









-
Citrus bacterial canker (CBC) is a plant bacterial disease caused by Xanthomonas citri (Xcc) subsp. citri, resulting in substantial losses to the global citrus industry[1]. CBC can spread through various pathways, making it difficult to prevent and control. The current disease control methods are ineffective; however, producing citrus canker-resistant citrus varieties through molecular breeding techniques can effectively control various diseases[2].
Plant lectins are a highly diverse group of carbohydrate-binding proteins that play roles in various biological processes. Based on phylogenetic analysis and domain structure, lectins are classified into 12 families, each comprising members derived from a single carbohydrate-binding-related domain[3]. The bulb-type mannose-specific binding lectin (B-lectins) belong to one of these families. These lectins can recognize and bind to mannose, modulating plant defense[4]. Mannose-binding lectins (MBLs) have been associated with plant growth. For instance, the G-type receptor-like kinase AsNIP43 interacts with the rhizobial effector nodulation outer protein P, which is essential for rhizobia-legume symbiosis[5]. Moreover, pollen development requires L-type lectin receptor kinase IV.2 (LECRK-IV.2)[6]. In Brassica, the S-locus receptor kinase containing a B-lectin domain modulates self-incompatibility responses[7].
The literature has validated that MBL plays a defensive role in many plant species. Furthermore, it is associated with salt stress in rice[8,9]. OsJAC1 overexpression increases Arabidopsis's resistance to ionizing radiation[10]. Moreover, in Arabidopsis, the MYB4-MAN3-mannose-MNB1 signaling cascade regulates cadmium tolerance via the GSH-dependent PC synthesis pathway[11]. MBLs are widespread in higher plants, and can interact with insect gastrointestinal systems, providing immunity against pests[12]. For instance, in transgenic crops, expressing snowdrop lectin GNA or feeding GNA provides resistance to lepidopteran pests[13,14]. In rice, GNA lectin gene overexpression increases resistance to the white-backed planthopper[15]. The upregulation of G-type lectin genes increases aphid resistance by inhibiting growth and fertility[16]. MBLs protect plants against pathogen attack by recognizing specific carbohydrates on pathogen surfaces and acting as a plant defense signaling mechanism. In rice, OsCERK1 plays a key role in lipopolysaccharide (LPS)-induced immune responses[17]. For example, the B-lectin domain-containing gene Pi-d2 confers resistance to rice blast[18]. Arabidopsis G-type lectin LORE, pepper G-type lectin genes CaMBL1 and CaGLP1 are involved in defense against Xanthomonas campestris pv. vesicatoria (Xcv)[19−21]. Moreover, in sunflowers, the jacalin-type MBL Helja interacts with fungal spore surfaces, inducing oxidative stress, and leading to cell death[22]. FaMBL1, an MBL gene, provides resistance to immature strawberry fruits against Colletotrichum acutatum[23].
Plants have various defense mechanisms, such as a complex immune system, to protect themselves against different pathogens[24]. Furthermore, several signaling molecules, such as reactive oxygen species (ROS), ethylene (ET), salicylic acid (SA), and jasmonic acid (JA), have been associated with plant defense networks[25]. The literature suggests that excessive ROS increases resistance against Xcc[26,27]. Moreover, JA and SA are key phytohormones that modulate plant immunity[28,29]. Further, plants confer resistance to citrus canker by regulating endogenous hormones such as JA and SA via multiple pathways[30]. Moreover, abscisic acid (ABA) has been found to negatively regulate resistance against pathogens, such as against Botrytis cinerea and Pseudomonas syringae in tomatoes[31,32]. Similarly, ET has a positive impact on plant disease resistance[33].
Various studies have investigated the association between MBL and biotic stress; however, its relationship with citrus canker remains unclear. Based on transcriptomic data from earlier infection experiments, using Xcc on citrus germplasms with contrasting resistance phenotypes, consistently higher expression of CsMBL1 in the citrus canker-resistant variety Kumquat (Citrus japonica) was observed compared to the citrus canker-susceptible variety Wanjincheng (Citrus sinensis), across multiple time points. This expression pattern suggested a potential role of CsMBL1 in CBC resistance, which became the focus of this study. To functionally characterize CsMBL1, genetic transformation and virus-induced gene silencing (VIGS) were performed in citrus. Integrated with biochemical assays, the present results reveal a mechanism by which CsMBL1 contributes to CBC resistance. It was observed that CsMBL1 contributes to the development of novel citrus canker-resistant citrus germplasm.
-
The Kumquat (C. japonica) and Wanjincheng (C. sinensis) citrus varieties were obtained from the National Citrus Germplasm Repository (Chongqing, China) and grown in a greenhouse at 28 °C for 3−5 months. Wanjincheng plants have greater susceptibility to citrus canker than Kumquat plants[34]. The Xcc variant, XccYN1, was isolated from infected citrus leaves and cultured at 28 °C in a peptone-yeast/malt extract medium augmented with 1.5% (w/v) D-glucose.
Bioinformatics analysis of CsMBL1
-
CsMBL1 coding sequence (CDS) was cloned and sequenced (Supplementary Table S1). This study acquired sequences from the CPBD database (
http://citrus.hzau.edu.cn )[35,36]. Gene Structure Display Server (GSDS V2.0,http://gsds.gao-lab.org ) was employed to predict gene structure[37]. Furthermore, functional domains were predicted using HMMER (http://hmmer.org )[38]. Moreover, MEGA X was utilized to construct a phylogenetic tree[39] using MBLs from different organisms (Supplementary Table S2).Exogenous hormone-induced expression in citrus leaves
-
To conduct exogenous hormone assays, leaf discs (with a diameter of 7 mm) were immersed in different hormones (10 μmol/L ABA, 100 μmol/L methyl jasmonate [MeJA], 10 μmol/L SA, 10 μmol/L ET), and then sampled at 0, 6, 12, and 24 h post-treatment (hpt).
Overexpression in citrus
-
The complete CDS of CsMBL1 was amplified using the following primers: FOEC-CsMBL1 (containing KpnI restriction site: GGTACCATGAAGATGTCATCTTCCTCAGC) and ROEC-CsMBL1 (containing 3 × flag tag and EcoRI restriction site sequence: GAATTCTTACTTATCATCATCATCCTTGTAATCCTTATCATCATCATCCTTGTAATCCTTATCATCATCATCCTTGTAATCCTTGTTGGGAGCCTTTATGA). The amplified sequence was then inserted into the pGLNe vector comprising the CaMV 35S promoter, which was then transformed into Agrobacterium tumefaciens EHA105. Furthermore, the segments of Wanjincheng's shoot were transformed according to the method of Peng[2]. Transgenic plants were confirmed by PCR and GUS staining, and thereafter assessed for CsMBL1 expression levels with qRT-PCR. Plants with empty vectors were set as controls.
Subcellular localization
-
The recombinant plasmid CsMBL1-EAQGFP was constructed based on the stop codon lacking CDSs of CsMBL1 using the following primer pair: FSC-CsMBL1 (containing homologous arms and an AgeI restriction site: CTGCCCAAATTCGCGACCGGTATGAAGATGTCATCTTCCTCAG), and RSC-CsMBL1 (containing homologous arms and an AgeI restriction site: TCCTTTGCTAGTCATACCGGTCTTGTTGGGAGCCTTTAT). Then, the fragments were cloned into the EAQGFP vector by following the protocol of the In-Fusion cloning kit (TaKaRa, Japan). Subsequently, the constructed plasmids were transfected into Nicotiana benthamiana protoplasts. After 12 h, the GFP signals were measured using a laser scanning confocal microscope (Olympus FV3000, Japan).
Virus-induced gene silencing (VIGS)
-
For VIGS fragment (Supplementary Table S1) amplification, the following primer pair was employed: FVIGS-CsMBL1 (containing homologous arms and an EcoRI restriction site: TTCTGTGAGTAAGGTTACCGAATTCGGTAAATTTATTTGGCAGAGTTTTGATTATCCA)/RVIGS-CsMBL1 (containing homologous arms and a BamHI restriction site: GACGCGTGAGCTCGGTACCGGATCCCGTGGGCCCCCAATCAA). The amplified region was incorporated into the TRV2 vector to produce TRV2-CsMBL1, and VIGS transformation was executed through infiltration with Agrobacterium tumefaciens EHA105[40]. After 30 d, Wanjincheng samples displayed green fluorescence under UV light, were pulverized in liquid nitrogen, and subsequently utilized for further analysis. The efficacy of VIGS silencing was assessed using qRT-PCR.
Citrus canker resistance analysis
-
Mature CsMBL1 overexpressing leaves and VIGS plants were punctured 24 times (6 punctures/group). Then, each puncture site was injected with 1 µL of XccYN1 (1 × 108 CFU·mL−1), and leaves were placed on wet cotton at 28 °C for 10 d. The citrus canker disease on the leaves was assessed by evaluating the disease index (DI), and lesion area[2].
qRT-PCR analysis
-
Total RNA was extracted using the RNA Mini Preparation Kit (Aidlab, China), and reverse transcribed into cDNA using the PrimeScript Kit (Takara, Japan), and a mixture of 1 µg RNA, 2 µL PrimeScript buffer, and RNase-free water (to a total volume of 10 µL). The reaction conditions were set as follows: 37 °C for 45 min, 85 °C for 5 s, and 4 °C for 5 min. The qRT-PCR thermal cycling program was as follows: pre-denaturation at 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s, and 56 °C for 30 s. The reaction system (12 µL) contained cDNA (100 ng), primers (0.3 mmol/L), and PCR premix (6 µL). The relative gene expression was assessed via the 2−ΔΔCᴛ method[41]. The primers employed for qRT-PCR analysis were designed using the NCBI Primer BLAST tool and included: FRT-CsMBL1 (GAGAGTTCGGGCCATACGTT) and RRT-CsMBL1 (AACGCAAGCTGGAATGGAGA). The primers for reference gene, CsGAPDH (CPBD ID: Cs5g06870)[42,43], were FRT-GAPDH (GCTTTCCGTGTACCCACTGT) and RRT-GAPDH (CTCTGACTCCGCCTTGATGG). The C. sinensis mRNA sequence was used as a standard for primer specificity evaluation.
Biochemical indices
-
Hydrogen peroxide (H2O2) and superoxide radicals (O2−) concentrations were measured using commercial kits (Suzhou Keming Biotechnology, China). Furthermore, the expression levels of JA, SA, and ABA were determined via UPLC-MS/MS, while ET concentration was measured using gas chromatography (GC). Peroxidase (POD), ascorbate peroxidase (APX), and respiratory burst oxidase homologs (RBOHs) activities were assessed using commercial kits (SinoBestBio, Shanghai, China).
Statistical analysis
-
Data were analyzed and visualized via GraphPad Prism V9.5 (San Diego, CA, USA), and depicted as means ± standard deviations (SDs). For intergroup comparison, a two-tailed t-test or Duncan's multiple range test followed by ANOVA was performed. p < 0.05 was considered statistically significant. qRT-PCR was performed in triplicate (n = 3).
-
CsMBL1 is present on chromosome 5 of sweet orange, and encodes a protein comprising 436 amino acids, with Bulb-type lectin, S-locus glycoprotein, and PAN domains. The CsMBL1 gene is 1,308 bp long and comprises a single exon (Fig. 1a). Phylogenetic analysis revealed that CsMBL1 is significantly associated with Poncirus trifoliata Ptrif.0003s1398.1.p. (Fig. 1b). The subcellular localization analysis revealed that CsMBL1 is primarily located in the nucleus (Fig. 1c).

Figure 1.
Expression analysis and bioinformatics characterization of CsMBL1. (a) Chromosomal localization of CsMBL1 was identified using CPDB (internal reference), and its exon-intron structure was determined using GSDS V2.0. CsMBL1's functional domains were identified using HMMER. (b) MEGA X (bootstrap: 500, Poisson model) was employed to construct a maximum likelihood phylogenetic tree of CsMBL1 and its homologous proteins. Tree branches were proportional, with lengths representing the number of substitutions per site. Source species and protein IDs are indicated on the right side of the phylogenetic tree. (c) CsMBL1 had a subcellular localization in Nicotiana benthamiana protoplasts. Images show GFP (green fluorescent protein), mCherry (red fluorescent protein), bright field, and merged channels. Scale bar = 10 μm.
Hormone treatments regulate CsMBL1 expression in plants
-
In plants, phytohormones generally modulate the expression of disease-related proteins[44]. To investigate the role of CsMBL1 in disease resistance signaling pathways, plants were treated with ABA, MeJA, SA, and ET, and then subjected to RT-PCR analysis to investigate CsMBL1 expression. After exogenous ABA treatment, CsMBL1 expression in both cultivars showed a consistent pattern of initial increase and then decrease (Fig. 2a). However, exogenous MeJA treatment induced contrasting CsMBL1 expression patterns, exhibiting considerable downregulation in Kumquat and pronounced overexpression in Wanjincheng (Fig. 2b). In Wanjincheng, SA treatment initially significantly decreased CsMBL1 expression, then increased, followed by a sharp decline at 24 h. Compared with Wanjincheng, CsMBL1 expression indicated a more gradual decline in Kumquat (Fig. 2c). Moreover, ET induction resulted in distinct temporal expression patterns of CsMBL1 between cultivars. In the Wanjincheng variety, CsMBL1 expression was significantly upregulated within the first 12 h post-ET-treatment, followed by a sharp decline by 24 h, whereas in the Kumquat variety, CsMBL1 expression was consistently elevated with no significant difference between 12 and 24 h timepoints (Fig. 2d). These results showed that CsMBL1 expression induces phytohormones, suggesting its functional involvement in certain hormone-mediated disease resistance signaling pathways.
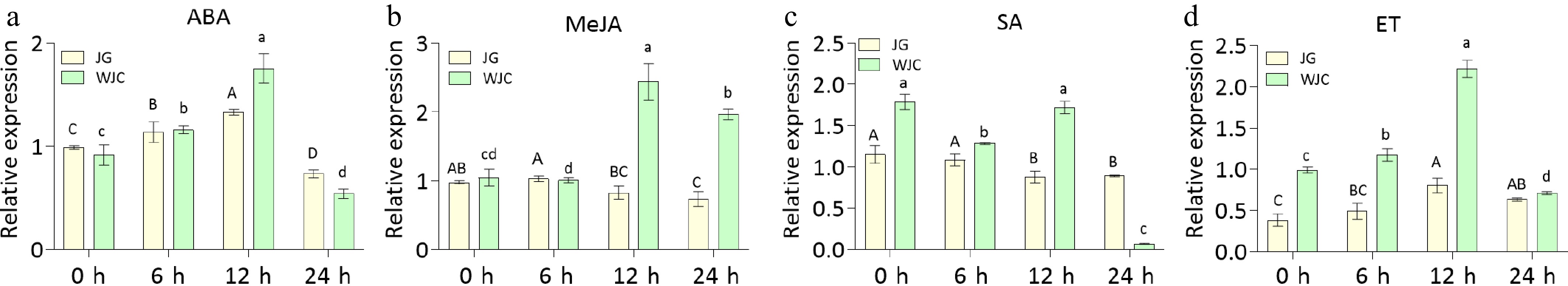
Figure 2.
Effects of phytohormone treatments on CsMBL1 expression in plants. qRT-PCR analysis of CsMBL1 expression in kumquat and Wanjincheng leaves following treatments with (a) ABA, (b) MeJA, (c) SA, (d) ET, and H2O (control). CsGAPDH (CPDB ID: Cs_ont_5g044290) was used as the internal reference gene. Statistical comparisons were performed using ANOVA with Duncan's multiple range test. WJC: Wanjincheng; JG: Kumquat.
CsMBL1 overexpression confers resistance against citrus canker
-
In this study, a CsMBL1 overexpression plasmid was designed to assess its effects (Fig. 3a). The GUS staining indicated blue color on the leaf disc edge in three transgenic plants (PLGNe-CsMBL1-1, -2, -3), while no staining was observed in control plants (PLGNe) (Fig. 3b). PCR analysis detected the target fragment in the transgenic and positive control samples, but not in the control samples (Fig. 3c). Furthermore, qRT-PCR analysis validated CsMBL1 overexpression in transgenic plants (Fig. 3d). Moreover, transgenic plants had normal growth rates (Fig. 3e), and reduced citrus canker symptoms compared to controls (Fig. 3f). Similarly, the lesion disease and area index were reduced by 11.5%−18.8% and 10.6%−15.4%, respectively (Fig. 3g and h). These data revealed that CsMBL1 overexpression increases citrus canker resistance.
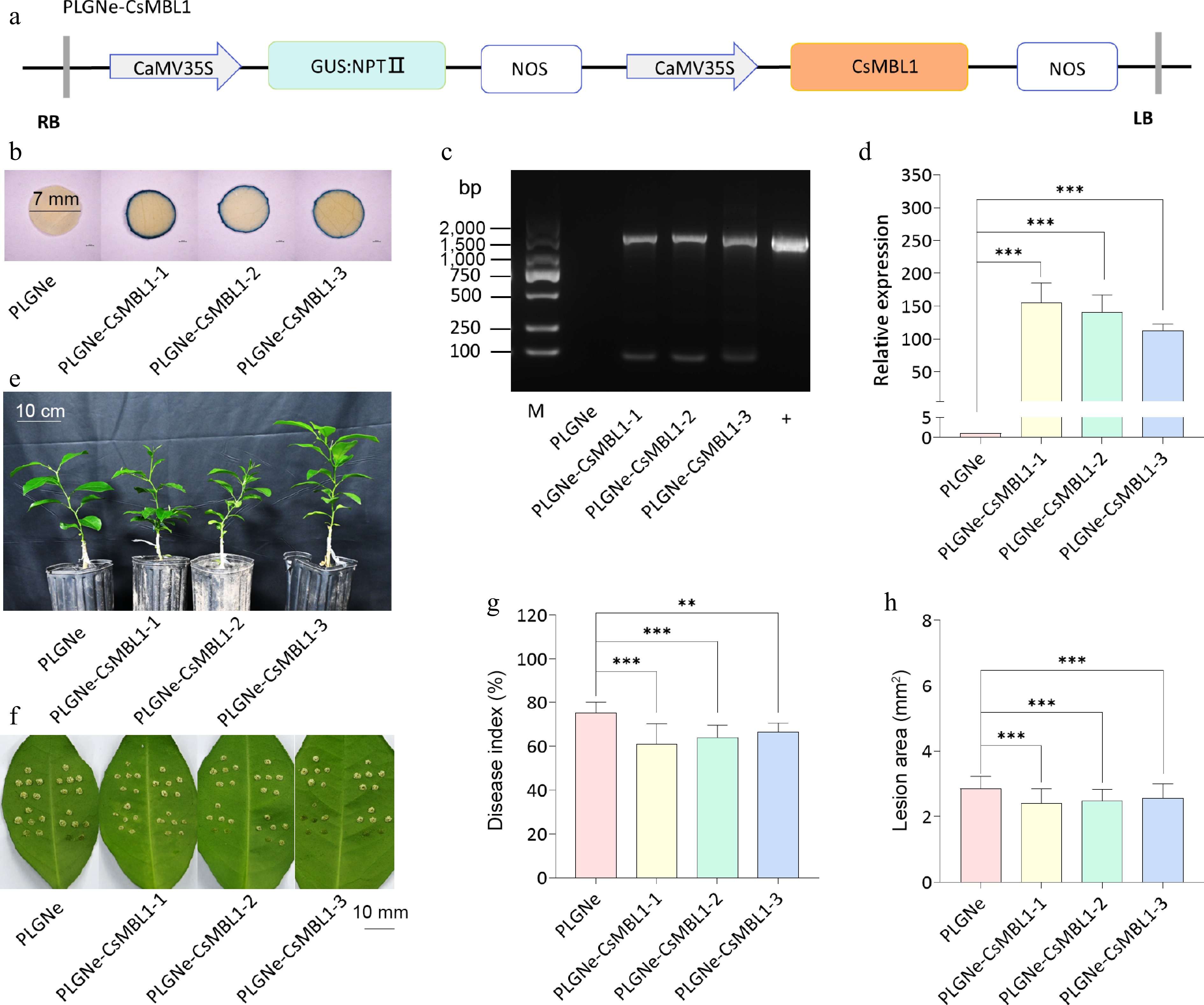
Figure 3.
Effects of CsMBL1 overexpression on citrus canker resistance. (a) CsMBL1 overexpression vector. (b) GUS staining results. Blue staining indicates transgenic plants. (c) PCR validation of transgenic plants. M: DNA marker; +: pLGNe-CsMBL1 plasmid. (d) The expression of CsMBL1 relative to CsGAPDH. The control value was set to 1. Data are presented as mean ± SD of three replicates. (e) Phenotype of transgenic plants. (f) citrus canker symptoms in Xcc-infected leaves at 10 dpi. (g) Lesion disease, and (h) area index in CsMBL1-overexpressing plants at 10 dpi. (b)−(h) pLGNe-CsMBL1-1, -2, -3: CsMBL1-overexpressing materials, while pLGNe: control plants carrying the pLGNe vector. Data was statistically analyzed using Tukey's multiple range test (p < 0.05). NPTII: neomycin phosphotransferase II, NOS: NOS terminator, 35S: CaMV 35S promoter, GUS: β-glucuronidase and npt-linked coding gene, LB: left border, RB: right border, dpi: days post-infection.
VIGS of CsMBL1 increases susceptibility to citrus canker
-
To evaluate the role of CsMBL1 in citrus canker resistance, CsMBL1 was silenced in Wanjincheng plants. A CsMBL1 VIGS plasmid was constructed using the TRV2 vector (Fig. 4a). Furthermore, CsMBL1 silencing in VIGS plants was validated by PCR analysis using TRV1 and TRV2-specific primers (Fig. 4b). Compared to controls, VIGS plants had reduced relative expression of CsMBL1 (Fig. 4c). Moreover, Xcc-infected VIGS plants had more severe citrus canker symptoms relative to control plants (Fig. 4d). Further, the calculated lesion areas (1.25, 1.18, and 1.32) (Fig. 4e) and disease index values (1.23, 1.17, and 1.33) of plants TRV2-CsMBL1-1, TRV2-CsMBL1-2, and TRV2-CsMBL1-3 were higher than that of controls (Fig. 4f). These results indicate that silencing CsMBL1 reduces resistance to citrus canker.
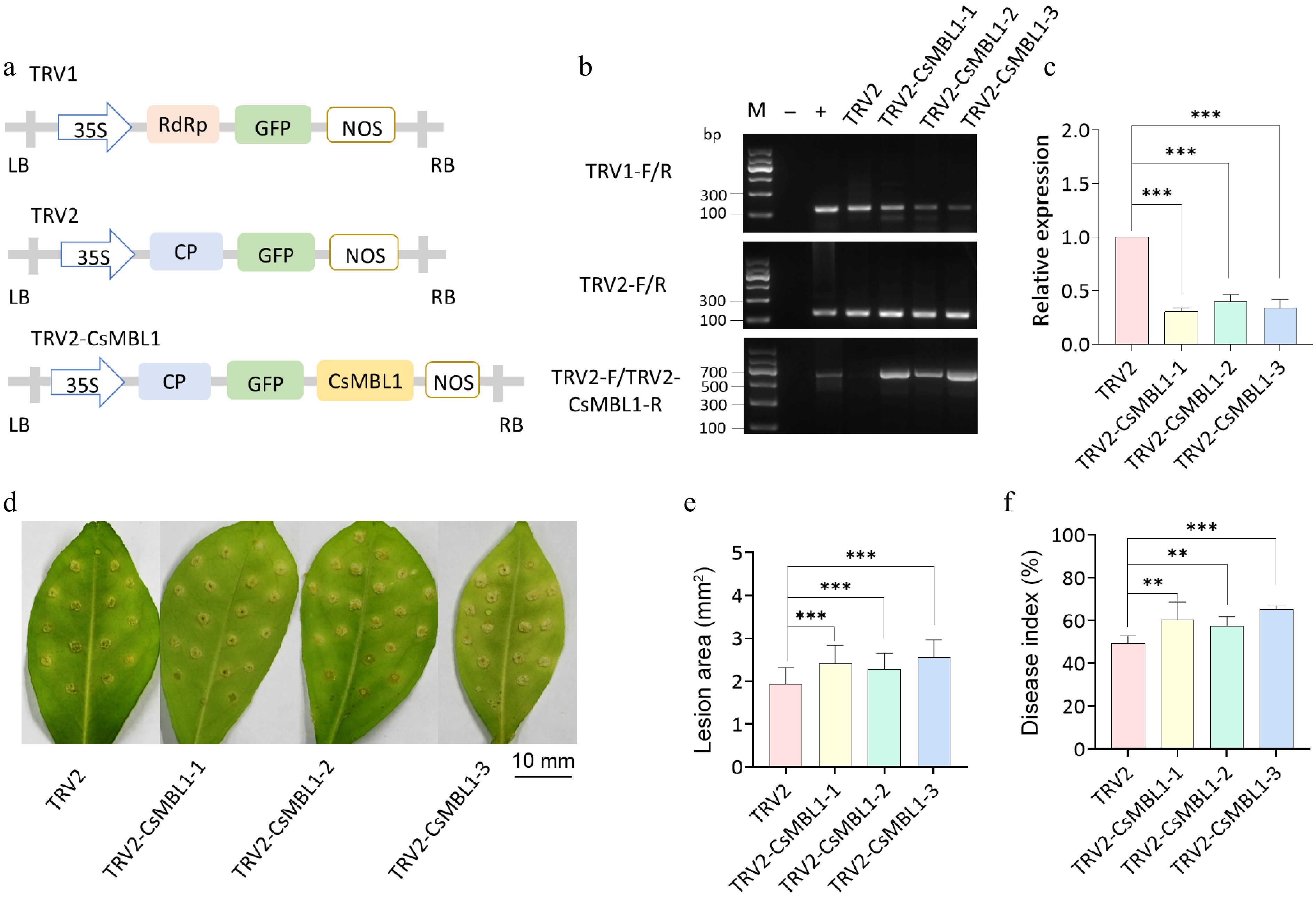
Figure 4.
Effect of CsMBL1 silencing on citrus canker resistance. (a) Plasmid used for VIGS. (b) Successful transformation of VIGS plants was confirmed by PCR. M indicates DNA Marker, "–" represents ddH2O, and "+" depicts plasmid. Primers used are shown on the left side of the gel electrophoresis image. (c) Relative CsMBL1 expression was assessed by qRT-PCR using CsGAPDH as a control. The control value was set to 1, and the data represent the average of three biological replicates. (d) Disease symptoms, (e) lesion area, and (f) disease index of Xcc-infected VIGS plants at 10 dpi. (b)–(f) TRV2-CsMBL1-1, -2, and -3: CsMBL1 VIGS plants; TRV2: control plants carrying the TRV2 vector (ANOVA with Tukey's multiple range test, p < 0.05). VIGS: virus-induced gene silencing (VIGS), 35S: Cauliflower mosaic virus 35S promoter, NOS: NOS terminator, GFP: green fluorescent protein, RdRp: RNA-dependent DNA polymerase, CP: coat protein, LB: left border, RB: right border.
CsMBL1 overexpression modulates ROS homeostasis in plants
-
In plants, ROS are the first line of defense against pathogens[45]. Furthermore, H2O2 and O2•− serve as signaling molecules that regulate diverse cellular processes[46,47]. Here, it was observed that both H2O2 (Fig. 5a), and O2− (Fig. 5b) levels were increased in CsMBL1-overexpressing plants. RBOHs are primary producers of ROS in plants[48]. Moreover, POD and APX participate in ROS scavenging[49,50]. This study examined the activities of POD, RBOHs and APX, and revealed significant downregulation of POD activity (Fig. 5c), an evident increase in RBOH activity (Fig. 5d), while no significant difference in APX activity was observed compared to controls (Fig. 5e). These findings revealed that CsMBL1 overexpression increases ROS levels through the coordinated regulation of POD and RBOH activities, therefore increasing resistance to Xcc.
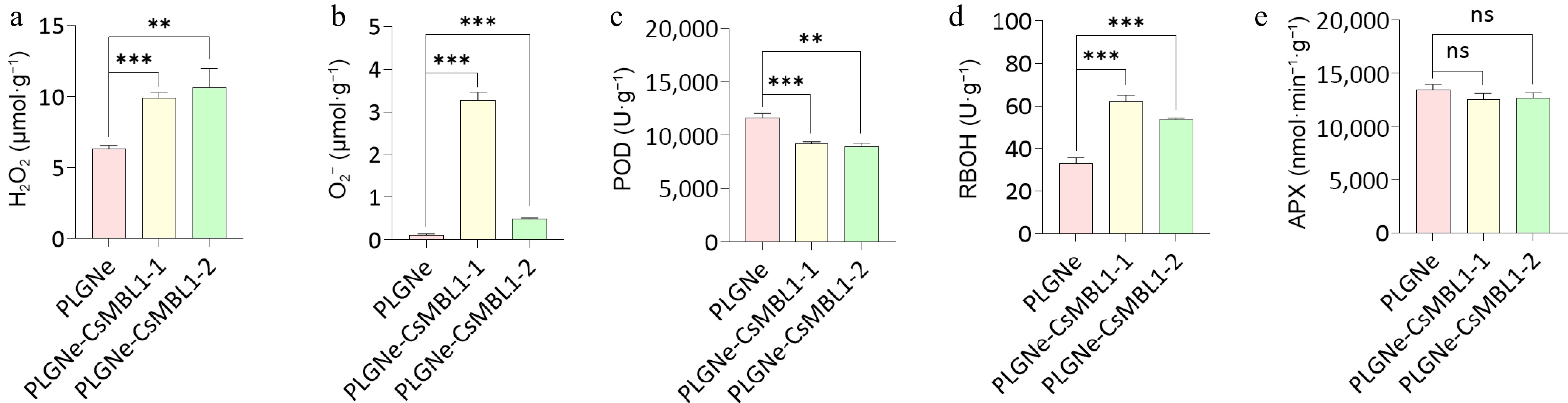
Figure 5.
Effects of CsMBL1 overexpression on plant ROS levels. The contents of (a) H2O2, (b) O2−, and activities of (c) POD, (d) RBOH, and (e) APX were measured in CsMBL1-overexpressing plants. PLGNe-CsMBL1-1/-2: CsMBL1-overexpressing transgenic lines, PLGNe: Control plants carrying the empty PLGNe vector. Data were analyzed by ANOVA with Tukey's multiple range test, p < 0.05.
CsMBL1 overexpression alters phytohormone levels
-
Phytohormones, including SA, JA, ABA, and ET, have been found to essentially modulate plant immunity[51−53]. To identify whether these hormones participate in CsMBL1 overexpression-mediated resistance, PLGNe-CsMBL1-2 transgenic plants with superior resistance were selected to assess phytohormone levels. It was observed that CsMBL1 overexpression significantly reduced ABA and JA levels (Fig. 6a and b), while increasing SA accumulation (Fig. 6c). There were no significant changes in ET content (Fig. 6d). These data suggest that CsMBL1 overexpression increases citrus canker resistance in Wanjincheng by modulating hormone homeostasis, decreasing JA and ABA levels while increasing SA accumulation.
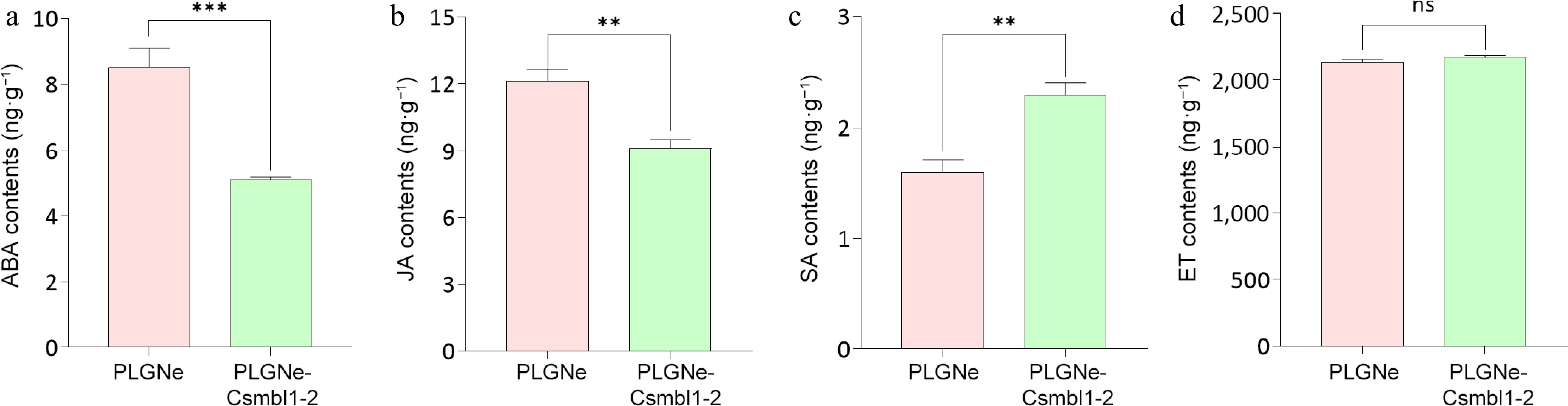
Figure 6.
Effects of CsMBL1 overexpression on phytohormone levels. Concentrations of (a) ABA, (b) JA, (c) SA, and (d) ET were measured in CsMBL1-overexpressing plants. In (a)−(d): PLGNe-CsMBL1-2: CsMBL1-overexpressing plants. PLGNe: Control plants with an empty PLGNe vector. Data were statistically analyzed by ANOVA with Tukey's multiple range test (p < 0.05).
-
The citrus canker is a destructive plant disease caused by Xcc and significantly adversely affects global citrus production[54]. Lectins are natural bioactive proteins with unique carbohydrate-binding abilities. Furthermore, they have indicated antibacterial properties and facilitate immune responses[55]. Mannose-binding B-lectins specifically recognize mannose and function in innate immunity. The anti-pathogenic activity of MBLs has been observed in various species, including Arabidopsis, pepper, orchid, rice, and strawberry[18,20,56,57]. To evaluate whether citrus MBLs confer citrus canker resistance, CsMBL1, a citrus canker-induced gene encoding a B-lectin, was analyzed. Based on the transcriptome data, we identified and functionally characterized the differentially expressed gene CsMBL1. The data showed that CsMBL1 overexpression alleviated canker symptoms, whereas VIGS exacerbated disease severity, suggesting that CsMBL1 positively regulates citrus canker resistance.
The ROS signaling plays a pivotal role in plant innate immunity[58], and its accumulation promotes a plant adaptive response to stress conditions[46]. In sweet orange, the CsAP2-09-CsWRKY25-CsRBOH2 cascade confers Xcc resistance by modulating ROS homeostasis[27]. In rice, OsCERK1 downregulation inhibits the formation of ROS generated by LPS, which is a principal microbe-associated molecular pattern (MAMP) recognized by both plants and animals[17]. Here, it was found that CsMBL1-overexpressing plants had elevated H2O2 and O2− levels, as well as decreased POD and enhanced RBOHs activities. This ROS accumulation constitutes the fundamental plant defense mechanism[59], demonstrating that CsMBL1 mediates citrus canker resistance via ROS production.
Phytohormones, such as ABA, SA, JA, and ET, regulate plant defense mechanisms against biotic and abiotic stressors[60]. It has been observed that SA significantly regulates hypersensitive response and systemic acquired resistance[61], while ABA antagonizes defense-related hormones[62]. Moreover, JA and ET jointly modulate Botrytis cinerea-induced systemic resistance[63]. Furthermore, the relationship between ABA, SA, JA, ET, and resistance to citrus canker is well-documented in the literature[34,54,64]. In this study, the hormonal analyses revealed that CsMBL1 positively regulates SA while negatively affecting JA and ABA levels. MBLs are considered the regulators of the SA/JA pathways. For example, the overexpression of pepper CaMBL1 stimulates the production of defense genes and the formation of SA to counteract Xanthomonas campestris pv vesicatoria (Xcv) infection[20]. Moreover, Arabidopsis AtLEC is involved in JA/ET/chitin-triggered defenses[65], and strawberry FaMBL1 overexpression decreases JA content[57].
These findings indicate that CsMBL1 acts as a modulator of ABA, JA, SA, and ROS homeostasis, thus increasing citrus canker resistance. Furthermore, CsMBL1 expression is inducible by MeJA in Wanjincheng; however, its overexpression inhibits JA levels, indicating a self-regulatory loop that may modulate JA-mediated defenses. However, the present study on CsMBL1 remains insufficiently comprehensive. The relationships between CsMBL1 and phytohormones, ROS, and other signaling molecules require further investigation. Subsequent research could identify CsMBL1-interacting proteins and examine the expression of downstream defense-related genes to refine the regulatory network. These efforts would provide additional data to support the role of CsMBL1 in resistance against citrus bacterial canker. Additionally, Xcc-induced gene expression experiments can be conducted in both the citrus canker-resistant variety Kumquat (C. japonica), and citrus canker-susceptible variety Wanjincheng (C. sinensis). This would allow for the construction of a hypothetical model that integrates cultivar-specific data and functional data to explain the enhanced resistance to CBC observed in Kumquat and in CsMBL1-overexpressing 'Wanjincheng' plants.
-
In summary, this study demonstrated that CsMBL1 increases citrus canker resistance by coordinated regulation of defense components: (1) upregulating CsMBL1 expression; (2) increasing SA and ROS (via RBOHs activation/POD suppression); and (3) reducing ABA/JA levels. Therefore, CsMBL1, as a positive regulator of citrus canker resistance, serves as a prospective molecular target for the development of canker-resistant citrus cultivars.
This study was funded by the National Key Research and Development Program of China (2022YFD1201600), National Natural Science Foundation of China (32472682, 32502634), National Natural Science Foundation of Chongqing (CSTB2025NSCQ-GPX0540), Earmarked Funds for the China Agriculture Research System (CARS-26) and Fundamental Research Funds for the Central Universities (SWU-XDJH202308).
-
The authors confirm their contributions to the paper as follows: designed the experiments: Li Q; performed the experiments: Li M, Song Q, Lin D, Zhang M; analyzed the data: Li M, Song Q, Li Q, He Y; wrote the article: Li M, Li Q, He Y. All authors reviewed the results and approved the final version of the manuscript.
-
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary materials.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Man Li, Qingwei Song
- Supplementary Table S1 Sequences used in this study.
- Supplementary Table S2 MBLs used for phylogenetic analyses.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li M, Song Q, Lin D, Zhang M, He Y, et al. 2026. The mannose-binding lectin gene CsMBL1 positively regulates resistance to citrus canker. Plant Hormones 2: e001 doi: 10.48130/ph-0025-0028 

The mannose-binding lectin gene CsMBL1 positively regulates resistance to citrus canker
- Received: 25 September 2025
- Revised: 10 November 2025
- Accepted: 17 November 2025
- Published online: 09 January 2026
Abstract: Citrus canker is a serious bacterial disease caused by Xanthomonas citri (Xcc) subsp. citri. It is a significant threat to the citrus industry. Plants have several defense proteins, including mannose-binding lectins (MBL), which modulate innate immune responses by recognizing and binding to carbohydrates on microbial surfaces. However, the role of MBL in citrus canker resistance remains elusive. Previous transcriptomic analysis revealed that in the leaves of the citrus canker-resistant variety Kumquat (Citrus japonica), and the susceptible variety WanJincheng (Citrus sinensis) treated with Xcc, higher levels of CsMBL1 were detected in the former compared to the latter, suggesting that CsMBL1 is associated with resistance to citrus canker. Therefore, this study analyzed and identified a mannose-binding B-lectin gene, CsMBL1, and obtained the following results. Subcellular localization results indicated that CsMBL1 is localized in the nucleus. Phytohormone induction experiments demonstrated that methyl jasmonate (MeJA) downregulated the expression of CsMBL1 in Kumquat, while upregulating it in Wanjincheng. Moreover, its overexpression increased citrus canker resistance, while virus-induced gene silencing (VIGS) increased susceptibility. Biochemical analyses revealed increased reactive oxygen species (ROS) content, reduced peroxidase (POD) activity, and increased respiratory burst oxidase homologs (RBOHs) activity in CsMBL1-overexpressing plants. Additionally, the levels of abscisic acid (ABA) and jasmonic acid (JA) were diminished, while the salicylic acid (SA) level was elevated. These findings suggest that CsMBL1 acts as a positive regulator of citrus canker resistance by modulating JA, ABA, SA, and ROS levels; therefore, CsMBL1 can be targeted for modulation to develop canker-resistant citrus cultivars.
-
Key words:
- Mannose-binding B-lectin /
- Citrus canker /
- Xanthomonas citri subsp. citri (Xcc) /
- Plant hormones /
- ROS