









-
Ganoderma lucidum, a representative medicinal mushroom in traditional Chinese medicine, belongs to the Basidiomycota, Agaricomycetes, Polyporales, and Ganodermataceae[1,2]. Known as the 'fairy mushroom' in ancient China due to its significant pharmacological activities, modern pharmacology studies have confirmed that G. lucidum possesses extensive medicinal values, including anti-tumor, immunomodulatory, anti-inflammatory, anti-aging, and cardio-cerebrovascular protective effects[1,3,4]. Its bioactive components mainly include polysaccharides and ganoderic triterpenoids[4,5], among which ganoderic triterpenoids are the core bioactive components, and their content has become one of the key indicators for evaluating the quality of G. lucidum germplasm. Currently, improving the yield of ganoderic triterpenoids in G. lucidum to meet the market demand for high-quality germplasm resources has become one of the core directions in G. lucidum research.
To promote the sustainable development of the G. lucidum industry, it is of great significance to breed superior strains with both high quality and high pharmacological activity to match market demand. The main breeding methods for G. lucidum currently include artificial selection[6], protoplast fusion[7,8], mutagenesis breeding[9], and molecular breeding[10]. Among them, physical or chemical mutagens can induce massive genetic variations in G. lucidum in a short time, thereby obtaining abundant genetically modified strains, and superior strains with target traits can be obtained after screening. Compared with artificial selection and physical mutagenesis, chemical mutagens directly act on genomic nucleotides, mainly inducing base point mutations. This process can simulate the variation characteristics of natural species evolution, usually without causing complete loss of gene function, but only changing the intensity of function, which facilitates the fine analysis of gene function[11−13].
Common chemical mutagens include lithium chloride[14,15], ethyl methanesulfonate (EMS)[16], sodium nitrite[14,17], and sodium azide (SA)[18]. In reported studies on chemical mutagenesis libraries of G. lucidum, Dong et al. confirmed that after mutagenizing G. lucidum protoplasts with 0.3% lithium chloride, the mortality rate reached 72.8%, and the germanium content of the 4th generation of mutagenized strains was the highest (174.88 μg/g), which was 269.02% higher than that of the parent strain[19]. Peng et al. obtained two mutant strains by treating G. lucidum protoplasts with lithium chloride alone, or in combination with Triton X-100, and the maximum yields of intracellular polysaccharides and ganoderic triterpenoids in these mutant strains increased by 568.45%, and 373.43%, respectively[20]. Although the above studies have successfully obtained G. lucidum mutants with superior traits through chemical mutagenesis, there is still a lack of methods for constructing G. lucidum mutant libraries using SA and efficient screening systems for mutant strains.
After constructing the G. lucidum mutagenesis library, screening strains with improved target traits is a core link, and the key is to efficiently and rapidly screen target strains from a large number of mutants. Common screening methods are mostly based on selectable markers, including drug resistance marker screening[21], molecular marker screening[22], fluorescent marker screening[23], and antagonistic screening[24].
Primary screening usually focuses on strain phenotypes, such as differences in mycelial growth rate, color, and other phenotypic characteristics, leading to relatively low screening efficiency. Phenotypic variation, in turn, originates from alterations in genetic material, which can significantly increase population genetic diversity. Genetic differences between different strains can inhibit the growth and proliferation of heterologous hyphae, a phenomenon manifested as the formation of antagonistic lines between strains in phenotypes, mostly occurring in the middle stage of strain growth and development; this trait can be used to clarify the genetic relationship between test strains and identify the mutation status of strains[24−26]. Based on this principle, co-cultivating mutagenized strains with the original strain, observing the occurrence and intensity of antagonistic lines can reflect the degree of genetic variation of G. lucidum mutant strains after mutagen treatment, and further reflect the differences in physiology and metabolism between mutants and the original strain.
Sodium azide (SA), as a highly efficient azide mutagen, can induce A/T→G/C base substitution, thereby causing point mutations[27]. SA is widely used in crop breeding and has successfully induced mutant populations in rice[28], wheat[29], barley[30], oats[31], and other species, but its application in G. lucidum breeding has not been reported. In this study, SA was used to mutagenize G. lucidum protoplasts, which, lacking a cell wall, serve as ideal recipients for chemical mutagenesis in fungal breeding. Genetically modified strains were screened by observing the antagonistic phenomenon between mutants and the original strain after co-cultivation, and mutagenized strains were subjected to cultivation experiments and total triterpenoid content determination to obtain high-triterpenoid-yielding strains.
-
The G. lucidum strain 'Xianzhi No.2' and 'Xianzhi No.3' used in this study were purchased from Zhejiang Shouxiangu Plant Medicine Research Institute Co., Ltd., and preserved in potato dextrose agar (PDA) medium in the laboratory. The liquid fermentation culture method of G. lucidum mycelia referred to previous research reports[32]; the cultivation experiment of G. lucidum mutant strains was carried out in Hangzhou Shouxiangu Planting Base in May 2025.
Protoplast preparation
-
G. lucidum mycelia cultured for 5−6 d (100 mL mycelial suspension in 250 mL conical flasks) were centrifuged at 5,000 r/min, 4 °C for 15 min, and the supernatant was discarded to collect the mycelia. The mycelial precipitate was resuspended in 0.6 mol/L mannitol, centrifuged at 5,000 r/min, 4 °C for 15 min, and the supernatant was discarded; this washing step was repeated twice. A 2% lywallzyme solution with a volume of 2−3 times that of the wet mycelia was added to the precipitate, mixed by pipetting, and enzymolyzed at 30 °C, 100 r/min for 2−3 h. The enzymolyzed solution was filtered through a sterile 0.45 μm filter cloth twice, and the filtrate was collected and centrifuged at 3,000 r/min, 4 °C for 5 min to discard the supernatant. The protoplast precipitate was resuspended in pre-cooled 0.6 mol/L mannitol, centrifuged at 3,000 r/min, 4 °C for 5 min, and the supernatant was discarded; this washing step was repeated twice. A 1−5 mL pre-cooled 0.6 mol/L mannitol solution was added to prepare protoplast suspension, which was counted under a microscope using a hemocytometer, and the protoplast concentration was adjusted to 1 × 107 cells/mL.
Sodium azide (SA) mutagenesis procedure
-
SA concentration gradients of 0, 1.0, 2.0, 3.0, 4.0, and 5.0 mM (six gradients in total) were set, and each concentration of SA solution was prepared with 0.1 mol/L phosphate buffer (pH = 3.0). 1 mL of each concentration of SA solution was used to treat 1 × 105 G. lucidum protoplasts, with three biological replicates per concentration, and the mutagenesis treatment lasted for 3 h. After mutagenesis, the mixture was centrifuged at 3,000 r/min, 4 °C for 5 min to discard the supernatant, and the precipitate was resuspended and washed twice with pre-cooled 0.6 mol/L mannitol. 500 μL pre-cooled 0.6 mol/L mannitol was added to suspend the protoplasts, which were evenly spread on protoplast regeneration medium and cultured in a constant temperature incubator at 28 °C for 6 d. The growth status of colonies was observed, and the number of regenerated colonies was counted. The half-lethal concentration was determined by colony counting, and the mutagenized strains at this concentration were picked, expanded, and preserved in PDA solid medium.
Antagonistic screening and cultivation of mutagenized strains
-
G. lucidum protoplasts were treated at the half-lethal concentration of SA, and the obtained mutagenized strains were uniformly expanded and cultured. Mycelial blocks of uniform size were prepared with a puncher and inoculated: the wild-type 'Xianzhi No.2' strain was inoculated in the center, the wild-type 'Xianzhi No.2' (negative control), and Xianzhi No. 3 (positive control) were inoculated above, and the mutagenized strains were inoculated below. After culturing at 28 °C for 6 d, the antagonistic phenotypes were observed and recorded. Mutagenized strains with antagonistic phenotypes and some strains without obvious antagonistic phenotypes were selected and inoculated into fungus bags for fruiting body cultivation.
Determination of total triterpenoid content
-
(1) Preparation of oleanolic acid standard curve: a 1 mg/mL oleanolic acid standard solution was prepared with methanol as the solvent. The standard solution was precisely measured at 0.02, 0.04, 0.06, 0.08, 0.1, 0.2, 0.3, 0.4, and 0.5 mL, placed in 10 mL stoppered test tubes, and the solvent was allowed to evaporate and cool to room temperature. Next, 0.2 mL of freshly prepared vanillin-glacial acetic acid solution was precisely added (weigh 0.5 g vanillin, add glacial acetic acid to make up to 10 mL), with 0.8 mL of perchloric acid, shaken well, heated in a water bath at 70 °C for 15 min, and immediately cooled in an ice bath for 5 min. After removal, 4 mL of ethyl acetate was precisely added, shaken well, and, using the corresponding reagent as blank control, the absorbance at 546 nm by ultraviolet-visible spectrophotometry was determined[33]. A standard curve with absorbance as the ordinate and oleanolic acid concentration as the abscissa was then drawn.
(2) Preparation and determination of test sample solution: the samples were dried to a constant weight at 60 °C, ground, and sieved through a 40-mesh sieve. Then, 60 mg of ground G. lucidum fruiting body powder was accurately weighed and mixed with 1.5 mL of ethanol. Ultrasonic extraction was performed at room temperature with a power of 200 W for 45 min. The mixture was subjected to ultrasonic treatment for 45 min, and this treatment was repeated twice. Subsequently, the mixture was centrifuged at 10,000 rpm at 4 °C for 5 min. An aliquot of 0.8 mL of the supernatant was transferred into a 2 mL centrifuge tube, and another 0.8 mL of ethanol was added and mixed well. Then 0.2 mL of the resulting solution was pipetted out and evaporated to dryness. To the dried residue, 0.2 mL of freshly prepared vanillin-glacial acetic acid solution and 0.8 mL of perchloric acid were added and mixed thoroughly. The mixture was heated in a water bath at 70 °C for 15 min, then immediately placed in an ice bath for cooling for 5 min and then taken out. Four milliliters of ethyl acetate was accurately added to the mixture and shaken well. Using the corresponding reagents as the blank reference, the absorbance was determined at a wavelength of 546 nm by ultraviolet-visible spectrophotometry.
The total triterpenoid content was calculated according to the standard curve. The t-test volcano plot analysis function provided by the online analytical platform (
https://cnsknowall.com/#/Home/OmicsFlowVolcanoMap?pid=330502001 ) was adopted to conduct visualization analysis and significance test for differences on the detection data of total triterpenoid content in different Ganoderma strains. Based on the input grouping information of Ganoderma samples and abundance data of total triterpenoids content, this platform can accurately calculate the t-test, p-value, and fold change (FC). The numerical differences and statistical difference characteristics of total triterpenoids content among different Ganoderma strains are intuitively visualized in the form of a volcano plot. This analytical approach enables efficient and accurate screening and identification of Ganoderma strains with statistically significant differences in total triterpenoid content among samples. -
The process for construction of a sodium azide-induced G. lucidum mutant library and screening of triterpenoid high-yield strains is shown in Fig. 1. To determine the optimal conditions for SA mutagenesis of G. lucidum protoplasts, different SA concentration gradients from 0 to 5 mM were set, and the protoplast regeneration rate was determined under the same treatment time (3 h). The results showed that the average regeneration rate of G. lucidum protoplasts in the control group was 257.5 colonies per 1 × 105 protoplasts; with the increase of SA concentration, the protoplast regeneration rate gradually decreased. The protoplast survival rate in the 2 mM SA treatment group was 53.30% ± 8.61% (Table 1, Fig. 2a, b), close to the half-lethal rate, so this concentration (median lethal concentration, LC50) was determined as the optimal concentration for SA mutagenesis of G. lucidum protoplasts. The mutagenized single colonies at this concentration were picked and inoculated on slant medium for expansion culture (Fig. 2c), and preserved at low temperature for later use.
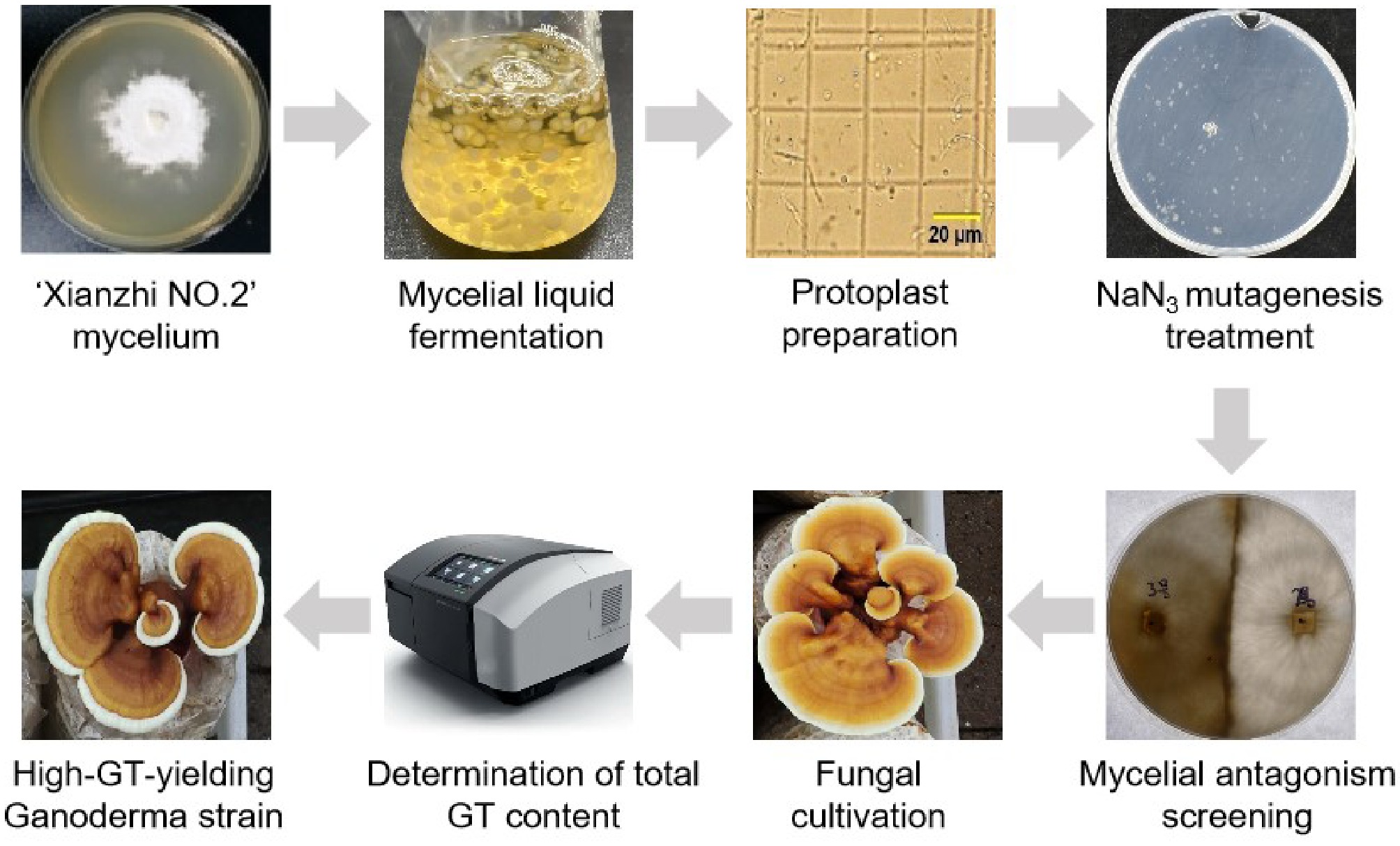
Figure 1.
Construction of a NaN3-induced mutant library of Ganoderma lucidum and screening for triterpene overproducers.
Table 1. Survival rate of Ganoderma lucidum protoplasts treated with different concentrations of sodium azide.
Treatment concentration
(mM)Biological replicate 1
(colonies)Biological replicate 2
(colonies)Biological replicate 3
(colonies)Relative regeneration
rate (%)0 > 300 254 261 100% 1 230 196 195 80.39 ± 7.74% 2 161 117 134 53.30 ± 8.61% 3 133 100 81 40.60 ± 10.22% 4 103 106 87 38.30 ± 3.97% 5 55 62 34 19.55 ± 5.66% Relative regeneration rate (%) = (Mean colony number of each treatment group/Mean colony number of the 0 mM control group) × 100%. 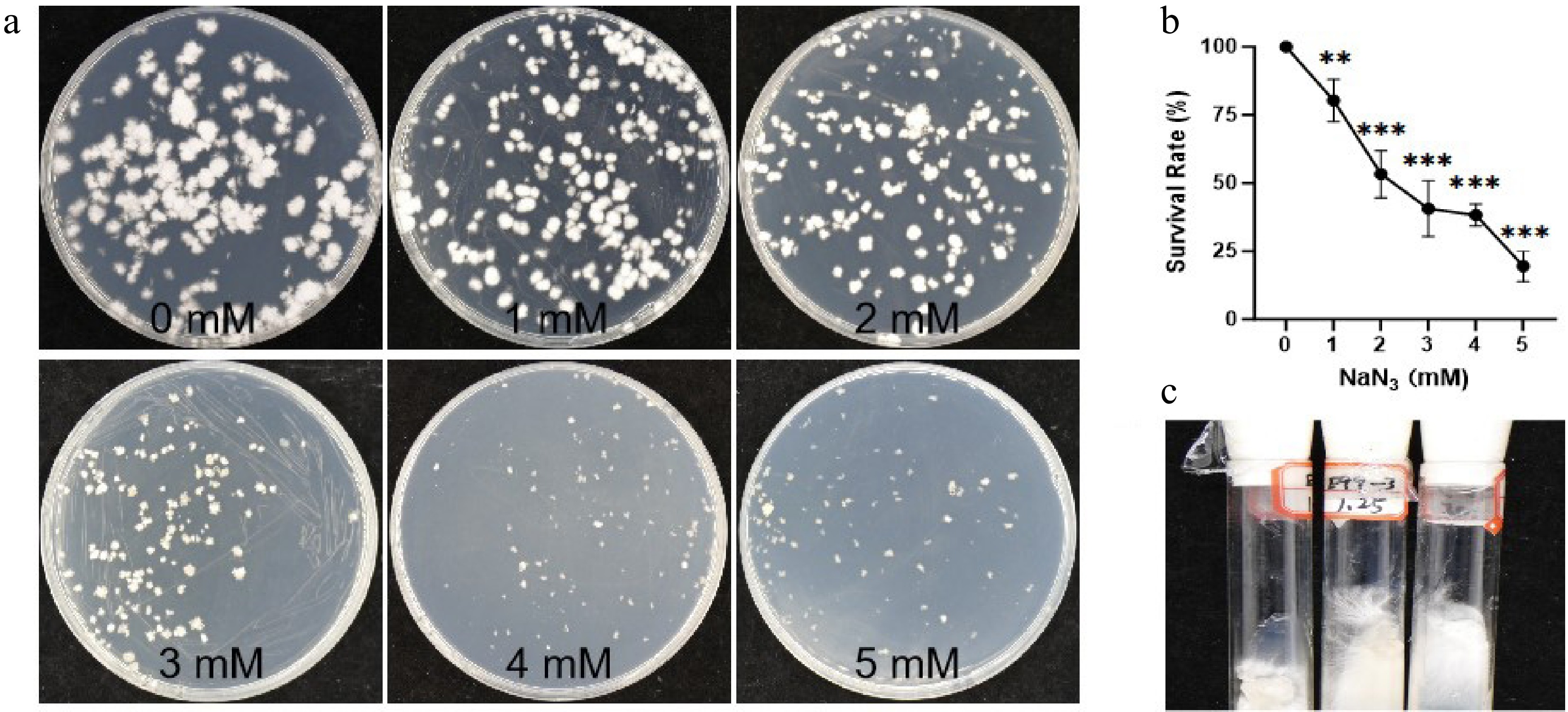
Figure 2.
Determining the LC50 through a NaN3 concentration gradient in Ganoderma lucidum protoplasts. (a) Mutagenesis and regeneration culture of protoplasts at different NaN3 concentration gradients (0, 1, 2, 3, 4, and 5 mM). (b) Survival rate (%) of G. lucidum at different NaN3 concentrations. The error bars indicate the standard deviations from three independent samples. ** p ≤ 0.01 indicates statistical significance, *** p ≤ 0.001 indicates statistical significance compared to the control group (CK). CK: G. lucidum strain 'Xianzhi No. 2'. (c) Scale-up cultivation of single colonies at the semi-lethal NaN3 concentration.
Screening results of antagonistic phenotypes of mutagenized strains
-
An antagonistic test is an important method to judge the somatic compatibility of filamentous fungi, which can be used to preliminarily infer the genetic relationship between strains[24,25]. In this study, the expanded mutagenized strains were inoculated on PDA solid medium, cultured for 5 d to prepare mycelial blocks, inoculated as shown in Fig. 3a, and cultured in a constant temperature incubator at 30 °C for 5 d. The width of the 'antagonistic line' formed by the positive control group ('Xianzhi No. 2' and 'Xianzhi No. 3') was 2.0−3.0 mm. Based on this, the antagonistic phenotypes of mutagenized strains and wild-type 'Xianzhi No. 2' were classified into Categories I−V: Category V had an antagonistic line width equivalent to that of the positive control group (2.0−3.0 mm); Category I had no obvious antagonistic phenomenon, with hyphae interlaced.
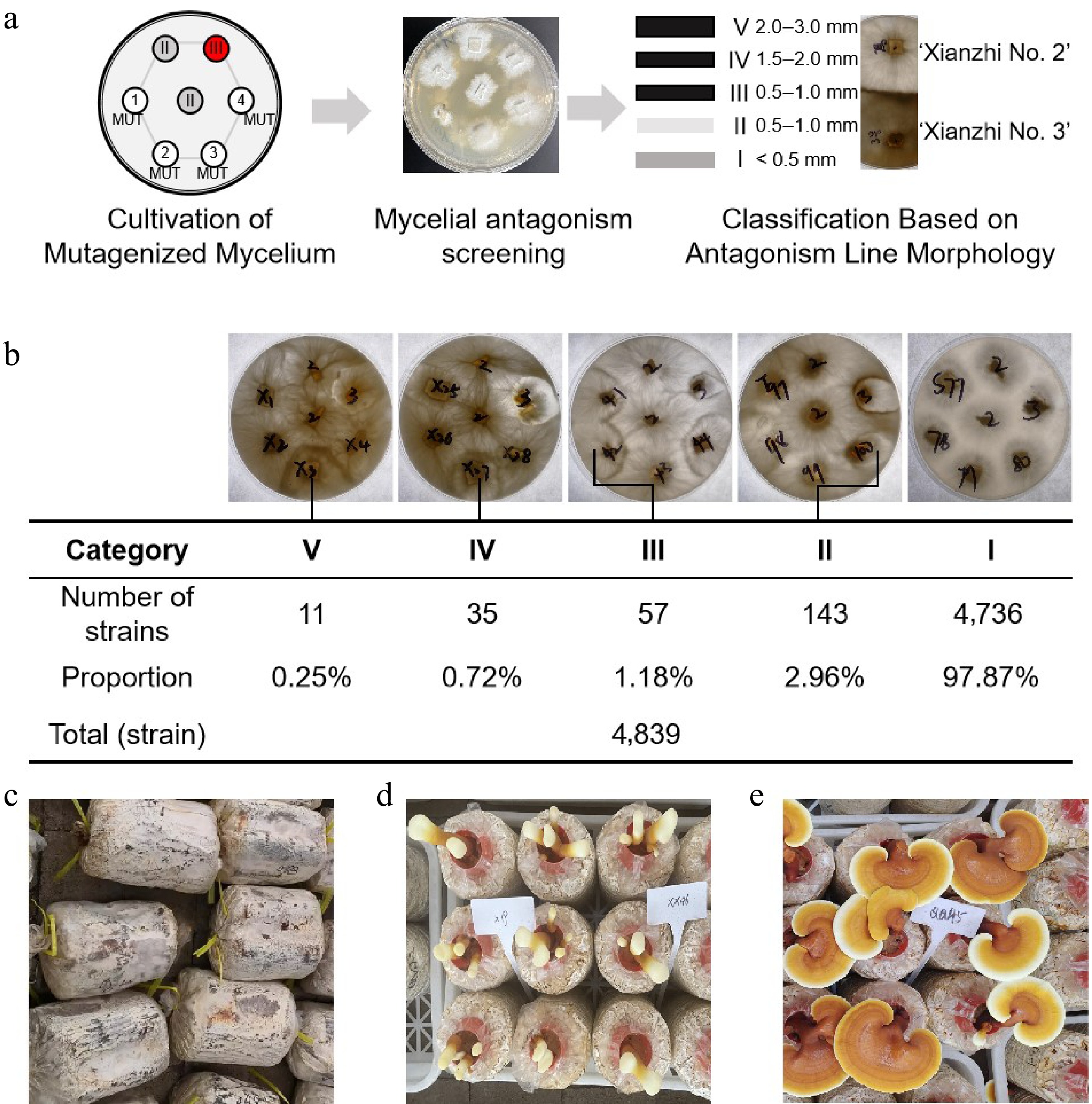
Figure 3.
Screening, cultivation, and selection of high-triterpenoid-yielding strains from NaN3-mutagenized Ganoderma lucidum. (a) Screening of G. lucidum mycelium with antagonistic phenotype. The grey circles represent the wild-type 'Xianzhi No. 2' strain; the red circles represent the wild-type 'Xianzhi No. 3' strain; and the numbers represent NaN3-induced mutant strains. (b) Screening and statistical analysis of antagonistic phenotypes in sodium azide-mutagenized Ganoderma strains. (c)−(e) Mycelial, photographic documentation, and observation of G. lucidum primordia and fruiting body growth stages.
Among 4,839 mutagenized strains, 11 strains showed Category V antagonistic phenotypes (accounting for 0.25% of the total), 35 strains for Category IV (0.72%), 57 strains for Category III (1.18%), 143 strains for Category II (2.96%), and the remaining 97.87% were Category I (no antagonistic line). A total of 217 strains with Category II−V antagonistic phenotypes and 259 randomly selected Category I strains (475 strains in total) were subjected to fruiting body cultivation, and finally 275 mutant strains capable of producing fruiting bodies were obtained (Fig. 3b−d).
Screening results of high and low triterpenoid-yielding mutant strains
-
Referring to the method for determining total triterpenoid content of G. lucidum in the Chinese Pharmacopoeia, a standard curve was established with oleanolic acid as the standard (Supplementary Table S1, Supplementary Fig. S1). The total triterpenoid content of 275 mutagenized strains was determined, and a volcano plot was drawn with the control group as reference (Fig. 4a, Supplementary Table S2).
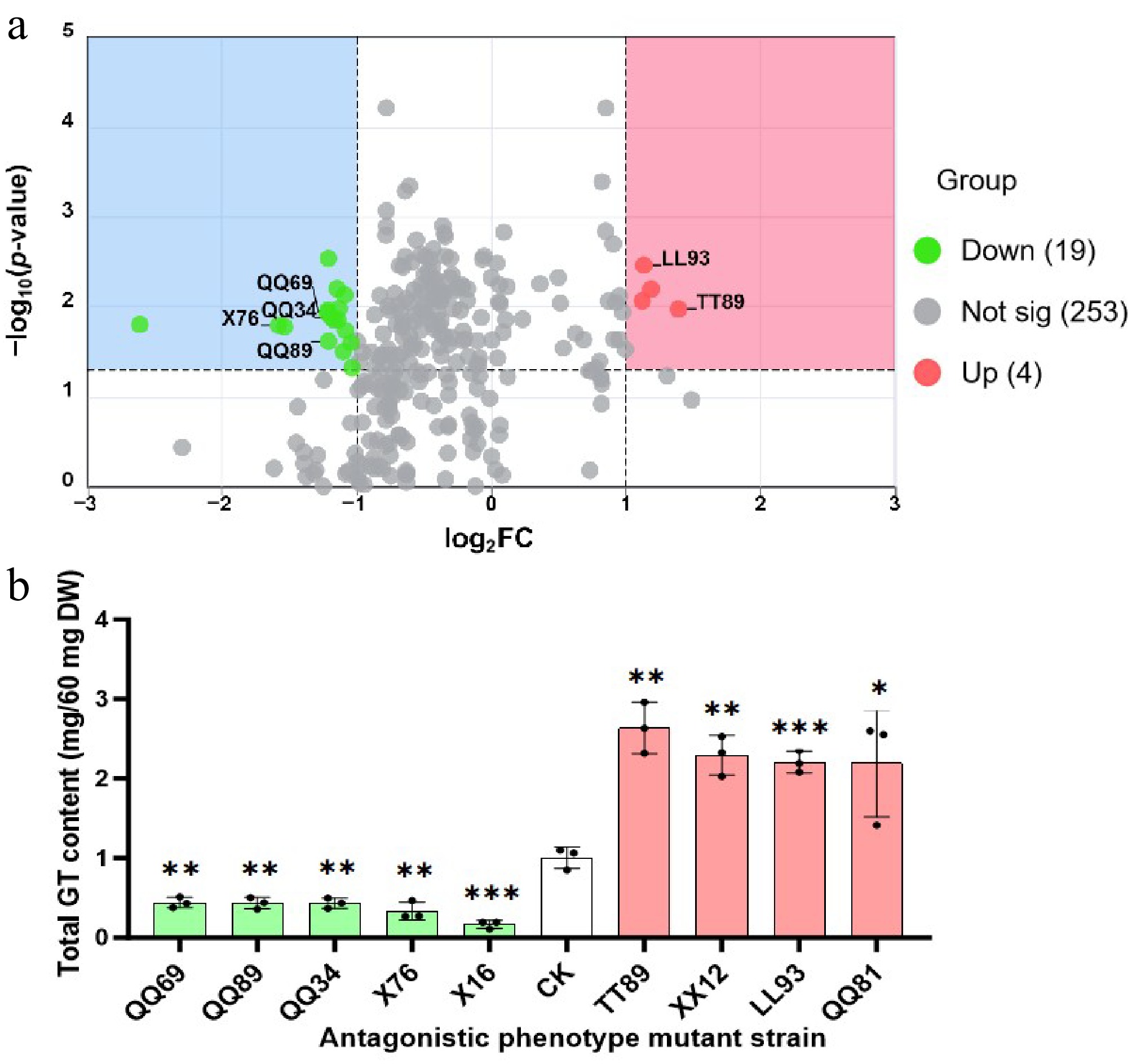
Figure 4.
Statistical analysis of total triterpenoid content in mutagenized strains. (a) Volcano plot of total triterpenoid content between G. lucidum high-triterpenoid-yielding mutants and the control group (CK). CK: G. lucidum strain 'Xianzhi No. 2'. This volcano plot illustrates the differential expression profiles of total triterpenoid content across 275 mutagenized strains. The x-axis represents the log2fold change |log2FC|, reflecting the relative abundance changes of triterpenoids between the two groups; the y-axis represents the −log10 p-value, which is used to evaluate statistical significance. Different metabolites with distinct significance levels and fold change magnitudes are distinguished by markers of varying colors and shapes in the plot: Red dots: represent strains with significant upregulation (p < 0.05 and log2FC > 1); Gray dots: represent strains with no substantial fold change or non-significant differences (|log2FC| ≤ 1 or p ≥ 0.05); Green dots: represent strains with significant downregulation (p < 0.05 and log2FC < −1). (b) Strains with significant differences in total triterpenoid content. Data are presented as mean ± standard deviation (mean ± SD) with three biological replicates per group (n = 3). The independent-samples t-test was performed to analyze the significance of differences between mutant strains and the control group (CK). * p ≤ 0.05 indicates a significant difference, ** p ≤ 0.01 indicates a highly significant difference, *** p ≤ 0.001 indicates an extremely significant difference, and ns indicates no significant difference (p > 0.05).
The volcano plot showed that four mutagenized strains with significantly upregulated total triterpenoid content were screened, namely TT89, XX12, LL93, and QQ81, all located in the upper right area of the plot, indicating that their content was significantly increased in the mutagenized group, with important biological significance (Fig. 4b). Among them, the total triterpenoid content of TT89 (Category II antagonistic phenotype) and LL93 (Category III antagonistic phenotype) was 262.27% and 219.48% higher than that of the control group, respectively, while that of the other two strains (XX12 and QQ81) was increased by 228.16% and 217.73%, respectively. Meanwhile, 19 mutagenized strains with significantly downregulated total triterpenoid content were screened, and some strains were labeled as follows: QQ69 (Category III), QQ89 (Category II), QQ34 (Category III), X76 (Category II), all located in the upper left area of the plot, indicating that their content was significantly decreased. Among them, the total triterpenoid content of X76 (the antagonistic strain) decreased to 33.41% of that in the control group, while the non-antagonistic strain X16 had the lowest total triterpenoid content, only 16.35% of that in the control group, which has important biological significance.
-
This study constructed a mutant library by SA chemical mutagenesis using G. lucidum protoplasts for the first time, obtained genetically modified strains by combining antagonistic screening technology, established a 'SA mutagenesis-antagonistic primary screening-total triterpenoid content verification' breeding system for G. lucidum, and obtained mutant strains with significantly increased total triterpenoid content through cultivation screening. The results showed that the survival rate of G. lucidum protoplasts was 53.30% ± 8.61% after treatment with 2 mM SA for 3 h, and 4.48% of the mutagenized strains showed antagonistic phenotypes under this condition. After cultivation validation, the total triterpenoid content of TT89 (Category II antagonistic phenotype) was the highest, 262.27% higher than that of the control group. This result confirms the feasibility of SA in G. lucidum mutagenesis breeding, and also verifies the efficiency of antagonistic screening as a primary screening method for G. lucidum genetic variation, providing a new technical path for G. lucidum germplasm innovation (Fig. 5).
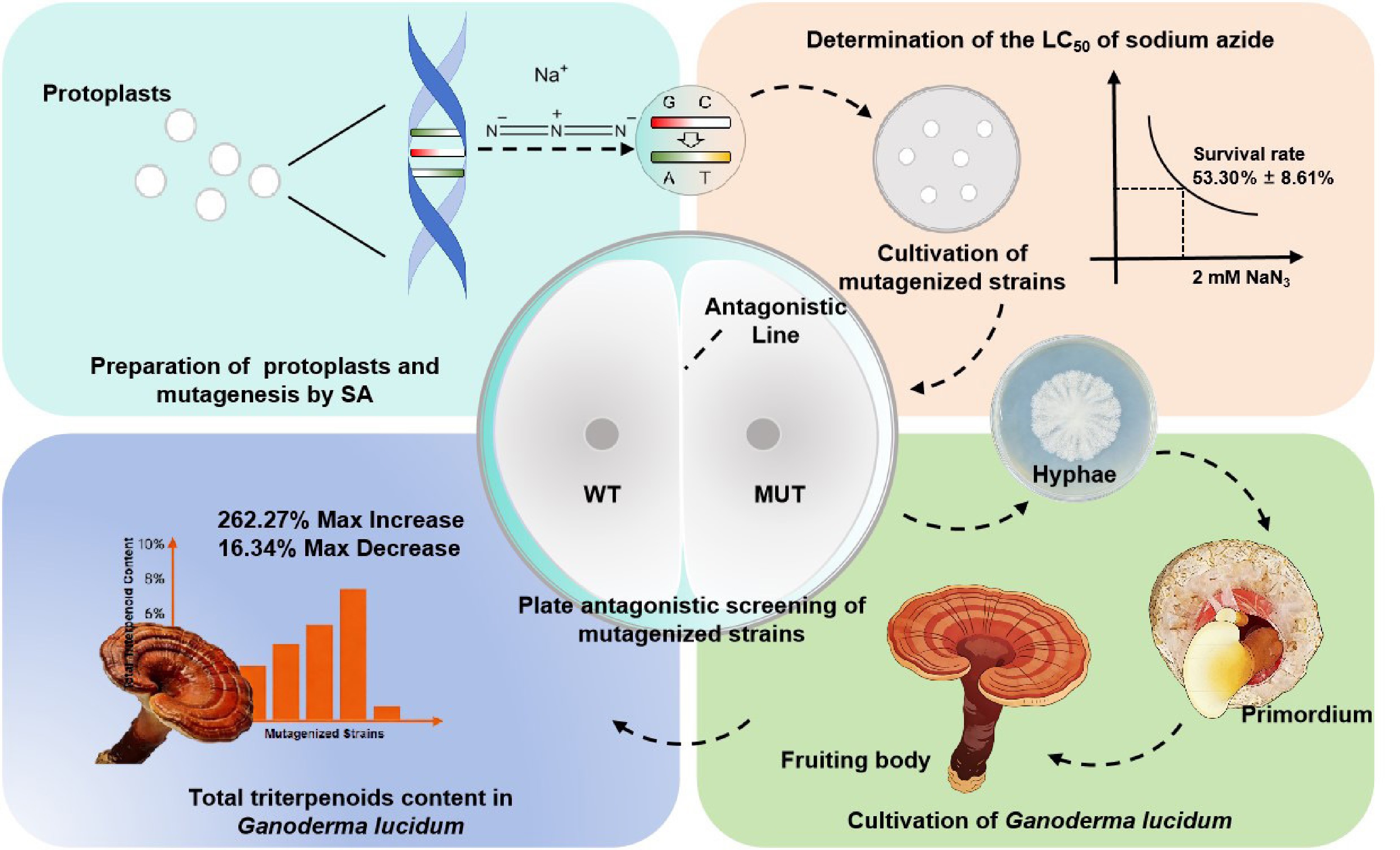
Figure 5.
Mutagenesis breeding with sodium azide and antagonistic screening of G. lucidum strains with high total triterpenoid content.
The selection of mutagen dosage is crucial for chemical mutagenesis breeding. Usually, the LC50 is used as the optimal mutagen dosage, which can induce high-frequency mutations while ensuring a sufficient screening population[34]. In this study, through concentration gradient experiments, 2 mM SA for 3 h was determined as the optimal condition for G. lucidum protoplast mutagenesis, further verifying the applicability of this mutagen dosage selection principle in medicinal organism breeding.
Existing studies mostly rely on molecular markers for mutant screening, which are cumbersome in operation, high in cost, and stringent in technical requirements[24,25,35]. In contrast, the 'SA mutagenesis-antagonistic primary screening' system established in this study utilizes the characteristic of filamentous fungi where hyphae inhibit each other's growth and form antagonistic lines, enabling rapid screening of genetically modified strains, significantly shortening the screening cycle, and being more suitable for industrial application. Meanwhile, the antagonistic effect not only reflects the degree of genetic variation within strains, but also enables the determination of phylogenetic relationships among strains and can also be used to assess whether mutations have occurred in strains[25,35]. In this study, 97.87% of the mutagenized strains exhibited no antagonistic phenotype. Among the four mutagenized strains with significantly upregulated total triterpenoid content (TT89, XX12, LL93, QQ81), TT89 and LL93 showed Class II and Class III antagonistic phenotypes, respectively. Specifically, the total triterpenoid content of TT89 showed the highest increase (262.24%) compared with the control group, while that of LL93 increased by 219.48%. Additionally, 19 mutagenized strains showed significantly downregulated total triterpenoid content, all of which decreased to less than 50% of that in the control group; four of these strains (QQ69, QQ89, QQ34, X76) displayed Class III, Class II, Class III, and Class II antagonistic phenotypes, respectively. These results indicated that there is no direct correlation between the presence/intensity of antagonistic phenotypes and total triterpenoid content in mutagenized strains with either upregulated or downregulated total triterpenoid levels, which is consistent with the random and non-directional nature of genetic material changes induced by chemical mutagenesis[36,37]. In this study, a total of 217 Ganoderma strains with antagonistic activity (classes II−V) were screened, and two high-triterpenoid-producing strains were finally obtained, with a screening efficiency of 1.84%. In contrast, among 259 randomly screened strains without an antagonistic phenotype, only two high-yield strains were obtained. These results indicate that although there is no direct correlation between antagonistic activity and triterpenoid yield, the antagonistic-based primary screening system can significantly enrich strains with genetic variation. Nevertheless, antagonistic phenotypes can serve as a primary screening indicator for genetic variation, facilitating the rapid enrichment of strains with significant genetic mutations, and thus the identification of strains with significant changes in total triterpenoid content, reducing subsequent screening efforts.
Point mutations induced by SA tend to fine-tune gene function rather than cause complete loss of function. It is speculated that such mutations may act on key rate-limiting enzyme genes in the G. lucidum triterpenoid synthesis pathway (e.g., the 3-hydroxy-3-methylglutaryl coenzyme A reductase gene HMGR) or branch pathways (e.g., the ergosterol synthesis pathway)[38−42], regulating total triterpenoid content by altering the synthesis of precursor substances. Previously, Peng et al. mutagenized G. lucidum using lithium chloride alone or in combination with Triton X-100, and the resulting mutant strains showed a maximum total triterpenoid content increase of 373.43%[20]. Dong et al.[19] mutagenized G. lucidum protoplasts with 0.3% lithium chloride, and the target component content of the obtained strains was 461.72% higher than that of the control group. In this study, the maximum total triterpenoid content increase in SA-mutagenized strains was 262.27%, which is lower than the above-mentioned studies, but still confirms the potential value of SA in G. lucidum triterpenoid breeding. The difference in the increase amplitude may be attributed to variations in the mechanism of action and mutation sites of different mutagens.
Notably, in addition to screening high-triterpenoid-yielding strains, this study also obtained mutant strains with significantly downregulated total triterpenoid content (the lowest being 16.34% of that in the control group). These negative mutant strains can serve as important experimental materials; by comparing their genomic, transcriptomic, and proteomic differences with the control group, key regulatory genes involved in triterpenoid synthesis can be accurately identified, providing a new perspective for studying the regulatory mechanisms of secondary metabolism in G. lucidum and filling the gap in current research on the negative regulation of triterpenoid synthesis in G. lucidum.
This study has certain limitations: First, antagonistic screening can only preliminarily determine the presence of genetic variation but cannot quantify the degree of variation. Furthermore, molecular biology techniques such as whole-genome sequencing and quantitative real-time PCR (qRT-PCR) are lacking to verify mutation sites and transcriptional changes of metabolic pathway genes. In addition, the heritable stability of the mutant strains has not been verified. Second, systematic determination of other phenotypic traits (e.g., biomass, contents of other bioactive components) in mutagenized strains was not performed. To address these shortcomings, future studies can use high-throughput sequencing technology to analyze the genetic variation characteristics of SA-mutagenized G. lucidum and clarify the expression changes of key genes in the triterpenoid synthesis pathway. Meanwhile, expanding the scope of phenotypic determination of mutagenized strains will help screen more specific superior strains. Moreover, it is necessary to carry out genetic stability verification of the screened mutant strains: continuous passage of the mutant strains for multiple generations (e.g., three to five generations) will be performed, and the phenotypic traits (such as ganoderic triterpenoid content) and genetic characteristics of each generation will be systematically detected to confirm that the excellent mutant traits can be stably inherited, laying a solid foundation for the industrial application of the mutant strains.
-
In conclusion, this study is the first to apply SA to G. lucidum protoplast mutagenesis breeding, establishing an efficient breeding system of 'chemical mutagenesis-antagonistic primary screening-total triterpenoid content determination' and successfully obtaining high-triterpenoid-yielding mutant strains. This study fills the application gap of SA in the mutagenesis of medicinal fungi, provides a theoretical basis and technical support for the innovation of germplasm resources of valuable medicinal fungi such as G. lucidum, and is of great significance for promoting the sustainable development of the medicinal fungi industry.
-
The authors confirm their contributions to the paper as follows: conceived the research: Xue Z; designed the study, performed the experiments, analyzed the data, and wrote the manuscript: Xu S; modified the manuscript: responsible for the determination of metabolite content and data preprocessing: Li Y; Liu L, Wei L; participated in the screening of the mutagenesis concentration system and preliminary validation: Guo X, Zang M, Diao Y, Liu S; coordinated and supervised the study: Zhou S, Liu C, Xu Z, Yang Z, Li Z. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
This research was supported by the Joint Funds of the Zhejiang Provincial Natural Science Foundation of China (Grant No. LHZSZ24H280003).
-
Zhou S and Li Z are researchers at Zhejiang Key Laboratory of Biological Breeding and Exploitation of Edible and Medicinal Mushrooms. In addition, Li Z holds shares in Zhejiang ShouxianGu Pharmaceutical Co., Ltd. and acts as a technical consultant for the company. These authors declare that despite these affiliations and financial involvements, all experimental work, data analysis and manuscript preparation in this study were completed independently, without any conflict of interest that could bias the research results.
-
accompanies this paper online at: https://doi.org/10.48130/mpb-0026-0009.
-
# Authors contributed equally: Shuhao Xu, Yanlin Li, Leijiao Liu
- Supplementary Table S1 Construction of the oleanolic acid standard curve.
- Supplementary Table S2 Determination of triterpenoid content in mutant Ganoderma lucidum strains.
- Supplementary Fig. S1 Construction of the oleanolic acid standard curve.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu S, Li Y, Liu L, Guo X, Wei L, et al. 2026. Construction, screening of a Ganoderma lucidum mutant library and identification of high-triterpenoid-yielding strains. Medicinal Plant Biology 5: e014 doi: 10.48130/mpb-0026-0009 

Construction, screening of a Ganoderma lucidum mutant library and identification of high-triterpenoid-yielding strains
- Received: 04 March 2026
- Revised: 05 April 2026
- Accepted: 22 April 2026
- Published online: 31 May 2026
Abstract: Ganoderic acids are the primary bioactive constituents of the medicinal fungus Ganoderma lucidum. Commercial cultivars typically have low ganoderic triterpenoid contents, creating an urgent industry need for high-triterpenoid germplasm. Here, we developed an efficient breeding pipeline for G. lucidum using protoplasts of strain 'Xianzhi No. 2' subjected to sodium azide (SA) mutagenesis, followed by antagonistic screening and total triterpenoid quantification to identify elite mutants. SA concentrations of 0–5 mM were tested, and 2 mM was selected as the half-lethal dose (53.30% ± 8.61% protoplast survival after 3 h treatment) for mutant library construction. Mutagenized isolates were categorized into five categories (I–V) by antagonistic phenotype. From 4,839 colonies, 217 with distinct antagonistic traits, and 259 randomly selected non-antagonistic isolates were cultivated for fruiting body production, yielding 275 fruiting-competent mutants. Quantitative analysis revealed that the high-yield mutant TT89 had a total triterpenoid content of 4.40%, a 262.27% increase over the wild-type (1.68% content). We also identified mutants with significantly reduced triterpenoid levels, including the non-antagonistic strain X16, which contained only 16.34% of the wild-type's triterpenoid content. This is the first report of an integrated breeding system for G. lucidum combining SA mutagenesis, antagonistic prescreening, and triterpenoid quantification. Our findings validate SA as an effective mutagen for G. lucidum and provide valuable mutant resources and technical support for germplasm improvement and triterpenoid biosynthesis research.