








-
The stomach, as a critical organ of the digestive system, performs multiple functions, including food storage, digestion, and regulation. The digestive system develops from the mesoderm and endoderm layers, with the endoderm giving rise to the primitive gut. The foregut within the primitive gut further differentiates into the esophagus, stomach, part of the duodenum, liver, gallbladder, pancreas, and respiratory organs[1]. During stomach development, various signaling pathways, including Wnt, bone morphogenetic proteins (BMPs), fibroblast growth factors (FGF), and retinoic acid (RA), play critical roles by coordinating the expression of transcription factors that govern cellular identity[2]. These pathways collectively contribute to the regulation of key developmental processes. However, the precise mechanisms by which these signaling pathways function in gastric development remain inadequately understood. Further investigation is necessary to clarify the molecular details and functional impacts of these regulatory pathways.
The stomach, as a key component of the digestive tract, is particularly susceptible to a range of pathogenic influences. Common gastric disorders include gastric ulcers, gastritis, and gastric cancer. Gastric ulcers and gastritis are frequently associated with Helicobacter pylori (H. pylori) infection, long-term use of certain medications, or psychological stress. In contrast, the development of gastric cancer is often driven by a complex interplay of genetic susceptibility, chronic inflammation, and environmental exposures[3]. These gastric conditions not only disrupt the normal physiological functions of the stomach, but may also impair the overall integrity and health of the gastrointestinal system.
As reported in the Global Cancer Statistics published in 2024, gastric cancer holds a significant position among all cancer types worldwide, with an estimated annual incidence approaching one million cases. It ranks among the top five leading causes of cancer-related mortality globally[4]. Notably, the epidemiological pattern of gastric cancer demonstrates marked regional disparities, with substantially higher prevalence observed in East Asia and Eastern Europe[5]. Although some high-incidence countries have achieved partial success in mitigating disease burden through the implementation of screening programs and lifestyle interventions, gastric cancer continues to impose a considerable challenge to global healthcare systems. The onset of gastric cancer is shaped by a variety of factors, such as H. pylori infection, dietary patterns, smoking, and genetic susceptibility[6]. In 1965, pathologist Laurén[7] proposed a histological classification method for gastric cancer, dividing it into intestinal-type, and diffuse-type. Additionally, there is a less common mixed type[7]. Current therapeutic approaches for gastric cancer are primarily tailored based on tumor stage, histopathological subtype, and the patient's overall clinical condition[8]. Surgery continues to be the primary curative approach for patients in the early and locally advanced stages, whereas radiotherapy and chemotherapy are integral to comprehensive multidisciplinary treatment strategies. Recent advances in systemic therapies for gastric cancer have been marked by the advent of molecular-targeted agents and, notably, by the development of immune checkpoint inhibitors such as programmed cell death protein 1 (PD-1) antagonists, which have yielded encouraging clinical outcomes[9]. However, the early diagnosis of gastric cancer is extremely challenging, and the disease is characterized by its complexity and heterogeneity, with treatment options being somewhat limited. Therefore, the development of effective models to study gastric tissue development, injury repair, and the pathogenesis, progression, and treatment of diseases, especially cancer, is crucial for improving the diagnosis, treatment, screening, and prevention of gastric cancer.
Commonly used models in gastric cancer research include two-dimensional (2D) cell lines, animal models, and emerging organoid models. However, due to prolonged in vitro cultivation, cell lines may undergo genetic and phenotypic alterations, which may not fully capture the heterogeneity of tumors or the complexity of the in vivo microenvironment[10]. Moreover, 2D cell cultures typically grow as a monolayer, which fails to replicate the complex spatial organization and intercellular interactions. To overcome the limitations of in vitro models in recapitulating the tumor microenvironment, researchers commonly use animal models such as xenograft models and genetically engineered mice, which provide greater physiological relevance in mimicking the in vivo development and therapeutic responses of gastric cancer[11,12]. While these animal models offer valuable in vivo experimental data, their establishment is complex, time-consuming, and costly.
Common approaches in three-dimensional (3D) cell culture include spheroid culture, scaffold-based culture, and organoid culture. Spheroids are formed by single-cell suspensions derived from tumor cell lines or primary cells, which spontaneously aggregate in low-adhesion plates or suspension systems[13]. This process typically does not require exogenous extracellular matrix (ECM) such as Matrigel; instead, spheroid integrity is maintained by cell–cell adhesion and mechanical tension. Spheroid culture is simple, cost-effective, and suitable for high-throughput drug screening. However, its structural simplicity and lack of a tissue-specific microenvironment limit its capacity to mimic in vivo functions. To enhance spatial organization, some studies have incorporated scaffolds—such as hydrogels, natural polymers, or 3D-printed structures—into spheroid cultures[14]. Nevertheless, these scaffold-based spheroids still fall short of organoids in terms of self-organization, lineage specification, and organ-specific functionality. In contrast, organoids are typically derived from pluripotent stem cells (e.g., iPSCs, ESCs) or adult stem cells, and cultured in ECM-rich environments like Matrigel. Under the guidance of specific signaling cues, they undergo self-organization to form highly biomimetic 3D structures that recapitulate the cellular composition, spatial architecture, and functional features of native organs. Organoids offer superior biological relevance and predictive value for modeling tissue development, disease progression, and personalized medicine.
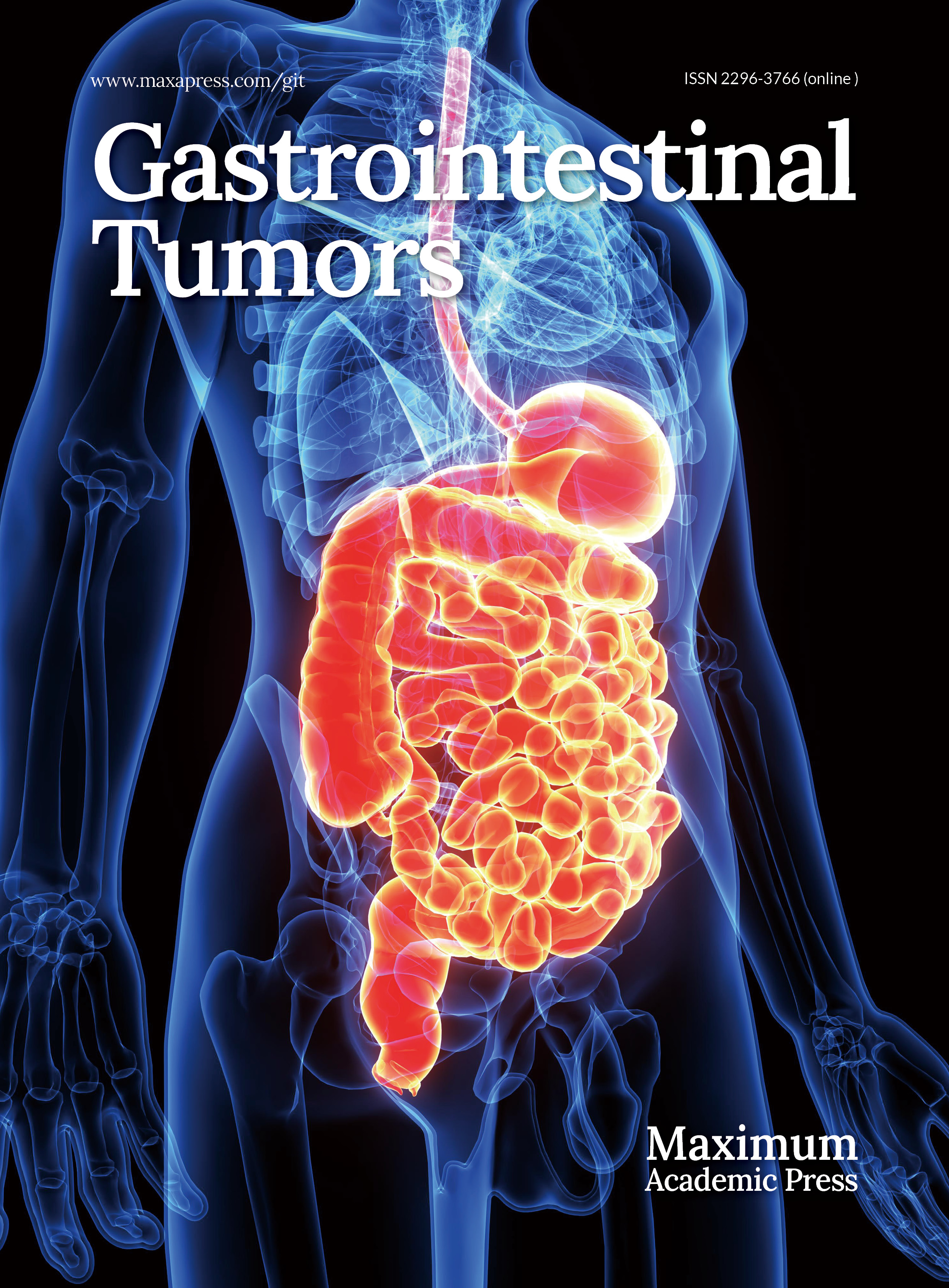
Since the introduction of this technology, organoid models have been rapidly and widely adopted across various research fields. Although several reviews have outlined the general development of organoid technology, a comprehensive account specifically focusing on the evolution of gastric organoid systems remains lacking; this review aims to fill that gap. In terms of applications, while previous literature has primarily emphasized the roles of organoids in disease modeling, drug resistance mechanisms, and personalized medicine, the present article further expands the discussion to include developmental biology and translational medicine, providing a broader perspective. Moreover, we systematically summarize recent advances in co-culture systems involving gastric organoids and immune cells. Finally, we offer an in-depth discussion of the current challenges and limitations faced by gastric organoid research, as well as potential future directions. This review will comprehensively cover the developmental trajectory, establishment methods, applications in developmental biology, modeling of H. pylori infection, gastric cancer and drug resistance studies, exploration of individualized therapeutic strategies, and potential use in regenerative medicine (Fig. 1).
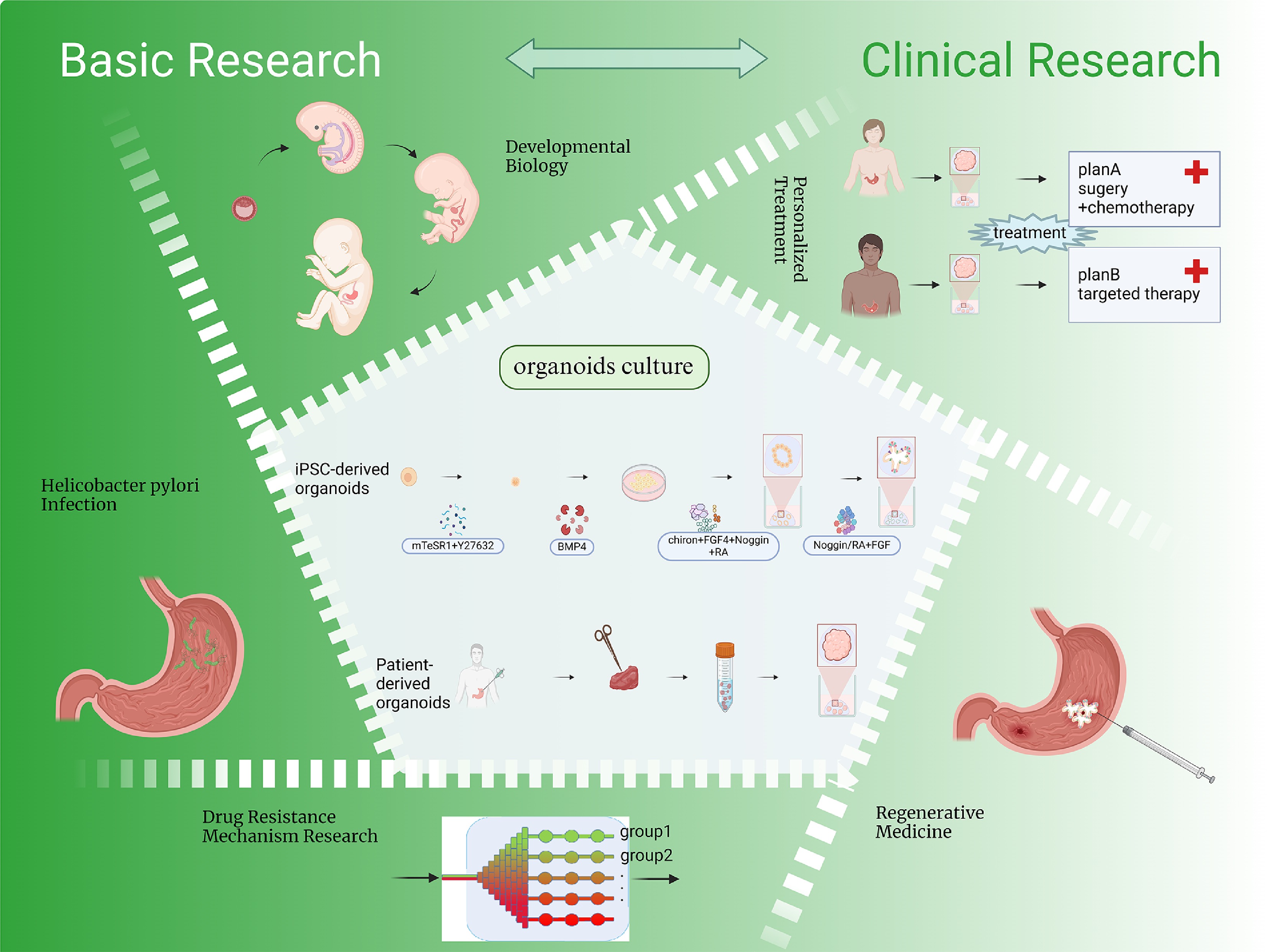
Figure 1.
Organoid generation and applications. The figure depicts the standard methodology for generating gastric organoids and highlights their applications in developmental biology, modeling of H. pylori infection, gastric cancer and drug resistance mechanism research, personalized treatment, and regenerative medicine. Created in BioRender. Chen, E. (2025)
https://BioRender.com/t04o468 . -
Since the first establishment of intestinal organoids by the Clevers group in 2009[15], organoid technology has rapidly advanced over the past decade from basic model construction to the recreation of complex tissue functions (Fig. 2). In 2009, Clevers et al. successfully developed intestinal organoids for the first time. The research team established a long-term culture system in which a single crypt or mouse Lgr5+ intestinal stem cell could develop into an epithelial structure with crypt-villus architecture, containing multiple differentiated epithelial cell types. To achieve long-term culture, the researchers combined multiple signals required for intestinal epithelial growth, including Wnt, epidermal growth factor (EGF) and Noggin, and used a Matrigel matrix to support cell proliferation[15]. The study by Barker et al.[16] in 2010 revealed that, in addition to the isthmus being the primary proliferative region in gastric glands, a population of Lgr5+ cells with significant proliferative capacity is also present at the base of mature pyloric glands. These cells are multipotent stem cells with self-renewal abilities, serving as the foundation for the long-term maintenance and regeneration of the gastric epithelium. The researchers successfully cultured single Lgr5+ cells in a three-dimensional system similar to that used for intestinal organoids, dependent on Wnt3a signaling. This approach resulted in the generation of pyloric organoids that exhibited stable structure, long-term growth, continuous budding, and well-maintained polarity[16]. In 2013, Stange et al. used flow cytometry to isolate single Troy+ cells from gastric corpus units and found that, after cultivation, these cells were also able to generate multi-unit organoids with continuous proliferation[17].
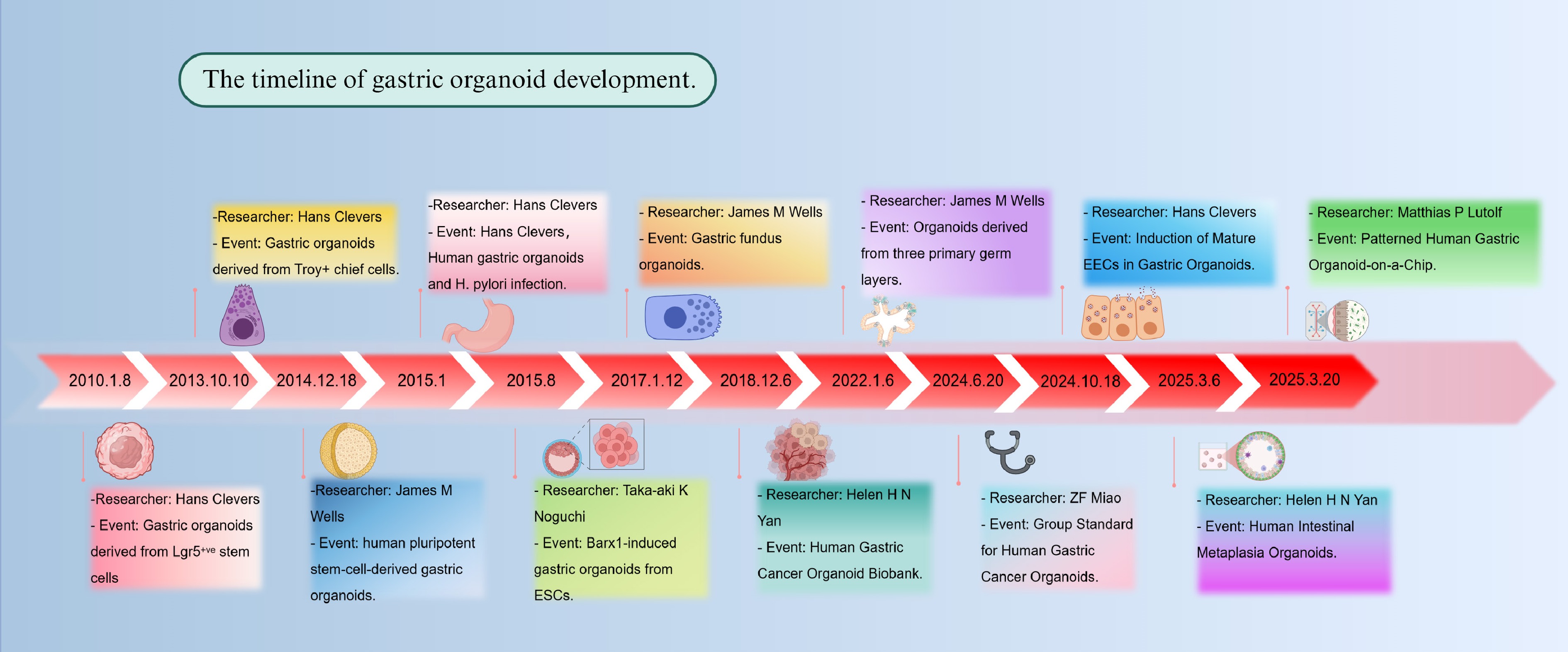
Figure 2.
Timeline of gastric organoid development. The figure illustrates the establishment of the first organoid and major milestones in the development and application of gastric organoid models from 2013 to 2025. Created in BioRender. Chen, E. (2025)
https://BioRender.com/9dh4twy .However, when studying human gastric development and disease, organoids derived from animal models face significant developmental and species differences compared to humans. Consequently, researchers began exploring whether human pluripotent stem cells could be cultured into organoids. To explore the developmental processes of different regions of the human stomach, McCracken et al. created a human antral organoid model in 2014 by guiding the differentiation of pluripotent stem cells. This approach enabled the in vitro reconstruction of 3D structures exhibiting early gastric gland characteristics and pit-like domains[18]. However, as this model primarily recapitulates the antral region, it does not fully capture the complexity of the entire stomach. In 2017, the same group further clarified the crucial involvement of the Wnt/β-catenin signaling pathway in controlling the differentiation of cells located at the base of gastric glands in mice. Building on this understanding, they successfully utilized this pathway to guide the differentiation of human pluripotent stem cells into organoids resembling fundic gland structures[19].
While significant progress has been made in modeling gastric development and physiological mechanisms using normal gastric organoids, there is an equally pressing need for more precise models to study gastric cancer. Gastric cancer organoids, as a novel tumor model, offer a more accurate representation of the disease's heterogeneity and its complex tumor microenvironment. Consequently, research on gastric cancer organoids has advanced substantially in recent years, emerging as a critical tool for investigating tumor initiation, progression, and personalized treatment strategies. In 2018, Yan et al. developed a biobank comprising 46 gastric cancer organoid lines, which collectively represent a wide range of the established molecular subtypes of gastric cancer[20]. Using high-throughput screening, the study assessed the efficacy of conventional chemotherapeutic agents commonly used for gastric cancer treatment, including 5-fluorouracil (5-FU), cisplatin, oxaliplatin, epirubicin, and paclitaxel. This work provides invaluable experimental evidence to support the development of personalized therapeutic approaches for gastric cancer.
In 2022, Eicher et al.[21] established a tri-germ-layer co-culture system based on human pluripotent stem cells (hPSCs), incorporating epithelial progenitor cells of ectodermal origin, mesenchymal cells derived from the mesoderm, and enteric glial cells originating from the endoderm. This approach enabled the generation of the most structurally advanced gastric organoids to date. The resulting organoids exhibited well-defined glandular architecture, smooth muscle tissue, and functionally mature enteric neurons capable of regulating muscle contraction. Further investigation revealed dynamic cross-talk among the different cellular components, highlighting that coordinated signaling across germ layers plays a critical role in orchestrating the spatially organized development of gastric tissue architecture[21].
Organoid technology has been widely applied in cutting-edge fields such as developmental biology, disease modeling, personalized medicine, and tissue regeneration, demonstrating substantial value in both research and clinical contexts. However, the lack of standardized protocols remains a major bottleneck limiting its broader application and clinical translation. To address this issue and improve reproducibility and comparability across studies, China Anti-Cancer Association developed an industry guideline for human gastric cancer organoid research[22]. This guideline provides standardized procedures covering the entire workflow—from tissue collection, transport, and processing to organoid establishment, expansion, cryopreservation, and recovery. Importantly, the guideline outlines key criteria for organoid identification and quality control, focusing on morphology, proliferative capacity, genetic stability, stemness maintenance, and functional phenotype. For instance, organoids should exhibit a cystic or spheroid structure with clear cellular stratification under light microscopy. They should be expandable for at least 5–20 passages while maintaining morphological and proliferative characteristics of the original tumor. Genetic stability is recommended to be assessed by karyotyping, with a normal diploid karyotype of 46, XX or 46, XY. In addition, the expression of proliferation marker Ki-67 and gastric cancer–associated biomarkers such as CA19-9, carcinoembryonic antigen (CEA), CA72-4, CA50, and CA24-2 is used for functional validation.
Enteroendocrine cells (EECs) are widely distributed within the gastrointestinal epithelium, where they secrete various peptide hormones to regulate digestive and metabolic functions. However, due to their low abundance and distinct features, EECs are typically difficult to identify and study. In 2024, the team led by Hans Clevers successfully induced the overexpression of NEUROG3 in gastric organoids using doxycycline, thereby promoting their differentiation into mature EECs[23]. The study demonstrated that these induced EECs not only exhibited mature secretory functions but also closely resembled primary human EECs in both phenotype and function. This technological advancement provides a robust experimental platform for investigating EEC biology, substantially advancing research in this field.
In 2025, gastric organoid research made significant breakthroughs, with notable advances in constructing complex microenvironments and modeling disease progression. During H. pylori-induced malignant transformation of normal gastric epithelium, intestinal metaplasia (IM) represents a critical intermediate stage marked by the emergence of intestinal-like epithelial cells[24]. However, progress in studying this stage has been limited due to a lack of suitable models. Yue et al. established a cohort of IM organoids, providing a valuable platform to elucidate the molecular mechanisms driving IM and its link to gastric carcinogenesis[25]. In the same year, another study focused on microenvironment reconstruction and disease modeling, proposing a novel organoid culture system. Hofer et al. developed a bilateral-access organoid system that allows differential delivery of growth factors to the apical and basal sides, effectively recapitulating the spatial gradient of mesenchyme-derived stem cell signals. Apical exposure to acidic conditions induced differentiation of mature gastric pit cells, which exhibited a previously unreported, cell type–specific response, underscoring the model's strength in capturing epithelial heterogeneity. Moreover, this platform supports H. pylori colonization and sustained infection for up to six days, offering a physiologically relevant tool for studying host–pathogen interactions in the gastric epithelium[26].
The past decade has witnessed substantial progress in the development and refinement of organoid systems, enabling the efficient generation of models that mimic both healthy and pathological tissues from induced pluripotent stem cells or patient-derived samples. These organoids recapitulate developmental cues within 3D culture systems, facilitating the formation of tissue architectures that closely resemble native organs. Compared to traditional cell and animal models, organoids offer several advantages, including improved biological relevance, greater stability, enhanced accessibility, and higher time efficiency. Since its inception, this technology has been widely adopted across a broad range of research disciplines.
-
As a major culture system, organoids encompass those derived from pluripotent stem cells, adult stem cells, and patient-derived tissue. Each type differs in its source, culture method, and ultimate application.
Pluripotent stem cells possess strong differentiation potential and can be used to model the development of various tissues and organs[27]. Compared to the other two types of organoids, iPSC-derived organoids are more challenging to culture, requiring stepwise differentiation protocols to develop into the desired model[28]. The main challenge in culturing this type of organoid lies in designing optimal culture conditions that enable the normal development of pluripotent stem cells. This requires researchers to have a thorough understanding of the gastric stem cell niche[29]. The process of generating gastric organoids in vitro begins with guiding pluripotent stem cells to differentiate into definitive endoderm, which then develops into foregut-like tissues. During this phase, activation of Wnt and FGF signaling pathways supports the formation of gut tube structures while limiting differentiation toward posterior gut lineages. To further promote foregut identity, the BMP pathway is suppressed—often through the addition of Noggin—leading to the development of foregut spheroids. Retinoic acid (RA) is then applied to further specify these spheroids toward a posterior foregut identity, while maintaining high SOX2 expression, a key marker indicative of commitment to the antral gastric lineage[18]. To generate organoids exhibiting fundic characteristics, activation of the Wnt/β-catenin pathway is introduced within a defined temporal window, along with supplementation of EGF and FGF10 to support spheroid maturation. During the later stages of differentiation, modulation of EGF concentration and co-treatment with BMP4 and a MEK inhibitor facilitate the development of functional cell types, including endocrine and parietal cells[30]. Embryonic stem cells (ESCs) can also be used to generate similar models. The process begins with forming embryoid bodies (EBs) on gelatin-coated dishes, which are cultured until day 6 to develop an endodermal cavity. Growth factors such as Sonic Hedgehog (SHH), DKK1, and Noggin are then applied to induce differentiation of the gut-like structures into gastric cell lineages. Finally, the treated organoids are transferred to a 3D matrix containing growth factors to promote maturation, resulting in more complex architectures and diverse cellular compositions[31].
The induction of adult stem cells to form gastric organoids is relatively simple and direct. The culture method employs an intestinal medium containing EGF, Noggin, and R-spondin1, supplemented with Wnt3A and FGF10, to promote budding morphology and expansion of the organoids[16]. To extend the lifespan of gastric organoids, Bartfeld et al. analyzed the effects of various molecules on culture longevity. Nicotinamide, as an optional additive, can enhance early organoid differentiation but gradually shortens their lifespan. Factors known to promote budding and extend lifespan include IGF, GSK3β inhibitors, TGFβ inhibitors, and p38 inhibitors[32]. CHIR99021 (CHIR), a GSK3β inhibitor, in combination with the HDAC inhibitor valproic acid (VPA), has been found to support stem cell maintenance and enhance their proliferative capacity—an effect also observed in gastric tissues. Both CHIR and VPA independently stimulate stem cell proliferation; however, their combined use yields the most robust outcomes. When used together, they not only suppress differentiation-associated gene expression but also upregulate stemness-related genes[33].
In recent years, advancements in tissue acquisition techniques have expanded the sources for establishing patient-derived gastric organoids (PDOs), which are no longer limited to surgically resected specimens. Image-guided biopsies, such as ultrasound or CT, now offer a viable alternative for collecting tissue suitable for organoid culture[34]. Moreover, endoscopic sampling has become a standard clinical practice due to its minimally invasive nature and high safety profile, ensuring patient comfort while enhancing the feasibility of precision medicine[35,36]. Tissues obtained through these methods are typically processed within 24 h for organoid generation, followed by drug sensitivity testing completed within approximately two weeks. This rapid turnaround time provides critical support for the timely development of personalized treatment strategies, underscoring the translational and clinical potential of PDOs in precision oncology[37]. However, two major challenges persist in the establishment of gastric PDOs: microbial contamination and overgrowth of normal cells. Due to necrosis in gastric cancer tissues, bacterial and fungal contamination is common, leading to a lower success rate of PDO establishment, which fails to meet clinical demands. Morimoto et al. suggest that avoiding necrotic regions and obtaining tissue samples from deeper and broader areas can significantly minimize contamination and improve the success rate[38]. Contamination by normal epithelial cells presents an even greater challenge. This issue refers to the phenomenon in which, for reasons yet to be fully elucidated, normal cells proliferate more rapidly than tumor cells during gastric organoid culture. Three strategies have been proposed to mitigate this issue: (1) manual removal of normal cell populations; (2) tumor cell enrichment via flow cytometry; and (3) for tumors harboring TP53 mutations, the use of the small molecule Nutlin-3, which stabilizes TP53 and selectively suppresses the growth of normal cells while permitting the expansion of tumor cells[39]. Tumor-derived organoids exhibit diverse growth dynamics and morphological heterogeneity, including both cystic and solid structures[40].
Recent advancements have significantly broadened the applications of organoid models by enabling their co-culture with microorganisms or immune system components, thereby enhancing their physiological relevance. In infection research, gastric organoids are commonly used as in vitro platforms for modeling host–pathogen interactions, particularly through the microinjection of H. pylori[41]. In cancer studies, co-culturing gastric cancer organoids with cancer-associated fibroblasts (CAFs), and natural killer (NK) cells has provided key insights into the complex dynamics of the tumor–immune microenvironment. CAFs have been shown to suppress NK cell activity, thereby facilitating immune evasion and promoting tumor progression[42,43]. Such multicellular organoid systems represent powerful experimental platforms for investigating tumor immunity and evaluating potential immunotherapeutic strategies[44].
-
As a muscular sac, the complexity of the stomach is often overlooked. The cellular composition and quantity vary across different regions of the stomach, as do their secretory profiles. During stomach development, mesenchymal-epithelial interactions are regulated by numerous signaling pathways and transcription factors[45]. Although there is a certain understanding of gastric development, replicating this process in vitro remains challenging.
Organoids have recently emerged as a robust 3D in vitro model capable of faithfully recapitulating human organogenesis and pathological processes. They have been widely utilized to investigate the fundamental mechanisms underlying gastric development. In 2014, McCracken et al. first demonstrated that human gastric organoids could be derived from human pluripotent stem cells (hPSCs) by precisely modulating key signaling pathways—such as FGF, Wnt, BMP, RA, and EGF—thereby replicating essential stages of early human gastric development in vitro[18]. Subsequent studies on the development of the gastric fundic glands have highlighted the crucial involvement of Wnt/β-catenin signaling in determining the regional patterning of the stomach. A 2017 study showed that activation of this pathway promotes the differentiation of hPSC-derived foregut progenitors into fundic-type epithelium, whereas its inhibition favors the formation of antral-type epithelium. These findings not only confirm the importance of Wnt signaling in establishing gastric regional identity but also underscore the value of organoids as a model system for dissecting the temporal dynamics of developmental signaling. By finely tuning signaling cues at distinct developmental stages, researchers successfully established human fundic gastric organoids that serve as a powerful platform for investigating epithelial morphogenesis and lineage-specific differentiation, thereby offering critical insights into the spatiotemporal regulation of human gastric development[19].
In 2020, Miao et al. employed human- and mouse-derived fundic gastric organoid models to systematically investigate the signaling mechanisms governing parietal cell differentiation. Their study identified the AMP Kinase (AMPK)–KLF4–PGC1α axis as a key regulatory pathway in the development and functional maturation of parietal cells, thereby providing valuable insights into the specification of this epithelial lineage[46]. Furthermore, by modeling gastric mucosal regeneration using organoids, the team discovered aberrant expression patterns of key paligenosis-associated factors, including Ifrd1 and Ddit4. These findings underscore the potential roles of these genes in organoid development and gastric tissue repair following injury[47].
These studies have not only deepened our understanding of human gastric development and regional specification but also highlighted the unique advantages of organoid systems in modeling key developmental events. Serving as a critical bridge between fundamental developmental biology and disease modeling, gastric organoids provide a powerful platform for investigating regenerative mechanisms and the regulation of epithelial cell fate.
-
H. pylori is a spiral-shaped, Gram-negative bacterium and one of the most prevalent pathogens in the human stomach. It is uniquely adapted to survive the highly acidic gastric environment, where it colonizes the mucosal surface, and induces chronic gastritis[48]. Persistent infection can lead to more severe gastric pathologies, including peptic ulcers, atrophic gastritis, and significantly elevates the risk of gastric cancer. Notably, H. pylori infection is recognized as a major risk factor for gastric carcinogenesis, particularly under conditions of prolonged colonization[49]. The pathogenicity of H. pylori involves a multifaceted mechanism. The bacterium produces urease to neutralize gastric acid, creating a localized neutral pH microenvironment that facilitates survival. It employs flagella to penetrate the gastric mucus layer and adheres to epithelial cells through interactions between bacterial adhesins and host receptors. Beyond colonization, H. pylori delivers virulence factors such as cytotoxin-associated gene A (CagA), and vacuolating cytotoxin A (VacA), which trigger a cascade of host cellular responses including inflammation, epithelial injury, and dysregulated signaling—all of which contribute to gastric tumorigenesis[50]. However, the detailed mechanisms underlying the development and progression of gastric cancer remain unclear. Studies have been conducted using cancer cell lines, short-term ex vivo primary cell cultures, and infected mouse models. Nevertheless, model organisms often differ significantly from humans and may fail to fully replicate the complex disease progression and biological processes of human gastric cancer. In addition, in vivo models are costly to maintain and require long experimental durations[51].
Since the development of organoid technology, Bartfeld et al. has introduced H. pylori into organoids, establishing a novel H. pylori model. This research revealed that NF-κB signaling is activated, a finding consistent with previous studies, thereby indicating that organoids are highly suitable for research in this area[32]. Consequently, their related applications are becoming increasingly widespread. Building on this work, McCracken et al. microinjected H. pylori into the luminal surface of gastric organoids to further model the early stages of infection, demonstrating that its pathogenicity is dependent on the virulence factor CagA. CagA is translocated into epithelial cells, where it binds to the c-Met receptor and triggers downstream signaling cascades[18]. In this organoid model, CagA-positive H. pylori strains were found to trigger β-catenin pathway activation, downregulate claudin-7—a key component of the epithelial barrier—enhance aberrant epithelial cell proliferation, and impair mucosal equilibrium, thereby implicating a potential role in the initiation of gastric cancer[52]. The organoid system also revealed that CD44, acting as a co-receptor of c-Met, cooperates with CagA to further enhance epithelial cell proliferation[53]. Furthermore, this model confirmed that low-densitylipoproteinreceptor-relatedprotein8 (LRP8), a member of the low-density lipoprotein (LDL) receptor family, interacts with both CagA and β-catenin to assist in the translocation of β-catenin into the nucleus, thereby enhancing the expression of pro-inflammatory genes and markers associated with cancer stem-like properties—highlighting a potential mechanistic link between H. pylori infection and tumor cell plasticity[54].
Expanding upon traditional 3D organoid approaches, a human gastric organoid-on-a-chip platform with dual-channel access has been engineered. In this system, gastric epithelial cells derived from organoids are cultured on a hydrogel scaffold with predefined geometry, while differential exposure to growth factors across the apical and basal chambers enables the spatial reconstruction of glandular and pit-like domains. Unlike enclosed organoid structures, the open design of this model allows for greater experimental manipulation and supports sustained epithelial homeostasis. Notably, exposure to physiologically relevant acidic conditions drives the generation of pit cells with enhanced maturation and facilitates lineage differentiation patterns that more faithfully mirror the native gastric epithelium[26].
Epithelial-only organoids offer an effective platform for investigating the interplay between H. pylori and the gastric epithelial layer. However, considering that immune responses are also involved in H. pylori infection in the human body, a co-culture system is required when investigating the role of the immune process in this context. Holokai et al. discovered that H. pylori utilizes CagA to upregulate PD-L1 levels in stomach epithelial cells by activating the SHH pathway. Through a co-culture approach involving human gastric organoids (HGOs) and immune cells, it was demonstrated that infection with H. pylori decreases apoptosis, while PD-1 blockade enhances the proliferation of cytotoxic T lymphocytes. This indicates that PD-L1 may have a protective function in regulating immune responses after H. pylori infection, possibly playing a role in the onset of gastric cancer[55]. In addition, gastric organoids can be co-cultured with macrophages. Through this model, the authors found that H. pylori infection leads to increased cytokine production, which is especially pronounced in the absence of Nod1. This indicates that Nod1 alters macrophage polarization to reduce the inflammatory response, facilitating immune evasion and protecting H. pylori[56].
In summary, this co-culture model is highly suitable for studying Helicobacter pylori infection and its pathogenic mechanisms. It has a short cultivation cycle and saves both time and resources. The model not only allows for the investigation of the direct interaction between H. pylori and epithelial cells but also enables the study of the role immune components play through co-culture systems.
-
Chemotherapy resistance in gastric cancer remains a major challenge in clinical treatment, significantly affecting patient prognosis and survival rates[57]. As treatment progresses, tumor cells often develop resistance to chemotherapy drugs through various mechanisms, thereby reducing the effectiveness of the therapy[58]. In addition, the high heterogeneity of gastric cancer complicates the mechanisms of drug resistance, with significant molecular differences observed between patients[59]. The advent of organoid technology has recently introduced an innovative platform for investigating mechanisms of drug resistance in gastric cancer. By recapitulating the biological characteristics of patient tumors, organoids can help identify key drivers of resistance and validate potential therapeutic targets[60]. This technological advancement provides a promising avenue for elucidating drug resistance mechanisms in gastric cancer and facilitating the development of innovative therapeutic strategies to overcome resistance.
As a first-line drug for gastric cancer, 5-FU plays a significant role in treatment; however, many patients still experience relapse after therapy[58]. Using patient-derived gastric cancer organoids, Ukai et al. established an in vitro model that simulates the development of resistance to 5-FU. This model revealed that KHDRBS3 plays a critical role in 5-FU resistance by regulating the expression of CD44 variants, thereby promoting stem-like properties in gastric cancer cells[61]. In addition, in PDOs from the 5-FU + CDDP-resistant subtype of gastric cancer (GC), ADAR1 regulates lipid metabolism-related genes through RNA editing, promoting self-renewal and chemoresistance. ADAR1 also modulates the mRNA stability of stearoyl-CoA desaturase (SCD1), thereby alleviating chemotherapy-induced endoplasmic reticulum stress. This regulation leads to the upregulation of β-catenin expression, ultimately contributing to chemoresistance and the enrichment of cancer stem cell populations. Inhibition of SCD1 can reverse chemoresistance and reduce the population of cancer stem cells[62]. In 5-FU-resistant cells, elevated expression levels of ferroptosis-suppressing genes (GPX4, SLC7A11, FTH1) along with STAT3 have been observed. STAT3 contributes to resistance by orchestrating a regulatory network that inhibits ferroptosis through its transcriptional regulation of these genes. Inhibition of STAT3 activity induces ferroptosis by increasing lipid peroxidation and iron accumulation. A STAT3 inhibitor, W1131, has been developed and exhibits significant antitumor effects in multiple models. This study proposes a novel therapeutic strategy for overcoming chemoresistance in gastric cancer[63]. Moreover, organoids not only serve as valuable models for investigating mechanisms of drug resistance, but also function as platforms for evaluating the antitumor efficacy of therapeutic agents, demonstrating considerable potential for clinical translation[64].
In addition to exploring mechanisms of resistance and identifying new potential drug targets, organoid technology also provides clinical guidance. PDOs used to screen drug sensitivity in diffuse gastric cancer (DGC) revealed that Aurora kinase inhibitors (AURKi) can induce a senescent phenotype in DGC cells. These senescent cancer cells secrete large amounts of MCP-1/CCL2 chemokines, which induce macrophage recruitment and polarization towards the M2 phenotype, thereby compromising the immune-mediated killing of cancer cells by macrophages. This drug-induced senescence has not yet been reported in gastric cancer studies, underscoring the utility of organoids in modeling unique drug responses of cancer cells. The study suggests that strategies for clearing senescent cells following AURKi treatment should be explored to prevent tumor recurrence or metastasis driven by these senescent cells[65].
Despite significant breakthroughs in understanding the molecular mechanisms of drug resistance, many unknown areas remain to be explored[66]. Organoid technology, as an innovative in vitro model, serves as a powerful tool for the development of personalized therapies and anti-resistance strategies. In the future, through in-depth research on the applications of organoids, combined with big data and multi-omics analyses, researchers are expected to further uncover the core mechanisms of gastric cancer resistance and develop more precise and effective treatment approaches, ultimately improving patient prognosis and survival rates[67].
-
Due to its high complexity at both the molecular and histological levels, GC exhibits marked heterogeneity. Based on comprehensive genomic analyses, The Cancer Genome Atlas (TCGA) classifies GC into four distinct molecular subtypes—Epstein-Barr virus (EBV)-positive, genomically stable (GS), microsatellite instability (MSI), and chromosomal instability (CIN)—to facilitate the development of precision therapeutic strategies[68]. Tumor heterogeneity is evident not only between individuals but also within the same individual, including differences between primary sites and metastatic lesions, and even within the same tumor. Even among patients classified under the same subtype, treatment responses can vary significantly, complicating the selection of subsequent chemotherapy regimens. This variability may also result in chemotherapy failing to achieve optimal therapeutic outcomes for some patients[69]. Precision treatment for gastric cancer aims to accurately stratify different subgroups to develop effective therapeutic strategies. In addition to next-generation sequencing (NGS), which has made precision treatment feasible, PDOs offer the ability to establish a 3D tissue structure in vitro that closely resembles the patient's tumor. PDOs retain the tumor's gene expression profiles and molecular characteristics, faithfully recapitulating tumor heterogeneity while maintaining stability during long-term culture[68,70,71].
Vlachogiannis et al. established an organoid biobank derived from patients with metastatic gastrointestinal cancers. Through histological and biochemical marker analysis, they found a high degree of concordance between the morphology and marker expression of the gastric cancer organoids and the original tissue biopsies. This indicates that the organoids serve as a reliable 'replica' of the tumor in gastric tissue. Furthermore, the researchers evaluated the drug responses of organoids and compared them to those observed in xenograft models derived from PDOs. They also utilized the PDO model to predict patient responses to various therapeutic approaches. This study demonstrates the promising potential of PDO models for advancing clinical precision medicine[72]. Zhao et al. also established gastric cancer organoids from different anatomical sites and TNM stages of the stomach, studying their responses to six chemotherapy drugs. Through transcriptome sequencing analysis, it was found that organoids sensitive to 5-Fu exhibited downregulation of genes related to proliferation and invasion, while genes associated with cellular senescence and the p53 signaling pathway were upregulated. Similarly, the authors tested these findings in PDO-derived xenograft models in mice and used PDOs to predict patient drug responses. The findings consistently support the idea that organoids are a valuable resource for drug screening in cancer patients[40]. Wang et al. investigated the consistency between drug sensitivity and patient prognosis using PDOs, further demonstrating that PDOs are well-suited for personalized treatment in clinical practice[73]. Yan et al. also established organoids with CLDN18-ARHGAP6 and CTNND1-ARHGAP26 fusions, which are absent in 2D cell lines. In the course of large-scale drug screening using the established PDO biobank, the screening process for each organoid was completed in under two weeks, greatly expediting the development of personalized treatment strategies[20]. However, when using organoids in clinical laboratory-developed tests (LDTs), strict standards and regulatory oversight are required to guarantee the accuracy and reliability of experimental results[74].
The tumor development process is heavily influenced by the immune microenvironment, yet traditional organoid models, which consist exclusively of epithelial cells, do not incorporate immune components. Chakrabarti et al. investigated the influence of myeloid-derived suppressor cells (MDSCs) on tumor organoids by co-culturing them with gastric organoids derived from patient samples. By incorporating immune components, this approach facilitates a more accurate assessment of tumor response to pharmacological interventions and contributes to the discovery of new therapeutic targets in gastric cancer[75]. Nonetheless, the approach may impose restrictions on the natural crosstalk between immune cells and organoid structures. To address this, the authors coated the culture surface with Matrigel, allowing the cells to adhere to the surface rather than embedding them within Matrigel for complete encapsulation[76]. In addition, patient-derived tumor cells and stromal cells can be cultured within a GelMA-Gelatin-Matrigel composite hydrogel, which facilitates their uniform distribution. These cells can then be divided into numerous identical assemblies, enabling standardized and scalable high-throughput drug screening. The models generated through this method closely mimic the tumor responses observed in patients[77].
In addition to published studies, several clinical trials are currently investigating the translational potential of gastric organoids. These efforts primarily focus on using PDOs to predict therapeutic response, guide drug selection, and evaluate the efficacy of neoadjuvant treatments (e.g., NCT05842187, NCT06196554, NCT06100003)[78]. Such exploratory applications underscore the promise of PDOs in bridging basic research and personalized clinical decision-making in gastric cancer.
In summary, patient-derived gastric cancer organoids not only retain the molecular characteristics and heterogeneity of the original tumors but also demonstrate significant potential in predicting drug sensitivity and guiding personalized treatment strategies. With the continued advancement of co-culture systems and high-throughput platforms, PDOs models are gradually transitioning from basic research to clinical application, offering a practical and robust tool to support precision oncology in gastric cancer.
-
Gastric subtotal and total gastrectomy are common surgical treatments for gastric cancer. Following these procedures, the jejunum is often used to replace the removed stomach, leading to a decrease in reservoir function and the loss of the anti-reflux barrier. As a result, patients may experience symptoms such as dumping syndrome and alkaline reflux gastritis[79]. However, due to a scarcity of donors and a significant risk of rejection, gastric transplantation is not a viable solution. Consequently, patients are left to endure a decline in quality of life after surgery[80]. The emergence of organoid technology offers hope for overcoming the challenges associated with transplantation. On one hand, patients would no longer need to endure long waiting times for donor allocation, and on the other hand, organoid technology could significantly reduce the issue of immune rejection.
In 2020, Yoshihara et al. successfully developed insulin-secreting pancreatic islet organoids with only endocrine function. When transplanted into diabetic mouse models, these organoids not only restored glucose homeostasis but also did not trigger immune rejection[81]. The Cambridge team, led by Ludovic, was the first to successfully apply bile duct organoid transplantation in regenerative medicine. Their work not only repaired chemically damaged bile ducts in mice but also restored damaged ducts in human donor livers. This pioneering achievement provides a prototype for the application of organoids in humans, advancing the potential of cell therapy in clinical settings[82].
Gastric organoids possess self-renewal and self-organization capabilities, which are crucial for gastric mucosal injury and ulcer repair. Researchers transplanted gastric organoids into aged mice and observed gradual repair of gastric ulcers[83]. The application of organoid-based regenerative medicine in the stomach can also be extended to organ transplantation. Before miniature gastric organoids can be used in organ transplantation, it is necessary not only to cultivate a complete structure that includes components like muscle, blood vessels, and nerves, but also to grow it to an appropriate size and anatomical configuration. After transplanting complex gastric organoids—containing glands, smooth muscle, and functional enteric neurons—into mice using microsurgical techniques, these organoids expanded a thousandfold, forming visible miniature organs. Eicher et al. suggests that using animal hosts to further develop human-derived organoids may ultimately serve as the pathway for organoid tissue transplantation in human patients. However, clinical use of such organoid transplants must adhere to Good Manufacturing Practice (GMP) standards, which may limit the feasibility of using animals as long-term hosts for organoid production[21].
Currently, the application of organoid technology in gastric regenerative medicine is still in its infancy, with a long way to go before achieving clinical implementation.
-
Although organoid technology has brought unprecedented breakthroughs in scientific research and clinical applications, it is not without limitations. Organoid models still exhibit significant shortcomings in certain areas, such as a simplified culture environment, lack of complete tissue structures, insufficient microenvironment simulation, and the absence of vascular integration. Additionally, challenges related to long-term cultivation and standardization of organoids remain critical bottlenecks affecting experimental reproducibility and clinical translation.
However, numerous exploratory solutions are already being pursued to address these limitations. First, the organoid culture environment is relatively simplistic, lacking the complex physiological signals present in the stomach, such as dynamic peristalsis, neural regulation, and the secretion of various substances. These factors play crucial roles in gastric development, function, and disease response in vivo, but are often inadequately replicated in organoid systems in vitro. Most importantly, organoids typically lack vasculature and form spherical or hollow structures. Organoids located at the center of the Matrigel have limited exposure to the external environment and culture medium, restricting the supply of nutrients and oxygen[84]. This not only affects the long-term culture and growth of organoids but also potentially limits their application in modeling the cancer microenvironment and studying the physiological and pathological mechanisms of gastric epithelial cells. The emergence of microfluidic technology offers a potential solution to this issue by integrating multiple external stimuli and controlling fluid flow to better simulate the physiological conditions within the human body[85]. Lee et al. developed a gastric organoid-on-a-chip model that, on a microfluidic imaging platform, uses a peristaltic pump to deliver culture medium through the lumen, achieving lumenization of the stomach and closely mimicking its physiological conditions[86]. By enabling the co-culture of pancreatic islet and liver organoids on the same chip, it is also possible to simulate the interactions between different organs, providing a more comprehensive model of inter-organ communication[87].
Additionally, the cellular composition of organoids is relatively simple, consisting only of a single layer of epithelial cells, lacking immune cells, stromal cells, and other components present in the microenvironment. This limitation hinders the full replication of the complex cell-cell interactions occurring within the stomach[88]. Moreover, the progression of gastric diseases is often closely associated with the immune system[89]. For example, macrophages not only participate in anti-tumor immunity but can also promote tumor growth and metastasis[90]. The development of organoid co-culture technology effectively addresses this issue (Table 1). The co-culture of organoids with diverse immune cell populations offers a more physiologically relevant model that closely recapitulates the in vivo microenvironment, thereby facilitating comprehensive studies of immune cell functions and their interactions within the tumor microenvironment[75]. In co-culture systems of organoids and immune cells, commonly used approaches include Transwell assays and Matrigel embedding. Transwell systems separate cells via a permeable membrane, making them suitable for studying paracrine signaling, while Matrigel co-embedding allows direct cell–cell contact, better mimicking in vivo interactions. Each model has its advantages, and the choice should be guided by specific experimental objectives.
Table 1. Co-culture models of gastric organoids with various cell types in recent literature.
Organoids Co-cultured cells Co-culture ratio Culture method Publication year Ref. PDOs CTLs Not mentioned Mix in Matrigel 2021 [91] HuFGOs DCs, CTL Not mentioned Mix in Matrigel 2019 [55] PDOs PBMCs 1:5 Not mentioned 2021 [92 Mouse gastroid Macrophages Not mentioned Transwell 2019 [56] PDOs Macrophages 300:105 Co-cultured in the medium 2024 [65] HuFGOs DCs 80:105 Organoids in Matrigel, overlaid with DCs 2019 [93] HuFGOs DCs Not mentioned GOFlowChip 2021 [94] Mouse and human gastroid DCs Not mentioned Transwell 2022 [95] PDOs CAFs 2:1 Mix in Matrigel 2024 [40] TCs 1:5 Mix in Matrigel Mouse and human gastroid ILC2s Not mentioned Mix in Matrigel 2024 [96] The table outlines the types of co-cultured cells, their respective ratios, and the co-culture methods used, offering a concise overview of technical approaches in the field. Although organoid–immune cell co-culture systems offer valuable tools to model tumor–immune interactions, several limitations remain. First, these models cannot fully recapitulate the complex in vivo tumor microenvironment, which involves dynamic interactions among immune cells, endothelial cells, fibroblasts, and others. While tumor-resident immune cells are more physiologically relevant, they are difficult to isolate, available in limited quantities, and challenging to expand in vitro. In contrast, peripheral blood–derived immune cells are more accessible but may not faithfully reflect local immune responses. Moreover, cell ratios and culture conditions in co-culture systems often fail to mimic in vivo settings. Second, to obtain sufficient numbers of tumor-specific T cells, repeated in vitro expansion is often required, which may lead to T cell exhaustion characterized by functional decline and upregulation of inhibitory receptors.
Moreover, the lack of standardization and reproducibility in organoid models leads to poor consistency and repeatability in experimental results. In organoid culture, Matrigel is a commonly used scaffold material. It is extracted from Engelbreth-Holm-Swarm (EHS) tumors and contains soluble basement membrane proteins. Matrigel can form a 3D gel at 37 °C, simulating the in vivo 3D environment of tissues[97]. Matrigel provides an ideal physical and chemical environment for the formation and maintenance of various organoid systems, including those derived from the pancreas, colon, and liver. However, different batches of Matrigel exhibit variations in biochemical properties, growth factor concentrations, and mechanical performance, which can potentially affect experimental outcomes.
In recent years, researchers have explored more controllable alternatives to Matrigel, among which synthetic or naturally derived hydrogels represent one of the most promising directions. Hydrogels are composed of hydrophilic polymer networks that can be crosslinked chemically or physically to form a 3D structure resembling the ECM. Compared to Matrigel, their major advantages lie in defined composition and tunable mechanical properties. Additionally, decellularized ECM-derived hydrogels can provide organ-specific biochemical cues that support organoid growth[98]. Despite improved controllability and reproducibility, hydrogels still face several limitations. Most synthetic hydrogels lack the complex bioactive signals present in native ECM, making it difficult to fully support organoid self-organization and long-term maintenance without further modification, such as the incorporation of adhesion peptides or natural proteins. Moreover, hydrogel formulation and crosslinking are often sensitive to environmental conditions, resulting in reduced experimental reproducibility and limited accessibility for general biological laboratories. Although no scaffold material currently matches the full bioactivity and supportive capacity of Matrigel, future improvements—such as the development of chemically defined hydrogels, organ-specific ECM-based materials, modular platforms, and standardized quality control systems—may help advance the standardization and scalability of organoid culture systems, facilitating broader application and clinical translation[99].
Beyond the technical limitations and advancements discussed above, several new technologies have been integrated with organoid systems, offering promising opportunities to study physiological processes and disease progression at the cellular and genetic levels. One such technology is CRISPR/Cas9, a gene-editing tool that can accurately target specific gene sequences and induce double-strand breaks in DNA. Through the cell's damage repair mechanisms, this enables precise gene knock-in or knock-out, expanding the potential of organoid-based research[100]. Leveraging CRISPR/Cas9-mediated knockout of ARID1A—an epigenetic regulator commonly altered across multiple cancer types—in organoid models enables researchers to explore its contribution to tumorigenesis and gain deeper insights into its role in disease progression[101]. Guenther et al. utilized CRISPR/Cas9 technology to knockout the cortactin gene in dysplastic organoids, establishing a model that simulates the malignant transformation process of gastric epithelium. By integrating an air-liquid interface (ALI) culture system, they successfully reconstructed the structural features of gastric epithelium in vitro. This approach revealed how the loss of cortactin disrupts cellular polarity, affecting the construction of epithelial layers and promoting tumor-like morphology, thereby providing a controllable organoid platform for understanding epithelial malignant progression[102]. Furthermore, integrating single-cell sequencing with organoid technology allows for a deeper understanding of the gene expression profiles of different cell types within organoids. This approach can reveal the roles of these cells in developmental processes and disease progression[103]. Looking ahead, as technology continues to advance, organoid models will increasingly integrate with other technologies, thereby expanding the scope of organoid applications across various fields.
To promote the application of organoid technology in drug screening, future developments must move toward automation and high-throughput capabilities. By incorporating technologies such as automated liquid handling systems, microfluidic chips, and machine learning, organoids can be cultivated and analyzed on a large scale with greater efficiency. This is especially critical in personalized medicine, where drug sensitivity tests on patient-derived organoids can aid in formulating individualized treatment plans. Moreover, to ensure the widespread use of gastric organoid technology in research and clinical applications, establishing standardized operating procedures and quality control systems is essential. In the future, researchers will need to develop unified standards for organoid cultivation, analysis, and identification, and extend these standards to preclinical and clinical studies. As organoid technology continues to mature, a key focus will be on how to effectively translate this technology into broader clinical applications.
-
The advent of gastric organoid technology has enabled in-depth investigation into the mechanisms underlying gastric development, physiological functions, and disease pathogenesis, while also showing great potential in personalized medicine and drug screening. Despite significant progress, challenges remain, such as insufficient tissue complexity, lack of vascularization, and limited integration of the immune system. Future advancements in bioengineering, genome editing technologies, and automation are anticipated to further enhance the applicability of gastric organoids in simulating disease processes, evaluating therapeutic compounds, and advancing precision medicine. By further optimizing culture conditions and standardizing operational procedures, the clinical translation potential of gastric organoids will be fully realized, contributing to the development of effective therapeutic modalities for gastric cancer and various gastrointestinal pathologies.
-
This review did not involve any studies with human participants or animals, and therefore, ethical approval was not required.
The authors thank the following support: Miao ZF was supported by National Natural Science Foundation of China (No. 82072724), and the Department of Education of Liaoning Province (QNZR2020004, 2021-YQ-07).
-
The authors confirm their contributions to the paper as follows: study conception and design: Zhao TT; literature search: Mu XR, Zhang JN, Li ZL, Yuan M, Chen WT; writing − draft manuscript preparation: Mu XR; figure preparation: Wang LL; writing − manuscript revision: Miao ZF, Zhao TT, Wang LL, Pang MJ, Tong QY. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xin-Ru Mu, Li-Li Wang
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This articleis an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Mu XR, Wang LL, Pang MJ, Tong QY, Zhang JN, et al. 2025. Exploring the applications of gastric organoids in disease modeling and personalized medicine. Gastrointestinal Tumors 12: e013 doi: 10.48130/git-0025-0013 

Exploring the applications of gastric organoids in disease modeling and personalized medicine
- Received: 15 February 2025
- Revised: 09 July 2025
- Accepted: 25 July 2025
- Published online: 31 July 2025
Abstract: Organoid systems are capable of forming three-dimensional self-organizing structures that closely recapitulate the cellular composition and physiological functions of native gastric tissue. Since their introduction, gastric organoid technology has rapidly advanced and become a powerful tool for studying gastric development, disease mechanisms, and therapeutic responses. This review summarizes key developments in the field, with a focus on five major areas of application: gastric developmental biology, modeling of Helicobacter pylori infection, research on gastric cancer and drug resistance, assessment of personalized therapy, and regenerative medicine. Distinct from previous reviews, this article systematically traces the evolution of gastric organoid technology and provides an in-depth analysis of current limitations in microenvironment reconstruction, standardized culture, immune co-culture modeling, and clinical translation. In addition, it discusses emerging strategies and integrated technologies aimed at overcoming these challenges, offering perspectives to advance organoid systems from basic research toward clinical application.
-
Key words:
- Stomach /
- Organoids /
- Tumor microenvironment /
- Preclinical trial /
- Personalized treatment