









-
Compared with grain crop yields, China's vegetable output has exceeded grain output, making vegetables the largest agricultural product since 2011[1]. The vegetable planting area in China has expanded from 6.3 × 106 hm2 in 1989 to 2.2 × 107 hm2 in 2022, accounting for 13% of the total crop sown area that year[2]. In 2022, vegetable output reached 8 × 108 t, accounting for more than 50% of the global total[3]. Vegetable fields are highly intensive agricultural land characterized by high water and fertilizer input and intensive rotation. The annual nitrogen fertilizer input exceeds 1,000 kg N·hm−2, far higher than the conventional application rate. A large amount of fertilizer nitrogen is lost from farmland into the surrounding environment, triggering a series of environmental problems, including N2O emissions[4]. Soil N2O emissions from protected vegetable fields are a major contributor to agricultural N2O emissions, with emissions from ordinary plastic greenhouses being approximately 6.9 times that of open-field soil[5]. Among them, protected vegetable fields account for more than 55% of the total vegetable planting area[6].
N2O is an important greenhouse gas with a global warming potential (GWP) about 273 times that of CO2 on a 100-year time scale[7]. Agricultural soil is one of the main anthropogenic factors driving increases in atmospheric N2O concentrations[8]. During crop production, long-term application of traditional chemical fertilizers can affect soil quality, including soil acidification and depletion of organic matter, thereby reducing crop yield and quality over time[9]. Organic fertilizers help maintain soil nutrients and improve soil structure, such as increasing soil total nitrogen (TN), soil organic matter (SOM), available phosphorus (AP), and available potassium (AK) contents, improving soil pH, and increasing soil water content and water-stable aggregate content[10]. Meanwhile, organic fertilizers have a slow-release effect. The combined application of chemical and organic fertilizers can coordinate the nutrient balance during crop growth, thereby optimizing nutrient utilization and improving crop yield. Studies have found that the levels of vitamin C and soluble sugar in tomatoes treated with a combination of Methylobacterium and a carbon-based organic fertilizer increased by 46.53% and 26.65%, respectively[11]. A weighted gene co-expression network analysis suggested that the application of organic fertilizers containing beneficial microorganisms regulates the expression of genes related to sugar and organic acid metabolism, alters sugar-acid metabolism, promotes sucrose accumulation, and citric acid degradation in fruits, and thus improves fruit quality[12]. Compared with the current situation in Denmark, where 70% of nitrogen input into farmland comes from organic fertilizers, China has even greater potential for reducing N2O emissions by integrating microbial technologies that utilize organic fertilizer resources. Current primary methods to reduce N2O emissions from farmland soil include adding nitrification inhibitors, applying biochar[13], and inoculating with plant growth-promoting rhizobacteria (PGPR) to mitigate N2O emissions. Inoculating bacterial agents with N2O-mitigating effects has emerged as a novel technology for decreasing emissions[14]. However, research on the 'microbe-organic fertilizer' synergistic system still has obvious shortcomings: existing studies mainly focus on the combined effects of a single bacterial strain and a specific organic fertilizer, and there is a lack of systematic investigation on the compatibility between different organic fertilizer substrates (such as livestock manure and decomposed straw) and functional bacterial strains. Gao et al.[15] found that inoculation with strain NRCB010 significantly promoted the growth of tomato seedlings. Yan et al.[16] showed that compared with the application of tobacco-specific compound fertilizer, the use of microbial fertilizer activated mineral elements in soil, promoted tobacco plant growth and development, increased potassium content and quality of tobacco leaves.
A study investigated the effects of co-inoculating Rhizobium spp. and Azospirillum brasilense on common bean cultivation, and found that the N2O flux in the inoculation treatment was significantly lower than that in the urea-only treatment, with an emission mitigating efficiency of 51%[17]. Bio-organic fertilizer containing T. guizhouense NJAU 4742 decreased bacterial amoA abundance, increased nosZ abundance, and decreased N2O emissions from soils[18]. The tested strain NRCB010 in this study is a PGPR carrying the nosZ gene. As a denitrifying bacterium that encodes the N2O reductase gene (nosZ), it can reduce N2O to N2 and is currently the only known biological sink for N2O[19]. Studies have shown that strain NRCB010 has strong nitrate reduction ability, as well as specific solubilizing capacities for inorganic calcium phosphate and zinc phosphate. Additionally, this strain exhibits resistance to drought and salinity under environmental stress. Therefore, it can promote crop growth while decreasing soil N2O emissions. Developing bacterial fertilizers with N2O-mitigating effects is one of the effective technical means for mitigating N2O emissions from farmland soil[20].
This study investigated the N2O-mitigating effect of strain NRCB010 inoculated into three different organic manures, aiming to provide practical guidance and a theoretical basis for mitigating greenhouse gas emissions from protected vegetable fields.
-
The candidate strain was Pseudomonas stutzeri NRCB010, isolated from paddy soil in Yixing, Jiangsu Province, China, and registered under the registration number CGMCC 19067. In previous studies, NRCB010 has demonstrated excellent N2O-mitigating and plant growth-promoting capabilities in both laboratory experiments and field in situ experiments, and it carries the nosZ gene. The NRCB010 strain was inoculated into nutrient broth medium (NBNS medium: beef extract 3.0 g·L−1, polypeptone 5.0 g·L−1, sodium nitrate 0.3 mmol·L−1, sodium succinate 4.4 mmol·L−1), and cultured with shaking at 28 °C and 180 r·min−1 for approximately 24 h. The OD600 value was adjusted to 1 for further use.
Sample collection
-
The soil was collected from a multi-span greenhouse vegetable field in the Full Mechanization Demonstration Base for Vegetable Production in Changshu, Jiangsu Province, China. The soil type is fluvo-aquic soil, and the previous crop was Bok choy, also known as Chinese cabbage (Brassica rapa subsp. chinensis). The collected soil samples were air-dried, ground, passed through a 2 mm sieve, and stored at 4 °C in a refrigerator before use. Gas samples were collected starting from the 2nd day of incubation, with sampling intervals of every 2 d at the initial active emission stage, and every 3–4 d in the later stage, yielding a total of 10 samplings.
Organic manures
-
Three types of organic manures were used, provided by Jiangsu Academy of Agricultural Sciences. In this study, the mushroom residue organic manure used mushroom residue as the core raw material; the high-temperature pretreated compost was prepared from livestock and poultry manure via high-temperature composting; the hydrochar carbon organic manure was produced from crop straw through hydrothermal carbonization. Mushroom residue organic manure, has total N, P2O5, and K2O ≥ 5%, organic matter ≥ 45%, pH of 7.37, EC of 5,810 μS·cm−1, total carbon (TC) of 18.59 g·kg−1, total nitrogen (TN) of 1.67 g·kg−1, and carbon-to-nitrogen ratio (C/N) of 11.13. High-temperature pretreated compost has a pH of 9.13, EC of 2,026.67 μS·cm−1, TC of 21.37 g·kg−1, TN of 2.66 g·kg−1, and C/N of 8.03. Hydrothermal carbon organic manure has N ≥ 1.6%, P2O5 ≥ 1.2%, K2O ≥ 2.2%, organic matter ≥ 40%, pH of 9.12, EC of 1,811 μS·cm−1, TC of 24.96 g·kg−1, TN of 2.42 g·kg−1, and C/N of 10.31.
Soil microcosm experiment
-
A total of seven treatments were set up in the experiment, with 16 replicates per treatment. The three organic fertilizers were added at equal nitrogen application rates, and each treatment also received 16 mL of sterile NBNS liquid medium (diluted 100 times). These seven treatments were no fertilizer (CK), supplemented with 16 mL sterile NBNS medium; mushroom residue organic fertilizer (MR), supplemented with 21 g MR and 16 mL sterile NBNS medium; high-temperature pretreated compost (CT), supplemented with 14 g of CT and 16 mL sterile NBNS medium; hydrothermal carbon organic fertilizer (HC10), supplemented with 15 g of HC10 and 16 mL sterile NBNS medium; the three combined treatments (MN, CN, HN) with each of three organic fertilizers received 16 mL NRCB010 bacterial solution. One hundred g of soil was weighed into a 500 mL culture flask, and organic fertilizer and bacterial solution (if applicable) were then added, then mixed thoroughly. Next, 100 g of soil was added, compacted, and then sterile water added to maintain soil moisture at 80% of the maximum field water-holding capacity. These soils were incubated in a 26 °C biochemical incubator in the dark. The conventional application rate of organic fertilizers in greenhouse vegetable fields in fluvo-aquic soil regions ranges from 300 to 600 t·hm−2. The converted field application rates in this study (364–546 t·hm−2) fall within this range and are consistent with actual production practices.
Quantitative real-time PCR (qPCR)
-
The gene copy numbers of 16S rDNA, amoB, AOA amoA, AOB amoA, nirS, nirK, nosZI, and nosZII in soil samples were determined using a quantitative real-time PCR instrument (Bio-Rad CFX96™ Optics Module, USA). Standard plasmids, with a concentration gradient of 102–108 copies·μL−1, were used as templates to construct the standard curve for qPCR amplification. The qPCR reaction system had a total volume of 25 μL, consisting of 5 μL of DNA template (diluted with double-distilled water and EASY Dilution at a specified ratio before addition), 12.5 μL of TB Green Premix Ex Taq II, 1 μL each of forward and reverse primers (10 μM), and 5.5 μL of sterile double-distilled water. For the negative control, sterile double-distilled water was used instead of the template DNA. All experimental results showed amplification efficiency greater than 90%, and a single peak in the melting curve. The primer sequences and reaction program for the different genes involved in the reaction are listed in Supplementary Table S1.
Determinations of greenhouse gas and physicochemical properties
-
N2O concentrations were analyzed by gas chromatography using an Agilent 7890B gas chromatograph equipped with an electron capture detector. The N2O emission flux and cumulative emission were calculated according to the formula. The pH of samples was measured using an HI 2211 pH meter (HANNA Instruments, Italy), and EC was measured using a DDS-307A conductivity meter (Inesa Scientific Instrument Co., Ltd, China). The NH4+-N content of samples was measured using a colorimetric method at 625 nm, whereas NO3−-N content was measured using a UV spectrophotometer, and the contents of TN and TC were analyzed using a Vario EL III element analyzer (Elementar, Germany).
DNA extraction and high-throughput sequencing of nosZI
-
Soil DNA was extracted using the HiPure Soil DNA Mini Kit (Magen, China). Based on the aforementioned qPCR analysis results, the nosZI region of 28 samples (forward primer sequence nosZ-F: CCCGCTGCACACCRCCTTCGA; reverse primer sequence nosZ-R: CGTCGCCSGAGATGTCGATCA) was sequenced on the Illumina Novaseq 6000/Miseq high-throughput sequencing platform. Raw paired-end sequencing data were assigned to each sample via unique barcodes. Quality control and clustering analysis were performed following standard protocols. Amplicon Sequence Variants (ASVs) are defined as exact sequence variants with only single-nucleotide differences, and were obtained using the DADA2 plugin in QIIME2 (
https://qiime2.org/ ). Briefly, after importing the sequencing data into the QIIME2 manifest file, demultiplexing was conducted with the q2-dmux plugin. Subsequently, the demultiplexed sequences were denoised and quality-filtered using the q2-dada2 plugin according to the standard DADA2 protocol, generating an ASV table and representative sequences. Chimeric sequences and singleton ASVs were removed, and the nosZI gene was functionally annotated using the GraftM tool. A total of 3,615,100 sequence reads were obtained from the 28 soil samples. After clustering analysis, 7,712 ASVs were identified.Data analysis
-
Data analysis and graphing were performed using Excel 2019 and GraphPad Prism 10.0, while IBM SPSS Statistics 23 was employed to conduct one-way analysis of variance (ANOVA), two-way ANOVA, and least significant difference (LSD) significance test for different treatments, with statistical significance of differences between treatments (p < 0.05). RStudio 2023.12.1 was used to generate stacked bar charts to visualize species abundance in bacterial communities and to analyze alpha and beta diversity indices. Beta diversity intra-group variation analysis implemented in R language using the vegan package, with PERMANOVA for statistical analysis, and the Bray-Curtis distance algorithm for dissimilarity calculation. The neutral community model was used to quantify the relative contributions of stochastic and deterministic processes to microbial community assembly. The co-occurrence network model was constructed using Gephi 0.9.2, with the calculation of network topological parameters (e.g., number of edges, average degree, connectivity). The structural equation model was analyzed using the linkET package in R.
-
The dynamic changes in soil N2O emission fluxes showed a consistent pattern (Fig. 1a–c), each treatment reached a peak on the second day of incubation. The CK had a peak of 218.23 ± 46.05 μg N·kg−1 dw soil·h−1, the MR was 739.22 ± 72.2 μg N·kg−1 dw soil·h−1, the MN was 339.39 ± 23.43 μg N·kg−1 dw soil·h−1, the HC10 was 558.78 ± 185.57 μg N·kg−1 dw soil·h−1, the HN was 460.56 ± 157.62 μg N·kg−1 dw soil·h−1, the CT was 744.14 ± 337 μg N·kg−1 dw soil·h−1, and the CN was 644.21 ± 266.56 μg N·kg−1 dw soil·h−1. The peak N2O emission of the HC10 treatment was compared with those of the CK and CT treatments using one-way ANOVA followed by the LSD multiple comparison method. The results showed that the peak emission of HC10 was significantly higher than that of the CK, and the peak emission of the MR was significantly higher than that of the CK (p < 0.05). These results indicate that applying organic fertilizer alone (especially HC10) significantly increases the peak N2O emission, and that different organic fertilizers have varying effects on the peak value. The peak N2O emission of the MN was significantly lower than that of the MR (p < 0.05), the CN was significantly lower than the CT (p < 0.05), and the HN was significantly lower than the HC10 (p < 0.05), demonstrating that inoculation with the NRCB010 strain could significantly decrease the peak N2O emission of each organic fertilizer treatment. Additionally, the peak N2O emission of the MN was significantly lower than that of the CN and HN (p < 0.05). The cumulative N2O emissions of the organic fertilizer treatments alone (MR, CT, HC10) were significantly higher than those of the non-inoculated control (CK) (p < 0.05). Among them, the HC10 treatment had the highest cumulative emission, followed by the MR treatment. The cumulative N2O emissions were significantly decreased (p < 0.05) after inoculation with the NRCB010 strain (Fig. 1d), with the mitigating rates in the order of MN (46.5%) > HN (27.8%) > CN (26.3%).
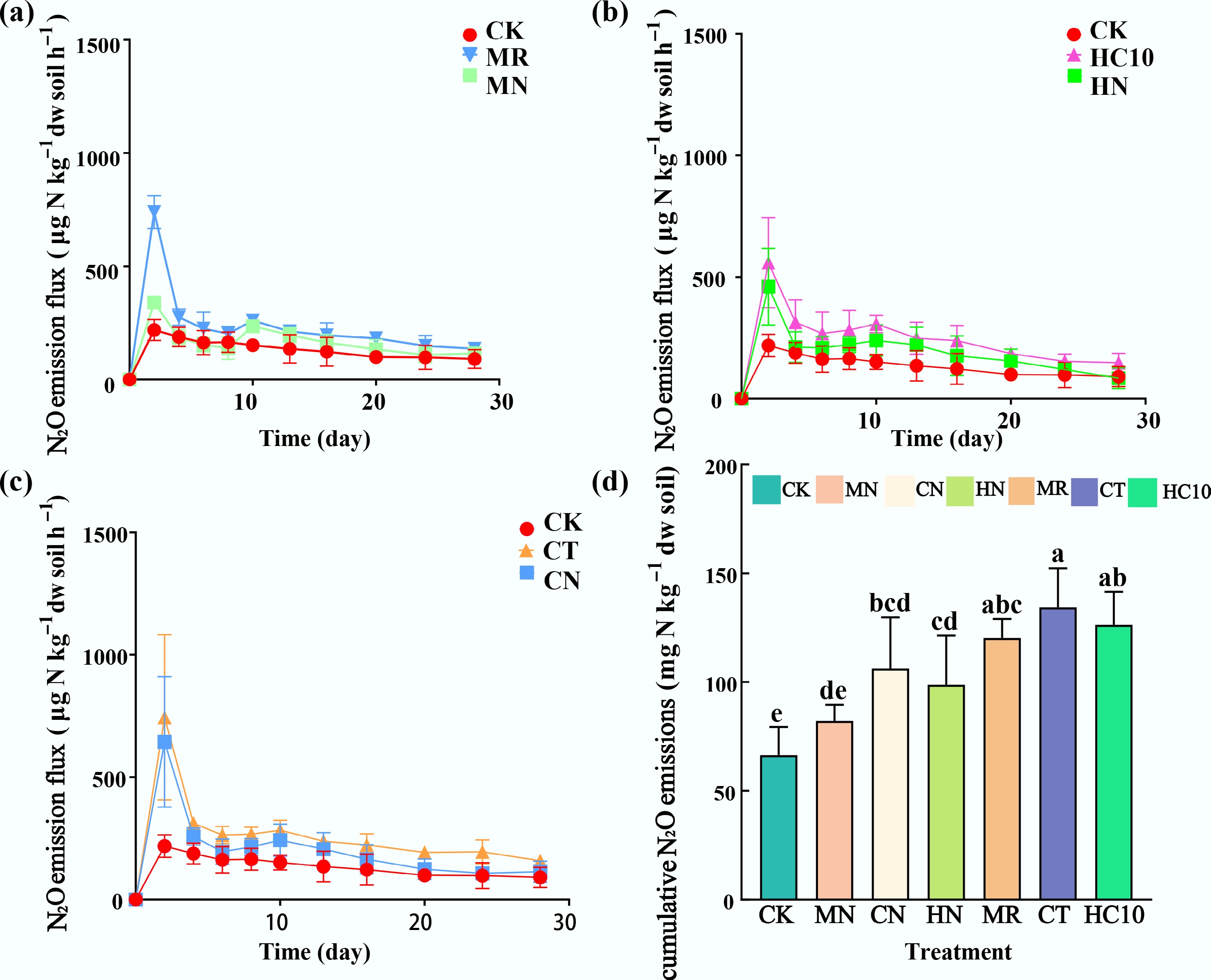
Figure 1.
(a)–(c) N2O emission flux, and (d) cumulative emission from soil. Different small letters above the bar indicated significant differences among treatments at the 5% level. CK: no fertilizer, MR: mushroom residue organic manure, CT: high-temperature pretreated compost, HC10: hydrothermal carbonized organic manure, MN: mushroom residue organic manure with NRCB010, CN: high-temperature pretreated compost with NRCB010, HN: hydrothermal carbonized organic manure with NRCB010.
Among the inoculation treatments, the MN treatment exhibited the optimal N2O mitigating effect—achieving a remarkable mitigating rate of 46.5% compared to the non-inoculated control, which is statistically significant. This outstanding performance not only demonstrates that the MN treatment is a highly promising combination for the practical control of N2O emissions from agricultural soils but also lays a critical foundation for the subsequent in-depth exploration of its underlying mechanisms. The enhanced N2O mitigation observed in the MN treatment is likely atrributable to the application of the mushroom residue organic manure, as it provides abundant readily available carbon substrates, promotes an optimal C/N ratio, and fosters an improved soil structure. These not only promote the proliferation of Pseudomonas stutzeri NRCB010—altering the abundance of the nosZI gene and modifying soil physicochemical properties (e.g., pH and EC)—but also contribute to efficient N2O mitigation through the enhanced nosZI gene abundance and synergistic interactions among functional microbial communities. The significant advantage of the MN treatment in N2O mitigation indicates a more efficient synergistic interaction between the mushroom residue organic manure and the NRCB010 strain. Gao et al.[15] reported that the combination of Pseudomonas stutzeri strains and organic fertilizers can reduce N2O emissions from vegetable soils by increasing the abundance of the nosZ gene. Future studies will systematically dissect the key biological processes driving its notable mitigating effect, thereby providing scientific support for the rational design and application of high-efficiency N2O mitigating technologies.
Soil physicochemical properties
-
Soil pH values were lower in the organic fertilizer treatments without bacterial inoculation (Table 1). The MR treatment (6.97) was significantly lower than the CK (7.11) (p < 0.05). After inoculation with the strain, the pH value of the MN treatment (7.02) was slightly higher than that of MR. The pH values of the CN (7.15) and HN (7.18) treatments were significantly higher than those of CT and HC10 (p < 0.05), with the HN treatment having the highest pH value. Regarding EC, the MR treatment (6.18 mS·cm−1) had a significantly higher EC than the other treatments (p < 0.05). After inoculation with the strain, the EC value of the MN treatment decreased to 4.45 mS·cm−1, a 28.0% reduction compared with MR (p < 0.05). It may degrade some soluble salts in the soil, thereby reducing the soil EC value, and improving the soil physicochemical environment[21]. This improvement reflects the synergistic effect between the strain and the mushroom residue organic manure. MR alleviates soil salinization stress through its high organic matter content, while the strain directly degrades soil-soluble salts via nitrate reduction metabolism under secondary salinization induced by nitrate. Both of them synergistically optimize the soil physicochemical environment, laying a foundation for N2O mitigation.
Table 1. Physicochemical properties of soil
Time Treatment pH (H2O) EC (mS·cm−1) NO3−-N (mg·kg−1) NH4+-N (mg·kg−1) 2 d CK 7.11 ± 0.02abc 2.84 ± 0.34e 28.41 ± 2.97e 11.44 ± 0.33b MR 6.97 ± 0.07d 6.18 ± 0.23a 68.15 ± 8.22ab 49.39 ± 9.93a CT 7.10 ± 0.06abc 3.70 ± 0.34d 48.77 ± 5.17cd 43.71 ± 13.14a HC10 7.07 ± 0.07bc 5.25 ± 0.28b 82.96 ± 14.95a 13.74 ± 0.95b MN 7.02 ± 0.06cd 4.45 ± 0.25c 50.95 ± 3.36bcd 42.28 ± 18.05a CN 7.15 ± 0.07ab 3.34 ± 0.46d 42.65 ± 7.54de 43.76 ± 21.72a HN 7.18 ± 0.08a 3.77 ± 0.37d 64.31 ± 24.38bc 13.28 ± 2.82b 10 d CK 6.81 ± 0.10d 3.45 ± 0.62d 31.78 ± 7.40e 8.62 ± 0.61c MR 6.82 ± 0.10d 5.96 ± 0.44a 85.35 ± 6.43a 51.28 ± 15.77a CT 7.12 ± 0.06b 3.49 ± 0.15d 46.26 ± 3.69cde 54.40 ± 17.12a HC10 6.93 ± 0.02c 4.92 ± 0.41b 39.08 ± 4.16de 26.78 ± 17.36bc MN 7.12 ± 0.04b 4.14 ± 0.28c 55.68 ± 20.75b 41.97 ± 25.72ab CN 7.24 ± 0.06a 3.25 ± 0.21d 50.64 ± 8.50bcd 51.48 ± 7.24a HN 7.15 ± 0.04ab 3.09 ± 0.34d 61.88 ± 8.70b 32.22 ± 12.32ab 20 d CK 6.67 ± 0.1d 4.14 ± 0.5c 28.91 ± 4.9b 11.39 ± 9.9b MR 6.79 ± 0.1c 6.08 ± 0.3a 53.392 ± 10.6a 28.37 ± 13.1a CT 6.94 ± 0.02b 3.56 ± 0.3d 49.23 ± 11.6a 30.46 ± 7.7a HC10 6.88 ± 0.03bc 4.99 ± 0.6b 61.70 ± 9.5a 22.89 ± 6.4ab MN 7.05 ± 0.1a 4.44 ± 0.3c 50.64 ± 8.2a 34.76 ± 17.1a CN 7.12 ± 0.1a 3.22 ± 0.3d 48.33 ± 11.5a 26.58 ± 10.6ab HN 7.07 ± 0.1a 3.55 ± 0.2d 50.18 ± 9.4a 29.18 ± 2.6a 28 d CK 7.42 ± 0.1a 2.86 ± 0.5c 34.98 ± 2.5c 8.45 ± 1.6c MR 6.97 ± 0.1bc 5.77 ± 1.0a 60.02 ± 12.7a 12.13 ± 1.1c CT 7.06 ± 0.1b 3.01 ± 0.1c 51.38 ± 6.1a 43.23 ± 10.4a HC10 6.84 ± 0.04c 4.63 ± 0.2b 57.71 ± 4.4a 14.28 ± 1.8c MN 6.85 ± 0.1c 4.08 ± 0.4b 51.59 ± 3.8ab 32.18 ± 11.6b CN 6.86 ± 0.2c 2.95 ± 0.4c 44.21 ± 8.78bc 35.45 ± 9.3ab HN 6.91 ± 0.1bc 2.62 ± 0.2c 45.61 ± 9.4bc 16.63 ± 3.7ac CK: no fertilizer, MR: mushroom residue organic manure, CT: high-temperature pretreated compost, HC10: hydrothermal carbonized organic manure, MN: mushroom residue organic manure with NRCB010, CN: high-temperature pretreated compost with NRCB010, HN: hydrothermal carbonized organic manure with NRCB010. Different lowercase letters indicate significant differences among treatments at the 5% level. Among the non-inoculated treatments, HC10 had the highest NO3−-N content, while MR had the highest NH4+-N content. After inoculation with the strain, the NO3−-N content in the MN treatment was significantly decreased by 25.2% compared with MR (p < 0.05), and the ammonium nitrogen content was slightly lower than that of MR.
The abundance of nitrogen cycling functional genes in soils
-
On the second day of the experiment, inoculation with strain NRCB010 significantly increased the abundance of the nosZI gene in the soil (p < 0.05) (Fig. 2g). Among the treatments inoculated with strain NRCB010 (MN, CN, and HN), the abundance of the nosZI gene was significantly higher than that in the non-inoculated treatments (MR, CT, and HC10) (p < 0.05). Specifically, the MN treatment exhibited the highest nosZI gene abundance (1.8 × 107 copies·g−1 dry soil), which was a 125% increase compared with the MR treatment (8 × 106 copies·g−1 dry soil). With extended incubation time, the abundance of the nosZI gene across all treatments showed a downward fluctuation. On the 28th day of incubation, the abundance of the nosZI gene in the MN treatment remained significantly higher than that in other treatments (p < 0.05).
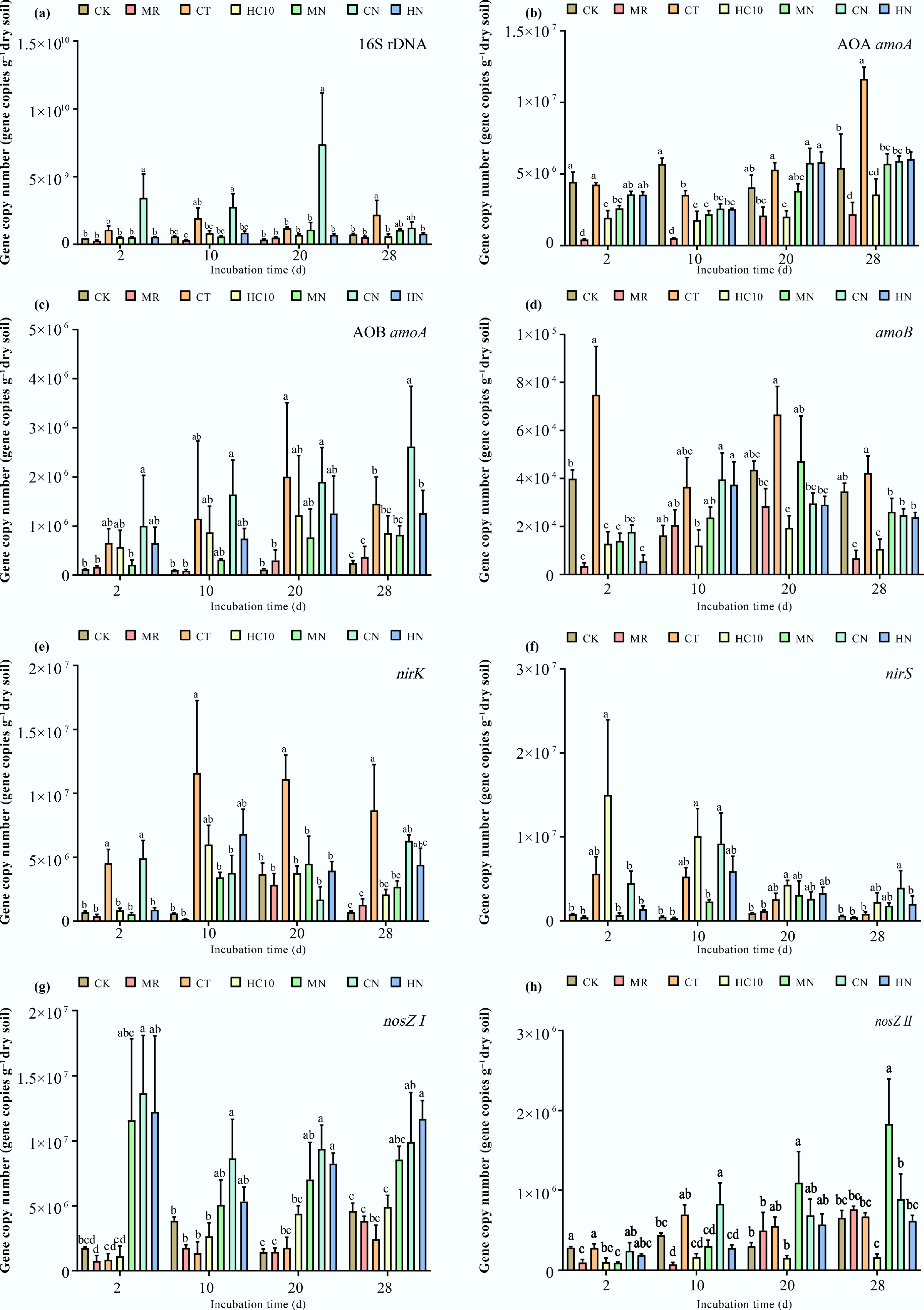
Figure 2.
Abundance of nitrogen cycling functional genes in soils. (a) The abundance of 16S rDNA in soils. (b) The abundance of AOA amoA genes in soils. (c) The abundance of AOB amoA genes in soils. (d) The abundance of amoB genes in soils. (e) The abundance of nirS genes in soils. (f) The abundance of f nirK genes in soils. (g) The abundance of nosZI genes in soils. (h) The abundance of nosZII genes in soils. Different lowercase letters above the bar indicated significant differences among treatments at the 5% level. CK: no fertilizer, MR: mushroom residue organic manure, CT: high-temperature pretreated compost, HC10: hydrothermal carbonized organic manure, MN: mushroom residue organic manure with NRCB010, CN: high-temperature pretreated compost with NRCB010, HN: hydrothermal carbonized organic manure with NRCB010.
Since the N2O reductase encoded by the nosZI gene is the key enzyme for converting N2O to N2[22], its high abundance can directly promote the reduction of N2O to N2. Therefore, the high expression level of the nosZI gene in the MN treatment may be the core mechanism underlying its optimal N2O-mitigating effect. The decrease in gene abundance over time may be related to the gradual consumption of soil nitrogen and to insufficient microbial metabolic substrates. The results also indicated that inoculation with NRCB010 significantly increased the abundance of the nosZI gene in the soil, thereby decreasing soil N2O emissions. A previous study showed that inoculation with NRCB010 decreases N2O emissions and promotes tomato growth simultaneously[15]. Additionally, n-hexadecanoic acid, a plant rhizosphere exudate, can not only decrease N2O emissions from farmland soils by itself, but also synergize with NRCB010 to promote plant growth and the colonization of PGPR in the plant rhizosphere[23].
Community composition and diversity of nosZI-carrying microorganisms in soils
-
To characterize the taxonomic composition of nosZI-carrying microorganisms at the phylum, class, and order levels, Fig. 3 illustrates their species composition. The dominant phyla may include Verrucomicrobiota, Pseudomonadota, Gemmatimonadota, Planctomycetes, etc. (Fig. 3a). Among them, Pseudomonadota and Verrucomicrobiota showed the most significant changes. In the treatments inoculated with strain NRCB010 (MN, CN, HN), the relative abundance of Pseudomonadota were significantly higher than those in the non-inoculated treatments (MR, CT, HC10) (p < 0.05). Specifically, the MN treatment had the highest proportion of Pseudomonadota (35%), which was 94.4% higher than that in the MR treatment (18%). Since strain NRCB010 belongs to Pseudomonas stutzeri and is classified in Pseudomonadota, its colonization and reproduction after inoculation are the main reasons for the increased abundance of this phylum in soils. The relative abundance of Verrucomicrobiota in the non-inoculated treatments were significantly higher than those in the inoculated treatments (p < 0.05). It is speculated that Verrucomicrobiota may prefer nitrogen and SOM-rich environments, and the introduction of the strain altered the soil microenvironment and inhibited bacterial growth in this phylum. At the class level, the relative abundance of Gammaproteobacteria in inoculated treatments increased significantly. In contrast, the relative abundance of Opitutae in Verrucomicrobiota decreased significantly, consistent with the changing trend at the phylum level (Fig. 3b). The dominant orders may include Pseudomonadales, Enterobacteriales, and Xanthomonadales. Pseudomonadales accounted for a prominent proportion in the MN and MR treatments, while Enterobacteriales were more dominant in the CK (Fig. 3c).
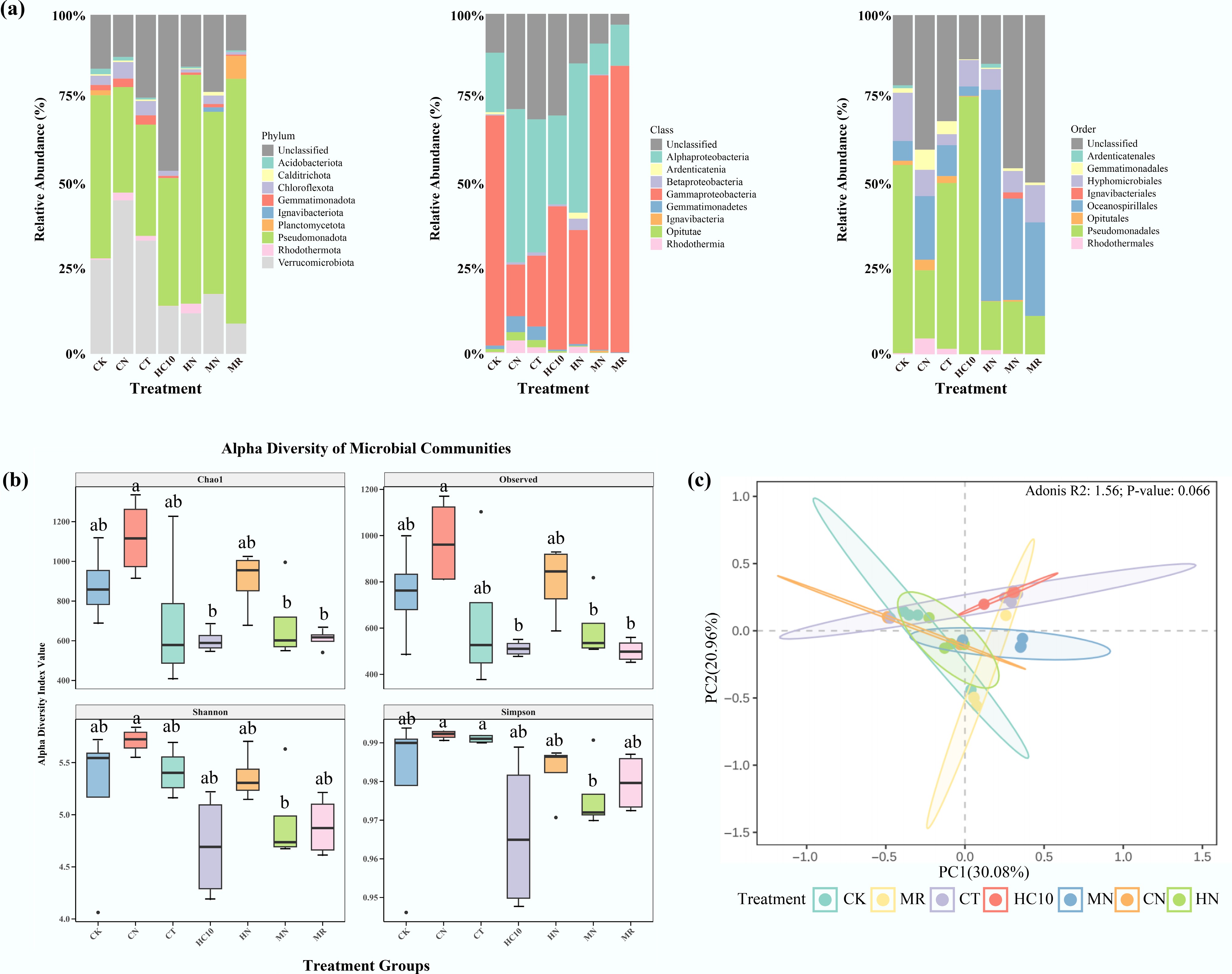
Figure 3.
Community composition of nosZI carrying microorganisms at (a) phylum, (b) class, and (c) order levels, and their (d) α diversity, and (e) β diversity in soils. CK: no fertilizer, MR: mushroom residue organic manure, CT: high-temperature pretreated compost, HC10: hydrothermal carbonized organic manure, MN: mushroom residue organic manure with NRCB010, CN: high-temperature pretreated compost with NRCB010, HN: hydrothermal carbonized organic manure with NRCB010.
The relative abundance of Verrucomicrobiota decreased significantly, dropping from 22%–25% in non-inoculated treatments to 10%–15% in inoculated treatments. Given that Verrucomicrobiota was enriched in non-inoculated environments with both high nitrogen and organic matter content, it is expected that certain members of this phylum may possess metabolic pathways for N2O production. The introduction of strain NRCB010 may have promoted the consumption of NO3− and SOM, thereby inhibiting the growth or metabolic activity of N2O-producing microorganisms.
Pseudomonadota is one of the main host phyla for the nosZI-carrying microorganisms in soils. In this study, the abundance of the nosZI gene in the MN treatment was significantly higher than that in the MR treatment. The enrichment of Pseudomonadota directly increased the abundance of the nosZI gene and enhanced the N2O-reducing capacity. Some taxa within the Verrucomicrobiota may harbor the amoA gene, whose expression is involved in ammonia oxidation and N2O production. The abundances of AOA amoA and AOB amoA genes in non-inoculated treatments (MR, HC10) were significantly higher than those in inoculated treatments. Specifically, the abundance of the AOB amoA gene in the MR treatment was 18.7% higher than that in the MN treatment. It is hypothesized that taxa carrying the amoA gene within Verrucomicrobiota proliferate extensively in non-inoculated environments, thereby promoting ammonia oxidation and thus increasing N2O emissions. In contrast, after inoculation with NRCB010, the soil pH tended to be neutral, and EC decreased, which altered the habitat for Verrucomicrobiota, inhibited its growth and the expression of the amoA gene, thereby decreasing N2O production.
Different treatments have significant effects on the community diversity of nosZI-carrying microorganisms as reflected in both α and β diversity. In terms of α diversity (Fig. 3d), the HC10 treatment significantly increased the Chao1 and Observed indices as well as Shannon and Simpson indices, whereas the MN and HN treatments significantly decreased community diversity. The HC10 treatment showed the most significant fluctuation in the Shannon index and an increase in the Simpson index, indicating that community stability was poor in the high-temperature pretreated compost environment and that competition among dominant species was intense[24]. The Chao1 indices for the CT treatment were relatively high, suggesting that high-temperature-pretreated compost may be more likely to increase species richness[25]. In terms of β diversity, the bacterial community compositions of the HC10, MN, and HN treatments were clearly separated from those of other treatments, indicating that these treatments significantly altered community composition. Among them, the HC10, MN, and HN groups showed strong internal aggregation, while the CK, MR, and CT treatments overlapped to some extent. The cumulative variance explained by PCoa1 and PCoa2 was 51.4% (Fig. 4e), which could well reflect the differences in community composition. Treatments of the same type were close in distance, while the distance between uninoculated treatments with different organic fertilizers was relatively far, indicating that the NRCB010 strain had a much greater impact on the community composition than the organic fertilizers. The introduction of the strain might be the core factor driving changes in the community structure[26]. The significance levels of β diversity-based community structure differences among treatment groups, determined by PERMANOVA, are shown in Supplementary Table S2.
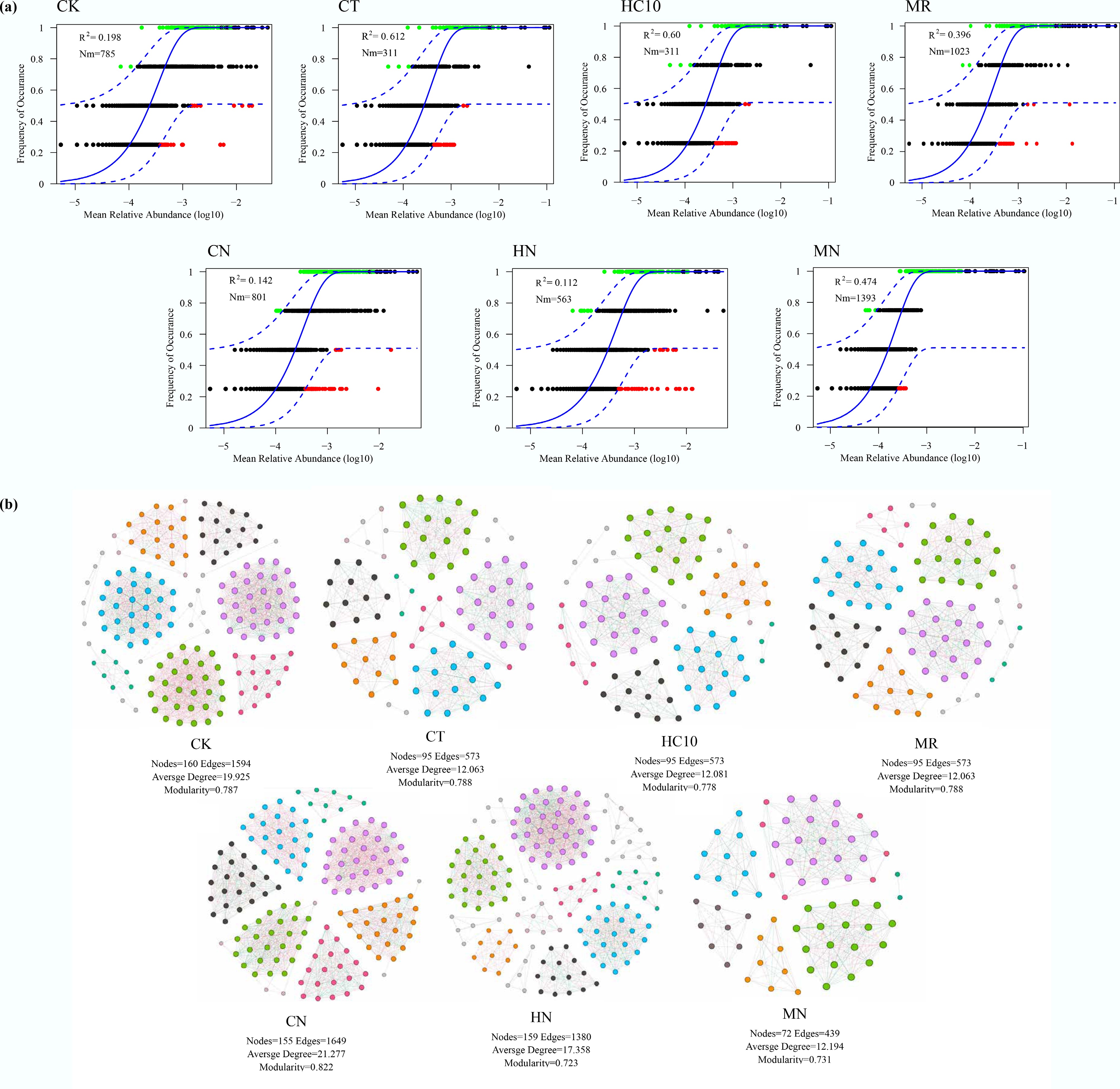
Figure 4.
The community assembly based on (a) neutral theory, and (b) co-occurrence network of nosZI carrying microorganisms. R2 indicates the model fit, and Nm represents the product of metacommunity size and migration time. CK: no fertilizer, MR: mushroom residue organic manure, CT: high-temperature pretreated compost, HC10: hydrothermal carbonized organic manure, MN: mushroom residue organic manure with NRCB010, CN: high-temperature pretreated compost with NRCB010, HN: hydrothermal carbonized organic manure with NRCB010.
Community assembly processes and key network modules of nosZI-carrying microorganisms
-
Neutral community modeling[27] analysis revealed that the CT and HC10 treatments exhibited the highest R2 values, followed by the MR and MN treatments (Fig. 4a). The community structures under these four treatments showed a better fit with the neutral model, and stochastic processes had greater explanatory power for community assembly. The low R2 values of the CK, CN, and HN treatments indicate that deterministic processes play a more important role. In the CK treatment, the absence of organic fertilizer leads to reduced soil nutrient availability and intense environmental filtering. In the CN and HN treatments, NRCB010 colonization alters soil pH, EC, and carbon-nitrogen supply, thereby reshaping the community structure through interspecific competition and synergistic interactions. The high R2 values of the CT and HC10 treatments suggest that stochastic processes dominate community assembly, which may be related to the weak selective pressure of these two organic fertilizers on microorganisms due to their physicochemical properties—such as the low C/N ratio of CT and the high carbon content of HC10. The MR and MN treatments had the highest Nm values, suggesting that inter-community species migration and diffusion capacities were strongest under these treatments, with higher species exchange frequency and a more uniform species distribution. A synergistic analysis showed that stochastic processes dominated the community assembly in the CT and HC10 treatments, with moderate species diffusion capacity; in the MR and MN treatments, stochastic processes mainly shaped the communities. Although species diffusion capacity was relatively strong in the CK, CN, and HN treatments, deterministic processes remained a key regulator of the community structure.
Through nosZI co-occurrence network analysis, the CK (160 nodes, 1,594 edges) showed significantly higher average degree (19.925) than the other treatments, indicating more frequent and intensive bacterial interactions (Fig. 4b). The CN (modularity = 0.822) had significantly higher modularity than the other treatments, suggesting clearer functional module division of the microbial community and potentially more distinct functional subgroups formation. In contrast, the HN (modularity = 0.723) had relatively low modularity with weak differentiation of community functional modules. The MN had the fewest nodes and edges, with both average degree and modularity at low levels, reflecting relatively sparse interactions among bacterial communities and weak modular organization in this treatment. These results indicate significant differences in the interaction patterns and the organizational levels of functional modules within bacterial communities. Such differences may be related to the cooperative metabolism, resource competition patterns, or the establishment of symbiotic relationships among microorganisms. It also implies that different treatments have differential shaping effects on the structure and functional network of microbial communities by influencing the interactions among microorganisms. Studies have shown that under different nitrogen input gradients, changes in the network structures and microbial traits of bacterial and fungal communities reveal that alterations in the life-history strategies of soil bacteria regulate the reduction in the complexity of their co-occurrence networks, uncovering the trait-based regulatory mechanism underlying microbial interspecific interactions and co-occurrence networks[28].
Correlation analysis between environmental factors and N2O emissions
-
The correlation analysis indicated that N2O emissions were significantly negatively correlated with the abundance of nosZI (Fig. 5a) and significantly positively correlated with soil NO3−-N, pH, and EC. EC indirectly modulates the expression level and activity of the nirS gene by regulating soil osmotic pressure and the community structure of nirS-type denitrifiers, thereby influencing the conversion efficiency of NO to N2O during denitrification and ultimately regulating N2O emissions. Greenhouse vegetable soils are highly susceptible to secondary salinization due to multiple factors, including long-term excessive nitrogen fertilization, inappropriate irrigation practices, and weak leaching under the enclosed conditions of greenhouses. The predominant anion in greenhouse vegetable soils is NO3−, accounting for more than 60% of the total soluble salts. NRCB010 is an N2O-reducing bacterium with strong NO3− reducing capability. Through its intrinsic NO3−- reducing metabolic pathway, NRCB010 can progressively decrease accumulated NO3− in soil, which is further converted into gaseous compounds such as N2. This process not only directly decreases the accumulation of NO3− in soil, thereby alleviating the degree of NO3−-dominated secondary salinization in the greenhouse vegetable soils and improving soil quality, but also decreases the substrate concentration for N2O production via dissimilatory NO3− reduction[15]. Both AOA amoA and AOB amoA were significantly positively correlated with N2O emission, suggesting that nitrification contributes to N2O production in greenhouse vegetable soils.
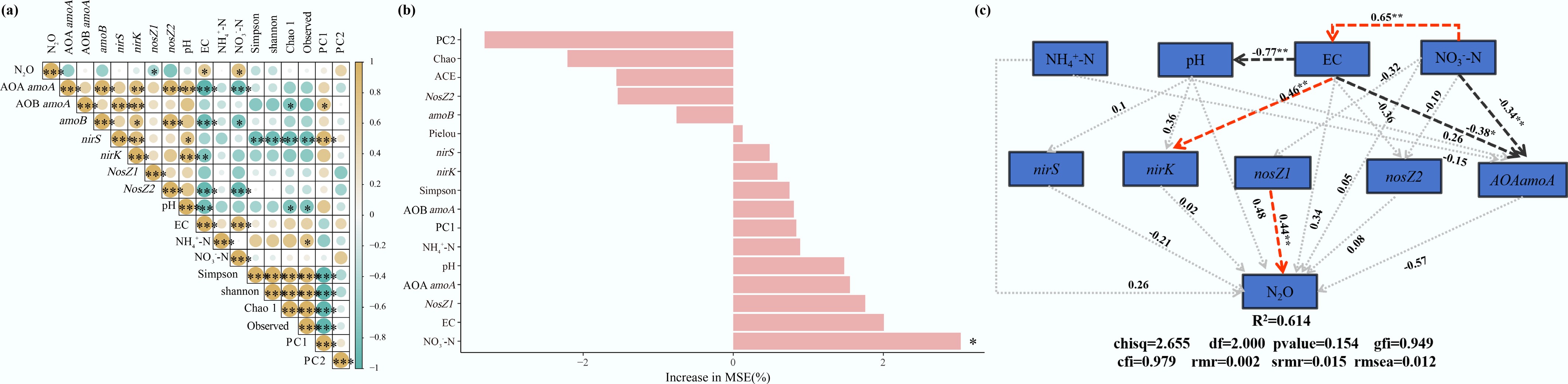
Figure 5.
(a) Correlation analysis, (b) random forest model, and (c) structural equation model analyses of environmental factors and N2O emissions from greenhouse vegetable soils. The red arrows and black arrows show positive and negative relationships, respectively. The numbers above the lines are standardized path coefficients. The width of the arrows is proportional to the strength of the path coefficients. * Significance at the p < 0.05 level, ** significance at the p < 0.01 level, and *** significance at the p < 0.001 level.
The importance ranking result of the random forest model reflects the contribution of each variable to the increase in percentage of the model's mean squared error (MSE), thereby indicating the variable importance (Fig. 5b). Among them, NO3−-N has the highest contribution to the increase in MSE and is a core factor affecting soil N2O emissions; the importance of subsequent variables such as nosZI and amoB decreases in turn. EC and NO3−-N are indirect drivers, as they influence N2O emissions by regulating the abundance of N cycling genes[29]. The structural equation model meets high reliability standards with satisfactory fitting performance, as evidenced by a reasonable degree of freedom, strong explanatory power of the Comparative Fit Index and Goodness-of-Fit Index, and minimal model-data deviation given that the Root Mean Square Error of Approximation and Standardized Root Mean Square Residual are well below the critical thresholds (df = 2.000, CFI = 0.97, GFI = 0.949, RMSEA = 0.012, SRMR = 0.015). The structural equation model further quantified the direct and indirect effects of each factor on N2O emissions: the nosZI gene exhibited a significant negative direct effect on N2O emissions (−0.44) (Fig. 5c). EC might indirectly promote N2O emissions by influencing the abundance of the nirS gene. Overall, nosZI is the key factor governing N2O emissions from greenhouse vegetable soils.
Synergistic effects of organic fertilizers and inoculation with NRCB010 on N2O emissions
-
The results showed significant differences in the N2O-mitigating effects among the combinations of different organic fertilizers and the strain. Among them, the MR-inoculated treatment (MN) exhibited the greatest N2O emission reduction, which was significantly higher than those of the CT-inoculated treatment and the HC10-inoculated treatment. This result reveals the key regulatory role of organic fertilizer type in the strain NRCB010 mitigating function. Studies have tested metabolic extracts from soil amendments and a mixture of four plant growth-promoting Bacillus strains, combined with different nitrogen fertilizers. The results showed that the microbial inoculants exhibited notable N2O-mitigating effects, which were dependent on the type of fertilizer[30]. Moreover, these inoculants could increase crop nitrogen uptake and promote crop growth.
From the perspective of the inherent characteristics of organic fertilizers, the physicochemical properties of MR provide a unique and suitable environment for the colonization and functional performance of strain NRCB010. On one hand, the pH of MR is close to neutral, while strain NRCB010 prefers a neutral to slightly alkaline environment. Neutral pH conditions can effectively alleviate acid-base stress on the strain, significantly improving its survival and reproductive efficiency in soil and laying a foundation for the strain to exert its N2O-reducing function continuously. On the other hand, MR has the highest organic matter content and a moderate C/N ratio. Compared with CT and HC10, MR can not only provide sufficient and easily utilizable carbon sources for the strain to meet its metabolic needs, but also reduce the accumulation of NO3−. Since NO3− is the core substrate for N2O production via denitrification, reducing the substrate directly lowers the potential for N2O generation. Moreover, MR has the unique advantage of improving soil structure. Previous studies have confirmed that microbial organic fertilizers derived from mushroom residue can enhance soil water and fertilizer retention capacity, increase bacterial activity, and even be used to remediate and improve various degraded soils[31].
In this study, the high organic matter content of MR further improved soil aeration, reducing the formation of anaerobic microenvironments. As denitrification occurs under anaerobic conditions, enhanced aeration can directly inhibit denitrifying bacterial activity in soil[32], thereby reducing N2O production through microenvironmental regulation. The mushroom residue organic manure used in the MN treatment could provide exclusive and highly bioavailable carbon sources for NRCB010. MR is rich in complex organic carbon compounds, including lignocellulosic derivatives, small-molecule organic acids, and amino acids, which are well matched to the metabolic preferences of Pseudomonas stutzeri. Compared with simple sugars, these carbon sources can more efficiently stimulate the metabolic activity of the strain[33]. They not only meet the energy requirements for its growth and reproduction, but also upregulate the abundance of nosZI, thereby enhancing their N2O-mitigating efficiency. In contrast, the labile organic carbon in the high-temperature pretreated compost has undergone thermal decomposition, and the carbon structure of the hydrothermal carbon organic fertilizer is recalcitrant to degradation; neither can provide such a specific carbon source. The high organic matter content of mushroom residue organic manure can promote the formation of soil water-stable macroaggregates. The aerobic outer layer of these aggregates can meet the aerobic metabolism and colonization requirements of NRCB010, while the moderately anaerobic inner microzones provide favorable conditions for denitrification[34].
Geng et al. reported that the application of bio-organic fertilizer effectively decreased N2O emissions from vegetable field soil, with a mitigation efficiency of 34%[18]. In contrast, the maximum N2O mitigating efficiency of NRCB010 tested in this study reached 46.5%, demonstrating a superior mitigating effect. The synergistic expression of the nosZ gene carried by strain NRCB010 and the soil-borne nosZI carrying microorganisms is the key molecular mechanism underlying the efficient N2O mitigation in the MN treatment. The N2O reductase encoded by the nosZI gene is the key enzyme for converting N2O into harmless N2, and its abundance shows a significant negative correlation with soil N2O emissions[35]. In this study, the abundance of the nosZI in the MN treatment was significantly higher than that in other treatments. The appropriate environment provided by MR not only promoted the colonization of NRCB010, but also enhanced the activity of native nosZI-carrying microorganisms, thereby enhancing the N2O reduction capacity and ultimately achieving efficient mitigation[36]. In contrast, the CN and HN treatments showed weaker N2O mitigating effects. Although the pH of the CN treatment decreased slightly after inoculation, it may still fall outside the suitable growth range of some strains, limiting their activity. Meanwhile, the low C/N ratio of CT readily leads to rapid accumulation of soil inorganic nitrogen, providing sufficient substrates for denitrification and offsetting part of the N2O mitigating effect. The high NO3−-N content of HC10 further increases the risk of N2O production, thereby limiting the N2O-mitigating impact of the HN treatment.
Overall, the combination of mushroom residue organic manure and NRCB010 not only achieves the optimal N2O mitigation effect but also offers multiple advantages, including improving soil quality, alleviating soil secondary salinization caused by NO3−, and increasing the utilization rate of agricultural wastes. However, limited by the simplified microcosm experimental system, the short experimental term, and the failure to account for field heterogeneity, there are restrictions on the extrapolation of the results. In the future, it is necessary to verify technical stability through long-term field experiments, analyze the mechanisms of strain-soil-crop interactions, optimize application forms, and expand research on adaptability across different environments. This will promote the application of this mitigating technology in actual production and provide more robust technical support for field use.
Underlying mechanism of microbial communities and NRCB010 in regulating N2O emission
-
The nosZI gene encodes N2O reductase and is mainly found in dominant bacterial phyla, such as Pseudomonadota[37]. The enzyme encoded by nosZI has a higher affinity for N2O and greater N2O-reducing efficiency, thereby playing a key role in N2O consumption. This also provides a theoretical basis for improving the abundance of the nosZI gene through genetic engineering methods in the future. The nirS gene encodes nitrite reductase, a key rate-limiting enzyme in denitrification. It reduces NO2− to NO, and NO is the direct precursor for the subsequent reduction to N2O[38]. A high abundance of the nirS means more active denitrification, increased NO production, and thus higher N2O generation. Both EC and NO3− content were positively correlated with the abundance of the nirS[39], suggesting that controlling water and fertilizer input may be an effective measure to inhibit the activity of the nirS gene and decrease N2O emissions[40].
NRCB010 was the primary driver of changes in soil microbial community composition, with its influence far greater than that of organic fertilizers. The colonization and reproduction of the strain directly altered the composition of the dominant bacterial phyla. NRCB010 belongs to the class Gammaproteobacteria of the phylum Pseudomonadota; the relative abundance of this phylum increased significantly after the inoculation with NRCB010. Most bacteria in Pseudomonadota are aerobic, and their widespread distribution can improve soil aeration[41], and inhibit the growth of anaerobic denitrifying bacteria (e.g., Verrucomicrobiota). The proportion of Verrucomicrobiota in non-inoculated treatments was about 22%–25%, which decreased to 10%–15% after inoculation. The decline of Verrucomicrobiota may further reduce denitrification activity and, in turn, decrease N2O emissions.
NRCB010 indirectly regulated community structure by altering the soil microenvironment through its metabolic activities. Soil EC decreased from 6.18 to 4.45 mS·cm−1, and pH increased from 6.97 to 7.02 after inoculation; optimizing soil physicochemical properties can promote the growth of beneficial microorganisms. Inoculation with NRCB010 not only achieves short-term N2O mitigation, but also improves soil ecological functions by optimizing community structure[15], providing support for the long-term sustainable use of greenhouse vegetable fields.
-
Inoculation with Pseudomonas stutzeri NRCB010 significantly decreased the cumulative N2O emissions from the greenhouse vegetable field. The N2O emissions were negatively correlated with the abundance of the nosZI gene (p < 0.05), suggesting that nosZI-carrying microorganisms were the most critical factor governing N2O emissions from the greenhouse vegetable soil. The combination of mushroom residue organic manure with P. stutzeri NRCB010 is an effective technical approach for decreasing N2O emissions from greenhouse vegetable fields, providing practical guidance for greenhouse gas mitigation and soil ecological restoration.
We sincerely thank the editor and reviewers for their insightful comments and suggestions.
-
It accompanies this paper at: https://doi.org/10.48130/nc-0026-0001.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: Wenjun Xie: writing − original draft, investigation, data curation; Zijian Qiu: validation, software, methodology, formal analysis; Dandan Li: validation, software, methodology, formal analysis; Zhouzhang Wang: Writing − review and editing; Nan Gao: validation, methodology; Ruonan Xiong: writing − review and editing; Adharsh Rajasekar: writing − review and editing; Xinhua He: writing − review and editing; Weishou Shen: writing − review and editing, validation, supervision, resources, project administration, funding acquisition, conceptualization. All authors provided revision comments on previous versions of the manuscript, reviewed the study results, and approved the final version of the manuscript.
-
The datasets used or analyzed during the current study are available from the corresponding author upon reasonable request.
-
This study was funded by the National Key Research and Development Program of China (Grant No. 2023YFF0805403), and the National Natural Science Foundation of China (Grant Nos 42377311 and 41771291).
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
Full list of author information is available at the end of the article.
- The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xie W, Qiu Z, Li D, Wang Z, Gao N, et al. 2026. Synergistic effects of Pseudomonas stutzeri NRCB010 and organic fertilizers on soil N2O emission reduction and microbial community structure in greenhouse vegetable fields. Nitrogen Cycling 2: e014 doi: 10.48130/nc-0026-0001 

Synergistic effects of Pseudomonas stutzeri NRCB010 and organic fertilizers on soil N2O emission reduction and microbial community structure in greenhouse vegetable fields
- Received: 31 October 2025
- Revised: 16 December 2025
- Accepted: 31 December 2025
- Published online: 04 February 2026
Abstract: Agricultural soils, particularly intensively managed greenhouse vegetable fields, are a significant source of the potent greenhouse gas nitrous oxide (N2O). While organic fertilization is essential for soil health, it can exacerbate N2O emissions. This microcosm study evaluated the efficacy of the denitrifying bacterium Pseudomonas stutzeri NRCB010, applied with three organic fertilizers (mushroom residue organic manure, high-temperature pretreated compost, and hydrochar organic manure), to mitigate N2O emissions and alter the microbial community. The results showed that inoculation with strain NRCB010 significantly decreased cumulative soil N2O emissions (p < 0.05). Among them, the mushroom residue organic manure inoculation treatment achieved the most significant emission reduction, with a mitigating rate of 46.5%, followed by the hydrothermal carbon organic manure inoculation treatment (27.8%) and the high-temperature pretreated compost inoculation treatment (26.3%). Soil N2O emissions showed a significant negative correlation with the abundance of the nosZI gene (p < 0.05), a key factor in decreasing N2O emissions, and a significant positive correlation with nitrate nitrogen content, pH, and electrical conductivity (EC). Furthermore, amplicon sequencing showed that inoculation significantly shifted the soil microbiome, increasing Pseudomonadota and decreasing Verrucomicrobiota, with the bacterial inoculant having a greater impact on microbial community structure than the type of organic fertilizer. It was concluded that the synergistic application of MR and P. stutzeri NRCB010 presents a robust, microbiome-mediated strategy for achieving significant N2O mitigation and ecological sustainability in protected agriculture.