17_18081 were identified as the miRNAs orchestrating these pathways. The mdm-miR408c had the highest number of target genes, mainly related to the biosynthesis and metabolism of fatty acids and unsaturated fatty acids. These genes included 3-oxoacyl-[acyl-carrier protein] reductase (MdfabG: MD03G1214200), [acyl-carrier-protein] S-malonyltransferase (MdfabD: MD03G1055800 and MD11G1056800), and very-long-chain enoyl-CoA reductase (MdTER: MD07G1197900 and MD01G1133600). In addition, novel_Chr17_18081 also targeted enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase (MdMEFP2: MD15G1181900) and was involved in regulating the metabolism and degradation of fatty acids and unsaturated fatty acids.
The amino acid biosynthesis regulatory network only involved one miRNA. The mdm-miR7125 targeted the cysteine synthase (MdcysK: MD14G1011300) to regulate amino acid biosynthesis.
The carotenoid biosynthesis regulatory network involved three miRNAs: mdm-miR390a, mdm-miR159c, and the novel miRNA novel_Chr03_29946. The mdm-miR390a targeted 15-cis-phytoene synthase (MdcrtB: MD03G1007200), mdm-miR159c targeted abscisate beta-glucosyltransferase (MdAOG: MD05G1085600), novel_Chr03_29946 targeted lycopene beta-cyclase (MdlcyB: MD00G1049000) involved in carotenoid biosynthesis.
The targeting relationship between these six miRNAs and 10 target genes are shown in Supplementary Table S4. However, the interaction between miRNAs and target genes needed further study.
Verification of miRNAs and target genes by qRT- PCR
-
To verify the miRNA-seq results, two miRNAs (mdm-miR408c and novel_Chr17_18081), and four putative target genes (MD03G1055800, MD11G1056800, MD15G1181900, and MD03G1214200) were selected for qRT-PCR. The qRT-PCR expression patterns of miRNAs were consistent with their TPM values derived from miRNA sequencing (Fig. 6), validating the reliability of the transcriptomic data. Furthermore, mdm-miR408c and novel_Chr17_18081 exhibited significant negative correlations with the expression levels of their putative target genes across, suggesting potential regulatory interactions between these two miRNAs and the four candidate target genes.
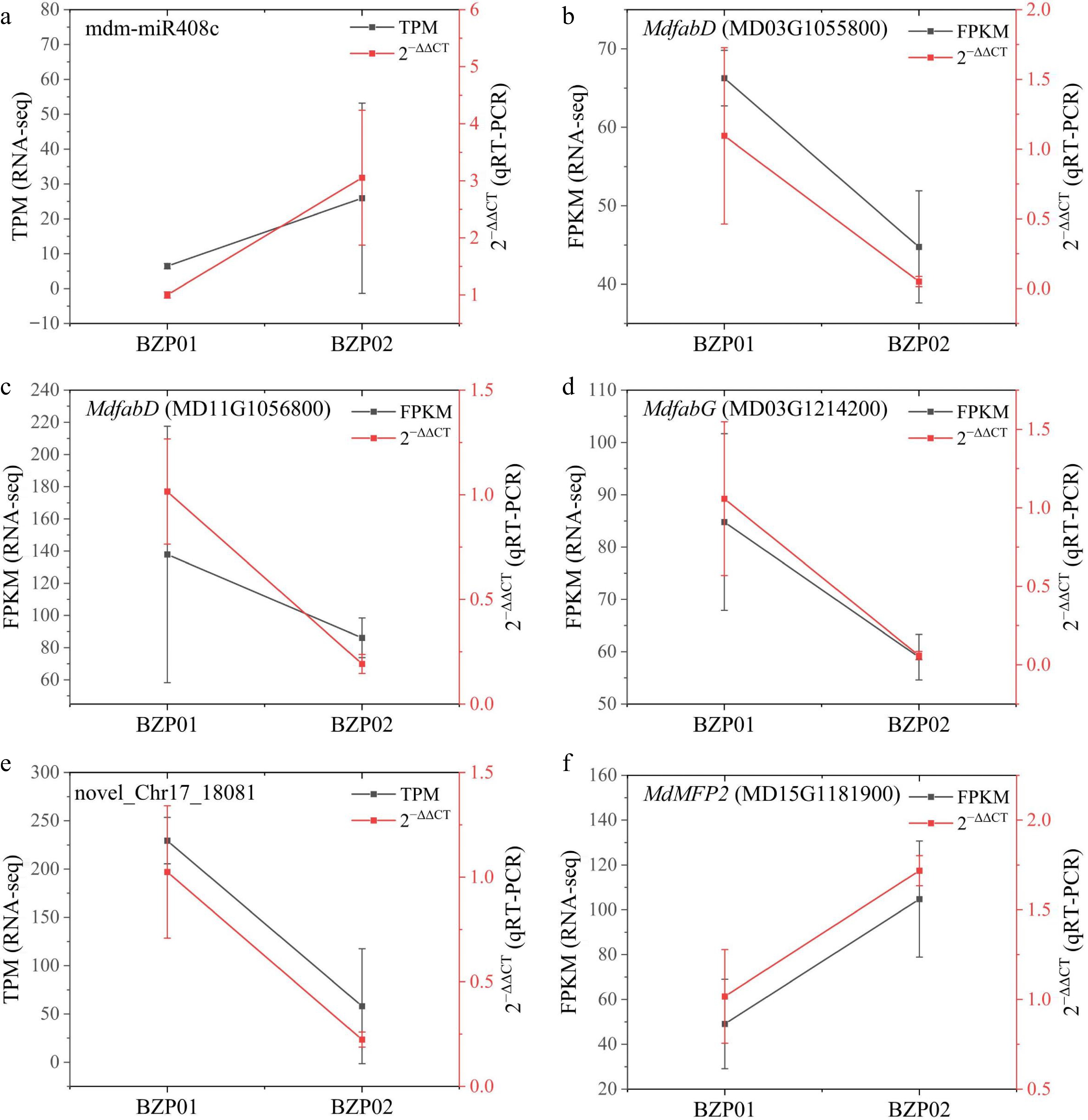
Figure 6.
Transcriptome expression levels and qRT-PCR of miRNAs and target genes. (a) The FPKM and 2−∆∆Ct of mdm-miR408c and its target gene (b) MdfabD (MD03G1055800), (c) MdfabD (MD11G1056800), and (d) MdfabG (MD03G1214200). (e) The FPKM and 2−∆∆Ct of novel_Chr17_18081 and its target gene MdMFP2 (MD15G1181900).
-
Fruits are rich in vitamins, dietary fiber, pectin, and carbohydrates, and are important food for humans[35]. Nutrient components, texture, flavor, and color, affect the market value of fruit. More importantly, fruit quality during the post-harvest storage stage directly affects their long-term value[36]. The fruit undergoes a series of physiological processes during post-harvest storage, such as ripening, ageing, and softening. These physiological processes are constantly occurring and changing the quality of the fruit, such as aroma production, coloring, and browning[37].
miRNAs, as non-coding RNAs, play roles in regulating plant biological processes by modulating target gene expression[38]. Studies have thus far confirmed the role of miRNAs in modulating the development and formation of fruit quality[38]. 'Binzi', as a natural hybrid of Mianpingguo (Malus pumila) and Shaguo (Malus asiatica), has a strong aroma but poor storage properties and is prone to softening and browning during post-harvest storage[39]. In this study, we identified 183 known miRNAs and 289 novel miRNAs, and 604 genes were predicted to be targeted by 29 DEMs. GO enrichment analysis showed that 604 target genes could be classified into three GO terms, including biological processes, molecular functions, and cellular components. Metabolic pathways and biosynthesis of secondary metabolites in KEGG enriched the most target gene amounts. This result indicated that miRNAs are involved in the complex physiological changes of 'Binzi' fruit during storage. Previous studies have shown that miR159-MYBs, miR172-AP2, and miR164-NACs are related to fruit ripening or senescence[40,41]. miR164, miR156, miR319, miR6478, miR528, and miR159 are associated with fruit browning in apple, banana, and strawberry[25,38]. In this study, miR159, miR172, miR164, and miR319 showed higher expression levels, which might lead to the storage intolerance of 'Binzi' fruit. Apples are a typical climacteric fruit, and the production of ethylene strongly affects the quality of apples during storage[29]. Studies showed that miR156 and miR157 controlled ethylene synthesis in apple fruit, while miR395 participated in ethylene mediated ripening[38]. In our study, miR156 and miR395 showed different expression patterns at different stages of storage. Therefore, it could be speculated that miRNAs were involved in regulating the physiological processes of 'Binzi' fruit during post-harvest storage, such as ripening, softening, ageing, and ethylene release, which further affected the production of 'Binzi' aroma.
Studies have shown that 'Binzi' produces a large amount of aroma during storage, with ethyl butyrate, ethyl 2-methylbutyrate, 6-methyl-5-hepten-2-one, and β-damascenone being the critical compounds. These aroma compounds are derived from fatty acids, amino acids, and carotenoid metabolism, respectively[32]. Saturated fatty acids, and unsaturated fatty acids are the two substrates of fatty acid metabolism[29]. Saturated fatty acids are degraded through β-oxidation pathways, and the volatile alcohols, aldehydes, and esters are the final products[42]. Alpha-linolenic acid is the main unsaturated fatty acid producing volatile compounds via the lipoxygenase pathway[29]. In this study, four known miRNAs and two novel miRNAs targeted ten genes, which were associated with the biosynthesis of unsaturated fatty acids, fatty acids, amino acids, carotenoids, metabolism of fatty acids, alpha-linolenic acid, and fatty acid degradation. Among them, five genes targeted by mdm-miR408c were related to the biological processes of fatty acids, including synthesis, metabolism, and degradation; three genes targeted by mdm-miR390a, mdm-miR159c, and novel_Chr03_29946 were involved in carotenoid biosynthesis, while one gene targeted by mdm-miR7125 was involved in amino acid biosynthesis. The relationship between miRNAs and mRNA was further confirmed by qRT-PCR. mdm-miR408c was negatively correlated with MdFabG (MD03G1214200), and two MdFabDs (MD03G1055800 and MD11G1056800); novel_Chr17_18081 was negatively correlated with MdMFP2 (MD15G1181900). In 'Binzi' fruit, ethyl butyrate with a pineapple, fruity, and apple flavor was the critical compound that differed from other apple cultivars, such as 'Golden Delicious', 'Delicious', 'Fuji', 'Ralls', 'Honeycrisp', 'Huahong' apples, etc[25−27,43]. The results indicated that mdm-miR408c and novel_Chr17_18081 might be jointly involved in regulating the characteristic aroma of 'Binzi'.
MiR408 is an ancient non-coding miRNA and conserved in plants and is the important regulator of plant vegetative growth and reproductive development[44]. At present, studies have found that the over-expression of miR408 could significantly promote the development processes of plants, including the growth and development of seeds, roots, leaves, and flowers[45,46]. The photosynthetic activity and seed yield were also increased by miR408[45]. Multiple environmental factors could also affect the expression level of miR408, such as cold, salinity, drought, nutritional deficiency, heavy metals, lipopolysaccharides, and disease[46−49]. The expression levels of cupredoxin and LAC3 targeted by miR408 were decreased, which increased the cold resistance of A. thaliana[46]. Tea trees could produce more volatile compounds when subjected to low-temperature stress[50]. After low-temperature storage, the aroma of 'Nanguo' pear weakened and the expression level of miR408 was downregulated[30]. miR408 could also reduce the expression levels of ROS and Cu-containing proteins to increase the endogenous availability of Cu[50]. Peñarrubia et al.[51] found that Cu homeostasis could affect changes in the levels of plant hormones (ABA and ethylene). This indicated that miR408 probably indirectly affected the production of VOCs by affecting the steady-state levels of endogenous Cu, thereby affecting ethylene production. We also found that mdm-mir408c was significantly up-regulated in this study, and the aroma content of 'Binzi' also increased[32]. However, our study did not establish a definitive correlation between miR408 expression and ethylene signaling or endogenous copper homeostasis. Whether miR408 influences aroma accumulation by adjusting these pathways is unknown.
FabD and FabG are the key enzymes in the fatty acids biosynthesis pathways. The first step in fatty acid biosynthesis is the conversion of acetyl-CoA to malonyl-CoA by acetyl-CoA carboxylase. The second step is the conversion of malonyl-CoA to malonyl-ACP through the enzyme FabD. Then the malonyl-ACP is converted to (3R)-3-Hydroxyacyl-ACP through the enzyme FabG. Ultimately, free long-chain fatty acids are produced by the enzyme fatty acyl-ACP thioesterase[52]. However, excessive production of free fatty acids probably delays the formation of aroma[53]. Shi et al.[54] found that low temperatures led to the up-regulation of fadD and fadG in 'Nanguo' pear, but its aroma content decreased. In our study, the up-regulated expression level of mdm-miR408c during storage negatively regulated the expression of MdfadD and MdfadG, and more aroma was produced. The down-regulation of these genes could reduce the content of free fatty acids and promote the production of volatile compounds. A similar result was found for the increased levels of linoleic and linolenic acids, which would affect the production of VOCs[55].
β-oxidation is the main process by which to degrade the straight-chain fatty acids in higher plants[56]. Fatty acids are degraded by the sequential removal of two carbon units. The enzyme MFP2, with the activities of 2-trans-enoyl-CoA hydratase and L-3-hydroxyacyl-CoA dehydrogenase activities, plays a role in catabolizing enoyl-CoA substrates in Arabidopsis thaliana[57]. Subsequently, volatile aldehydes, alcohols, and esters are formed under the catalysis of aldehyde dehydrogenase (ALDH), alcohol dehydrogenase (ADH), and alcohol acyltransferase (AAT) enzymes. In addition, MFP2 could promote the production of methyl jasmonate, which in turn up-regulates the expression of lipoxygenase (LOX), thereby increasing the accumulation of volatile compounds[58]. In this study, the gene MdMFP2 targeted by novel_Chr17_18081 was up-regulated during storage, while the novel_Chr17_18081 was down-regulated. An increase in the expression level of MFP2 was found with the improvement of the flavor of Rougui tea (Cinnamomum cassia) and Oolong tea (Camellia sinensis)[59,60].
-
Our research showed that the aroma production of 'Binzi' was jointly regulated by mdm-miR408c, novel_Chr17_18081, and their target genes (MdfabD, MdfabG, and MdMFP2). During the storage of 'Binzi', the up-expressed mdm-miR408c inhibited the expression of MdfabD and MdfabG, leading to a decrease in the content of free long-chain fatty acids and further promoted the production of more aroma compounds. At the same time, the down-expression in novel_Chr17_18081 led to the up-expression of MdMFP2, which promoted the conversion of long-chain fatty acids to short-chain fatty acids, thereby promoting aroma production (Fig. 7). Our study provides new insight into miRNAs in mediating aroma formation, and mdm-miR408c could be used as an important research object to uncover more secrets about the aroma of 'Binzi' apple.
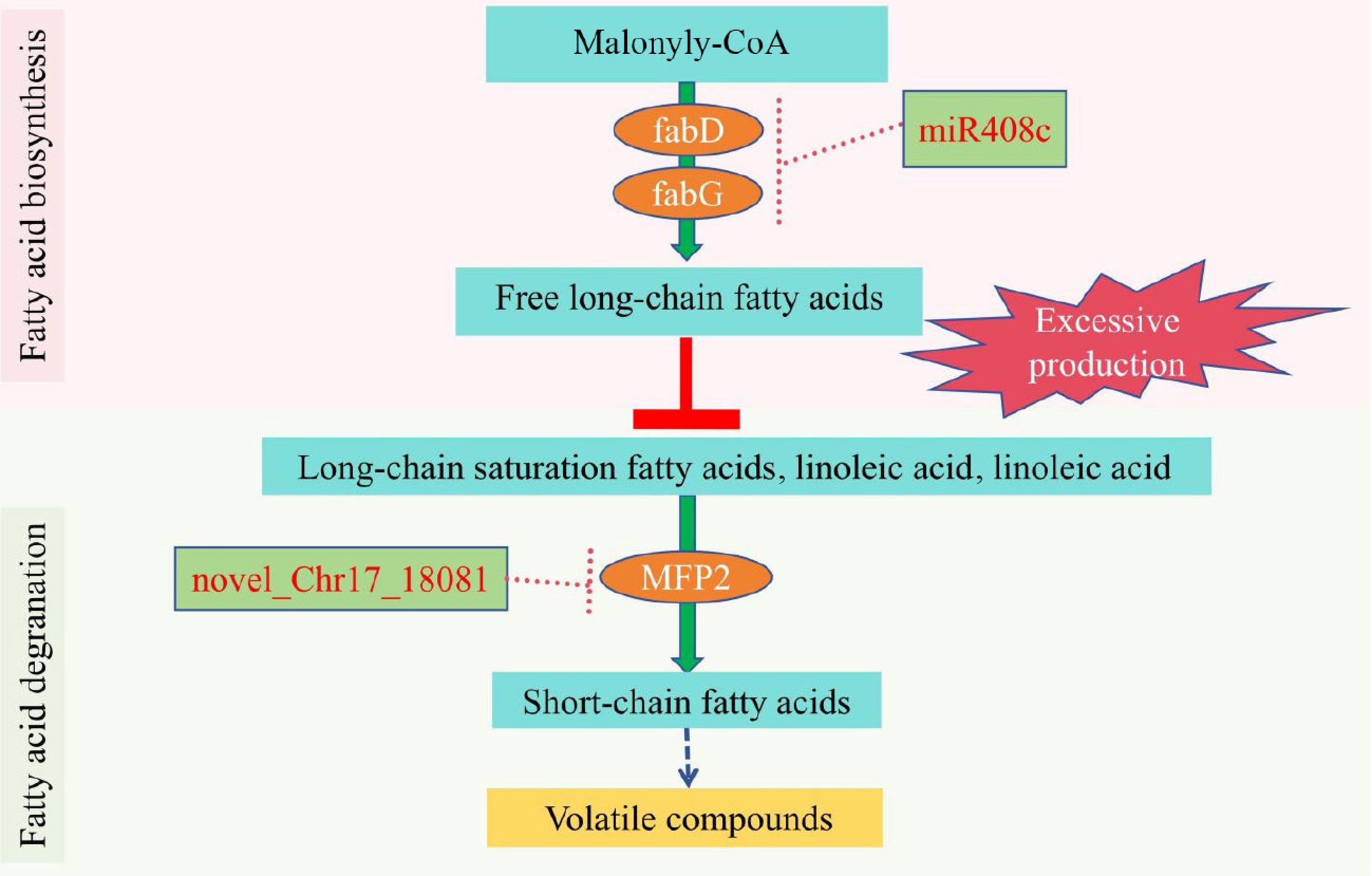
Figure 7.
A working model on the role of miR408c, novel_Chr17_18081, MdfabD, MdfabG, and MdMFP2 in regulating volatile aroma compounds synthesis in 'Binzi' apple. MdfabD: [acyl-carrier-protein] S-malonyltransferase. MdfabG: 3-oxoacyl-[acyl-carrier protein] reductase. MdMFP2: enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase.
We would like to thank OmicShare tools, a free online platform for plotting (
www.omicshare.com/tools , accessed on 31 December 2024), the Hiplot team (https://hiplot.com.cn , accessed on 31 December 2024) for providing technical assistance and valuable tools for data analysis and visualization. This work was supported by the National Key Research and Development Plan (Grant No. 2023YFD120010402), and the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (Grant No. CAAS-ASTIP-2021-RIP-02).-
The authors confirm contribution to the paper as follows: study conception and design: Wang D, Wang K, Feng J; data collection: Lu X, Li Z, Guo H, Li L, Wang L; analysis and interpretation of results: Lu X, Liu Z, Tian W; draft manuscript preparation: Lu X, Shang W, Gao Y, Sun S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files, are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primer sequences for real-time fluorescence quantitative PCR.
- Supplementary Table S2 Statistics on clean reads mapping against reference genome.
- Supplementary Table S3 Expression of miRNAs in each sample (TPM).
- Supplementary Table S4 Targeting relationship between miRNA and target genes.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lu X, Liu Z, Feng J, Guo H, Shang W, et al. 2025. MiRNAs play important roles in aroma formation during room temperature storage of 'Binzi' apple (Malus domestica). Fruit Research 5: e024 doi: 10.48130/frures-0025-0014 

MiRNAs play important roles in aroma formation during room temperature storage of 'Binzi' apple (Malus domestica)
- Received: 04 January 2025
- Revised: 09 April 2025
- Accepted: 15 April 2025
- Published online: 13 June 2025
Abstract: MicroRNAs are non-coding RNAs that play a crucial regulatory role in plant growth, development, and environmental stress response. 'Binzi' is an ancient apple cultivar renowned for its unique aroma, however, the mechanisms of miRNAs underlying aroma formation remain largely unknown. In this study, we performed small RNA sequencing to investigate the role of miRNAs during post-harvest storage of 'Binzi' apple. Peel samples from 'Binzi' fruit (namely BZP01 and BZP02) at two storage stages (20 ± 1 °C) were sequenced. A total of 29 DEMs (15 miRNAs uniquely detected at specific storage periods and 14 miRNAs differentially expressed in different storage periods) were predicted to regulate aroma compound biosynthesis in 'Binzi' apple peels. A total of 604 genes were predicted to be targeted by 29 miRNAs. GO enrichment analysis was performed on the target genes, with 'cellular process', 'metabolic process', 'single-organism process', and 'catalytic activity' being the over-enriched biological processes. KEGG analysis showed that three pathways related to aroma synthesis ('fatty acid metabolism', 'biosynthesis of unsaturated fatty acid biosynthesis', and 'carotenoid biosynthesis') were enriched in the top 20 of the KEGG enrichment. We verified the regulatory relationship between miRNAs and their target genes by qRT-PCR. mdm-miR408c was negatively correlated with three target genes (two MdfabDs and one MdfabG) and was involved in the regulation of fatty acids biosynthesis. Furthermore, novel_Chr17_18081 was negatively correlated with MdMFP2 and was involved in the regulation of the fatty acid degradation. Our research indicated that mdm-miR408c, novel_Chr17_18081, and their target genes (MdfabDs, MdfabG, and MdMFP2) were collectively involved in 'Binzi' aroma production.
-
Key words:
- Apple /
- 'Binzi' /
- miRNA /
- Fatty acids biosynthesis /
- Fatty acids degradation /
- Aroma formation.