









-
Global climate change has led to more frequent extreme temperatures and water resource scarcity, making drought a major environmental factor that restricts plant growth and production[1]. Drought stress hinders photosynthesis, exacerbates oxidative stress, disrupts cell membrane integrity, and disturbs ion homeostasis within plants, ultimately stunting growth and resulting in mortality[2]. In addition, drought inhibits enzyme activities and affects leaf development, disrupting normal physiological and metabolic processes[3]. Blueberries (Vaccinium spp.) are among the most beneficial fruits due to their high contents of flavonoids and other bioactive compounds, which exhibit antioxidant, anti-aging, anti-cancer, and vision-enhancing properties[4,5]. However, due to their shallow and slender root system, blueberries struggle to absorb water from deeper soil layers, leading to high soil moisture requirements and increased susceptibility to drought stress[6,7]. Under drought stress, blueberry branches tend to thin and weaken, leaves may turn red, and are prone to early defoliation, thereby adversely affecting normal plant growth[8,9]. Drought stress significantly reduces fruit set in blueberries, leading to a marked decrease in fruit yield and posing a serious threat to blueberry production[10]. Therefore, enhancing drought tolerance in blueberries is crucial for safeguarding yield and quality, as well as promoting industrial development.
During drought stress, plants initiate a series of complex adaptive mechanisms to cope with the impacts of water deficit[11]. Reactive oxygen species (ROS), acting as signaling molecules involved in diverse plant biometabolic processes, play a crucial role in plants' defense against drought stress. 'ROS' is a collective term for oxygen-containing free radicals and peroxides prone to free-radical formation, which are associated with oxidative metabolism in living organisms[12]. Under normal growth conditions, plants maintain a dynamic balance between ROS accumulation and scavenging. However, when drought stress disrupts this balance, a large amount of ROS accumulates, affecting membrane structure and function and leading to membrane damage. To sustain normal growth, plants regulate their antioxidant systems to maintain ROS homeostasis. For instance, TaWD40-4B.1C interacts with catalases to reduce H2O2 levels under drought conditions, thereby enhancing drought tolerance in wheat (Triticum aestivum L.)[13]. In poplar (Populus ussuriensis), overexpression of PtrGSTU23 increases glutathione S-transferase (GST) activity while reducing ROS accumulation, leading to improved drought tolerance[14]. In apple (Malus sieversii), MsNAC022 activates MsPOD, enhancing ROS-scavenging enzyme activity and strengthening drought tolerance[15]. Therefore, identifying ROS-scavenging genes in blueberries is of great research value for mitigating oxidative damage and improving drought tolerance.
The MYB transcription factor family is one of the largest in plants and plays a crucial role in various processes, including growth, development, secondary metabolism, and responses to biotic and abiotic stresses[16,17]. Previous studies have demonstrated that MYB transcription factors are involved in activating ROS scavenging mechanisms, thereby enhancing tolerance to abiotic stress[18]. For instance, the apple (Malus domestica) transcription factor MdMYB23 binds to the promoter of MdANR, activating its expression, which promotes proanthocyanidin accumulation and ROS scavenging in apple calli, thus improving cold tolerance[19]. Additionally, MYB88 and MYB124 facilitate anthocyanin accumulation and reduce ROS levels, positively regulating cold tolerance and the expression of low-temperature response genes in apples[20]. In upland cotton (Gossypium hirsutum), GhMYB3 negatively regulates drought tolerance; silencing this gene promotes the biosynthesis of anthocyanins and proanthocyanidins, reducing ROS accumulation and improving drought tolerance[21]. TT2-type MYB transcription factors are well known for their role in proanthocyanidins biosynthesis[22−25]. Recent studies also suggest their involvement in plant response to environmental stress. For instance, overexpression of BpMYB123 enhances drought tolerance in birch (Betula platyphylla) by upregulating BpLEA14 expression, thereby improving cell membrane stability[26]. Similarly, heterologous expression of cotton GhTT2 in Arabidopsis increases proanthocyanidin content and activates stress-responsive genes, enhancing osmotic stress tolerance in transgenic seedlings[27]. While TT2 function has been characterized in other species, its role in blueberry drought tolerance remains unexplored.
In this study, a TT2-type MYB transcription factor from blueberry, VcTT2, was identified, which responds to drought, salt, and low-temperature stresses. While VcTT2 overexpression did not enhance salt or low-temperature tolerance, it significantly improved drought tolerance in blueberries by upregulating VcAPX1, VcAPX2, VcSOD1, VcSOD2, VcPOD1, and VcPOD2 under drought stress, thereby increasing APX, SOD, and POD activities to scavenge excess ROS. Conversely, suppression of VcTT2 reduced drought tolerance. These findings establish VcTT2 as a key regulator of drought tolerance in blueberries, providing valuable insights into the underlying mechanisms and offering a crucial genetic resource for breeding drought-resistant cultivars.
-
The seedlings of northern highbush blueberry (Vaccinium corymbosum) cultivar 'Legacy' were cultivated in a greenhouse with 16 h of light/8 h of darkness at 24 °C and 60%−70% relative humidity. Thirty-day-old seedlings were subjected to drought stress (10% PEG6000), salt stress (200 mM NaCl), and cold stress (4 °C). After 0, 3, and 12 h of treatment, the leaves and roots were frozen in liquid nitrogen and stored at −80 °C for RNA extraction. Each experiment included three biological replicates, with three technical replicates performed for each biological replicate.
Blueberry 'Legacy' seedlings used for genetic transformation were cultured in WPM medium containing 1.0 mg/L ZT in a greenhouse at 24 °C with 16 h of light/8 h of darkness and were sub-cultured once a month. As previously reported, 'Northland' blueberry calli used for genetic transformation were obtained through leaf wounding induction and cultured in WPM medium containing 1.0 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D) and 0.4 mg/L 6-benzylaminopurine (6-BA) in the dark at 24 °C, with subculturing every two weeks[28].
RNA extraction and Real-Time Quantitative PCR (RT-qPCR)
-
Total RNA was extracted from calli, as well as leaves and roots of blueberry seedlings, using the Plant Tissue Total RNA Isolation Kit (Vazyme, China). cDNA synthesis was performed using the 5× All-In-One RT MasterMix (Applied Biological Materials, Canada). The PowerUp™ SYBR™ Green Premix Kit (Applied Biological Materials, Canada) was used for RT-qPCR, with VcUBC28 serving as the internal reference gene. Relative expression levels were calculated using the 2−ΔΔCᴛ method. The primer sequences were presented in Supplementary Table S1.
Analysis of cis-acting element of promoters
-
The cis-acting elements in the promoters of TT2-type MYBs (2,000 bp upstream of the transcription start site) were predicted and analyzed using PlantCARE website (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ).Transformation of blueberry calli and seedlings
-
The coding sequence (CDS) of VcTT2 was cloned and inserted into the pMDC85 vector to construct the overexpression vector 35S: VcTT2. Additionally, both the sense and antisense fragments of VcTT2 were inserted into the pRI101 vector to construct the 35S: VcTT2-RNAi suppression vector. According to previous studies, blueberry transgenic calli and seedlings were obtained using the Agrobacterium-mediated method[28,29]. The primer sequences employed in this study are detailed in Supplementary Table S1.
Different stress treatment of blueberry calli
-
A total of 0.1 g of wild-type (WT) and transgenic blueberry calli, which had been cultured for approximately 20 d, were weighed and placed on the succession medium as a control. These were then incubated in the dark at 26 °C for 10 d. Blueberry calli with similar growth were weighed to equal masses and treated with different stress. For drought stress, WT and transgenic calli were placed on the medium containing 5% and 10% PEG6000, respectively, and incubated in the dark at 26 °C for 10 d. For salt stress, the calli were placed on the medium containing 200 mM NaCl and incubated in the dark at 26 °C for 10 d. For low-temperature stress, the calli were placed in the dark at 4 °C for 10 d.
Natural drought treatment of blueberry transgenic plants
-
WT and transgenic blueberry seedlings were transplanted into pots and cultivated at 24 °C with 16 h of light/8 h of darkness for 30 d. Thirty-day-old WT and transgenic seedlings were dehydrated for 20 d. Subsequently, the phenotypes were observed, and samples were collected to determine physiological parameters. Each experiment included three biological replicates, with three technical replicates performed for each biological replicate.
Diaminobenzidine (DAB) and 3,3'-Nitrotetrazolium blue chloride (NBT) staining
-
Leaves of WT and transgenic blueberry seedlings under normal growth and drought stress were stained with DAB (Nanjing Jiancheng, China) and NBT solution (Nanjing Jiancheng, China) for 2 h. Photographs were taken after decoloration with 95% ethanol solution. The NBT staining time of the calli under different stress was 20 min. The brown and blue spots represented the production of H2O2 and O2·−, respectively[30,31]. Each experiment included three biological replicates, with three technical replicates performed for each biological replicate.
Measurement of physiological parameters
-
Following drought treatment, 0.1 g of blueberry calli or leaves from WT and transgenic lines were weighed and ground in liquid nitrogen for the measurement of physiological parameters. Malondialdehyde (MDA) content was determined by the thiobarbituric acid measurement method[6]. Hydrogen peroxide (H2O2) content was determined using the KI method[32]. Proline content was measured using the sulfosalicylic acid method[33]. Superoxide dismutase (SOD) activity and ascorbate peroxidase (APX) activity were determined using NBT (nitrogen blue tetrazolium) photoreduction method[34]. Peroxidase (POD) activity was determined using the guaiacol method[35]. Relative water content, chlorophyll, and relative conductivity in leaves were determined according to previous studies[36−38]. Each experiment included three biological replicates, with three technical replicates performed for each biological replicate.
Statistical analysis
-
Each result was based on three parallel experiments with at least three technical replicates to ensure the accuracy of the results. Error bars show the standard deviation (SD) of three replicates. Statistically significant differences are analyzed using one-way ANOVA and Student's t-test with GraphPad Prism or Excel software, and denoted with different lowercase letters or with asterisks.
-
To identify TT2-type MYB transcription factors in blueberry, a BLASTP search using the amino acid sequence of AtTT2 was conducted to identify TT2-type MYB proteins in the Vaccinium corymbosum cv. Draper v1.0 genome. A total of 509 TT2-type MYB proteins were identified, of which only 18 contained the SG5 motif (Supplementary Table S2). Based on the PlantCARE website, cis-acting elements in the promoter were predicted and analyzed. The results revealed that the promoter of maker-VaccDscaff32-augustus-gene-55.27 contained both a low-temperature responsive element and a defense and stress-responsive element (Fig. 1a), and it had the highest number of these elements among the 18 TT2-type MYB promoters (Supplementary Fig. S1). This suggests that the gene may be involved in the response of abiotic stresses. Thus, this gene was designated as VcTT2 (also known as VcMYBPA2.1).
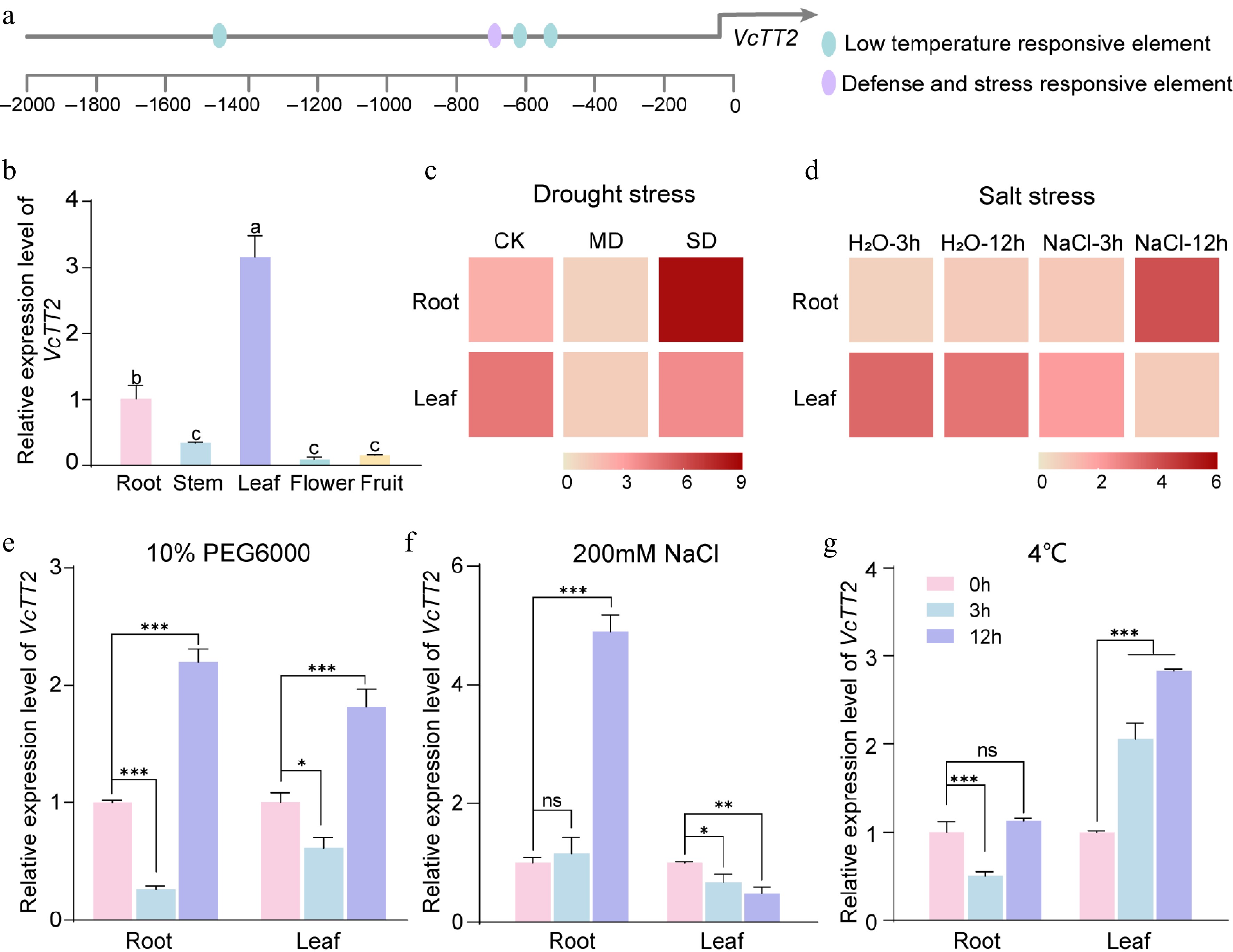
Figure 1.
VcTT2 responds to drought, salt, and low-temperature stress. (a) Schematic drawing depicting the cis-acting elements related to stress-responsiveness in the promoter of VcTT2. (b) RT-qPCR assay showing the relative expression levels of VcTT2 in various tissues of blueberry, including root, stem, leaf, flower and fruit. Error bars represent the standard deviation of three biological replicates. Different letters above the error bars indicate significant differences at the p < 0.05 level (one-way ANOVA). (c) Heatmap showing the expression profiles of VcTT2 in blueberry roots and leaves under different levels of drought stress. Normal watering treatment (75%−80% soil water content (SWC)) was used as control (CK), 55%−60% SWC as moderate stress (MD), and 30%−35% SWC as severe stress (SD). (d) Heatmap showing the expression profiles of VcTT2 in blueberry roots and leaves under salt stress at different time points. Salt stress was simulated with 200 mM NaCl treatment, and water treatment was used as control. RT-qPCR assay of relative expression levels of VcTT2 in blueberry leaves and roots under (e) drought stress, (f) salt stress, and (g) low temperature. Drought stress was simulated by 10% PEG6000 treatment, salt stress by 200 mM NaCl treatment, and low-temperature stress by 4 °C treatment. Asterisks denote statistical significance determined by Student's t-test, * p < 0.05, ** p < 0.01, *** p < 0.001.
To determine the tissue-specific expression of VcTT2, its expression was measured in blueberry roots, stems, leaves, flowers, and fruits. RT-qPCR analysis showed that VcTT2 was expressed in all plant tissues, with the highest expression levels observed in leaves and roots (Fig. 1b). Transcriptome profiling of blueberry seedlings under drought stress was previously reported[39], as well as NaCl treatment[40]. Based on these transcriptome data, it was found that the expression of VcTT2 in both roots and leaves initially decreased and then increased with increasing drought stress (Fig. 1c). However, as the duration of NaCl treatment increased, VcTT2 expression was upregulated in roots but downregulated in leaves (Fig. 1d). To further investigate the response of VcTT2 to various abiotic stresses, its expression levels were measured in blueberry leaves and roots following treatments with 10% PEG6000, 200 mM NaCl, and 4 °C for 0, 3, and 12 h. The expression of VcTT2 in both roots and leaves increased under drought treatment (Fig. 1e). Under salt treatment, VcTT2 was significantly induced in roots, while its expression decreased in leaves (Fig. 1f). Under low-temperature treatment, VcTT2 expression in roots initially decreased and then recovered, showing no significant difference compared to the untreated control, while its expression in leaves was gradually upregulated (Fig. 1g). These findings suggest that VcTT2 expression is induced by drought, salt, and low-temperature stresses.
VcTT2 enhances drought tolerance in transgenic blueberry calli
-
To investigate the role of VcTT2 in regulating drought tolerance in blueberry, the VcTT2-overexpressing (VcTT2-OE) blueberry calli were generated. RT-qPCR analysis revealed that the expression level of VcTT2 was significantly upregulated in VcTT2-OE calli (Fig. 2b). WT and VcTT2-OE calli were then treated with different stresses, including drought stress (simulated with 5% PEG6000), salt stress (200 mM NaCl), and low-temperature stress (4 °C), while those grown under normal conditions were used as the control. Under control conditions, both WT and VcTT2-OE calli exhibited comparable growth, with no significant differences observed (Fig. 2a). Notably, under 5% PEG6000 treatment, VcTT2-OE calli showed superior growth compared to WT, with fresh weight approximately 1.6 times greater than that of WT (Fig. 2c). In contrast, 200 mM NaCl and 4 °C treatments significantly inhibited the growth of both WT and VcTT2-OE calli, showing no significant difference in fresh weight (Fig. 2a, d, e). These findings suggest that overexpression of VcTT2 enhances drought tolerance but not salt tolerance or low-temperature tolerance in blueberry calli.
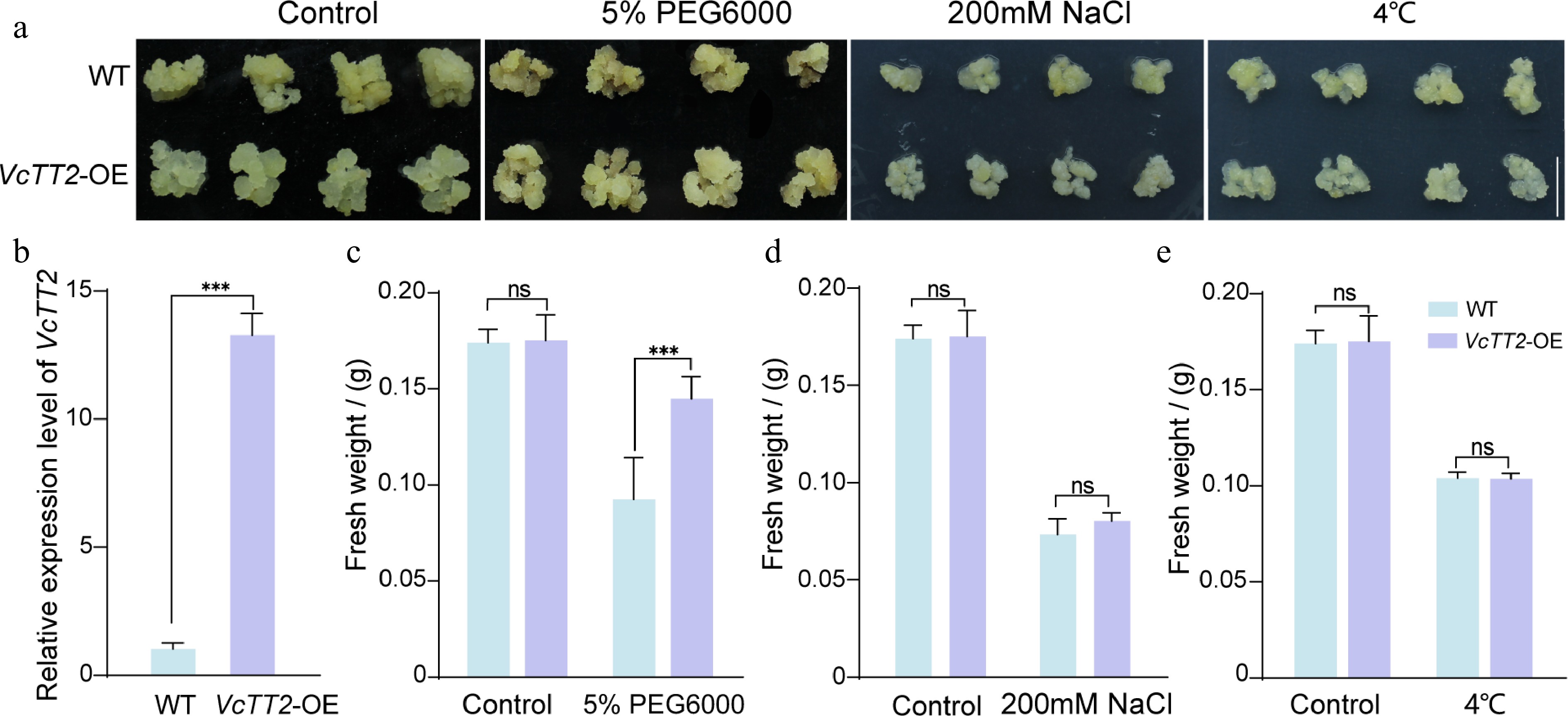
Figure 2.
Overexpression of VcTT2 enhances drought tolerance in blueberry calli. (a) Phenotypic comparison of wild-type (WT), and VcTT2-overexpressing (VcTT2-OE) blueberry calli under drought stress (5% PEG6000 treatment), salt stress (200 mM NaCl treatment), and low-temperature stress (4 °C treatment). WT and VcTT2-OE blueberry calli grown in normal conditions were used as controls. Scale bar = 1 cm. (b) RT-qPCR analysis of relative VcTT2 expression levels in WT and VcTT2-OE blueberry calli. Fresh weight of WT and VcTT2-OE calli following 10 d of (c) drought stress, (d) salt stress, and (e) low-temperature stress. Asterisks indicate statistical significance determined by Student's t-test, *** p < 0.001.
VcTT2 improves ROS scavenging ability in transgenic blueberry
-
It has been reported that MYB transcription factors can enhance drought tolerance in plants by improving their ability to scavenge ROS[41]. To investigate whether VcTT2 promotes drought tolerance by scavenging ROS under drought stress in blueberry, WT and VcTT2-OE calli were stained with NBT after 5% and 10% PEG6000 treatment to assess H2O2 levels. The staining results showed that both WT and VcTT2-OE calli exhibited lighter coloration under control conditions. Compared with 5% PEG6000 treatment, NBT staining intensified in both WT calli and VcTT2-OE calli when the concentration of PEG6000 was increased to 10%, but the staining in VcTT2-OE calli remained lighter than in WT calli (Fig. 3a). To further confirm whether VcTT2 enhances drought tolerance by scavenging ROS, the levels of the contents of malondialdehyde (MDA), hydrogen peroxide (H2O2), and the osmoregulatory substance proline were measured in both WT and VcTT2-OE calli. The results showed that although the contents of these three substances increased after 5% PEG6000 treatment, the proline content in VcTT2-OE calli was higher than that in WT calli, whereas the MDA and H2O2 levels were significantly lower in VcTT2-OE, with the H2O2 content being approximately half that of WT (Fig. 3d). When the calli were subjected to 10% PEG6000 treatment, the MDA and H2O2 levels in VcTT2-OE calli remained lower than in WT, while the proline content significantly increased to 1.68 times that of WT (Fig. 3b−d). These results suggest that the VcTT2-OE blueberry calli were less damaged than WT calli under drought stress.
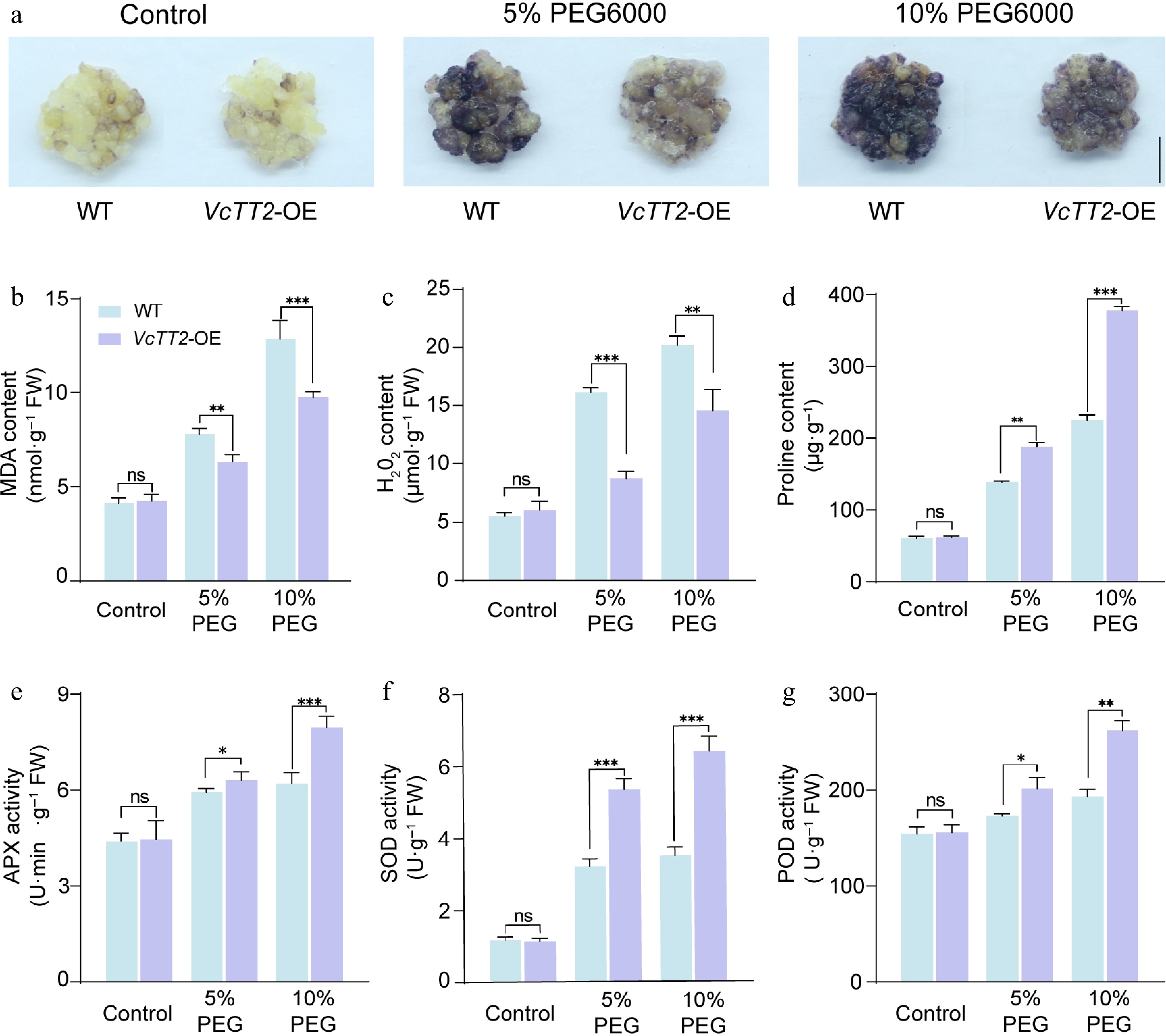
Figure 3.
Overexpression of VcTT2 enhances the ability of blueberry calli to scavenge reactive oxygen species (ROS). (a) NBT staining of wild-type (WT) and VcTT2-overexpressing (VcTT2-OE) blueberry calli under drought stress simulated by 5% and 10% PEG6000 treatments. WT and VcTT2-OE calli grown under normal conditions served as controls. Scale bar = 1 cm. Contents of (b) MDA, (c) H2O2, (d) proline, and activities of antioxidant enzymes (e) APX, (f) SOD, and (g) POD in WT and VcTT2-OE blueberry calli under drought stress as shown in (a). Asterisks indicate statistical significance determined by Student's t-test, * p < 0.05, ** p < 0.01, *** p < 0.001.
To determine whether overexpressing VcTT2 enhances drought tolerance in blueberry calli by increasing antioxidant enzymes activities, the activities of ascorbate peroxidase (APX), superoxide dismutase (SOD), and peroxidase (POD) were measured. The results showed that the activities of APX, SOD, and POD were significantly higher in VcTT2-OE calli compared to WT calli after 5% or 10% PEG6000 treatments. Notably, after 10% PEG6000 treatment, the SOD enzyme activity in VcTT2-OE calli was 1.83 times higher than in WT (Fig. 3e−g). These findings indicate that overexpression of VcTT2 effectively enhances drought tolerance in blueberry calli by improving their ability to scavenge ROS under drought stress.
VcTT2 positively regulates drought tolerance in transgenic blueberry seedlings
-
To further investigate the effect of VcTT2 on drought tolerance in blueberry plants, transgenic seedlings overexpressing VcTT2 (VcTT2-OE#1, 2, 3, 4) and suppressing VcTT2 (VcTT2-RNAi#1, 2, 3, 4), were generated. RT-qPCR showed that the expression level of VcTT2 was significantly upregulated in the VcTT2-OE seedlings while downregulated in VcTT2-RNAi seedlings (Fig. 4b). WT, VcTT2-OE, and VcTT2-RNAi blueberry seedlings were then subjected to natural drought treatment, with normal watering treatment used as the control. Twenty days after watering was stopped, VcTT2-OE lines exhibited better growth and significantly less wilting compared to WT#2, while VcTT2-RNAi lines displayed yellowing leaves and the greatest wilting (Fig. 4a). Under drought treatment, the relative electrical conductivity of VcTT2-OE leaves increased to approximately 29%, which was significantly lower than that of the WT (Fig. 4c). The relative water content of VcTT2-OE leaves was about 1.26 times greater than that of WT, while the chlorophyll content was approximately 1.28 times higher than that of WT (Fig. 4d, e). Conversely, the relative water content and chlorophyll content of VcTT2-RNAi leaves were significantly lower than those of WT, whereas the relative electrical conductivity was higher than that of WT. These data suggest that VcTT2 enhances the drought tolerance of blueberry seedlings.
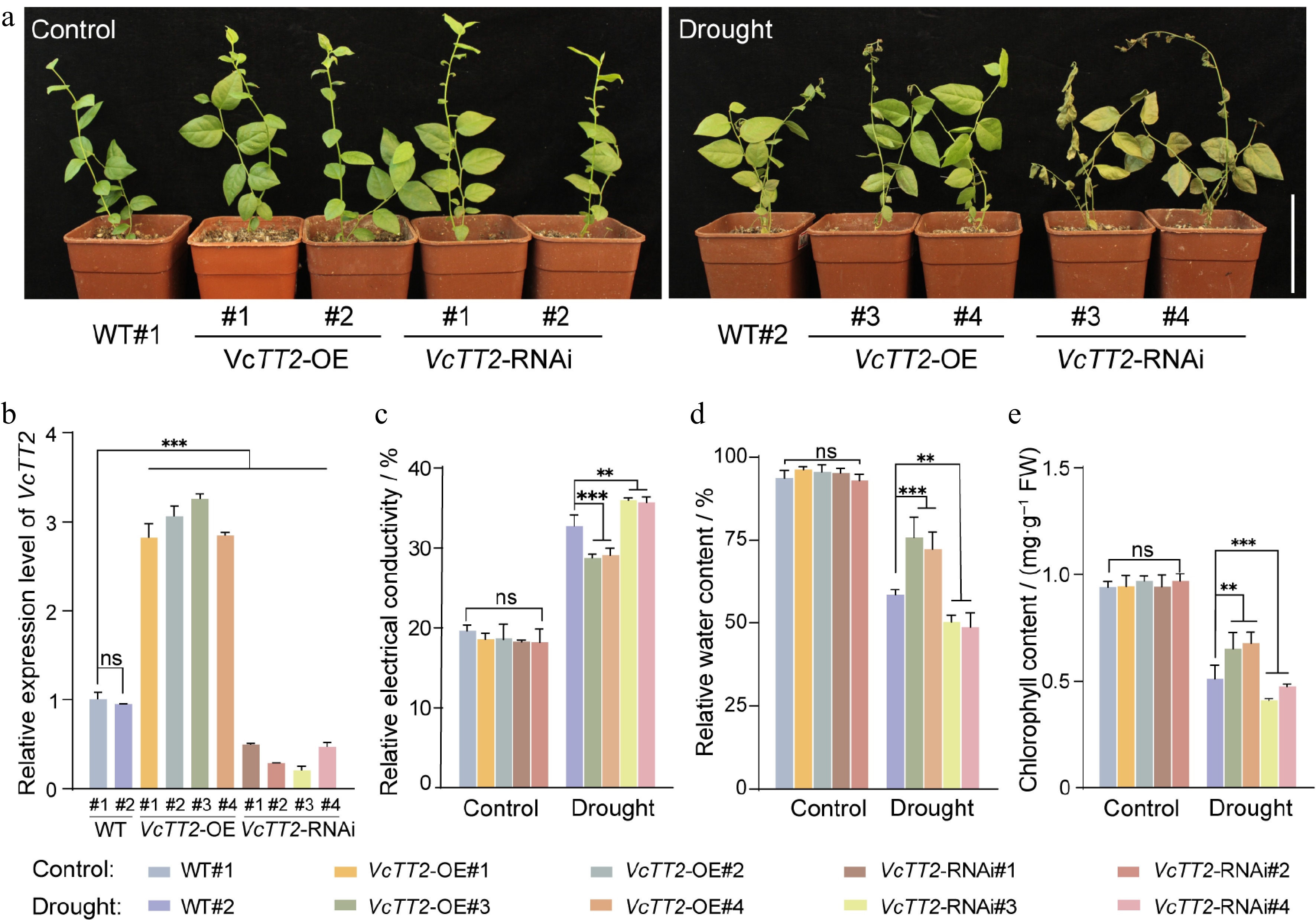
Figure 4.
VcTT2 positively regulates drought tolerance in blueberry seedlings. (a) Phenotypic comparison of wild-type (WT) and transgenic blueberry seedlings (VcTT2-OE and VcTT2-RNAi) after 20 days of natural drought treatment, with normal watering treatment used as the control. VcTT2-OE: VcTT2-overexpression; VcTT2-RNAi: VcTT2-suppression. Scale bar = 10 cm. (b) RT-qPCR analysis of relative VcTT2 expression levels in WT and transgenic blueberry seedlings. (c) Relative electrical conductivity, (d) relative water content, and (e) chlorophyll content of WT and transgenic blueberry seedlings under natural drought treatment, as shown in (a). Asterisks indicate statistical significance determined by Student's t-test, **p < 0.01, ***p < 0.001.
VcTT2 positively regulates the ability of transgenic blueberry seedlings to scavenge ROS
-
To further investigate whether VcTT2 enhances the ability to scavenge ROS under drought stress, the accumulation of O2·− and H2O2 in the middle leaves from WT and VcTT2 transgenic seedlings were measured using NBT and DAB histochemical staining, respectively. The staining results showed no significant differences among all leaves under control conditions. However, under drought stress, OE#4 leaves exhibited lighter staining compared to WT, while RNAi#4 leaves showed darker staining (Fig. 5a, b), suggesting that VcTT2 may inhibit ROS accumulation. Quantification of ROS levels revealed that H2O2 and MDA contents were lower in OE#4 leaves than that in WT under drought stress (Fig. 5c, d), while the proline content was approximately 1.75 times greater than in WT and significantly lower in RNAi#4 (Fig. 5e). To further verify whether overexpressing VcTT2 enhances drought tolerance by increasing antioxidant enzyme activities in blueberry seedlings, the activities of antioxidant enzymes APX, SOD and POD in the leaves were measured. Under drought stress, the activities of APX, SOD, and POD in OE#4 leaves were significantly higher than those in WT, whereas RNAi#4 leaves exhibited the opposite trend (Fig. 5f−h). These results suggest that VcTT2 may scavenge ROS by enhancing the activities of antioxidant enzymes, thereby improving the drought tolerance of blueberry.
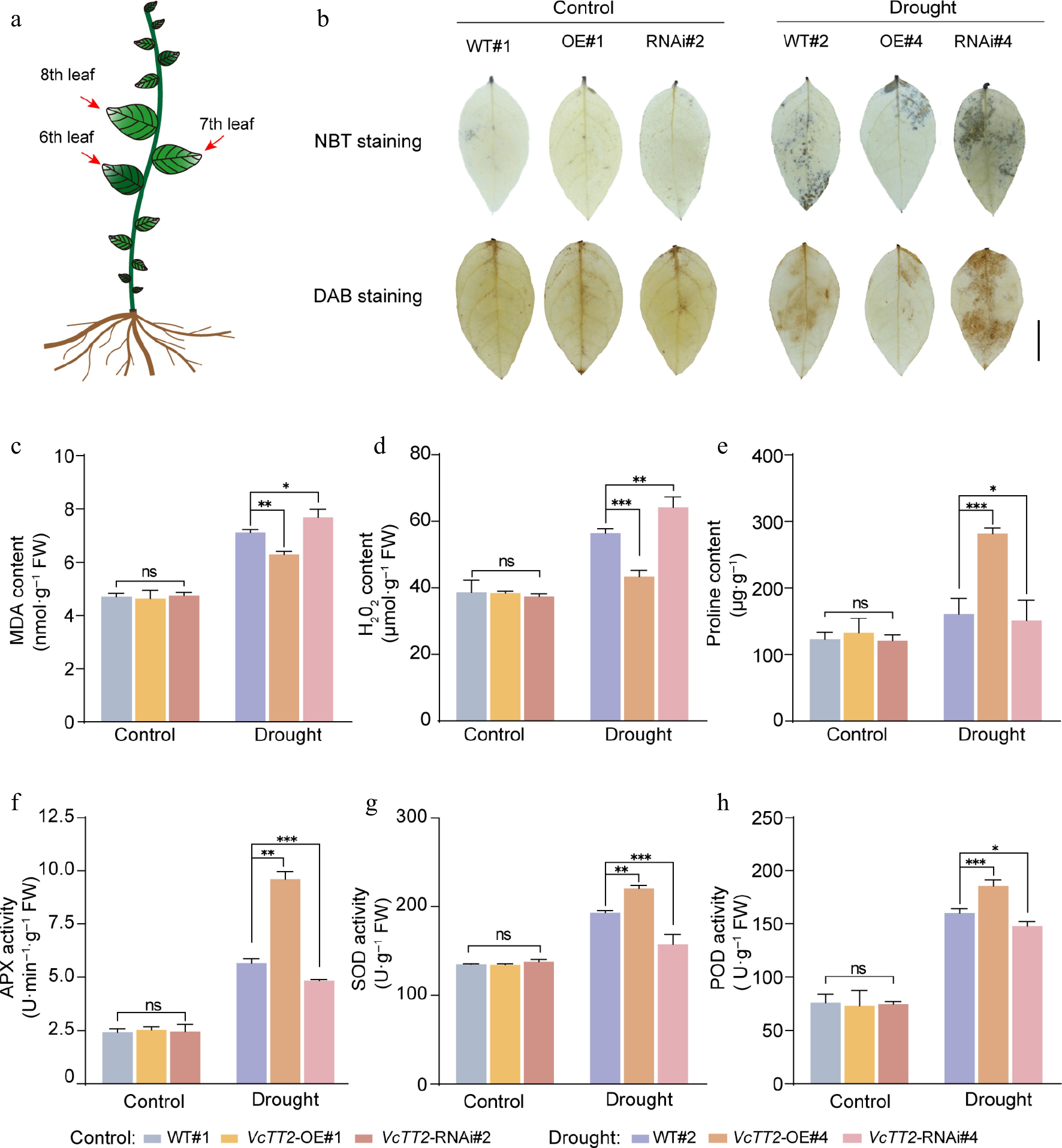
Figure 5.
VcTT2 positively regulates the ability of blueberry seedlings to scavenge ROS. (a) The red arrows indicate the experimentally sampled leaves. (b) DAB and NBT staining of leaves from wild-type (WT) and VcTT2-overexpressing (VcTT2-OE) blueberry seedlings after 20 d of natural drought treatment. Contents of (c) MDA, (d) H2O2, (e) proline, and activities of antioxidant enzymes (f) APX, (g) SOD, and (h) POD in leaves of WT and VcTT2-OE blueberry seedlings under drought stress, as shown in (a). Asterisks indicate statistical significance determined by Student's t-test, * p < 0.05, ** p < 0.01, *** p < 0.001.
VcTT2 promotes the expression of antioxidant enzyme genes under drought stress
-
To investigate whether VcTT2 enhances antioxidant enzyme activities by regulating the expression of antioxidant enzyme genes under drought stress, the expression levels of VcSOD1, VcSOD2, VcPOD1, VcPOD2, VcAPX1, and VcAPX2 were measured in the leaves of WT and transgenic blueberry seedlings subjected to natural drought treatment. Under control conditions, the expression levels of VcSOD2, VcPOD2, VcAPX1, and VcAPX2 in OE#4 leaves were significantly higher than those in WT (Fig. 6b, d−f). However, no significant differences were found in the expression levels of VcSOD1 and VcPOD2 between WT and OE#4 (Fig. 6a, c). Notably, under drought stress, the expression of all enzyme genes was significantly upregulated in the OE#4, with VcPOD2 expression being approximately 37 times higher than that in WT (Fig. 6d). These results suggest that under drought stress, VcTT2 promotes the expression of antioxidant enzyme genes (especially VcPOD2), increases antioxidant enzyme activities, and enhances ROS scavenging, thereby improving drought tolerance in blueberry.
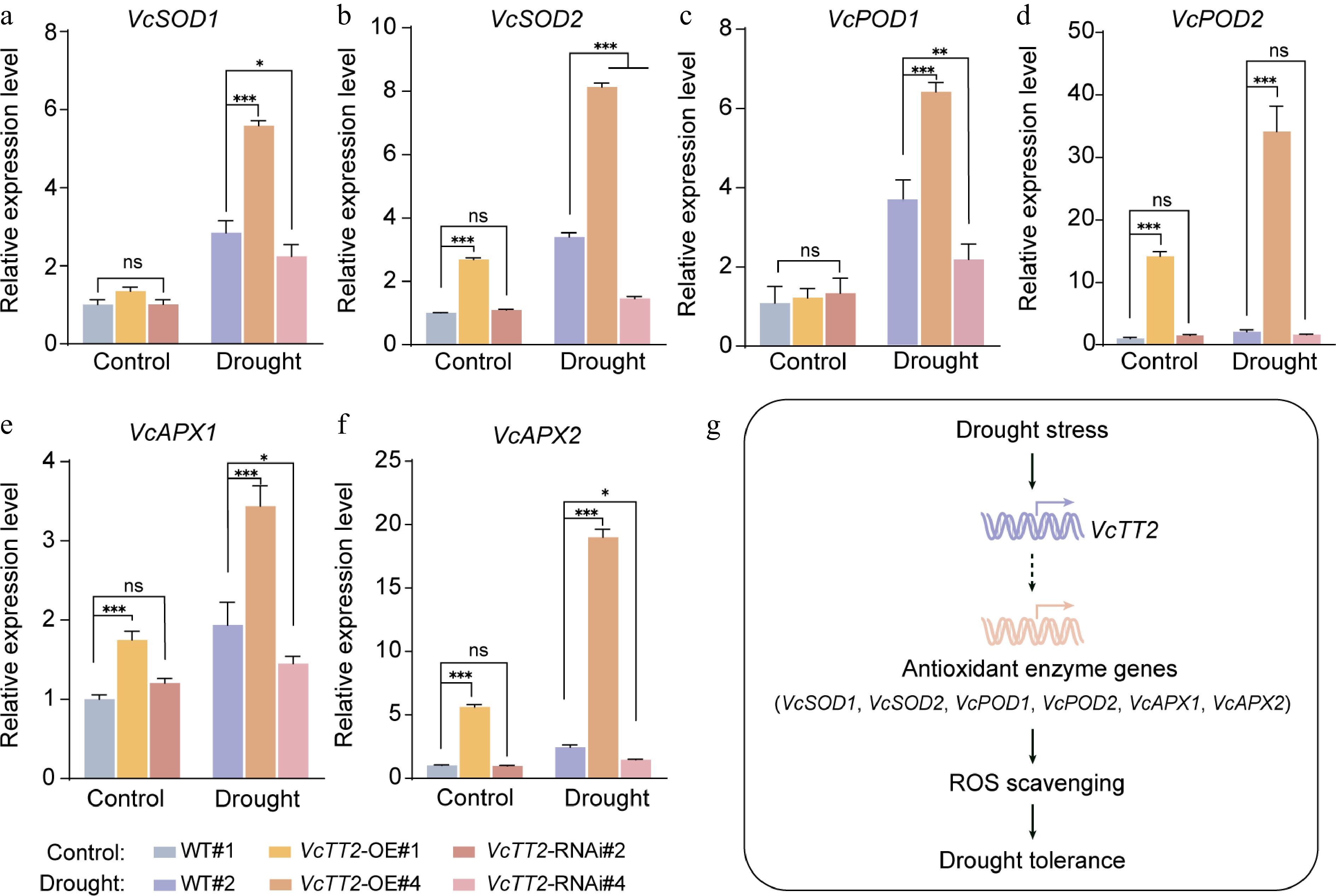
Figure 6.
VcTT2 promotes the expression of antioxidant enzyme genes under drought stress. RT-qPCR analysis of relative expression levels of (a) VcSOD1, (b) VcSOD2, (c) VcPOD1, (d) VcPOD2, (e) VcAPX1, and (f) VcAPX2 in leaves from wild-type (WT) and VcTT2-overexpressing (VcTT2-OE) blueberry seedlings after 20 d of natural drought treatment. Asterisks indicate statistical significance determined by Student's t-test, * p < 0.05, ** p < 0.01, *** p < 0.001. (g) The working model of VcTT2 regulation of blueberry drought tolerance. Black arrows indicate positive regulation.
Based on these findings, a working model in which drought stress induces the expression of VcTT2 is proposed, leading to the upregulation of antioxidant enzyme genes, including VcSOD1, VcSOD2, VcPOD1, VcPOD2, VcAPX1, and VcAPX2. This upregulation enhances the activities of antioxidant enzymes (SOD, POD, and APX), improves the plant's ability to scavenge ROS, and ultimately increases drought tolerance in blueberry (Fig. 6g).
-
Drought stress is a major constraint on global agricultural sustainability, posing a significant challenge to crop productivity[42,43]. Previous studies have demonstrated that TT2-type MYB transcription factors play a crucial role in regulating anthocyanin and proanthocyanidin biosynthesis, with antioxidant and stress-resistant functions across various plants. For example, in rose (Rosa rugosa), the TT2-type MYB transcription factor RrMYB10 promotes proanthocyanidin accumulation in transgenic roses, enhancing their tolerance to injury and oxidative stress[44]. Similarly, in Panax ginseng, overexpression of PgTT2 in Arabidopsis positively regulates ABA biosynthesis and signaling, leading to enhanced ROS scavenging[45]. In Gossypium hirsutum, heterologous expression of GhTT2 in Arabidopsis increases proanthocyanidin content and induces stress-responsive genes, thereby improving osmotic stress tolerance[27]. Given that anthocyanins and proanthocyanidins act as effective antioxidants in scavenging ROS and protecting cells, it is hypothesized that VcTT2 regulates the drought-induced metabolic pathways of anthocyanins or proanthocyanidins in blueberry, thereby enabling blueberries adaption to drought conditions. Unlike these studies that used heterologous expression in Arabidopsis, this study focuses on the native function of VcTT2 in blueberries. A TT2-type MYB gene, VcTT2, was identified, whose expression was upregulated under drought, salt, and low-temperature stresses (Fig. 1c−e). Overexpression of VcTT2 significantly enhanced drought tolerance in blueberry calli but did not affect salt or low-temperature tolerance (Fig. 2). Furthermore, overexpression of VcTT2 improved the drought tolerance in blueberry seedlings, while suppression of VcTT2 reduced drought tolerance (Fig. 4). These results indicate that VcTT2 plays a positive role in drought stress tolerance in blueberry, sharing functional similarities to TT2-type MYB genes in other plants.
In rice (Oryza sativa), the expression of OsPrx114 is transiently downregulated under PEG6000 treatment, followed by a significant rebound, peaking at 9 h[46]. In this study, VcTT2 expression was similarly downregulated at 3 h of PEG6000 treatment but markedly upregulated at 12 h, aligning with the pattern observed for OsPrx114. It is hypothesized that the initial downregulation may serve as an energy conservation strategy during the early stages of drought stress. The subsequent upregulation likely reflects the activation of drought resistance mechanisms in blueberry seedlings, particularly those related to the antioxidant enzyme system.
Drought stress induces excessive accumulation of ROS, and an imbalance between ROS production and scavenging leads to oxidative damage and cell death. Therefore, maintaining ROS homeostasis is essential for alleviating oxidative stress and enhancing drought tolerance in plants[12,47,48]. Previous studies have shown that overexpression of PuC3H35 in poplar significantly reduced MDA, H2O2, and O2·− levels in roots under drought stress, thereby enhancing ROS scavenging and improving drought tolerance[49]. In this study, VcTT2 overexpression significantly reduced MDA and H2O2 levels in both blueberry calli and seedlings under drought stress compared to WT plants (Figs 3, 5), suggesting that VcTT2 effectively suppresses H2O2 accumulation, alleviates membrane lipid peroxidation, and minimizes cell membrane damage, thereby enhancing drought tolerance.
Proline is a crucial osmoprotectant that plays a key role in plant responses to abiotic stress. Its accumulation under drought stress helps mitigate dehydration by regulating cellular osmotic potential and maintaining turgor pressure[50]. Additionally, proline scavenges free radicals, interacts with ROS, reduces oxidative stress, and protects cellular structures[51]. Here, overexpression of VcTT2 significantly increased proline content in transgenic blueberry calli and seedlings, effectively preventing excessive cellular dehydration and further enhancing drought tolerance (Figs 3d & 5d). These findings are consistent with previous research on BplMYB46, which also enhanced proline accumulation by regulating P5CS expression to improve stress tolerance in poplar[52].
In the antioxidant enzyme system, SOD, POD, and APX play crucial roles in the catabolism of ROS[53]. Among these enzymes, the activity of SOD, a core metalloenzyme, is frequently utilized as a critical indicator of plant stress tolerance[54,55]. Overexpression PlMYB108 from herbaceous peony (Paeonia lactiflora) in tobacco has been shown to enhance flavonoid content and antioxidant enzyme activities, reducing ROS levels and improving water retention, thereby increasing drought tolerance[56]. Similarly, the cotton gene GbMYB5 has been reported to enhance drought tolerance by increasing antioxidant enzyme activities and activating the expression of stress-responsive genes[57]. Ectopic expression of the RrANR gene from rose in tobacco increased the activities of antioxidant enzymes, including POD, SOD, and catalase (CAT), by upregulating the expression of ROS scavenging-related genes, thereby improving the plant's ability to scavenge ROS and increasing its tolerance to oxidative stress[58]. It was also observed that under drought stress, the activities of SOD, POD, and APX were elevated in VcTT2-OE blueberry calli and seedlings. The expression levels of VcSOD1, VcSOD2, VcPOD1, VcPOD2, VcAPX1, and VcAPX2 were significantly upregulated in VcTT2-OE seedlings compared to WT, indicating that VcTT2 promotes the expression of antioxidant enzyme genes. This leads to enhanced antioxidant enzyme activities, effective scavenging of excessive ROS, regulation of cell redox homeostasis, and ultimately improved drought tolerance in blueberry. Notably, the expression levels of VcAPX, VcSOD2, and VcPOD2 were upregulated in VcTT2-OE blueberry seedlings under normal conditions, suggesting that these genes may be downstream targets of VcTT2. However, further investigation is needed to determine whether VcTT2 directly enhances the expression of VcAPX, VcSOD2, and VcPOD2 by binding to their promoters, thereby promoting drought tolerance in blueberry.
-
In summary, this study demonstrates the role of the TT2-type MYB gene, VcTT2, in enhancing drought tolerance in blueberry. VcTT2 was rapidly induced under drought stress, and its overexpression upregulated the expression of a series of antioxidant enzymes genes, including VcSOD1, VcSOD2, VcPOD1, VcPOD2, VcAPX1, and VcAPX2. This, in turn, enhanced antioxidant enzyme activities, effectively scavenged excessive ROS, attenuated oxidative damage, and improved drought tolerance in blueberry. These findings confirm the role of VcTT2 in enhancing drought tolerance from multiple perspectives, contributing to the understanding of the functional diversity of TT2-type MYB transcription factors in plant stress tolerance. In conclusion, this study provides valuable genetic resources for breeding drought-tolerant blueberry germplasm.
This work was supported by the Key R&D Program of Shandong Province (Grant No. 2024LZGCQY025), the Natural Science Foundation of Beijing (Grant No. 6222041), and 5·5 Engineering Research & Innovation Team Project of Beijing Forestry University (Grant No. BLRC2023B08).
-
The authors confirm contribution to the paper as follows: study conception: Zhang L; study design: Zhang L, Cao Y; performing experiments and results analysis: Li X, Xu Y, Liu K, Li Z; draft manuscript preparation: Li X, Xu Y; manuscript revision: Zhang L, Cao Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xinyu Li, Yaxin Xu
- Supplementary Table S1 All primers used in this study.
- Supplementary Table S2 TT2-type MYBs identified in the blueberry genome.
- Supplementary Fig. S1 Analysis of stress-related cis-elements in 18 TT2-type MYBs promoters.
- Copyright: © 2025 by the author(s). Published byMaximum Academic Press, Fayetteville, GA. This articleis an open access article distributed under Creative CommonsAttribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li X, Xu Y, Liu K, Li Z, Cao Y, et al. 2025. VcTT2 enhances drought tolerance in blueberry by regulating ROS scavenging. Fruit Research 5: e029 doi: 10.48130/frures-0025-0016 

VcTT2 enhances drought tolerance in blueberry by regulating ROS scavenging
- Received: 09 March 2025
- Revised: 08 April 2025
- Accepted: 15 April 2025
- Published online: 31 July 2025
Abstract: Blueberries are highly valued for their nutritional richness and distinctive flavor. However, they are susceptible to drought stress, which adversely affects growth, yield, and fruit quality. Therefore, improving drought tolerance in blueberries is crucial for industrial development. In this study, VcTT2 (also named VcMYBPA2.1), a gene encoding a TT2-type MYB transcription factor, was identified, and its expression was found to be induced by drought, salt, and low-temperature stresses. Overexpression of VcTT2 in blueberry calli enhanced drought tolerance by increasing the activities of superoxide dismutase (SOD), peroxidase (POD), and ascorbate peroxidase (APX) enzymes, promoting proline accumulation, and reducing malondialdehyde (MDA) and hydrogen peroxide (H2O2) levels. Furthermore, overexpression of VcTT2 improved drought tolerance in blueberry seedlings, whereas suppressing this gene resulted in decreased drought tolerance. Overexpression of VcTT2 significantly upregulated the expression of genes related to reactive oxygen species (ROS) scavenging under drought stress, including VcSOD1, VcSOD2, VcPOD1, VcPOD2, VcAPX1, and VcAPX2. Collectively, under drought stress, VcTT2 overexpression enhanced the activities of antioxidant enzymes APX, SOD, and POD by promoting the expression of downstream antioxidant enzyme genes, thereby scavenging the over-accumulated ROS and improving the drought tolerance of blueberries. These findings reveal the function and mechanism of VcTT2 in enhancing drought tolerance in blueberry by regulating ROS levels, providing a theoretical basis and genetic resources for drought-tolerant germplasm improvement in blueberry.
-
Key words:
- VcTT2 /
- ROS scavenging /
- Antioxidant enzymes /
- Drought tolerance /
- Blueberry