









-
Floral induction is an important development stage in flowering plants, which significantly impacts subsequent flowering, and fruit formation, fundamentally affecting yield, quality, and economic benefits[1]. Plants regulate flower formation by sensing multiple endogenous and exogenous signals through a complex regulatory network. Different pathways interact and connect with each other, converging the perceived flowering signals on two main flowering pathway integration factors, namely FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (SOC1)[2]. FT-FD complexes further activate the downstream floral meristem (FM) identity genes within the shoot apical meristem (SAM), such as FRUITFUL (FUL)[3], to generate FM on the flank of the inflorescence meristem (IM), which together determine the developmental direction of the inflorescence meristem[4].
Sugar content is a determining factor in floral induction and floral organ development[5]. As a storage organ, the induction and subsequent morphological differentiation of floral meristem are strongly influenced by carbohydrate supply[6]. An increase in carbohydrate levels can be observed during floral induction in plants such as strawberries[7] and Citrus sinensis[8]. In apple (Malus domestica Borkh.), floral induction was proved to be involved in sugar metabolism, biosynthesis, and transport processes[9]. Sucrose spray can significantly increase the size and fresh weight of apple buds, sucrose content in buds and leaves, and flower number[10]. The application of exogenous sugars has been shown to affect the floral transition by influencing flowering-related genes such as FT, SOC1, and LEAFY (LFY)[11]. Genes related to sugar metabolism and transfer have also been shown to affect flower formation, such as the sugars will eventually be exported transporter (SWEET) protein[12] and trehalose-6-phosphate synthase (TPS)[13]. However, the specific role they play in floral induction, changes in sugar content, and whether they are related to floral meristematic genes such as SOC1 are still unclear.
Leaves are the main organs of photosynthesis, the most important source of carbohydrate output, and essential for floral induction[14]. The lack of competition for carbohydrates between vegetative and reproductive organs may lead to failure or delay flower bud and fruit development[15]. As the main transport form of carbohydrates, sucrose is the initiating signal and an important promoter of floral induction[9,16]. During floral induction in plants such as Sinapis alba and Perilla frutescens, sucrose levels in phloem and shoot tips increase rapidly but briefly, indicating that sucrose induces floral transition in meristematic tissues[17]. High sucrose levels can induce early flowering by upregulating FT expression in the phloem of leaves. Meanwhile, the FT signaling pathway can also activate the transcription of sucrose uptake/efflux carriers during the floral transition process[18]. It indicates competition for carbon sources between flower buds and vigorous growing parts, and carbon assimilation and redistribution are crucial in flower formation. However, the role of sugar in blueberry flower bud differentiation has not been reported in detail; thus, the topic deserves further research.
Highbush blueberry plants (Vaccinium corymbosum L.) belong to the Ericaceae family. The small berries the plant produces are popular for their antioxidant properties[19], and are produced from the apical area of annual branches. Proper pruning is necessary for controlling the crowding of the canopy of greenhouse blueberries after harvesting[20]. As an effective pruning technique to relieve apical dominance, short pruning can significantly accelerate the flowering process and fruit yield[21], by increasing sugar transport and promoting the growth of axillary buds[22]. The upright-growing shoots are located at the top of nutrient partitioning and stop growing later. They cannot form flower buds at the front end. If not pruned, it will seriously waste the nutrients of the tree and affect the light of the lower flowering branches[23]. In autumn, only oblique shoots can naturally differentiate into flower buds, the main part of the flowers in the spring of the second year. Previous research has shown that pruning upright-growing autumn shoots can significantly increase the number of flower buds by about four times[24]. For blueberry trees, pruning promotes healthy vegetative growth during the remainder of the growing season[25], and the differentiation of more flower buds for the following spring improves the weight and quality of the fruit for the coming year[26]. Summer pruning of Rabbiteye blueberries (Vaccinium virgatum Ait.) induced the transition of vegetative buds to reproductive buds in the lower part of the cut position, consequently producing fruits in the next summer[27,28] and improved the quantity, quality, and weight of fruits[29]. From 25% to 75%, with the increase of pruning intensity, the single fruit weight gradually increases, but the number of fruits decreases, so moderate pruning is recommended to achieve higher yields[30,31]. Pruning lemon (Citrus limo burm.) and guava (Psidium guajava L.) can result in higher single-branch flowering and fruit production[32,33]. Is the flower bud formation of such shoots related to the adjustment of carbohydrates by pruning? How are they regulated? The regulatory mechanism is still unclear.
Cultivation modes, environmental conditions, hormones, and other conditions may all affect blueberry flower bud formation. In the complex flower bud formation regulation networks, what is the role of sugar? Regarding this issue, the differences in flower bud induction and differentiation processes of autumn blueberry shoots under different treatments, such as pruning, reducing leaf area, and sucrose spray, were compared. The non-structural carbohydrates (NSCs) content, and the expression patterns of genes related to sugar transport and metabolism were analyzed across different tissues. In addition, yeast two-hybrid and transient transformation experiments were combined to analyze the regulatory relationship between sugar-related genes and flowering genes such as FT. The aim is to explore the possible carbohydrate metabolism and transport processes during floral induction, laying the foundation for further research on the regulation mechanism of sugar in blueberry floral induction.
-
The upright growing autumn shoots on the Northern highbush blueberry (Vaccinium corymbosum L.), cultivar 'M7', planted in a greenhouse in the eastern suburbs of Beijing (40°23'42'' N, 116°47'30'' E) were used as plant materials. The natural flower bud differentiation of this blueberry cultivar occurs on the oblique shoots in mid to late September, while the upright growing shoots are still in the vegetative growth period. The average length of the upright growing shoots is about 81.5 cm, and the average diameter of the base is about 5 mm. The sampling is completed in two stages (Fig. 1). The first stage of sampling was conducted from September to October 2018, hereinafter referred to as short pruning treatment (Fig. 1a). Three-hundred and twenty shoots were short pruned, with the top 1/3 of new shoots removed. Another 380 shoots were not pruned as the control group; 60 shoots were picked and brought back as an initial sample on September 13. The labeled shoots were sampled once a week for a total of five times, 60 shoots each, every 20 shoots as a repetition. The non-pruned shoots were evenly divided into three segments from the base: lower, middle, and upper. The upper and middle parts were named the head of non pruned shoot (NP) and the middle of non pruned shoot (MP) group, respectively. The upper part of the pruned shoots had been removed, and the top half of the remaining shoots were named the head of the pruned shoot group (HP). The shoot samples were divided into stem segments, leaves, and axillary buds and stored in a −80 °C refrigerator for later use.
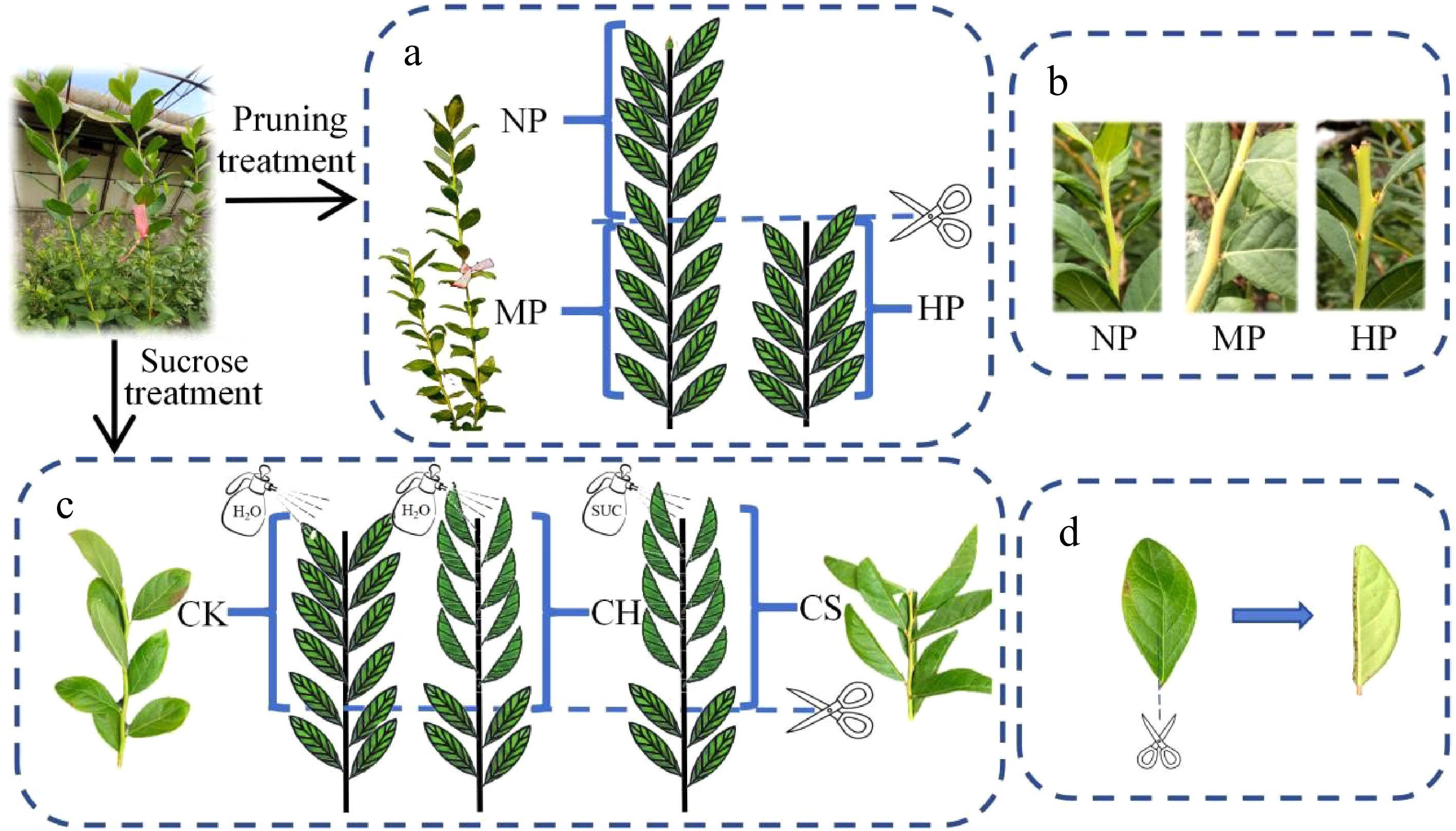
Figure 1.
Schematic diagram of processing method. (a) Schematic diagram of the sample collection sites for the pruning treatment group, where NP, MP, and HP represent the sample collection sites for the uncut and sheared upper parts, respectively. (b) Growth status of NP, MP, and HP shoot on October 6. (c) Schematic diagram of the collection sites for the sugar treatment group, where CK, CH, and CS represent the sample collection sites for the only pruning group, cut half a half and pruning group, and cut half a half and pruning and sucrose treatment group, respectively. (d) Schematic diagram of the half-leaf cutting processing method. The dashed line represents the cutting position.
The second stage sampling was conducted from September to October 2023, hereinafter referred to as sucrose treatment (Fig. 1c). On the basis of the same pruning method as above, half-leaf cutting and sucrose spray were added. On September 25, three groups were designed, namely, only pruning (CK), pruning and cut half a leaf (CH), and pruning and cut half a leaf and sucrose spraying (CS) groups. Cutting off half of the leaves along the leaf midvein of the CH and CS treatment groups while retaining the leaf midvein (Fig. 1c, d). The CK and CH groups were treated with whole plant spraying of water solution until a large number of water droplets drip down, while the CS group was sprayed with the same volume of 1.5% (W/V) sucrose water solution in the same way. Sprays were administered to the entire tree with a low-pressure hand sprayer. The CK treatment group, which only sprayed water without leaf cutting, was set as the control group. Samples were taken five times on days October 01, 05, 09, 14 & 23. 60 shoots were taken as samples each time, and every 20 shoots as a repeat for a total of three repetitions. After rinsing with water three times, the surface was dried with filter paper. The leaf midvein was cut off along both sides of the leaf midvein. The stem segments, axillary buds, non-leaf midvein leaves, and leaf midvein were stored separately. Five axillary buds at the morphological upper end of each shoot were taken to calculate the flower bud rate. Ten bud samples from each sampling period were randomly selected for routine paraffin sectioning. The daily sunshine duration and the lowest and highest temperatures (Supplementary Fig. S1) are attached.
Morphological changes and anatomical observation of axillary buds
-
On October 23, the longitudinal and transverse diameters of axillary buds on each shoot were measured using an electronic caliper. The axillary bud samples were preserved with FAA solution (7.5:1:1:1 ratio of 50% ethanol : formalin : glacial acetic acid : glycerol). The dehydration process was a range of concentrations of ethanol immersion (70% for 2 h, 80% for 1.5 h, 95% for 1 h, 100% for 1 h, and 100% for 0.5 h). Then the Turpentine oil (TO) was used to replace the ethanol as follows: TO : Ethanol (1:2) for 1 h; TO : Ethanol (1:1) for 1 h; TO : Ethanol (2:1) for 1 h; TO, changed twice, for 1 h and each; paraffin-saturated TO solution at 37 °C for 24 h; paraffin wax (58−60 °C), changed eight for 30 min and each. Then, tissues were embedded in paraffin blocks. The embedded samples were trimmed before sectioning. The trimmed samples were mounted on a microtome and cut into thin slices, each with a thickness of 10 µm. After fixation, dehydration, wax dipping, embedding, section, dewaxing, dyeing, and other steps of paraffin section[34], the sections were stained using the Salarbio Plant Tissue Staining Kit (PAS-N method) (G4809; Salarbio). After staining, the cell walls of the sections appeared purple-red; starch granules were purple-red particles. The bud morphology of the sections was observed under a biological microscope (DM2500) and manually photographed.
Determination of non-structural carbohydrate content
-
The sucrose, fructose, glucose, and water-soluble sugars of the two treatment groups were measured, and the starch contents were additionally measured in the sucrose treatment group. The specific method is as follows: Accurately weigh 0.1 g of fresh plant sample, add 4 mL of distilled water, water bath at 100 °C for 20 min, invert and mix three times during this period, centrifuge at 4,000 × g for 10 min, transfer the supernatant and add 2 mL of distilled water to the precipitate, repeat the above steps twice, make up the supernatant to 10 mL for carbohydrate content determination, and dry the starch in a cool place for starch content determination. The water-soluble sugar (WSS) content was determined using the anthrone colorimetric method[35], hereinafter referred to as soluble sugar, while the sucrose and fructose content was determined using the hydrochloric acid resorcinol colorimetric method[36]. The glucose content was determined using the glucose oxidase method (specific steps refer to the instruction manual A154-1-1 of the Nanjing-built glucose test kit), and the starch content was determined using the Boxbio starch content detection kit.
Total RNA extraction and cDNA synthesis
-
Total RNA of the sucrose treatment group was extracted from stem, leaves, leaf midvein, and axillary buds using the Vazyme Universal Plant Total RNA Isolation Kit. Reverse transcription was accomplished using the Vazyme ALL in one RT SuperMix Perfect for qPCR Kit for subsequent qRT-PCR experiments and sequence cloning[37].
qRT-PCR
-
The primers were designed using PrimerPremier 5.0 software and synthesized by Ruibio BioTech Co., Ltd. (Beijing, China). Genes were divided into three categories: (1) Genes in the flowering pathways, including VcFT, VcSOC1, VcFUL, and VcSVP (SHORT VEGETATIVE PHASE). (2) Genes related to sugar transport, including VcSWEET1, VcSWEET16, VcSTP (Sugar Transporter Proteins), and VcSUC (SUCROSE TRANSPORTER). (3) Genes related to sugar metabolism, including VcAMY (α-amylase gene family), VcINV (invertase gene family), VcTPS1, VcTPS9, VcTPS11, and VcSnRK1 (sucrose nonfermenting-1-related protein kinase) (Primers are listed in Supplementary Table S1). According to StepOnePlus Systems (Thermo Fisher, China), the relative expression level of different genes was calculated using a double standard curve. VcUBC28 was used as a reference gene[37]. The 2−ΔΔCᴛ method was used to analyze the relevant gene expression data.
Yeast two-hybrid system
-
From the Blueberry Genome Database (
www.vaccinium.org ), the CDS sequence of the target gene was retrieved, and a seamless cloning method was used to clone the VcFUL, VcSOC1, VcFT, VcSVP, VcSUC, VcTPS1, and VcAMY genes into pGADT7 and pGBKT7 vectors. The fusion pGADT7 and pGBKT7 vectors were then transformed into the Y2HGold yeast strain (YC1002S; Weidi). The transcriptional auto-activation activity was detected on SD/-Trp/-Leu plates. The pGADT7-gene and pGBKT7-gene transformed colonies were tested for possible interactions on SD/-Leu/-Trp/-His/-Ade plates. Primers are listed in Supplementary Table S1.Transient transformation in blueberry
-
The full-length VcTPS1 gene was cloned into the pSuper1300-GFP vector, sequenced and validated, and then transformed into Agrobacterium tumefaciens. Positive colonies were selected and cultured in liquid YEB medium containing 50 μg/mL Kana (kanamycin), 20 μg/L Rif (Rifampicin), and 200 μg/L As (Acetosyringone) at 37 °C and 200 rpm on a shaking incubator with a rotational radius of 10 cm. After centrifugation, they were resuspended in a liquid WPM medium containing As and adjusted to OD600 at around 0.6. These were kept in the dark for 3 h, and then the blueberry tissue culture seedlings in the bacterial solution were vacuum-filtered for 20 min. After drying the surface bacterial solution, they were wrapped in plastic wrap and incubated in the dark for 24 h. Subsequently, they were cultured for 5 d under 16 h of light and 8 h of darkness. After ensuring that the plants were healthy and free from physical damage, RNA was extracted from the leaves. The qRT-PCR method was used to test the gene delivery efficiency. The pSuper1300-GFP empty vector of Agrobacterium tumefaciens for infection was a negative control group, following the method reference[38] with slight modifications.
Data analysis
-
The experiment was conducted with triple biological replicates for each parameter. The significant difference was analyzed using SPSS 26; relevant graphs were drawn using Origin and Prism 10.1.2 software. Microsoft Excel was used to organize initial data and conduct preliminary analysis.
-
To investigate whether short pruning promotes floral induction of autumn shoots is related to sugar, the sugar content of each part of the pruning treatment group was measured. After pruning, the soluble sugar content increased in buds and leaves, while it decreased in the stem compared to those without pruning (Fig. 2j−l). Pruning increased the sucrose content in leaves and the glucose content in buds of the HP (the head of the pruned shoot) group but significantly decreased the sucrose and glucose content in the stem (Fig. 2a−f) from September 13 to September 28. According to previous research[24]; the floral induction period of the strong upright blueberry autumn shoots after pruning starts from September 13 to 28, and the formation period of flower organs starts from September 28 to October 20. During the floral induction period, the sucrose content in leaves and the sucrose and soluble sugar contents in buds increase from September 13 to 21. The glucose content in buds increases from September 21 to September 28, while they significantly decrease in the stem. The sucrose content changes in various parts earlier than those in monosaccharides. The sucrose content in leaves and buds of the HP group increased by 42.39% and 87.41% on September 21, compared to September 13, respectively (Fig. 2a & c). However, in the stem, it significantly decreases by 54.55% (Fig. 2b). Thus, sucrose seems to be the main trigger for the formation of flower buds in the upper parts of the strong upright blueberry autumn shoots after pruning.
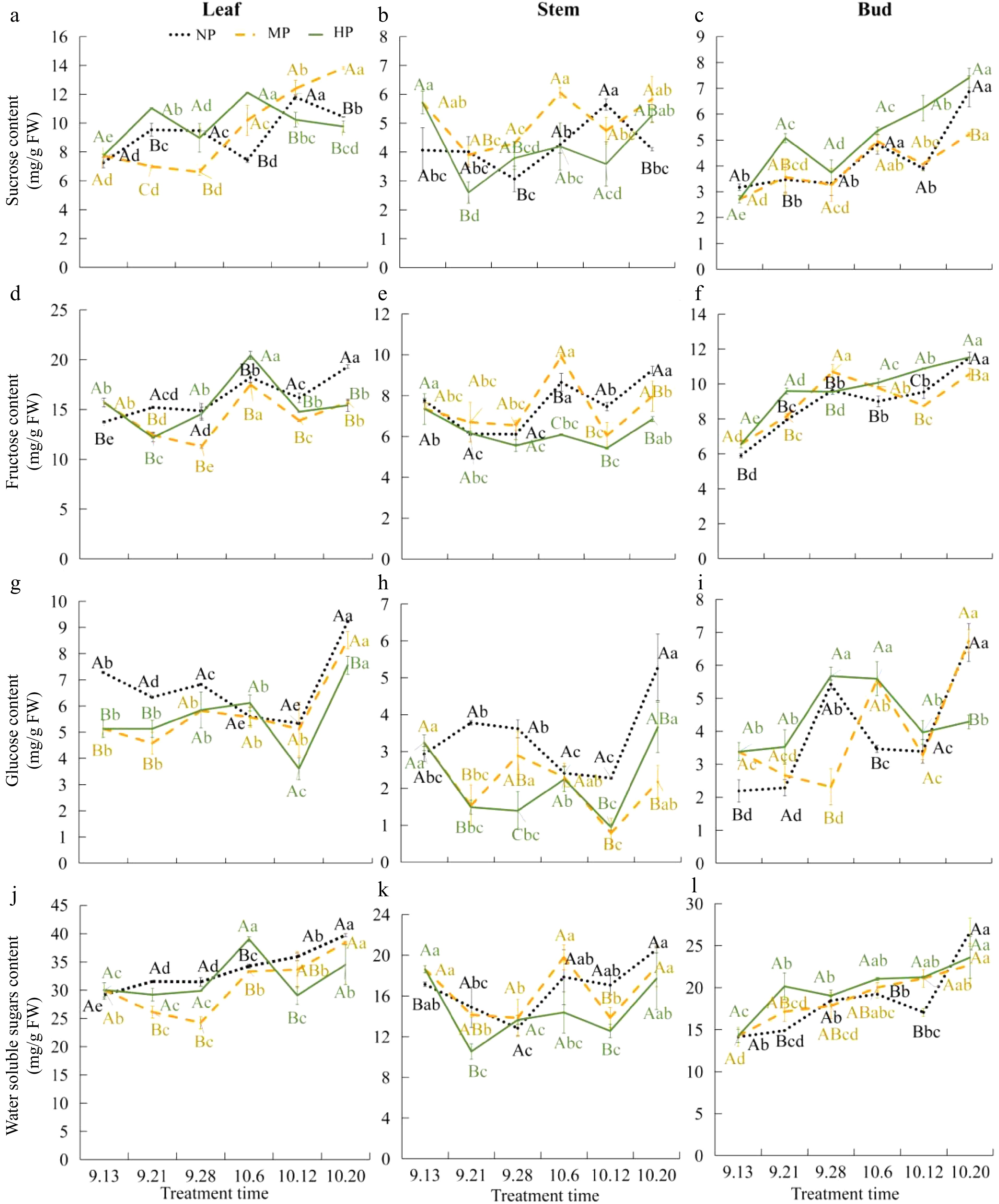
Figure 2.
Sugar content in different parts of autumn short 14 pruning. (a)−(c) Sucrose content in leaves, stems, and buds. (d)−(f) Fructose content in leaves, stems, and buds. (g)−(i) Glucose content in leaves, stems, and buds. (j)−(l) Water soluble sugar content in leaves, stems, and buds, respectively. NP indicates the head of the non-pruned shoot, MP indicates the middle of the non-pruned shoot, and HP ndicates the head of the pruned shoot. Duncan's multiple range test was used for significance analysis. Different lowercase letters indicate significant differences at the p < 0.05 level between the same treatment and different treatments, while different uppercase letters indicate significant differences at the p < 0.05 level between different treatments at the same time. The same applies below.
Cutting leaves and sucrose supplements affect flower bud differentiation process
-
To determine the effect of the sucrose treatment on flower bud differentiation process of axillary buds, the morphological and anatomical structure of the buds were observed and statistically analyzed. The longitudinal and transverse diameters of axillary buds in the CK (only pruning) group were significantly higher than those in the CH (pruning and cutting half a leaf) and CS (pruning and cutting half a leaf and sucrose treatment) groups. Significant swelling of axillary buds was observed in the CK and CS groups on October 5 and 14, respectively (Fig. 3a).
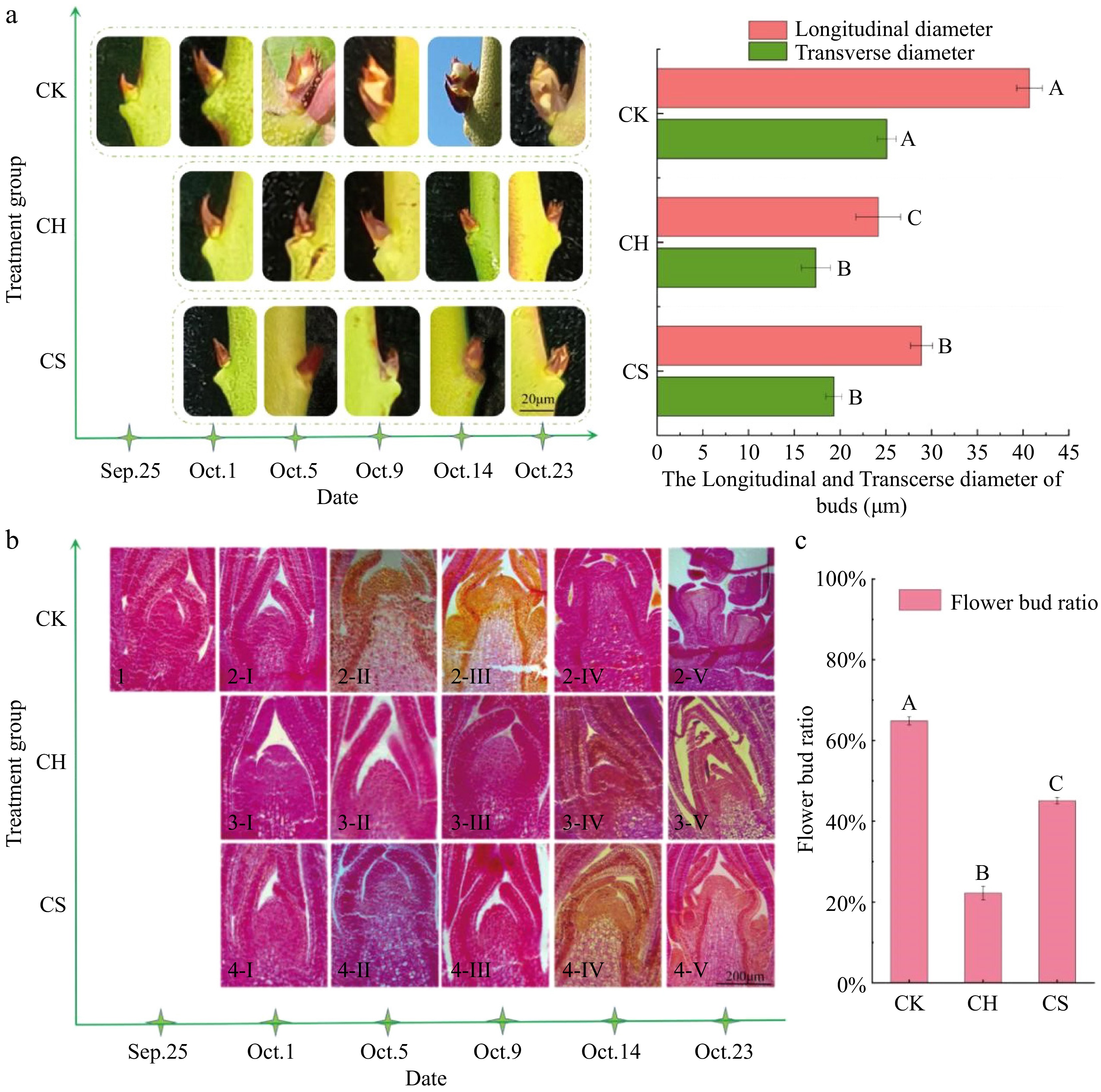
Figure 3.
Observation of bud morphology and flower bud ratio. (a) The directly observed morphology of axillary buds and the transverse and longitudinal diameters of buds on October 23. (b) The morphology of paraffin sections. (b-1) The buds on September 25. (b-2-I−V)−(2-I−V) The buds from October 1 to 23 of CK, CH, and CS, respectively. (c) The proportion of the total number of flower buds in each treatment to the total number of axillary buds. Different uppercase letters indicate significant differences at the p < 0.05 level between different treatments. CK represents the only pruning group, CH represents the pruning and cutting half a leaf group, and CS represents the pruning and cutting half a leaf and sucrose treatment group.
On September 25, the growth point of axillary buds on autumn shoots was still tightly held by bud scales, and the growth point was conical with no signs of differentiation (Fig. 3b-1). The floral induction period of the CK group started from September 25 to October 01. On October 5, the scales of axillary buds tightly embracing the growth point began to loosen, the growth point increased, gradually became round at the top, and the cells were larger (Fig. 3b-2-II). The buds developed to the initial differentiation stage, the number of slices in this stage accounted for 80% of the total. On October 23, the buds in the CK group differentiated into two small protrusions at the intersection of the inner side of the sepal primordia (Fig. 3b-2-V), developed to the stage of petal primordia differentiation. The buds of the CH group showed no signs of differentiation until October 23 (Fig. 3b-3-V). As shown in Fig. 3b-4-IV, the buds of the CS group developed the initial differentiation stage on October 14, during which the number of slices accounts for 60% of the total, about 9 d later than the CK group. A large number of purple-red starch granules were observed in both the CK and CS groups during the initial differentiation stage and the inflorescence primordium differentiation stage (Fig. 3b-2-II−2-III & 4-IV−4-V). The proportion of flower buds in the pruning treatment group is 64% (Fig. 3c). Cutting off half a leaf significantly affects the number of flower buds formed in the lower part of the cut. Exogenous sucrose spray remedied some of the flower bud formation but still could not recover to the level of the CK group.
Sucrose in leaves promotes floral induction
-
To confirm whether the effects of leaf pruning and exogenous sucrose treatment on floral induction were achieved by affecting NSCs contents, especially sucrose, the NSCs content in various parts of the sucrose treatment group was measured.
Sucrose increases in leaves and buds during the floral induction
-
Exogenous sucrose spray significantly increases the sucrose and starch content in leaves and buds of the CS (pruning and cutting half a leaf and sucrose treatment) group, as well as the total water-soluble sugar content in various parts (Fig. 4a−d). The sucrose content in buds of the CK (only pruning) group was higher than that in the CH (pruning and cutting half a leaf) group at all stages with a similar trend of change and reached a peak of 3.73mg/g FW at the initial differentiation stage on October 5, second only to the peak (Fig. 4d). In the CS group, and it reached a peak of 5.37mg/g FW at the initial differentiation stage on October 14 (Fig. 4d). Sucrose content in leaf midvein of the CK group reached its highest level of 3.57mg/g FW during the floral induction period, while in the CH group it remained at a relatively stable level (Fig. 4b). During the floral induction period, the sucrose content in CK and CS leaves and buds significantly increased, while the sucrose content in stem significantly decreased (Fig. 4d).
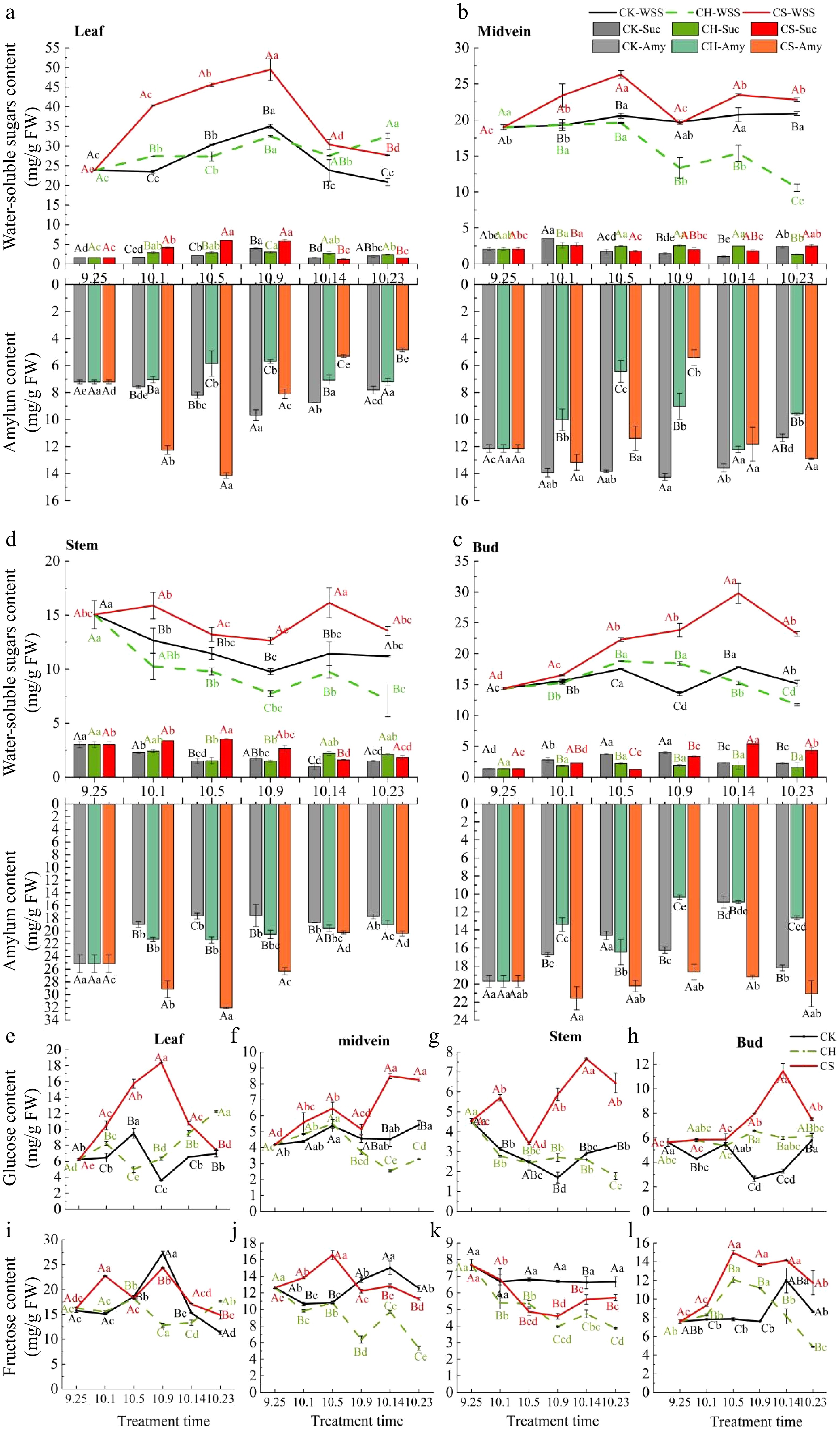
Figure 4.
NSCs content in various parts of the sucrose treatment 36 group. (a)−(d) Sucrose, water-soluble sugar (WSS), and amylum (Amy) content of leaves, midvein, stem, and buds. The X-axis represents the processing date. Above the X-axis of each graph are WSS (line graphs) and sucrose contents (bar graphs), and below are amylum contents (bar graphs). (e)−(h) Glucose content in leaves, midvein, stem, and buds, respectively. (i)−(l) Fructose content in leaves, midvein, stem, and buds, respectively. CK represents the only pruning group, CH represents the pruning and cutting half a leaf group, and CS represents the pruning and cutting half a leaf and sucrose treatment group.
Fructose and glucose change during the morphological differentiation period
-
After treatment, the fructose content in buds of the CH and CS groups rapidly increased and reached the peak on October 5 at 12.09 mg/g FW and 14.96 mg/g FW, respectively. Subsequently, it decreased rapidly in the CH group, while in the CS group it remained at a certain level (Fig. 4l). There was only a brief increase on October 14 in CK group, and it remained at a certain level for the rest of the time (Fig. 4l). Fructose content in leaves of CK and CS groups significantly increased during the floral induction period and then decreased during morphological differentiation period (Fig. 4i), while showing a stable level in leaves and leaf midvein of CH group (Fig. 4i, j). Sucrose spray increased the fructose content of leaves. The fructose content in buds increased and then decreased during the morphological differentiation period. The glucose content in leaves of both CK and CS groups increased during the floral induction period and decreased during the initial morphological differentiation period in buds, with a stable level in the CH group (Fig. 4e & h). The decrease of glucose in leaves and buds of the CK group is always one to two stages earlier than fructose (Fig. 4d−i). It indicates that the utilization degree or sequence of the two monosaccharides in buds is different during the differentiation stage.
Soluble sugar decreases in the stem during the floral induction
-
The soluble sugar content in leaves, leaf midvein, and buds of the CK group increased significantly during the floral induction period, while it decreased in the stem (Fig. 4a−d). After entering the stage of morphological differentiation, the soluble sugar content in buds fluctuated at a certain level, but in leaves significantly decreased (Fig. 4a & d). The changes in the leaf midvein and stem may indicate an increased degree of sugar transport at this time. The soluble sugar content in CK and CS groups show similar trends of change. Although the soluble sugar content in leaves and stems of the CH group also significantly increases after treatment, it is not as significant as that in the CK and CS groups (Fig. 4a, c & n).
Starch content in leaves increases during the floral induction
-
During the floral induction period, the starch content in the leaf midvein of CK and CS groups increases, while in leaves and leaf midvein, it slightly increases and then decreases (Fig. 4b−d). In the CH group, the starch content in various parts first decreases and then stabilizes at a certain level. After entering the stage of morphological differentiation, the starch content in buds fluctuates, while in leaves and leaf midvein, it slightly increases and then decreases, and in stem, it remains at a certain level (Fig. 4b−d). The starch stored in the stem may be an important source of carbohydrates required for this flower bud formation. During the morphological differentiation period, intense starch synthesis and metabolic reactions occur in the buds.
VcFT upregulates expression during floral induction and is induced by sucrose
-
Flowering-related genes are objective indicators that reflect the process of flower bud differentiation. Sucrose treatment significantly increases the relative expression level of VcFT in various parts and the level of VcFUL and VcSOC1 in buds of the CS group, while in the CH group, there was no significant increase (Fig. 5a−c, f & i). The relative expression level of VcFT in both buds and leaf midvein of CK and CS groups increased during the floral induction period, reaching their peak on October 5 and 14, respectively (Fig. 5b, c). VcFUL was mainly expressed in the CK group, whose expression level increases during the floral induction period in buds and morphological differentiation period in buds and leaf midvein (Fig. 5g−i). The relative expression level of VcSOC1 in CK and CS group buds increases during the initial differentiation period, while in the leaves and leaf midvein, it mainly plays a role during the morphological differentiation period (Fig. 5d−f). It indicates that during the floral induction and differentiation period, VcFT in the leaf midvein and VcSOC1, VcFUL, and VcFT in the buds may play a major role. Sucrose treatment reduces the relative expression level of VcSVP in various parts of the CS group on October 1 (Fig. 5j−l). The relative expression level of VcSVP in buds, leaves and leaf midvein of the CK group decreases during the floral induction period, then increases during the morphological differentiation period, similar to the expression level of VcFUL(Fig. 5j−l).
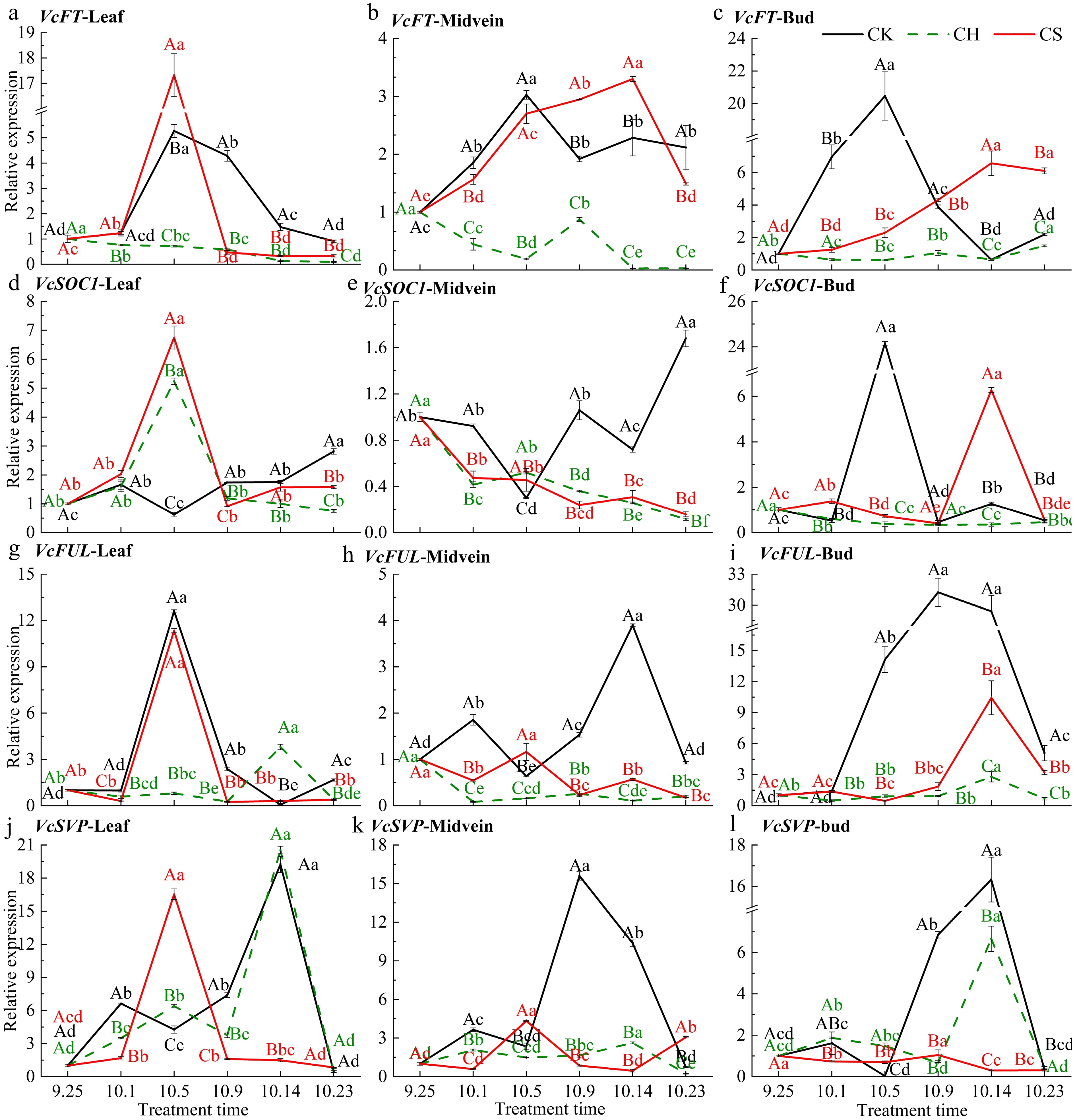
Figure 5.
Relative expression level of genes related to flowering pathways in various parts. (a)−(c) Level of VcFT in leaf, midvein and bud, respectively. (d)−(l) Relative expression level of VcSOC1, VcFUL, and VcSVP in the above-mentioned parts respectively. CK represents the only pruning group, CH represents the pruning and cutting half a leaf group, and CS represents the pruning and cutting half a leaf and sucrose treatment group. Duncan's multiple range test was used for significance analysis. Different lowercase letters indicate significant differences at the p < 0.05 level between the same treatment and different treatments, while different uppercase letters indicate significant differences at the p < 0.05 level between different treatments at the same time. The same applies below.
Sugar metabolism-related genes participate in flower regulation
-
To clarify the expression characteristics of carbohydrate metabolism-related genes during the flower regulation process and their corresponding effects on exogenous sucrose, the expression levels of VcTPS and other genes were detected. Exogenous sucrose increases the expression level of VcSNRK1 in the CS (pruning and cutting half a leaf and sucrose treatment) group (Fig. 6a, b & d). The relative expression level of VcSNRK1 in the buds of CK (only pruning) and CS groups significantly increases at the initial morphological differentiation stage on October 5 and 14, respectively (Fig. 6c), indicating that VcSNRK1 in the buds has a promoting effect of the morphological differentiation. T6P is a precursor of trehalose and can serve as an indicator of plant sugar status, synthesized by TPS catalysis. Sucrose treatment significantly increases the relative expression level of VcTPS1 and VcTPS11 in CS group buds, while in buds and leaves, the level of VcTPS9 decreases (Fig. 6e, g−i & k). VcTPS1 is mainly upregulated during the induction and morphological differentiation period in buds, while VcTPS9 is mainly upregulated in buds and leaves of the CH (pruning and cutting half a leaf) group (Fig. 6e, g & h). The response of VcTPS11 to exogenous sucrose is the most significant (Fig. 6i−k), whose level in buds and leaves of the CK group increases from October 5 to 23, the morphological differentiation period (Fig. 6i & k). It indicates that VcTPS11 may play an important role in promoting flower bud morphological differentiation. VcINV can irreversibly convert sucrose into glucose and fructose for plant cell utilization. The relative expression levels of VcINV in the buds of both CK and CS groups significantly increase at the initial differentiation stage, following the same trend as VcSUC (Fig. 6m−o). The expression level of VcINV in the leaf midvein of the CK group increases continuously with the process of flower bud differentiation, and it is higher than that of CH and CS groups (Fig. 6n). VcAMY is upregulated during both the floral induction and morphological differentiation periods in the CK and CS groups (Fig. 6l).
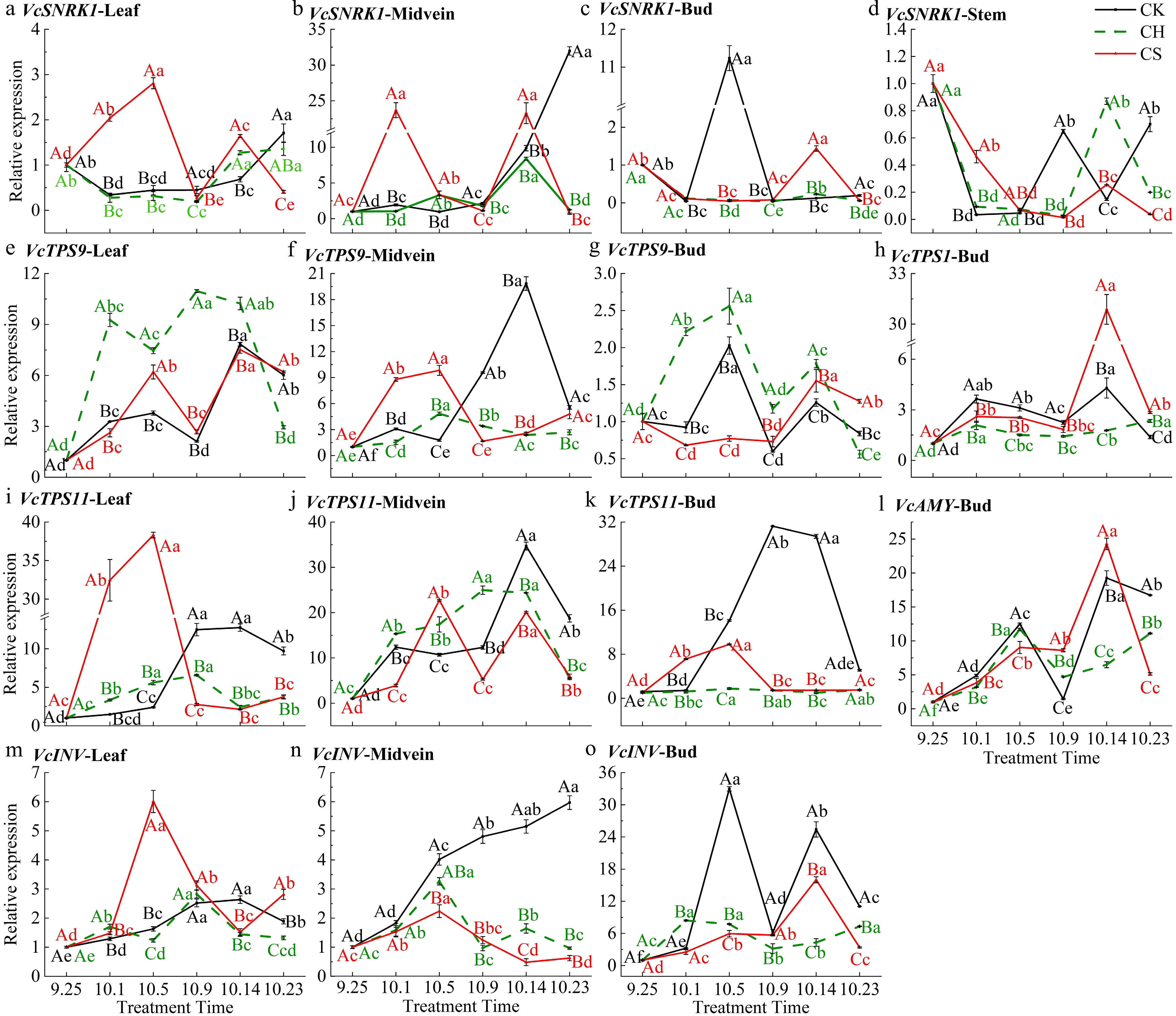
Figure 6.
Relative expression level of carbohydrate metabolism-related genes in various parts. (a)−(d) Relative expression level of VcSNRK1 in leaf, midvein, bud, and stem. (e)−(h) Relative expression level of VcTPS9 in lea, midvein, bud, stem, and VcTPS1 in bud. (i)−(l) Relative expression level of VcTPS11 in leaf, midvein, bud, and VcAMY in bud. (m)−(o) Relative expression level of VcINV in leaf, midvein, and bud. CK represents the only pruning group, CH represents the pruning and cutting half a leaf group, and CS represents the pruning and cutting half a leaf and sucrose treatment group.
Sugar transport-related genes upregulate during morphological differentiation
-
Sugar transport-related genes can reflect the sugar transport status in different parts of the plant. Sucrose treatment significantly increases the relative expression level of VcSWEET1 in various parts of the CS (pruning and cutting half a leaf and sucrose treatment) group on October 1 (Fig. 7a−d). VcSWEET1 was mainly differentially expressed in the leaf midvein and stem at the initial differentiation stage (Fig. 7b, c). In the CK (only pruning) group, the relative expression level significantly increases on October 5 and 9 in the leaf midvein (Fig. 7b), while in the CS group, the level significantly increases on October 1 and 14 of stem (Fig. 7c). VcSWEET16 is homologous to AtSWEET16 and is downregulated in buds and leaves during the morphological differentiation period, compared to the CH (pruning and cutting half a leaf) group (Fig. 7e−h). The expression level of VcSUC in CS group leaves is higher than that in CK and CH during the period from September 25−October 5 (Fig. 7m). In the buds, it significantly increases under all three treatments but then decreases to a lower level in the CH group on October 1 (Fig. 7). The expression level of VcSUC in the buds of CK and CS groups reaches two peaks on October 5 and 14, respectively (Fig. 7o), may be related to the increase in sucrose content in the buds of CK and CS groups. The relative expression level of VcSUC in the leaf midvein of the CK group significantly increases from October 1 to 23 (Fig. 7n). The CS group maintains a lower relative expression level after treatment, which may indicate the efficiency of sucrose transport in the leaf midveins. VcSTP is a type of sugar transporter protein in the MST family that has the ability to take up hexose, highly expressed in the buds and leaf midvein of the CK group on October 5, consistent with the trend of the relative expression level of VcINV in the buds of the CK group (Figs 6i & 7j−l).
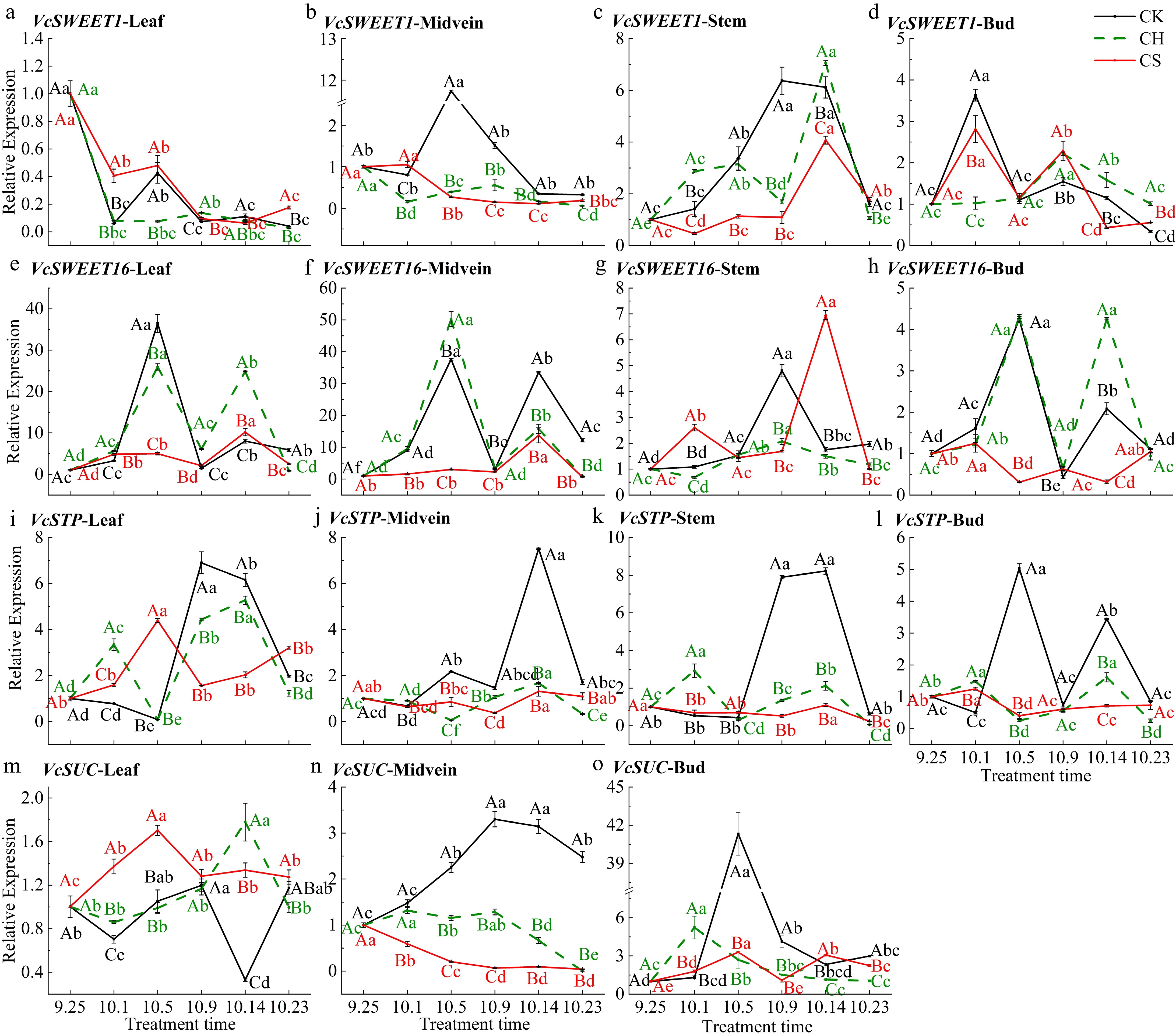
Figure 7.
Relative expression level of carbohydrate transport-related genes in various parts. (a), (e), (i) and (m) Relative expression level of VcSWEET1, VcSWEET16, VcSTP, and VcSUC in leaf. (b), (f), (j) and (n) Relative expression level of VcSWEET1, VcSWEET16, VcSTP, and VcSUC in midvein.(c), (g) and (k) Relative expression level of VcSWEET1, VcSWEET16, VcSTP in the stem. (d), (h), (l) and (o) Relative expression level of VcSWEET1, VcSWEET16, VcSTP, and VcSUC in bud, respectively. CK represents the only pruning group, CH represents the pruning and cutting half a leaf group, and CS represents the pruning and cutting half a leaf and sucrose treatment group.
VcSOC1 interacts with VcTPS1 protein
-
To confirm whether sugar-related proteins regulate floral through physical interactions with flowering proteins, a yeast two-hybrid experiment was conducted. The yeast strains with VcFUL, VcSOC1, VcFT, VcSVP, VcTPS1, VcSUC, and VcAMY have no transcriptional self-activation activity (Fig. 8a). The flowering proteins VcSOC1-VcFUL, VcFUL-VcFT, VcSVP-VcSOC1, VcSOC1-VcFT, and VcFT-VcSVP yeast strains can grow normally on the SD/-His/-Leu/-Trp medium, but cannot grow on the SD/-Ade/-His/-Leu/-Trp medium. It indicates that there are interactions between them, but they are weak. The verification results of VcSOC1-VcSUC, VcSOC1-VcAMY, VcSOC1-VcTPS1, and VcFT-VcSUC are the same as above, indicating that weak interactions exist (Fig. 8b).

Figure 8.
Self-activation detection and interaction verification of the yeast two-hybrid. Yeast two-hybrid assay revealing the interaction of flowering proteins with VcTPS1, VcSUC and VcAMY proteins. (a) Self-activation detection. (b) The interaction verification. SD/-Trp-leu/, SD/-His-Trp-Leu/, and SD/-Ade-His-Trp-Leu/ are DDO, TDO and QDO media, respectively. This experiment was replicated three times.
VcTPS1 promotes the expression of VcSOC1 in blueberries
-
The transient transformation experiment was conducted to investigate whether VcTPS1 has the ability to affect the expression of flowering genes and other sugar-related genes in blueberries. The growth status of blueberries is shown in Supplementary Fig. S2. Based on the hygromycin tag on the vector, the effectiveness of transformation by agarose gel electrophoresis was verified (Supplementary Fig. S3). The relative expression level of VcTPS1 in transformed plants is higher than that in the control group (Fig. 9a). Except for VcSNRK1, whose relative expression level decreases after transformation (Fig. 9c), the relative expression level of VcSOC1, VcSUC, VcINV, and VcAMY in the transformed plants significantly increase (Fig. 9b & d−f). It suggests that VcTPS1 may have the ability to promote starch metabolism, sugar transport, and floral processes through the flowering gene VcSOC1, as well as carbohydrate metabolism and transfer genes.
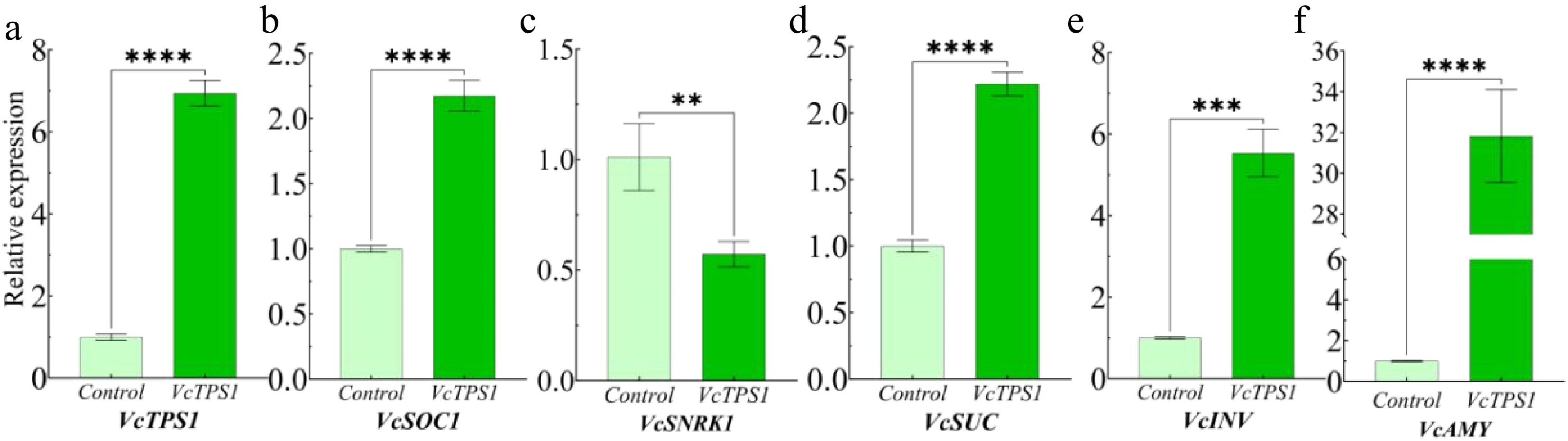
Figure 9.
Transient conversion expression analysis. (a)−(f) Relative expression level of VcTPS1, VcSOC1, VcSNRK1, VcSUC, VcINV, and VcAMY genes in blueberries transiently transformed with VcTPS1. ** represents very significant difference (p ≤ 0.01), *** represents extremely significant difference (p ≤ 0.001), **** represents extremely significant difference (p ≤ 0.0001).
-
Sugar is the initial signal for floral induction[39]. During the transition from vegetative growth to reproductive growth in plants, the sucrose level in the phloem and shoot tips undergo a rapid but brief increase phase[40]. Studies in Sophora tonkinensis Gagnep[34] and strawberries (Fragaria × ananassa cv. 'Kordestan') also show that soluble carbohydrate content may play a decisive role in floral induction[7]; that is, an increase in soluble sugar content within the bud is necessary to achieve a transition towards reproductive growth[41]. However, the existence of apical of the stem inhibits axillary bud outgrowth during the vegetative growth period[42], which can be eliminated by the plant decapitated[43]. Following pruning and during the floral induction period, the glucose, sucrose, and water-soluble sugar content in the buds significantly increases, and the soluble sugar and starch content in the stem significantly decreases (Fig. 4). Short-pruning may promote the transfer of non-structural carbohydrates from stem to buds, the stem may be important sources of carbohydrate elevation in buds. The increase in carbon supply to axillary buds can occur a few hours after treatment[22,44]. The starch synthesized during the inflorescence development stage of the Oncidium orchid is ultimately degraded during the flower development stage[45]. Anatomical observation shows that a large number of starch granules accumulate in the buds during the initial stage of flower bud differentiation (Fig. 3). It indicates that, in addition to the transportation from source to sink, the possible short-distance transport in buds is also necessary to meet the nutritional needs of the bud differentiation process, and intense carbohydrate metabolism occurs in the buds[46].
Flower bud differentiation is accompanied by intense carbohydrate metabolism
-
Strong starch and sucrose metabolism reactions occur within the buds during floral induction and morphological differentiation. The dynamic changes of carbohydrates are important signals for plant floral transition, which cannot be separated from the transmission of signals and the synthesis of related metabolic enzymes. Studies have shown that sucrose/starch metabolism is involved in the regulation of litchi (Litchi chinensis Sonn.) flower bud differentiation[47]. SnRK1 plays a core role in regulating cellular responses to endogenous energy and carbon states[48]. In peach (Prunus persica), the activity of SnRK1 can be promoted by appropriate concentrations of sucrose[49] and participate in the upregulation regulation of genes involved in the starch biosynthesis pathway[50]. This is consistent with the significant increase in starch content observed in the research, where the expression level of VcSNRK1 rapidly increases in leaves and leaf midvein after sucrose treatment. In rice (Oryza sativa), low sucrose content induces an increase in OsSnRK1a expression and facilitates starch degradation and sugar transport in the phloem[51]. Upregulation of VcSNRK1, VcINV, and VcAMY was observed during the bud morphological differentiation period following short pruning (Fig. 6). At the same time, the starch content in the buds also shows a lower level, which may be related to the increase in soluble sugars content in the buds. The expression levels of these genes exhibited dynamic changes, with peak expression observed on October 5 and 14. This can further prove that the changes in carbohydrate content during floral induction and differentiation are dynamic; that is, the accumulation of nutrients may have multiple processes, which respectively meet the requirements of floral induction and differentiation. After treatment, the expression of VcSNRK1 in the leaves and leaf midvein of the CS group is significantly upregulated, followed by a decrease, consistent with the trend of changes in sucrose and starch content in the leaves and leaf midvein during this period (Fig. 4). This suggests that the addition of exogenous sucrose may promote the expression of SNRK1[52], thereby promoting the outward transport from the source and the carbohydrates accumulation in buds. The content of sucrose or starch is closely related to the expression of VcSNRK1. T6P, as an important sugar signaling substance, is synthesized by TPS class I protein catalysis, which is opposite to the signal emitted by SnRK1 activation[53]. TPS I has been shown to promote floral induction[54], while TPS II protein can also inhibit SnRK1 kinase activity and nuclear SnRK1 signaling[52], indicating that TPS is involved in sugar-controlled flowering induction[13]. The expression of VcTPS1 was also significantly upregulated during the floral induction and initial morphological differentiation period (Fig. 6), which was consistent with the changes in sucrose content in the buds during this period. High levels of sucrose simultaneously meet the energy and signaling requirements for floral induction[17,55]. Based on the transient transformation experiment in blueberries, the increase in VcTPS1 expression level is speculated to promote the expression of VcSOC1, which corresponds to the increase in VcSOC1 expression level in buds during the initial morphological differentiation period, and the two proteins may then form a protein complex to promote flower formation. VcTPS11 is upregulated during the morphological differentiation period in leaves, leaf midvein, and buds, indicating that VcTPS11 may be involved in the morphological differentiation process of flower buds (Fig. 6).
Short pruning promotes sugar efflux from the stem and midvein
-
Stem may be an important source of carbohydrates of axillary bud during the induction and morphological differentiation period. The apical dominance controls the direction of sugar movement, released after removing the top bud. Leaves are the source of induction signals that promote axillary bud germination, and the application of exogenous sucrose promotes bud germination[56]. Exogenous sucrose can promote the loading and unloading of sucrose in the phloem of mature tomato (Solanum lycopersicum L.) leaves[57]. The expression level of VcSNRK1 in the stem of the CS (pruning and cut half a leaf and sucrose spraying) group was higher than that of the CH (pruning and cut half a leaf) group after sucrose treatment (Fig. 6), indicating a possible increase in stem, leaf and leaf midvein carbon output at this time. Studies have also shown that SNRK1 is necessary for the sheath-to-panicle transport of NSCs in rice[51]. Sucrose is the main form of carbohydrate transport, and intercellular transport at different tissue junctions is believed to occur through extracellular vesicles[58]. Activation of extracellular sugar supply promotes the growth of axillary buds[59], and the upregulation of VcINV, VcSWEET, and VcSUC expression level in buds of CK (only pruning) and CS groups seems to confirm the existence of this transport mode. SWEET is a type of efflux carrier in phloem thin-walled cells[60]. AtSWEET1 has been shown to be localized to the vacuolar membrane[61]. In this research, VcSWEET1 was mainly upregulated in buds during floral induction and the leaf midvein and stem during morphological differentiation (Fig. 7). This suggests that pruning may promote the efflux of carbohydrate from the lower stem part after pruning by affecting the expression level of VcSWEET1 and VcSNRK1. VcSEWEET16 does not seem to play a role in the regulation of flower bud differentiation, but the relative expression level of various parts in the CS group decreases after spraying sucrose, indicating that the application of exogenous sucrose may have inhibited the efflux of sugars from vacuoles (Fig. 7). SUC sucrose transporter protein absorbs sugars from the thin wall tissue of the phloem and absorbs them into the SE/CC complex, playing a role in phloem loading and unloading together with SWEET[62]. In Arabidopsis, exogenous sucrose repressed phloem loading activity in leaves by inhibiting SWEET expression but promoting SUC expression[63], which is consistent with the results. These sucrose transporters have different physiological functions in different tissues[64]. In this experiment, VcSUC mainly plays a role in leaf midvein and bud during the induction and initial differentiation period, indicating the occurrence of sugar transport at this moment. It is also consistent with the research results in lily (Lilium spp.), that the expression of FT may be positively influenced by SUC[65]. In addition to the SUC-mediated sucrose transport pathway, sucrose in the plastid region transported through the phloem is hydrolyzed by cell wall convertase (INV). The resulting hexose is then transported to sink tissue by sugar transporter proteins (STPs)[66]. The relative expression level of VcSTP in the buds of the CK group increases at the initial differentiation and sepal differentiation primordia, similar to the expression trend of VcINV mentioned earlier. This suggests that the buds may have an increased demand for monosaccharides at this time or that the sugar transport mode may have changed (Fig. 7).
Expression characteristics of flowering genes during flower bud differentiation
-
FT and SOC1 are considered integrated factors of multiple flowering regulatory pathways, promoting the floral transition in Arabidopsis[2,67]. Sucrose mainly plays a role in the leaf phloem to enhance the production of floral meristematic characteristic genes such as FT at flowering sites[40]. FT has also been shown to be mainly expressed in the leaf vein phloem[68]. In this experiment, VcFT in the CK (only pruning) group first shows upregulation in the leaf midvein and buds (Fig. 5), consistent with the transient increase in sucrose content. Subsequently, the sucrose content in the leaves increases significantly, accompanied by an increase in VcFT expression level (Fig. 5). After reaching the morphological differentiation stage during development, the VcFT expression level in leaves and leaf midvein of the CK group gradually decreases with the decrease of sucrose content. VcFT reaches its peak expression level at the Initial morphological differentiation in the CK and CS groups. This suggests that VcFT may be mainly regulated by endogenous sucrose levels but can also be induced by exogenous sucrose. FT positively regulates SOC1 and FUL in Arabidopsis to promote flowering[69]. Meanwhile, the function of SOC1 is partially independent of FT[70]. AtSOC1 is considered the earliest floral transition signal in Arabidopsis[71]. VcSOC1 was upregulated in the buds at the initial flower bud differentiation stage and in the leaves during morphological differentiation, similar to the results of studies in peonies and pears[72]. This indicates that VcSOC1 plays a role downstream of VcFT in buds and integrates upstream signals to initiate the differentiation processes, which are signals of floral transition. In Litchi (Litchi chinensis Sonn.), SOC1 is mainly expressed in mature leaves[73], FUL and SOC1 jointly regulate flower induction, but FUL is mainly expressed in the inflorescence meristem[74]. In rice, AP1 and FUL participate in the transition from meristematic tissue to floral meristematic tissue[4]. VcFUL was upregulated in buds during the morphological differentiation in the CK and CS groups, which seems to confirm this conclusion. In addition to floral regulation, FUL is also involved in leaf development[75], and may participate in the transportation and distribution of carbohydrates[76]. The expression levels of VcFUL in leaves and leaf midvein demonstrate this. As a flowering-promoting factor, FUL is inhibited by the MADS-box transcription factor SVP[67], but in roses, the expression level of SVP increases during floral induction[77]. In Arabidopsis, SOC1, SVP, and AGL24 are involved in the regulation of the floral meristematic tissue differentiation[78], So although SVP inhibits the flowering of most species, it is necessary for the formation of floral meristematic tissues. When the expression level of both SVP and FUL are upregulated, FUL-SVP dimer can inhibit the inhibitory effect of SVP on flowering and promote flowering[79]. In general, the upregulation level of VcFT in the leaf midvein of blueberries is considered the initial signal of floral induction. VcSOC1 is upregulated after integrating multiple upstream signals to initiate morphological differentiation. Subsequently, the expression levels of VcFUL and VcSVP are upregulated to promote the subsequent differentiation of inflorescence primordia and the formation of floral meristematic tissue. Based on these results, a hypothetical regulatory network model was proposed, which involves sucrose as the core response to exogenous and endogenous cues involved in blueberry floral transition (Fig. 10).
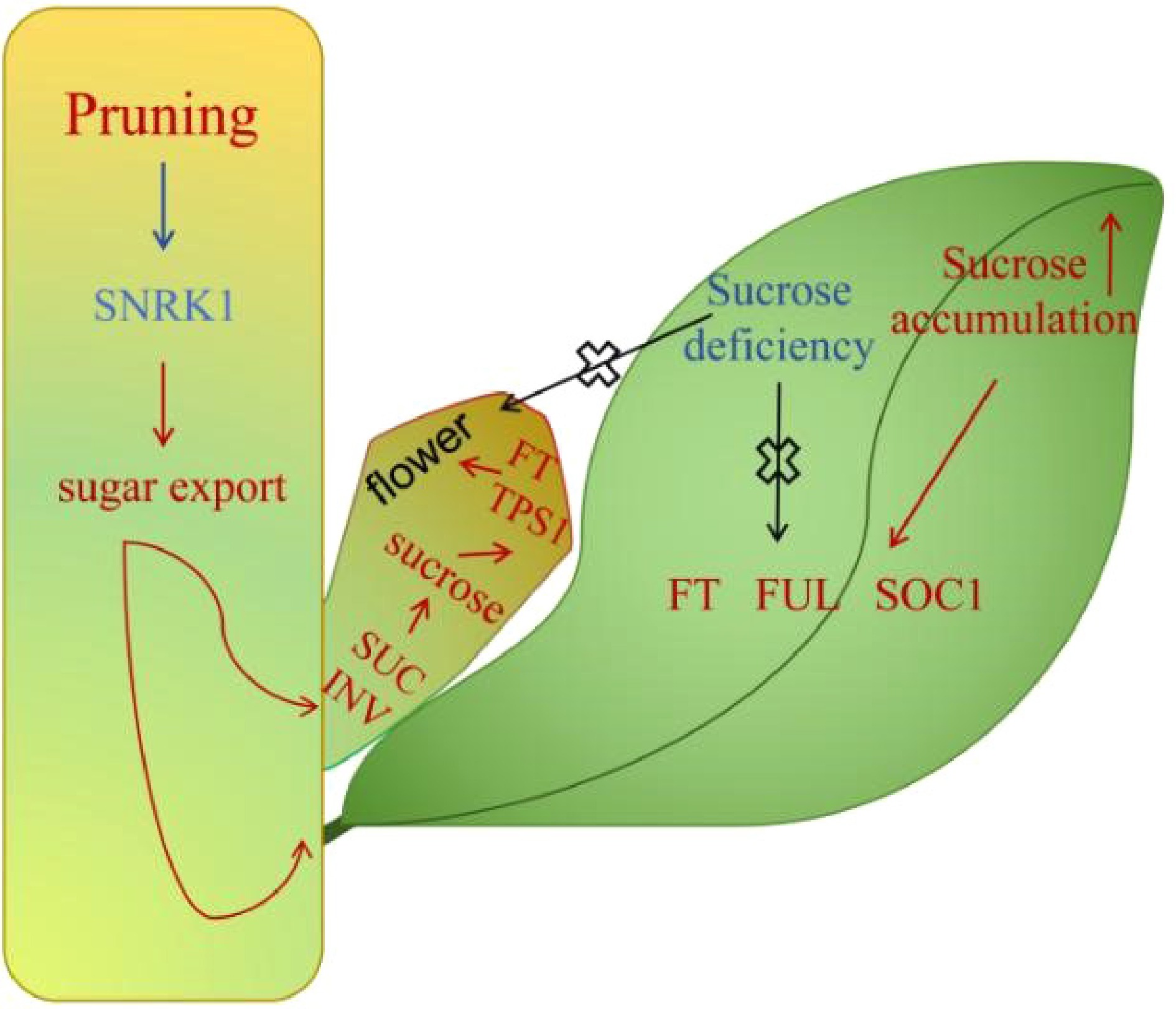
Figure 10.
Hypothetical model depicting the flowering regulation promoted by pruning in blueberries. After pruning, the sugar content in the stem decreases, and the expression levels of VcFT, VcFUL, and VcSOC1 increase as the leaf sucrose continues to accumulate to a certain level. The increased expression levels of VcSUC and VcINV promote sucrose accumulation in buds, and the participation of VcFT and VcTPS1 jointly induces flowering. However, if the sucrose content in the cutting leaves is deficient, the expression of the above genes cannot be induced, and the process of flower bud induction cannot begin.
-
Short pruning can significantly increase the flower bud ratio of strong upright growing highbush blueberry autumn shoots to 64% by promoting carbohydrate transport and metabolism, which is affected by cutting half a leaf and sucrose supplement. During the floral induction period, sucrose and related genes play a promoting role, especially VcAMY, VcINV, and VcSUC. VcFT and VcTPS1 are mainly upregulated in buds, while VcFT is also upregulated in the leaf midvein, all influenced by sucrose content. During the morphological differentiation of flower buds, the expression levels of VcSOC1, VcFUL, VcSUC, and VcSTP are up-regulated, and at this time, the sucrose content in leaves, leaf midvein, and buds gradually decreases, and the glucose and fructose contents in buds show alternating fluctuations. The relative expression level of VcSOC1, VcSUC, VcINV, and VcAMY significantly increase in blueberry overexpressing VcTPS1, while the relative expression level of SNRK1 decreases. The yeast two-hybrid results also show a weak interaction between VcSOC1 and VcTPS1 protein.
This research was supported and funded by the Beijing Natural Science Foundation (Grant No. 6252023), and the Beijing Forestry University Hot Spot Tracking Project (Grant No. 2022BLRD07).
-
The authors confirm their contribution to the paper as follows: study conception and design: Hou Z, Wu X, He S; data collection: Li Y, Xie J, Qie M, Wu X, Zhang S; technical assistance: Jiang Y, Cao M; analysis and interpretation of results: Wu X; draft manuscript preparation: Wu X, Feng X. All authors reviewed the results and approved the final version of the manuscript.
-
he datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xinliang Wu, Yang Li
- Supplementary Table S1 Primer sequences for qRT-PCR.
- Supplementary Fig. S1 Sunshine duration and daily maximum and minimum temperatures from September 23rd to October 23rd, 2023 (source from China Meteorological Administration) (https://www.cma.gov.cn/).
- Supplementary Fig. S2 The growth status of Transforming blueberries. On the left is the pSuper1300 control group, and on the right is the pSuper1300-VcTPS1 treatment group.
- Supplementary Fig. S3 Agarose gel map of the control and VcTPS1 transient expression group.
- Copyright: © 2025 by the author(s). Published byMaximum Academic Press, Fayetteville, GA. This articleis an open access article distributed under Creative CommonsAttribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wu X, Li Y, Zhang S, Qie M, Feng X, et al. 2025. Sucrose participates in the flower bud differentiation regulation promoted by short pruning in blueberry. Fruit Research 5: e025 doi: 10.48130/frures-0025-0017 

Sucrose participates in the flower bud differentiation regulation promoted by short pruning in blueberry
- Received: 12 December 2024
- Revised: 18 April 2025
- Accepted: 20 April 2025
- Published online: 18 June 2025
Abstract: Autumn shoot pruning is a significant production measure in Highbush blueberry (Vaccinium corymbosum L.) that promotes floral differentiation, but the mechanism is still unclear. To explore the role of carbohydrates in floral induction and differentiation promoted by short pruning, carbohydrate content, and the transport and metabolic-related genes were compared and analyzed. It is found that sucrose might be an important trigger for pruning to promote floral induction. Compared with only pruning the shoots, cutting half of the leaves significantly reduces the flower bud rate and delays the flower differentiation process, while further adding 1.5% sucrose spray alleviates this phenomenon partly. During floral induction, the soluble sugar content in buds increases significantly, while the sucrose content in leaf midvein and buds increases earlier. The starch and soluble sugar content in the stem decreases significantly. At the early flower bud morphological differentiation stage, a large number of starch granules accumulate in the buds, sucrose in leaves increases, glucose in buds decreases, and then fructose decreases. There is an upregulation of VcFT, VcSOC1, VcFUL, VcSUC, and VcINV expression in the leaf midvein, as well as an upregulation of VcFT, VcSOC1, VcTPS1, VcINV, and VcSWEET1 expression in buds during floral induction. At the same time, interaction between VcSOC1 and VcTPS1 proteins exists. The level of VcSOC1 and VcAMY can also be upregulated by VcTPS1. Sucrose plays an important role in blueberry floral induction through flowering factors and sugar-related genes, especially VcFT, VcSOC1, VcTPS1, VcSUC, and VcINV. The results of this study laid a strong foundation for further research on the regulation mechanism of flower bud differentiation in blueberry and other woody plants.
-
Key words:
- Autumn shoot /
- Blueberry /
- Flower bud differentiation /
- Short pruning /
- Sucrose /
- VcTPS1