









-
Blueberries are originally defined based on their blue or black blue fruit skin in the genus Vaccinium of the family Ericaceae. Blueberry is commercially pointed to V. corymbosum and V. ashei in a restrictive meaning. People favor the fruits for their unique flavor and high contents of anthocyanins and other diverse antioxidant compounds[1,2]. Blueberries exhibit remarkable anti-inflammatory, antioxidant, anticancer, and vision-enhancing properties. These bioactive advantages not only underscore their substantial economic and healthcare significance, but also establish the fruit as a compelling subject for multidisciplinary research in nutritional science and phytomedicine[3−6]. Although blueberry was introduced to China at the end of 19th century, the cultivated area and production is presently about 96 thousand hectares, and 780 thousand tons, respectively. More than 70% of the blueberry harvest in China is used for fresh-market fruits, making fruit firmness one of the most critical breeding traits for blueberry.
Fruit firmness profoundly influences blueberry mechanical harvesting efficiency, intrinsic quality, transportability, and shelf life. It serves as a decisive indicator of freshness preservation particularly for fresh-market blueberries. Blueberries with enhanced firmness exhibit good cellular resilience during mastication, delivering a superior crisp texture highly valued by consumers. Nevertheless, the scarcity of high-firmness cultivars has emerged as a primary bottleneck constraining the sustainable development of the blueberry industry, compromising both post-harvest logistics and consumer satisfaction. Fruit firmness is intrinsically linked to the cell wall architecture of mesocarp cells and their associated physiochemical alterations during development[7−9]. Typically exhibiting progressive decline with advancing ripening stages, or prolonged storage duration, premature softening can also be triggered in developing fruits by mechanical injuries, phytopathogen attacks, and abiotic stresses. Optimal ripening firmness improves consumer-oriented textural properties, whereas excessive softening compromises intrinsic quality attributes, and elevates postharvest losses during harvesting and transportation[10]. While the ripening-associated firmness reduction represents an inevitable physiological process, developing fruits with superior pre-ripening firmness demonstrate a remarkable capacity to retain residual firmness characteristics through maturation phases[11,12].
The main cause for fruit firmness decrease is the degradation of cell wall components. Several cell wall-metabolizing enzymes containing polygalacturonase (PG), pectin methyl esterase (PME), and pectin lyase (PL) are involved in the degradation process[10,13]. Previous studies suggested that lateral organ boundaries domain (LOB or LBD) transcription factors directly or indirectly regulate fruit firmness. LOB transcription factors are unique to higher plants, characterized a conserved LOB domain at the N-terminus[14]. LOB domain is comprised of three conserved motifs, including a CX2CX6CX3C zinc finger motif, a GAS motif made up of conserved amino acids Gly-Ala-Ser, and a Leucine Zipper-like Motif (LZLM)[15]. The zinc finger and GAS motifs bind to the promoters of downstream genes[16], while the LZLM is associated with protein dimerization[17,18]. The LOB family can be classified into two subfamilies based on the domain variations, i. e. members in subfamily I have the complete LOB domain structure, whereas members in subfamily II lack the LZLM[19]. Most LOB members fall into subfamily I. There are 43 LOB members in Arabidopsis thaliana, with only seven belonging to subfamily II[19], and of the 46 LOB members in tomato (Solanum lycopersicum), only six are part of the subfamily II[20].
LOBs play critical roles at diverse steps during plant growth, development, and metabolism. SlLBD3, SlLBD20, SlLBD22, and SlLBD47 were involved in fruit development and ripening, among which SlLBD20 and SlLBD47 were found exclusively expressed in tomato fruit[21]. SlLOB1 regulates the softening process in tomato fruits by directly controlling the activity of the EXP1 promoter[22]. An analysis of the correlation between LOB gene expression patterns and fruit firmness in melon (Cucumis melo) revealed a significantly negative correlation between CmLOB2 gene expression and fruit firmness during ripening[23]. MaLOB1/2/3 in banana (Musa acuminate) can be activated by ethylene and directly bind to MaEXP1/2 promoters, participating in the fruit ripening and softening processes[24]. However, there is no report on LOB transcription factors regulating fruit firmness in blueberries. Whether the fruit firmness differences between firm- and soft-flesh blueberries were regulated by LOBs need to be elucidated. With genome-wide identification and expression pattern analysis of blueberry LOB genes, we identified VcLOB33, VcLOB74, VcLOB107, and VcLOB114 as the top-related genes that affect fruit firmness. The biological function of the VcLOB107 gene was further validated using transgenic tomatoes. Our findings lay foundation for further studies on the molecular mechanism of VcLOB genes regulating fruit firmness during blueberry development, and firmness differences between firm and soft-flesh blueberry cultivars.
-
Blueberry fruits were obtained from the germplasm repository at Zhejiang Normal University. The study focused on two southern highbush blueberry cultivars, 'Star' (firm-flesh) and 'O'Neal' (soft-flesh), which are widely cultivated in the middle and lower reaches of the Yangtze River region. 'O'Neal' serves as the male parent of 'Star', and despite their similar genetic background, the two cultivars exhibit significant differences in fruit firmness. Following the developmental stage classification for blueberries established by Zifkin et al.[25], which is based on fruit size and skin coloration, fruits from both cultivars were harvested at six developmental stages (S3 to S8) for analysis (Fig. 1). Fruit firmness of both 'Star' and 'O'Neal' initially increased, and then decreased (Supplementary Fig. S1). Specifically, fruit firmness peaked at stage S4, reaching 1,061 g·mm−1 for 'Star', and 1,207 g·mm−1 for 'O'Neal'. Fruit firmness declined sharply from stage S4 to S6, with 'O'Neal' showing a more rapid decrease than 'Star'. The fruits were left at room temperature for 1 h to eliminate field heat after harvesting, damaged and diseased fruits were removed. Fruits with similar size and developmental state were chosen for liquid nitrogen treatment and stored at −80 °C for later use. Stems, leaves, flowers, and flower buds from these two blueberry cultivars were also collected, treated using liquid nitrogen and preserved at −80 °C for later use.

Figure 1.
'O'Neal' and 'Star' fruits at different developmental stages.
Transcriptome sequencing
-
Total RNA from blueberry fruits and different organs was extracted using the modified CTAB method, and the first-strand cDNA synthesis was performed using a reverse transcription kit purchased from Nanjing Novozan Biotechnology Co., Ltd. (Vazyme, Nanjing). For each sample, 1 μg of fruit RNA was used to construct sequencing libraries with the NEBNext® Ultra™ RNA Library Prep Kit (NEB, USA). The library concentration and insert sizes were respectively determined using Qubit 2.0 and Agilent 2100. High-throughput sequencing was carried out on the Illumina NovaSeq 6000 platform with a read length of paired-end 150 base pairs. Reads were filtered to remove those with a greater than 10% incidence of Ns, and those with a base quality score of Q ≤ 10 comprising over 50% of the read length, retaining other reads for transcriptome assembly. Sequence assembly and annotation was accomplished using HISAT2[26] and StringTie[27] with the blueberry 'Draper' genome[28]. Adjusted p-values (padj. < 0.01) of multiple tests were corrected using Benjamini and Yekutieli's approach for controlling the false discovery rate (FDR)[29]. Transcripts Per Kilobase of exon model per Million mapped reads (TPM) were calculated using RSEM[30].
Identification of the blueberry LOB genes
-
Arabidopsis LOB amino acid sequences were retrieved and downloaded from the National Center for Biotechnology Information (NCBI,
www.ncbi.nlm.nih.gov ) to serve as query sequences. The BLAST software package was used to search for homologous sequences in the blueberry proteome database (www.vaccinium.org/analysis/49 ), with an e-value set to 1 × 10−20 and all other parameters remaining at default settings. The Hidden Markov Model for the LOB domain (PF03195) was downloaded from the InterPro database, and homologous sequence analysis was conducted using the HMMER software package. The combined, non-redundant results from BLAST and HMMER searches were taken as candidate members of the blueberry LOB family. The NCBI Conserved Domain Search service (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ) was used to inspect the conserved domains of the candidate members, and sequences with missing or incomplete domains were manually removed. The TBtools v.2.034 software package was used to calculate the basic physicochemical properties of LOB family member proteins. The gene serial numbers of the aforementioned genes can be found in Supplementary Table S1.Construction and classification of the phylogenetic tree of blueberry LOB genes
-
A multiple sequence alignment analysis of LOB amino acid sequences from blueberry and Arabidopsis was conducted using MEGA 11[31]. The best model for amino acid substitution was evaluated using IQ-TREE 2.2.0, and the phylogenetic tree was constructed using the maximum likelihood method with 1,000 replicates bootstrap[32]. Following the classification method for Arabidopsis LOB family members, the blueberry LOB family was divided into two major subfamilies. The phylogenetic tree visualization was enhanced with the online tree drawing tool iTOL (
https://itol.embl.de/ ).Chromosomal localization, gene structure, and conserved motif analysis of blueberry LOB genes
-
TBtools was used to extract information on the chromosomal localization of VcLOB from the blueberry genome sequence and annotation files. The blueberry LOB genes were named based on their physical positions on the blueberry genome. The MG2C online tool (
http://mg2c.iask.in/mg2c_v2.1/ ) was employed for the visualization analysis of chromosomal localization information. MEME 5.5.1 software (http://meme-suite.org/tools/meme ) was used to predict and analyze the conserved motifs of blueberry LOB proteins, with the number of motifs set to 15. TBtools was applied for the visualization analysis of the conserved motifs within the blueberry LOB family.Synteny analysis of blueberry LOB genes
-
TBtools was also used for the visual analysis of the gene structure within the blueberry LOB family[33]. MCScanX[34] and Circos[35] were applied to estimate the syntenic relationships of VcLOB genes and to draw synteny maps. To understand the selective pressure on the VcLOB genes, we calculated the Ka/Ks ratio (non-synonymous to synonymous substitution rates) for each pair. Duplicated gene pairs were identified using DupGen finder, and their coding sequences (CDS) were aligned with MAFFT (v7.505) using default parameters. Ka/Ks values were then computed using Tbtools.
Expression pattern analysis of blueberry LOB genes during different fruit developmental stages and in various organs
-
The relative expression fold change of the same VcLOB gene between the two blueberry cultivars was calculated by dividing the TPM value of 'O'Neal' and 'Star' at the same developmental stage. The results were used to generate a gene heatmap using TBtools. Based on the coding sequence (CDS) information of the VcLOB genes, quantitative primers for VcLOB were designed using the Primer Designing Tool (
www.ncbi.nlm.nih.gov/tools/primer-blast ), with VcGAPDH serving as the internal reference gene. cDNA synthesis was carried out in accordance with the manual of the HiScript® Reverse Transcriptase Kit by Vazyme. Quantitative PCR (qPCR) reactions were conducted using the 2 × SYBR Green qPCR Mix (High ROX) Kit. The 10 μL reaction system consisted of: 2 × SYBR Green qPCR Mix (High ROX) 5 μL, upstream primer (10 μmol·L−1), downstream primer (10 μmol·L−1) 1 μL each, cDNA 1 μL, and ddH2O 2 μL. PCR reaction conditions were as follows: initial denaturation at 95 °C for 4 min; denaturation at 95 °C for 30 s, annealing at 57 °C for 30 s, extension at 72 °C for 30 s, for 40 cycles, followed by a melt curve analysis. The relative expression levels of genes were calculated using the 2−ΔΔCᴛ method[36]. Each sample was examined in three biological replicates and four technical replicates. Details of the primers can be found in Supplementary Table S2. After obtaining the relative expression levels of genes, a t-test was performed on the VcLOB gene expression levels using IBM SPSS Statistics 23. A p-value less than 0.05 indicates a significant difference, and less than 0.01 indicates an extremely significant difference.Measurement of blueberry fruit firmness, pectin content, and PG activity
-
A sufficient quantity of fruit (≥ 75) was selected for firmness measurement using the Firmtech FT7 Fruit Firmness Tester (UP GmbH Firmensitz, Germany). The firmness tester's probe was applied perpendicularly to the equatorial plane of the fruit, with deformation threshold modes of 1.0 and 2.0 mm combined. The fruit firmness was recorded using FT7 Control software (UP GmbH Firmensitz, Germany)[37]. Cell wall material (CWM) extraction was carried out following the method by Chen et al.[38]. Pectin extraction was performed according to the method by Kozioł et al.[39]. Pectin content was determined using the carbazole method[40], with the procedure repeated three times for consistency. Fresh blueberry fruits were selected to measure the PG activity using a polygalacturonase assay kit (Grace Biotechnology, Suzhou).
Overexpression of VcLOB107 in tomato
-
Cloning primers were designed using Primer Premier 5 based on VcLOB107 sequence (Supplementary Table S3). PCR amplification was performed using PrimeSTAR Max DNA Polymerase (TaKaRa, Dalian), with mixed cDNA from 'O'Neal' fruit at stages S3 to S8 as the template. The target fragment was purified using an agarose gel recovery kit (Haoxin Biotech, Hangzhou), and ligated into the pMD19-T vector (TaKaRa, Dalian). The ligated product was transformed into E. coli DH5α, and positive clones were screened and sequenced (Shangya Biotech, Zhejiang) to obtain strains containing the target gene.
For overexpression vector construction, the empty pCAMBIA1302 vector and plasmids containing VcLOB107 were double-digested with EcoRI and HindIII to generate compatible sticky ends. The target fragments were ligated into the digested pCAMBIA1302 vector using T4 DNA Ligase (TaKaRa, Dalian) (Supplementary Fig. S2). The recombinant plasmids were transformed into E. coli DH5α, and positive clones were screened and sequenced (Shangya Biotech, Zhejiang) to obtain strains containing the recombinant plasmids.
The recombinant plasmids were then introduced into Agrobacterium tumefaciens GV3101 using the freeze-thaw method. Positive strains were screened, and the activated Agrobacterium culture was used to infect wild-type MicroTom tomato via hypocotyl infection. Briefly, the cotyledons of tomato seedlings were removed and the cut surfaces were repeatedly treated with the infection solution. The seedlings were transferred to normal growth conditions after 12 hours of dark incubation.
-
A total of 150 VcLOB genes were identified in the blueberry genome. They were named from VcLOB1 to VcLOB150 according to their physical localization on the scaffold. A phylogenetic tree was constructed using 43 AtLOB protein sequences, and all VcLOB protein sequences, to investigate the subfamilies within VcLOBs. Based on the identified AtLOBs phylogenetic affiliations, the 150 VcLOB proteins could be divided into Class I and Class II with 112 and 38 VcLOBs within two classes, respectively. Class I was further separated into four subfamilies (Ia, Ib, Ic, and Id) while Class II contained IIa and IIb based on the phylogenetic relationship (Fig. 2). Furthermore, VcLOB numbers within each subfamily were unevenly distributed. The members in Class I subfamilies ranged from 23 to 34, with an average of 28. The subfamilies in Class II showed dramatic differences in member numbers with Class IIa and IIb containing 11 and 27 VcLOBs, respectively.
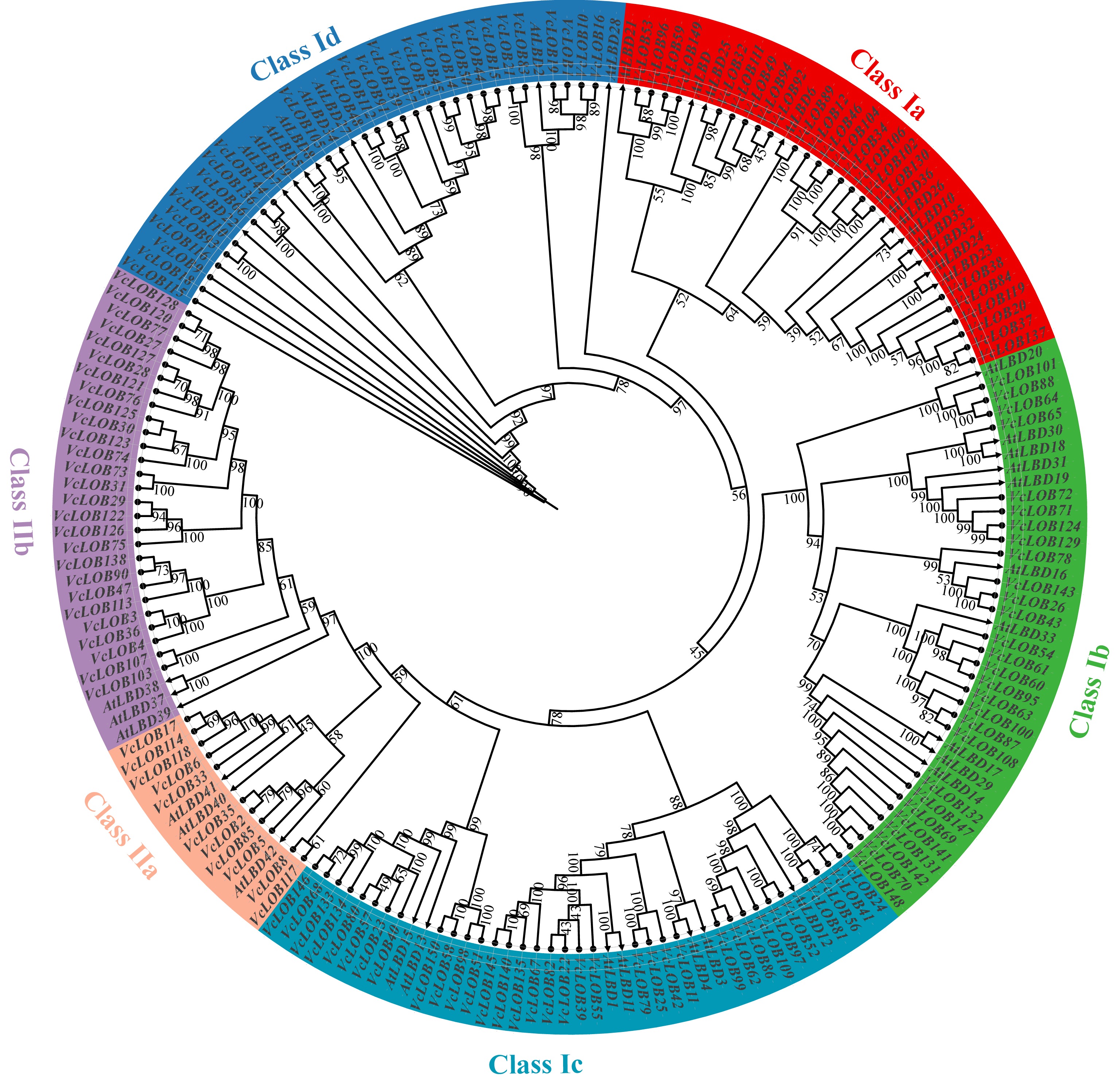
Figure 2.
Phylogenetic tree of LOB proteins in blueberry and Arabidopsis thaliana. LOB proteins of blueberry and Arabidopsis are marked with circles and triangles, respectively. The digits on the branches represent bootstrap values for 1,000 replicates.
Chromosomal localization and collinearity analysis of the VcLOB genes
-
The 150 VcLOB genes are distributed on nearly all chromosomes except for chromosomes 38 and 39 (Fig. 3). The VcLOB gene numbers of each chromosome ranged from one to seven, with an average number of three. Five chromosomes including chromosomes 11, 15, 19, 23, and 24 showed a higher number of VcLOB distribution than other chromosomes. The VcLOB genes usually exhibited a clustering style on those gene-rich chromosomes. Eleven chromosomes contained only one VcLOB gene. Further analysis of chromosomal localization indicated that Class I members are predominantly found on chromosomes 11, 15, 24, and 43. Class II is mainly found on chromosomes 12, 23, 40, and 41. Thirty two VcLOB genes form 10 gene clusters distributed on 10 chromosomes, with four clusters composed of Class IIb members and four clusters contained Class Ib members, suggesting a clustering distribution trend among gene members of the same subfamily.
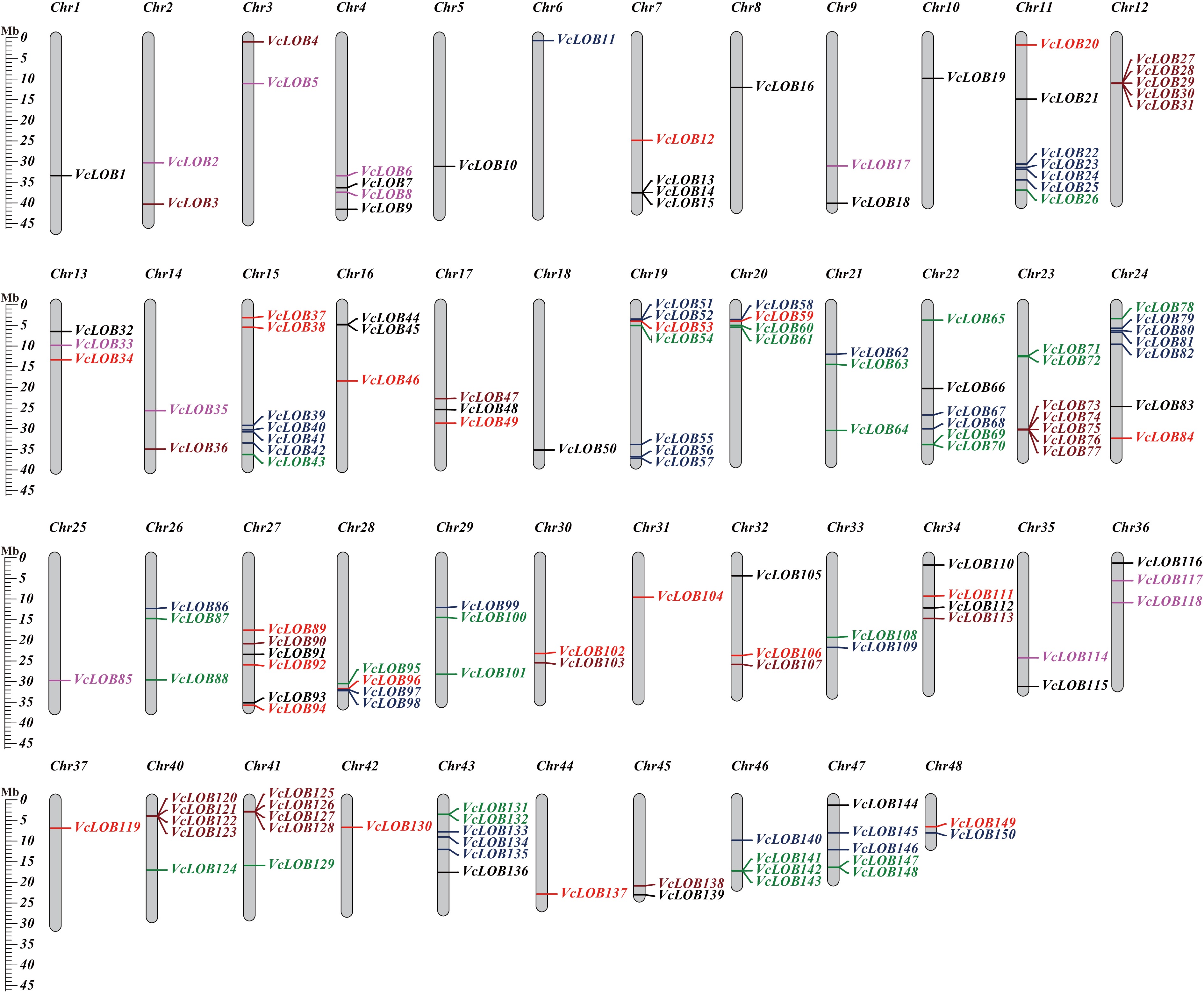
Figure 3.
VcLOB gene distribution on 48 blueberry chromosomes. Class Ia, Class Ib, Class Ic, Class Id, Class IIa, and Class IIb genes are shown in black, red, green, blue, brown, and purple, respectively.
Collinearity results identified 303 pairs of collinear genes with whole-genome duplication or segmental duplication events, occurring among different chromosomes (Fig. 4), with 80.6% of the duplications occurring within Class I genes. The calculation of Ka/Ks values for collinear gene pairs (Supplementary Table S4) revealed the evolutionary selective pressure on VcLOB genes. The results suggested that most genes may have undergone purifying selection during the course of evolution.
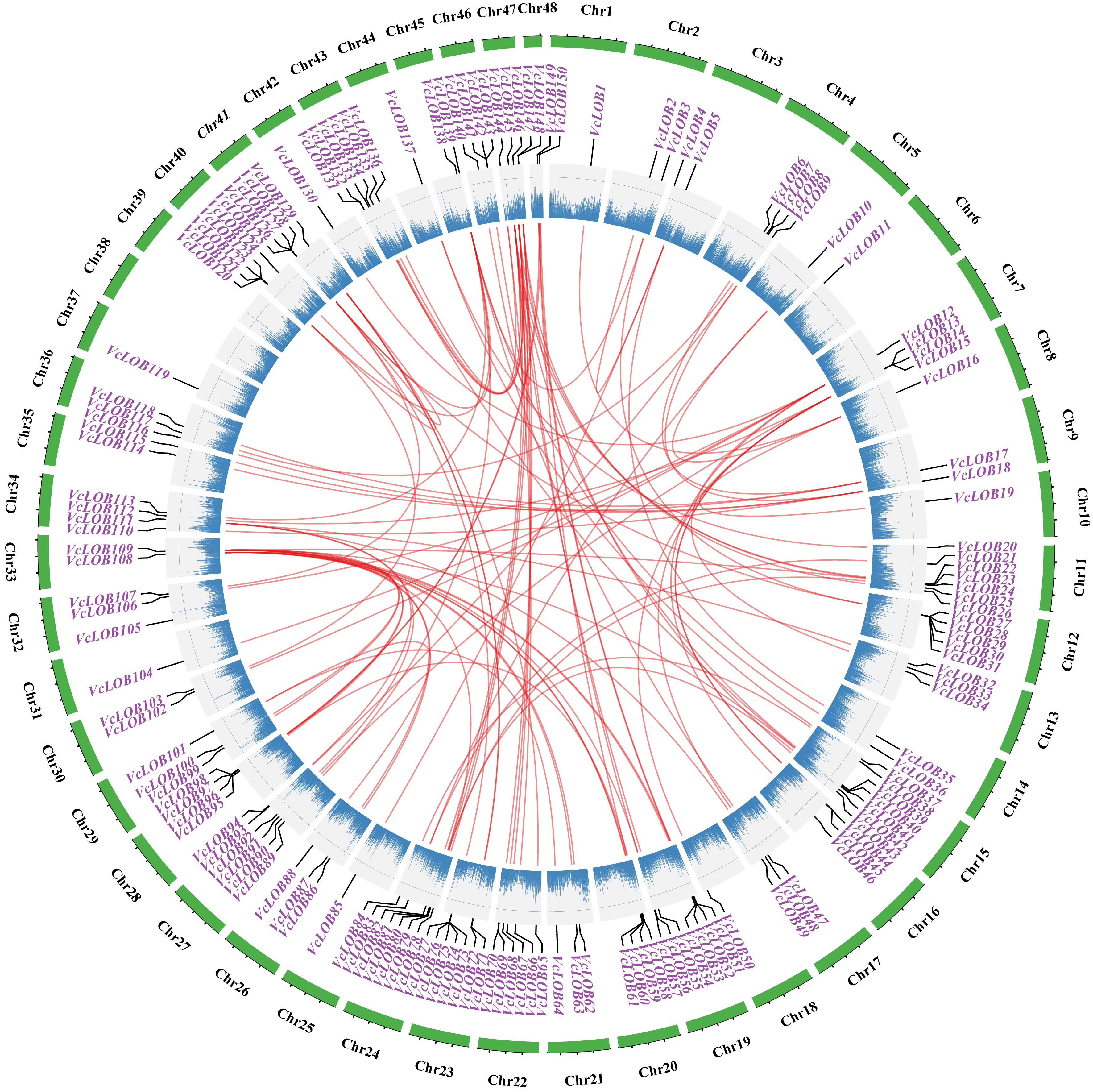
Figure 4.
Collinearity analysis of 150 VcLOB genes in blueberry genome. Red represents gene pairs, and blue scatter represents the gene frequency on the chromosome.
Gene structure and conserved motifs of VcLOB genes
-
The lengths of VcLOB genes range from 512 to 12,505 bp, with an average length of 2,316 bp. Significant differences in gene structures between members of Class I and Class II were found (Fig. 5). Class I members have shorter gene lengths than Class II members. Class Ia members have an average gene length of 3,016 bp, among which VcLOB16 (12,505 bp), and VcLOB19 (10,620 bp) are significantly longer than other genes. Except for VcLOB1, VcLOB10, VcLOB16, VcLOB19, and VcLOB105, the remaining genes have one or two exons. Class Ia members had exon numbers ranging from one to three, and an average gene length of 1,699 bp. VcLOB34 (5,989 bp) and VcLOB106 (4,773 bp) were significantly longer than other genes in Class Ia. Only VcLOB34 and VcLOB106 possess untranslated regions (UTR) structures. The average gene length of Class Ib members was 2,098 bp, among which VcLOB65 (6,727 bp) is the largest, and exon numbers ranged from two to four. Class Ic members have two or three exons, with an average gene length of 3,079 bp. Compared to Class I, Class II members are more conservative in their gene structure. The average gene length of Class IIb members is 1,385 bp, with VcLOB73 (2,751 bp) being the longest. VcLOB73 and VcLOB31 have three exons each, while the rest have two exons. Apart from VcLOB73 and VcLOB30, the rest genes possess untranslated regions (UTR). The exon number of Class IIa members ranged from two to five, with an average gene length of 2,368 bp. VcLOB117 (5,647 bp) and VcLOB8 (5,556 bp) gene length were significantly longer than other members and neither gene has UTR structures.
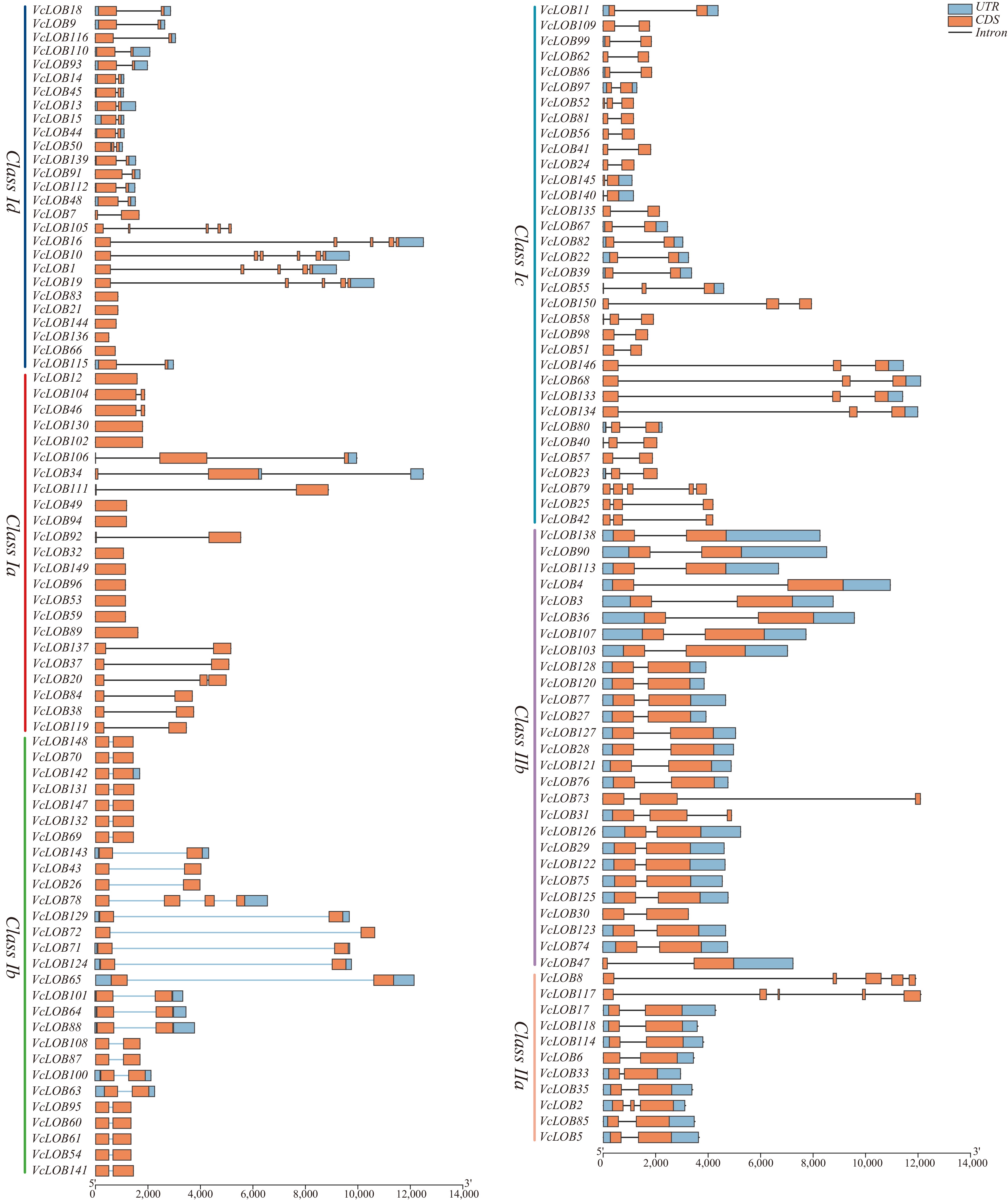
Figure 5.
Gene structure analysis of 150 VcLOB genes.
We performed a conserved motif analysis of VcLOB proteins (Fig. 6) and identified 15 conserved motifs (Supplementary Table S5). Class I contained seven unique motifs, among which motif 5 is only found in Class Ib, while motif 12 is exclusive to Class Id. Motifs 6, 7, and 9 are specific to Class II, with motifs 6 and 7 present only in Class IIb members. A significant portion (93.3%) of VcLOB members contain motifs 1, 2, 3, and 4. Motif 5 (YPQDVQSWFQSQNPNIIPQLDPNTASNFGNNPFYGNGIMNQ) has the most conserved amino acid sequence, whereas motifs 9 and 10 exhibit higher variability at their amino acid positions. Motif 4 has the shortest amino acid sequence, comprising only 15 amino acids.
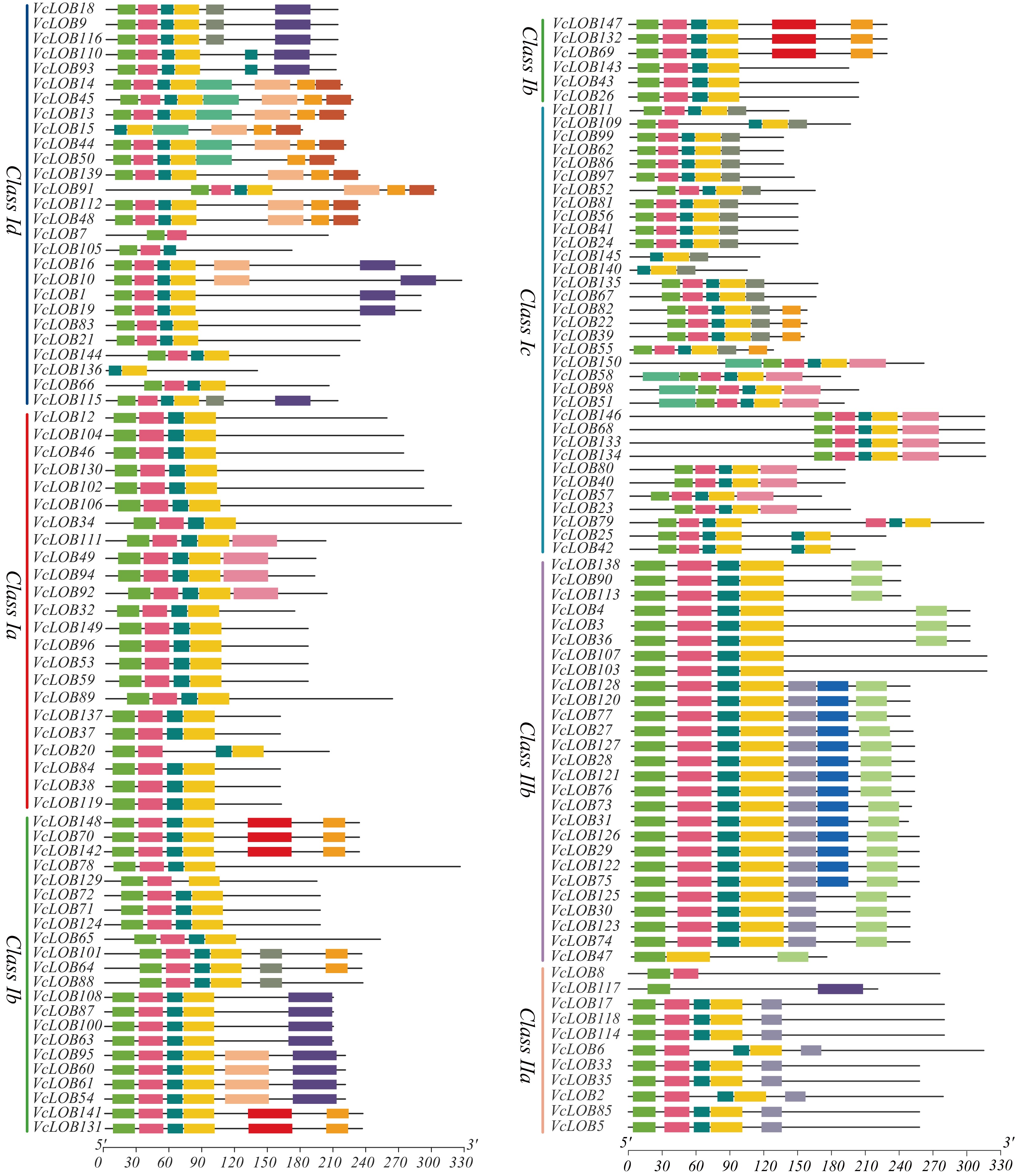
Figure 6.
Conserved motif analysis of 150 VcLOB proteins.
Transcript abundance differences and expression patterns of VcLOB genes at various fruit developmental stages
-
We randomly selected 30 genes based on the TPM values (Supplementary Table S6) of 'O' Neal' and 'Star' from S3 to S8, and calculated the fold change in TPM values between the two blueberry cultivars at the same developmental stage by calculating the 'O'Neal' to 'Star' TPM ratios. A ratio heatmap was created to illustrate their transcript abundance differences (Fig. 7). If the genes showed more transcript abundance in 'O'Neal' than that in 'Star', the values in the heatmap were indicated with ≥ 0, conversely presented with < 0. The transcript abundance of 22 genes was significantly higher in O'Neal' than those in 'Star' during the S3 stage. In 'O'Neal', VcLOB78 and VcLOB11 showed significantly higher expression levels than 'Star' at stage S4, whereas VcLOB103, VcLOB102, and VcLOB107 had significantly higher expression at stage S7. VcLOB32 and VcLOB16 had notably higher expression in 'O'Neal' compared to 'Star' at stage S6, and VcLOB10 in 'O'Neal' exhibited significantly greater expression than 'Star' at stage S8. The expression levels of these 30 genes at the S5 stage were higher in 'Star' than that in 'O'Neal'.
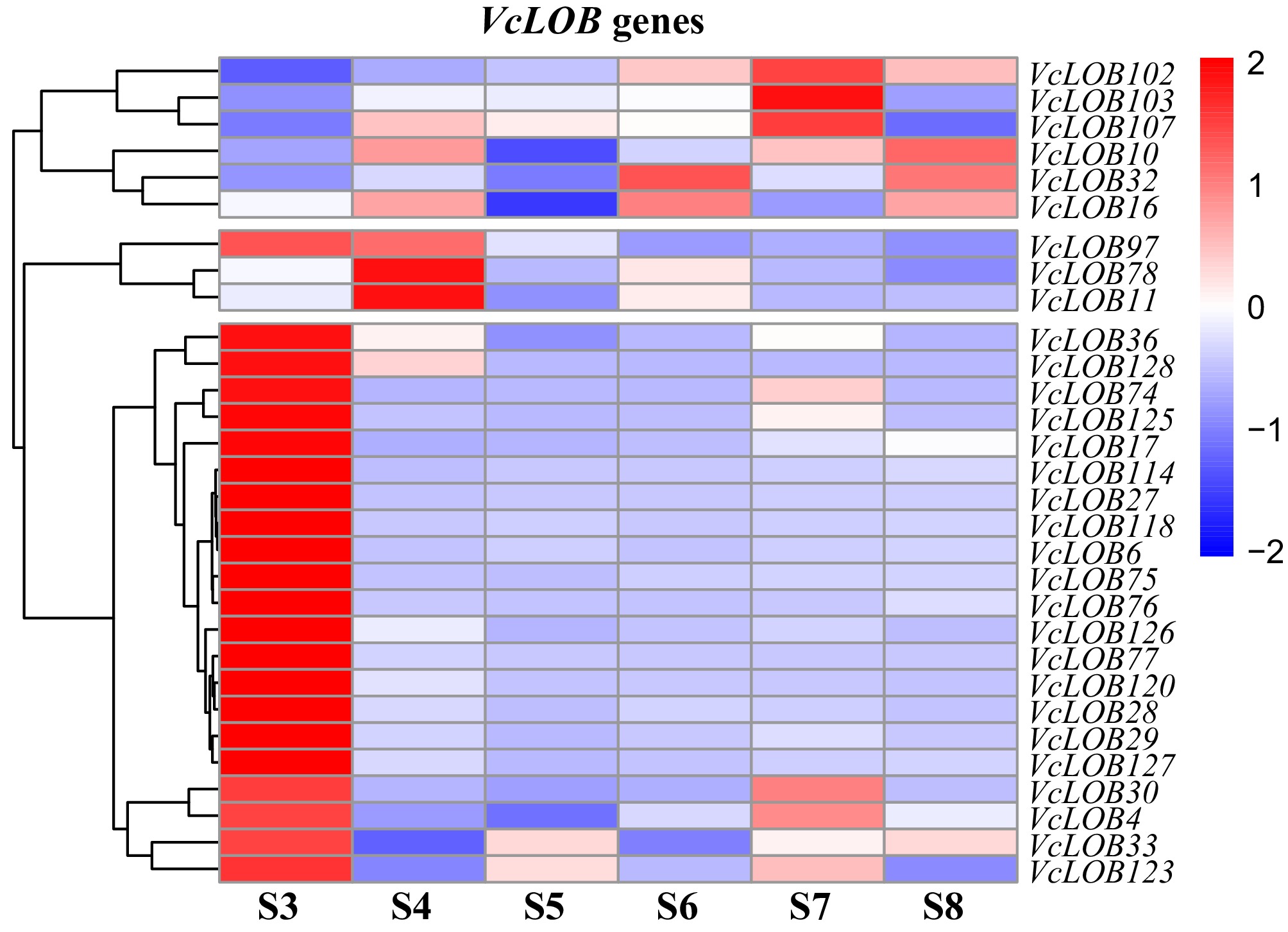
Figure 7.
TPM ratio heatmap of 30 VcLOB genes between 'O'Neal' and 'Star' blueberry fruits during the ripening process.
We found four genes that showed significant differences in TPM values between stages and cultivars. They were further analyzed using quantitative PCR. The qPCR results revealed that the relative expression level of VcLOB33 in both blueberry cultivars displayed a trend of initially increasing and then decreasing during fruit development (Fig. 8). The expression level peaked at the S6 stage in 'Star' while it reached a maximum value at S5 stage in 'O'Neal'. Significant differences in VcLOB33 expression levels were observed at stages S4, S5, and S6 between 'Star' and 'O'Neal'. The expression values of VcLOB33 in 'Star' at stages S4 and S6 were 3.00 times compared to that in 'O'Neal'. VcLOB74 showed a higher expression level in 'O'Neal' than that in 'Star' after stage S5, especially at stages S7 and S8. The expression values of VcLOB74 in 'O'Neal' at stages S4 and S6 was 0.98 times compared to that in 'Star'. VcLOB107 reached its maximum expression level of 8.28 at stage S7 in 'O'Neal', which was 4.45 times compared to that in 'Star'. The peak expression value of VcLOB107 in 'Star' was found at stage S5, which did not show significant differences with that in 'O'Neal'. Low expression levels were found for the VcLOB114 gene at stages in both cultivars except for stage S4. Dramatically different expression levels were observed at stage S4 between 'O'Neal' and 'Star'. The expression levels were 2.09 and 14.8 in 'O'Neal' and 'Star', respectively. Although significant differences in expression level were also detected at stages S5, S7, and S8, the VcLOB114 gene showed lower expression values than other VcLOB genes.
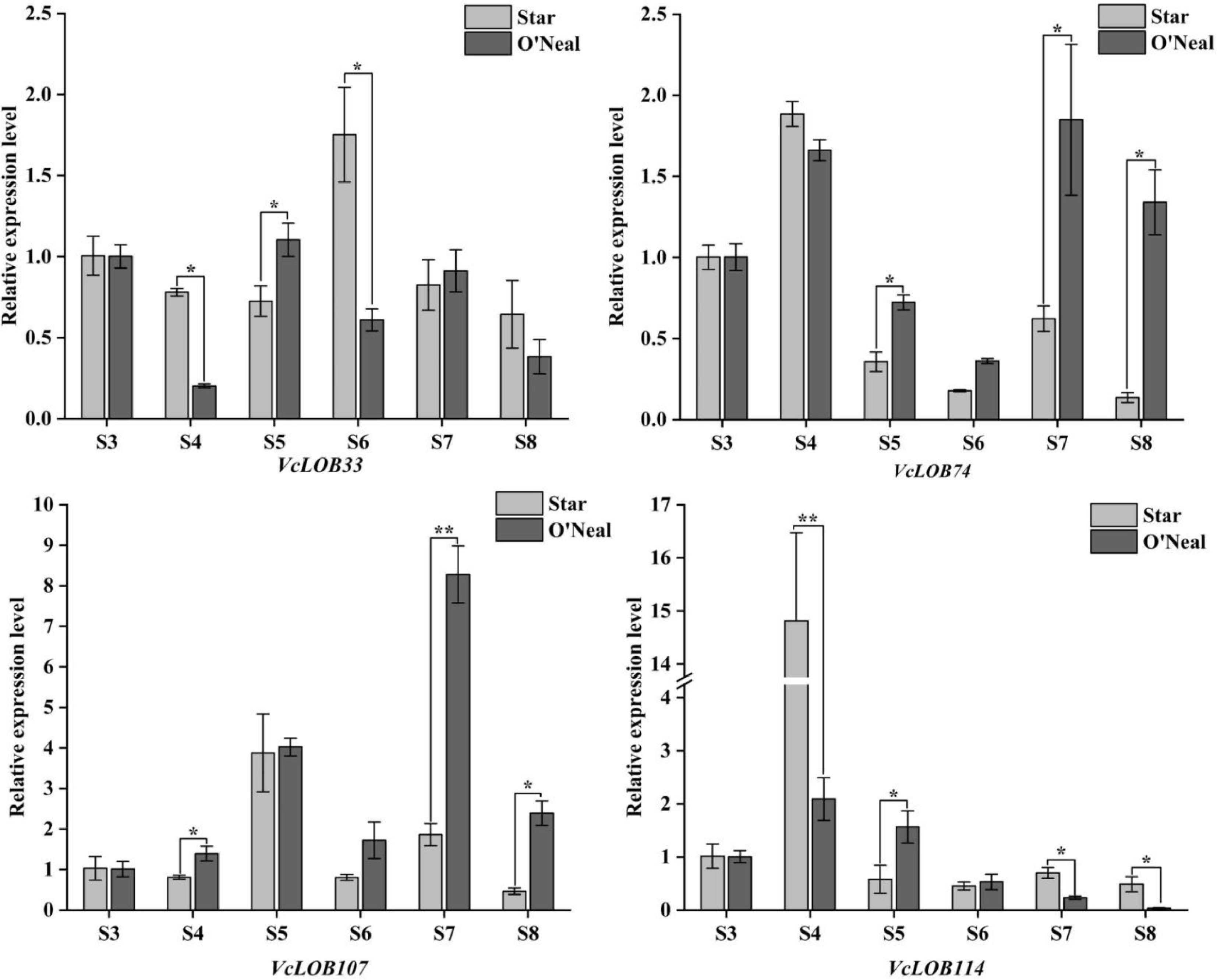
Figure 8.
Relative expression of VcLOB genes during fruit development (t-test, * p < 0.05, ** p < 0.01).
Expression patterns of LOB genes in different tissues of blueberry
-
The expression patterns of VcLOB genes in fruits, flowers, leaves, and stems of 'O'Neal' and 'Star' blueberry were investigated (Supplementary Fig. S3). Generally, VcLOB33, VcLOB74, VcLOB107, and VcLOB114 showed distinct expression patterns among tissues in those two blueberry cultivars. All four genes were highly expressed in 'O'Neal'. VcLOB114 exhibited the highest relative expression level in the 'O'Neal' flowers. Interestingly, those four genes were expressed at lower values in 'Star' than that in 'O'Neal' with an exception of VcLOB107 in the shoots of blueberry 'Star'. Significant differences in expression levels were detected between the leaves and shoots of the two cultivars. The expression levels of VcLOB33 and VcLOB114 showed additional differences between flowers of the two blueberry cultivars. Due to the mixed sampling of fruits in this step, no significant expression differences were detected for the four genes in fruits between 'Star' and 'O'Neal'.
PG activity and pectin content at different stages between two blueberries
-
We measured polygalacturonase activity of the blueberry fruits from stages S3 to S8 (Fig. 9a). The PG activity showed an increasing trend accompanied with the fruit development, and both reached summit values at stage S8 with values of 1.06 and 1.15 mg·h−1·g−1 fresh weight for 'Star' and 'O'Neal', respectively. Notably, The PG activity in 'O'Neal' fruit was significantly higher than that in 'Star' at stages S5 to S7 which corresponds to fruit firmness decrease in those two blueberries. We also tested various types of pectin at different stages in the fruit, including water-soluble pectin (WSP), chelator-soluble pectin (CSP), and dilute-alkali-soluble pectin (DASP). The WSP content between 'Star' and 'O'Neal' showed significant differences among fruit developmental stages, with stages S3 and S7 as exceptions (Fig. 9c). 'Star' fruits only contained significantly higher WSP than 'O'Neal' at stage S4. The WSP contents in the 'O'Neal' fruit was higher than that of 'Star' at late ripening stages except for stage S7. The WSP content in 'O'Neal' rapidly increased at S5, and reached a maximum of 74.06 mg·g−1 cell wall material at S7. The CSP content between the two cultivars showed similar trends with WSP, while only stages S4 and S6 showed significant differences between the two (Fig. 9d). 'Star' fruit contained more CSP than 'O'Neal' fruit at stages S3 and S4, but more CSP was detected in 'O'Neal' fruits from stage S5 to S8. The highest CSP content was observed at stage S7 with a value of 58.75 mg·g−1 CWM. The DASP content displayed a different trend with both WSP and CSP, which presented its peak at stage S3, with both cultivars reaching maximum values at S3 (Fig. 9e). No notable differences were investigated for DASP contents between 'O'Neal' and 'Star' throughout the developmental stages.
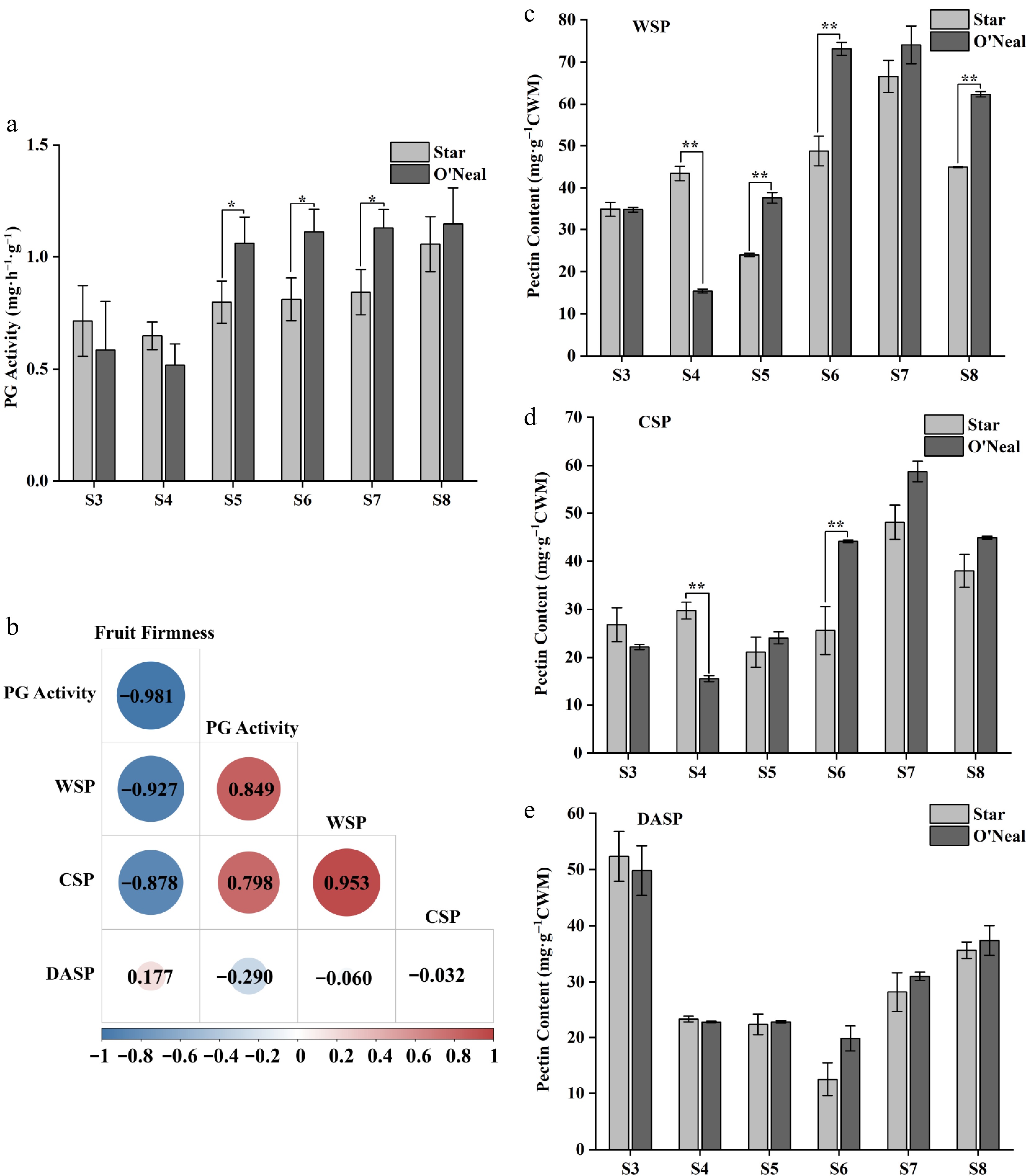
Figure 9.
PG activity, correlation between fruit firmness and physiological indicators, and pectin contents at different developmental stages of 'Star' and 'O'Neal' fruits. (t-test, * p < 0.05, ** p < 0.01).
A correlation analysis was conducted on the hardness of blueberry fruits and their physiological indices (Fig. 9b). The results indicate that fruit firmness of blueberry is negatively correlated with the WSP content, CSP content, and PG activity with correlationship coefficients of −0.927, −0.878, and −0.981. No significant correlation between fruit firmness and DASP dilute content was found. Additionally, the WSP content of fruit showed a positive correlation with the CSP content and PG activity, with correlation coefficients of 0.953 and 0.849, respectively. There is no significant correlation between CSP content and either DASP content or PG activity. Similarly, DASP content had a weak correlation with PG activity.
Overexpression of the VcLOB107 gene delays tomato fruit softening
-
Among the four candidate genes identified, VcLOB107 exhibited the strongest association with the difference in fruit firmness between 'Star' and 'O'Neal'. To further investigate the biological function of VcLOB107 in blueberries, we overexpressed this gene in tomato. Three independent transgenic lines were selected to analyze the relative expression level of VcLOB107, and to find the phenotypic characteristics of tomato fruits at different ripening stages (Fig. 10). The fruit firmness, PG activity, and WSP content of transgenic lines were also detected (Fig. 11). Phenotypic analysis revealed that tomato fruits overexpressing VcLOB107 turned yellow and ripened faster than the control group. The expression level of VcLOB107 in transgenic plants was significantly higher than in the control, ranging from 3.5 in VcLOB107-OE12 to 5.3 in VcLOB107-OE10, with an average expression value of 4.5 (Fig. 11a). Transgenic tomatoes exhibited an average fruit firmness of 409.46 g·mm−1, significantly higher than that of the control. Furthermore, PG activity and WSP content in the transgenic lines was lower than that in the control group (Fig. 11b, c & d). These findings suggest that VcLOB107 regulates fruit firmness by modulating PG activity and subsequently influencing WSP content. Phenotypic comparisons between transgenic and control tomatoes further demonstrated that fruits overexpressing VcLOB107 switched into the mature green stage earlier than those of the control group.

Figure 10.
Fruit phenotype alteration of the overexpressed tomato. MG: mature green, BR: breaker stage, BR+3: 3 d after breaker stage, BR+7: 7 d after breaker stage, BR+15: 15 d after breaker stage. dpa: days post-anthesis.
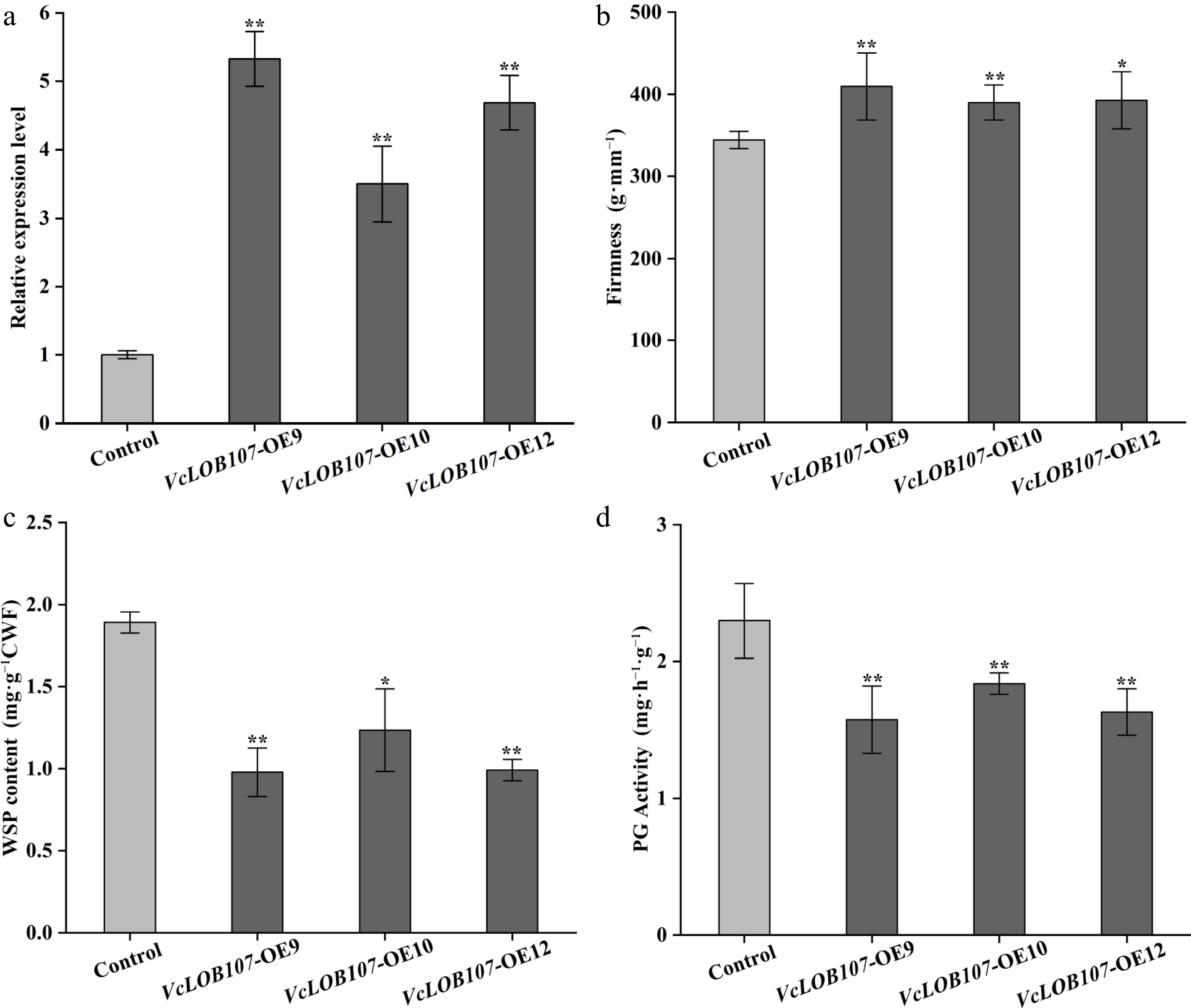
Figure 11.
VcLOB107 expression, fruit firmness, WSP content, and PG activities in the overexpressed tomato fruits (measured at 15 d after breaker stage). (t-test, * p < 0.05, ** p < 0.01).
-
LOB proteins play a pivotal role in the development of lateral organs and hormonal metabolic pathways in higher plants. The LOB genes were identified in multiple species, with 43 and 46 LOB members in Arabidopsis[41], and tomato[20], respectively, 35 members in rice[41], 67 members in apple[42], and 57 members in poplar[43]. One hundred and fifty LOB genes were identified in the blueberry genome in the present study, which is a notably higher number than that reported in other species. Previous studies indicated that blueberry has undergone at least two whole-genome duplication (WGD) events during its speciation[44], which have been crucial for the rapid expansion and evolution of the blueberry gene families. Hence, it is speculated that the amplification of LOB genes in blueberry was caused by WGD events experienced during its evolutionary history. Furthermore, we also discovered that 97.8% of the collinear gene pairs within the family exhibit a Ka/Ks ratio of less than 1, indicating that external pressures also played a significant role in the expansion of the VcLOB gene family.
Potential functions of VcLOB33, VcLOB74, VcLOB107, and VcLOB114 in blueberry
-
Previous studies have indicated that LOB proteins are involved in the regulation of lateral organ development in higher plants[45], including the formation of flowers, stems, leaves[18,46], and fruit development[22]. Tissue-specific gene expression can lay the foundation for a deeper understanding of their functions in growth and development processes. In this study, we analyzed the expression patterns of VcLOB genes in blueberry stems, leaves, flowers, and fruit, with different VcLOB genes displaying relatively high expression levels in specific tissues (Supplementary Fig. S3). VcLOB33 and VcLOB74 showed higher relative expression in leaves, suggesting a function related to leaf development; VcLOB114 and VcLOB107 were highly expressed in flowers and stems, respectively, suggesting a potential role in flowering and stems in addition to fruit firmness. A cluster analysis of the aforementioned four genes with functionally identified LOB proteins from other species (Supplementary Fig. S4), showed that VcLOB107 clusters with MdLOB13, which acts as a negative regulator in the anthocyanin biosynthesis process in apples[47], suggesting that VcLOB107 may have a similar function in blueberries; VcLOB33 and VcLOB114 cluster with SlLOB40, which is involved in the jasmonic acid methyl ester (JA) signaling pathway, and plays a negative regulation role in tomato drought resistance[48], suggesting that VcLOB33 and VcLOB114 may be involved in blueberry JA signaling transduction and drought resistance pathways; VcLOB74 clusters with AtLOB39, implicated in anthocyanin synthesis pathway and nitrate metabolism in Arabidopsis[49], leading to the conjecture that VcLOB74 may perform a similar role in blueberries.
VcLOB107 delays the softening of blueberry fruit
-
Polygalacturonase acts on demethylated polygalacturonic acid residues, causing changes in cell wall structure and thereby promoting fruit softening[50]. In this study, we found that PG activity and WSP content of 'O'Neal' fruits were significantly higher than that of 'Star' at stages S5 to S8 of blueberry fruit development, while fruit firmness was significantly lower than that of 'Star'. A gradual increase in PG activity during fruit development and ripening was also found in pear and tomato[51]. PG activity is closely related to the expression of PG genes. During the softening of strawberries[52], the expression of the FvPG1 gene increases, leading to a rise in PG activity and a consequent reduction of fruit firmness. The PG2 gene is highly expressed in pears[53], playing an important role in the degradation of fruit pectin and softening of the fruit flesh. Meanwhile, LOB can regulate the expression of downstream genes, such as the induction of EXP and other cell wall-degrading enzyme coding genes by transient expression of CsLOB1[54]. The VcLOB107 gene was significantly upregulated in the later stages of 'O'Neal' blueberry fruit development and downregulated in the later stages of 'Star' blueberry fruit development. The differential expression patterns of VcLOB107 between the two blueberry cultivars could be one of the reasons causing the difference in fruit firmness. Similar results were found in tomato that SlPG2a was notably up-regulated in SlLOB1 RNAi lines[22,55]. Therefore, we hypothesize that the difference in the fruit firmness of blueberry fruit between the two cultivars is partially attributed to the differential expression pattern of the VcLOB107 gene, which leads to variations in the expression levels of downstream VcPGs genes, ultimately resulting in disparities in the WSP content. However, further studies are needed to explore the regulation possibility of VcLOB and VcPGs genes.
LOB genes can respond to plant hormones including auxins[56], brassinosteroids (BR)[57], and jasmonic acid (JA)[58]. It is possible that the hormone content in 'Star' and 'O'Neal' blueberry fruit is different during the same period, leading to different extents of response to plant hormones by the LOB genes, hence resulting in different expression patterns. Our preliminary study has found significant differences in the levels of indole-3-acetic acid (IAA) during the growth and development of the two blueberry cultivars (unpublished data). Treatment with exogenous auxin analogs such as 1-naphthaleneacetic acid (NAA), and indole-3-butyric acid (IBA) on blueberry fruit at specific developmental stages could reveal the regulatory effects of auxins on LOB genes, and further elucidate the regulatory mechanism behind blueberry fruit firmness in the future. The transcription factor VcLOB107 delayed the softening of blueberry fruit possibly by regulating PG gene expression, thereby maintaining fruit firmness. Leveraging the role of VcLOB107 in delaying fruit softening, and an early answer that VcLOB33 interacted with the VcPG102 promoter (unpublished data), future studies could identify more VcLOB genes and molecular markers associated with VcLOB107 expression through association analysis, accelerating the breeding of firm-flesh blueberry cultivars. Furthermore, bulked segregant analysis (BSA) could be employed in the filial generation of 'Star' and 'O'Neal' to pinpoint key functional loci linked to VcLOB107, providing additional genomic regions and candidate targets for fruit firmness.
-
The 150 VcLOB genes identified in the blueberry genome were unevenly distributed on 46 chromosomes. 303 collinear gene pairs were found and most of the VcLOB family members had experienced purifying selection pressure. VcLOB107, VcLOB33, VcLOB114, and VcLOB74 showed different expression patterns between firm and soft flesh blueberry cultivars during fruit development. The fruit firmness of 'O'Neal' was significantly lower than that of 'Star' at stages S5 to S8, while the PG activity and WSP content were significantly higher than those of 'Star'. VcLOB107 overexpression in tomatoes had significantly higher fruit firmness, and lower PG activity and WSP content than that of the control. VcLOB107 genes acted as an important factor of fruit firmness difference between blueberry cultivars.
This work was supported by National Key Research and Development Program of China (Grant No. 2024YFE0214000), the Natural Science Foundation of Zhejiang Province, (Grant No. ZCLY24C1502), and the Key Research Project of Department of Science and Technology of Zhejiang Province, (Grant No. 2021C02066-9).
-
The authors confirm contributions to the paper as follows: study conception and design: Zong Y, Guo W; data collection: Huang Q, Dong J, Hu Y; analysis and interpretation of results: Huang Q, Lu X, Li Y, Xu L, Sun P; draft manuscript preparation: Huang Q, Zong Y. All authors reviewed the results and approved the final version of the manuscript.
-
The RNA-seq dataset of blueberry fruit ripening generated in this study has been deposited in the National Genomics Data Center (NGDC) database and is accessible through the GSA accession number CRA010224 (https://bigd.big.ac.cn/gsa/browse/CRA010224). All data analyzed during this study are included in its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 LOB gene accession numbers in Arabidopsis thaliana and blueberry.
- Supplementary Table S2 VcLOB primers for quantitative PCR.
- Supplementary Table S3 Primers used for VcLOB107 coding sequence cloning.
- Supplementary Table S4 The Ka/Ks distribution of VcLOB genes.
- Supplementary Table S5 Conserved motifs of VcLOB proteins.
- Supplementary Table S6 TPM values of VcLOB genes.
- Supplementary Fig. S1 Fruit firmness of 'Star' and 'O'Neal' at different fruit development stages.
- Supplementary Fig. S2 Schematic diagram of overexpression vector of VcLOB107.
- Supplementary Fig. S3 Relative expression of VcLOB genes in different organs of blueberry.
- Supplementary Fig. S4 Phylogenetic tree of LOB proteins in blueberry and other species.
- Copyright: © 2025 by the author(s). Published byMaximum Academic Press, Fayetteville, GA. This articleis an open access article distributed under Creative CommonsAttribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang Q, Dong J, Hu Y, Lu X, Li Y, et al. 2025. VcLOB107 acts as an important regulator of fruit firmness differences between firm and soft flesh blueberries. Fruit Research 5: e030 doi: 10.48130/frures-0025-0021 

VcLOB107 acts as an important regulator of fruit firmness differences between firm and soft flesh blueberries
- Received: 27 November 2024
- Revised: 21 April 2025
- Accepted: 29 April 2025
- Published online: 05 August 2025
Abstract: Fruit firmness is a significant trait for blueberry breeding as it affects machinery harvest, consumer preference, and transportability. LOB transcription factors play a vital role in the development of lateral organs and growth in higher plants; yet, few studies on LOB transcription factors have been reported in blueberries. We performed a genome-wide identification analysis of the blueberry LOB gene family. Using fruit at different developmental stages from the firm-flesh cultivar 'Star', and the soft-flesh cultivar 'O'Neal', the fruit firmness, polygalacturonase (PG) activity, and pectin content of different types were detected. A total of 150 VcLOB genes were identified in the blueberry genome, unevenly distributed across 46 chromosomes. They could be divided into six subfamilies, with members in each subfamily sharing similar gene structures and motifs. Based on the transcriptomic data and fruit firmness of two blueberry cultivars, four VcLOB genes functioning during the critical period of fruit firmness variation were selected, and their expression patterns were dissected, revealing differences in the expression pattern across various tissues and throughout the fruit development stages in blueberries. The fruit firmness of 'O'Neal' is significantly lower than that of 'Star' from stages S5 to S8 during fruit development. Additionally, the PG activity and water-soluble pectin content in 'O'Neal' is significantly higher than those in 'Star'. VcLOB107 overexpressed tomato plants exhibited notably higher fruit firmness, lower PG activity, and WSP content compared to the control group, indicating VcLOB107 is a regulator of fruit firmness differences between firm and soft flesh blueberries.
-
Key words:
- Lateral organ boundaries domain /
- Fruit firmness /
- Blueberry /
- Genome-wide identification