









-
Floral nectar production is a critical trait in plant growth, as it mediates interactions with pollinators, flower-visiting antagonists, and microbes through its chemical composition[1]. Nectar is produced in specialized secretory cells known as nectaries. These structures can be located on various parts of the plant but are most commonly found within the flower[2]. Despite the critical role of nectar in attracting pollinators, promoting outcrossing, and enabling plants to achieve maximum seed set, the molecular regulation of nectar secretion, particularly during the pre-secretory stage, remains poorly understood. In most plants, floral nectar is synthesized from sucrose transported via the phloem[3,4]. Depending on the plant species, a portion of the sucrose delivered by the phloem is hydrolyzed into glucose and fructose within the nectaries[5]. In some species, a portion of the glucose produced is temporarily stored as starch within the nectaries, and later converted back into sugar during nectar secretion[5]. Developmental studies of nectaries in various species have shown that starch accumulation is often a prominent feature[6,7]. The rates of nectar synthesis and secretion can be influenced by both endogenous and exogenous factors, at least partially under the regulation of phytohormones. Jasmonic acid (JA) has been demonstrated to positively regulate nectar secretion in both floral and extra floral nectaries[8]. JA levels are positively correlated with increased nectar production, reaching peak concentrations just prior to nectar secretion in Brassica napus[9].
Antagonistically, gibberellic acid (GA) regulates the volume of nectar produced by altering the availability of sugars, thereby directly controlling the osmotic potential, and the volume of the nectar secreted[10]. GA-deficient or GA-signaling mutants have nectaries that are full of starch but produce very little nectar. Application of GA can promote nectar secretion in some plants, but it is often insufficient without the initial JA signal[11,12]. The relationship between JA and GA in this process is not independent; they engage in a sophisticated hormonal crosstalk, often acting antagonistically. DELLAs promote JA (for initiation), and inhibit GA's starch-breaking function[13]. Removing DELLAs via exogenous GA switches the process from initiating secretion to maximizing nectar volume. Similar to these phytohormones, auxin does play a role in the nectar secretion network, but its function is more nuanced and appears to be primarily upstream and indirect, often acting as a master regulator of floral development that sets the stage for the more direct JA/GA crosstalk. Mutants with disrupted auxin signaling or transport often show absent or malformed nectaries. The auxin response marker DR5::GUS shows strong expression in developing nectary primordia[14]. In Arabidopsis, AUXIN RESPONSE FACTORS 6 and 8 (ARF6/8) play a key role in promoting and coordinating nectary formation[15], and Aquilegia[16]. Although these taxa exhibit differences in nectary formation, they share a common response to hormone signaling, highlighting the central role of phytohormones. Auxin further influences nectar secretion through the expression of PIN-FORMED 6 (PIN6), which is positively correlated with nectar production[17]. Furthermore, both auxin and JA are regulated by gibberellic acid (GA), underscoring the intricate interplay among auxin, JA, and GA[15]. Although it is known that auxin can influence JA biosynthesis, providing a potential link between the early patterning and the later secretory signals, does auxin mediate early crosstalk for nectar synthesis at the pre-secretory stage remain unclear? To answer this, the genetic basis of carbohydrate accumulation induced by exogenous auxin was investigated, followed by its degradation into nectary sugars, in the day flowering herbal plant Platycodon grandiflorus.
-
Platycodon grandiflorus seedlings obtained from a nursery in a potting composed of a 3:1:1 (peat : perlite : vermiculite) mixture, were grown in the open fields at National Institute of Forest Sciences (NIFOS), Biotechnological Resources, Suwon campus (1st week of May 2024). The plants were grown for nearly 50 d, until the buds were seen (temperature: daytime 25 ± 1 °C, and nighttime 18 ± 1 °C; relative humidity measured varied 60% ± 5% throughout the study period; light cycle of 16/8 h photoperiod). For the treatments, individual buds of uniform size (~1.5 cm) (pre-secretory stage) were selected and sprayed with 1 and 100 μM auxin solution (from a 1 mm stock solution made up to 5 ml and sprayed using a transparent bottle), and control (water alone). The buds were initially covered with polyethylene transparent bags for 3 h after spraying (to prevent immediate transpiration). The plastic bags were removed, and the treated buds were left for 48 h. The floral bud samples were collected at two time points (24 and 48 h after treatment), respectively. Collected samples (four replicates each) were immediately stored at 4 °C (for histochemical sectioning), and −80 °C (for RNA sequencing).
Histochemical sectioning
-
The anatomical structure of floral buds was examined microscopically as follows. At the end of the treatments (24 and 48 h), the floral buds were washed with distilled water, and fixed with an FAA fixative embedded in paraffin[18]. Briefly, samples were immediately immersed in freshly prepared FAA (Formalin-Acetic Acid-Alcohol: 10% formalin, 5% glacial acetic acid, 50% ethanol, 35% distilled water; v/v), for a minimum of 24 h at 4 °C. Fixed tissues were dehydrated through a graded ethanol series at room temperature (RT) with gentle agitation (70% ethanol-1 h; 80% ethanol-1 h; 95% ethanol-1 h; 100% ethanol-1 h). Dehydrated samples were cleared by sequential incubation in an ethanol : xylene (1:1) mixture (1 h), and transferred to pure xylene (two changes, 1 h each) until tissues became translucent. Cleared tissues were infiltrated with molten paraffin wax (Paraplast X-TRA, melting point 56–58 °C) in an oven at 60 °C. Infiltrated tissues were oriented in pre-warmed metal molds filled with fresh paraffin. Blocks were solidified on a chilled plate (4 °C) to prevent crystallization. Paraffin blocks were sectioned at 1 mm thickness using a rotary microtome (Leica RM2235, Germany). Ribbons were floated on a 42 °C water bath, and mounted on pre-cleaned, charged glass slides. Histochemical staining was performed in triplicate.
Staining for starch analysis
-
Starch staining was performed as previously described[19]. Ultra-sectioned and fixed bud samples were dipped in Lugol's iodine solution (Fischer) for 2 min, after a 1-min vacuum infiltration in the same. Sections were subsequently rinsed in deionized water, twice, to wash off excess stain and subsequently imaged under a stereo microscope (Nikon- SMZ18, Japan). Starch quantification (area/optical density) was carried out using the Image J online tool.
Estimation of sugars using HPLC-RID
-
A 1 g sample was dissolved in 25 mL of HPLC-grade water. The solution was then centrifuged at 16,000 rpm for 10 min. The resulting supernatant was collected, and filtered through a 0.2 µm nylon filter prior to analysis. Sugar analysis was performed on a Shimadzu LC-20AD/T HPLC system equipped with a refractive index detector (RID-10A). Separation was achieved using a reverse-phase LC-NH2 column (25 cm × 4.6 mm, 5 μm) maintained at 35 °C. The mobile phase was an isocratic mixture of acetonitrile and water (85:15, v/v for one set of analyses, and 65:35, v/v for another), at a flow rate of 1.5 mL/min. The injection volume was 20 µL. Standard curves for fructose and sucrose (1%–5% w/v), and glucose (1%–10% w/v) were prepared in HPLC-grade water. All samples and standards were analyzed in triplicate. The sugar analysis was conducted at the National Instrumentation Center for Environmental Management (NICEM), Seoul National University, South Korea.
RNA extraction and differential gene expression (DEG) analysis
-
An illumina next generation seq 1000 platform was used to sequence the library. The quality of the raw data obtained by paired-end sequencing of the normalized cDNA library was assessed using FastQC software (
www.bioinformatics.babraham.ac.uk/projects/fastqc ). The de novo assembly, the RNA-Seq transcriptome library preparation, and the DEG analysis were carried out using an already established pipeline[20].Correlation analysis between RNA-Seq data and quantitative PCR
-
Total RNA was isolated from control and auxin-treated floral buds using the Ribospin Plant RNA isolation kit (GENEALL, Korea). Complementary DNA (cDNA) was synthesized from the same RNA samples used for RNA-Seq analysis, with the iScript Reverse Transcriptase mix (Bio-RAD, USA). Quantitative reverse-transcription PCR (qRT-PCR) was then performed using the IQ SYBR Green Supermix RT qPCR Kit (Bio-RAD, USA). Six DEGs (three up-/three down-) from the transcriptome data were used for RT-qPCR with a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA), according to the following program: 95 °C for 2 min, followed by 40 cycles at 95 °C for 10 s, 61 °C for 30 s, and 72 °C for 30 s. All RT-qPCR reactions were performed in three biological replicates per biological sample. The expression ratios were calculated by the 2−ΔΔCᴛ method[21]. ACTIN served as the internal reference gene for normalization. The expression patterns obtained from qRT-PCR were validated against the RNA-Seq data by calculating the Pearson correlation coefficient using R. All primer sequences used in this study are provided in Supplementary Table S1.
Statistical analysis
-
Data are presented as mean ± SE. Statistical significance was determined by one-way ANOVA, followed by Tukey's least significance difference (LSD) test in R (v.3.5.1). Differences were considered significant at p < 0.05. The field experiments were conducted twice (simultaneously) during the study period.
-
Starch buildup is required in pre-nectaries prior to nectar synthesis, and is rapidly broken down to produce sucrose and hexoses during the post-secretory phase[22]. Knowledge regarding the starch degradation into sucrose, glucose, and fructose, with the help of sucrose synthases, is available and clearly understood; however, information regarding the starch buildup during the early phases of nectary synthesis are limited, and not fully understood[5]. In particular, phytohormones such as JA and auxin are known to increase the overall nectar volume, but whether they influence pre-nectary starch accumulation is still unanswered[8].
To test this, Platycodon grandiflorus individual buds (~1.5 cm) (pre-secretory stage) were sprayed with 1 and 100 μM auxin solution, and water alone (control). Buds treated with auxin were collected fresh after 24 and 48 h, respectively, and immediately cold-stored at 4 °C. The stored buds were then fixed in FAA fixative, and the μm-thick sections were made using an ultra-microtome (Leica, Germany). The obtained tissue sections were stained with 5% lugol stain, and left for 1 min at room temperature. The stained sections were observed through a confocal light microscope (Nikon, Japan). No changes were observed in buds after 24 h, irrespective of the treatments (data not shown), similar to floral buds treated with 1 μM auxin (Fig. 1), whereas bud treatment with 100 μM auxin showed enhanced starch accumulation 48 h after treatment, compared to the control (Fig. 1). Quantitative analysis of starch accumulation, prior to secretion, revealed a significant effect of auxin treatment. ImageJ analysis of starch-specific staining (lugol stain) showed that the mean starch granule area per cell (mean optical density per glandular area) was significantly higher in tissues treated with 100 μM auxin, compared to the control (Fig. 2). This image-based quantification directly correlated with the observations from anatomical sectioning (Fig. 1). Histological sections of control tissues showed sparse, small starch granules in the pre-secretory cells, whereas sections from 100 μM auxin-treated tissues displayed a marked increase in both the number and size of starch granules within the same cellular compartments (Fig. 2). From these results, auxin was confirmed as being able to build up starch storage at the pre-secretory cells; however, the molecular mechanism behind this rapid carbohydrate storage is unknown.
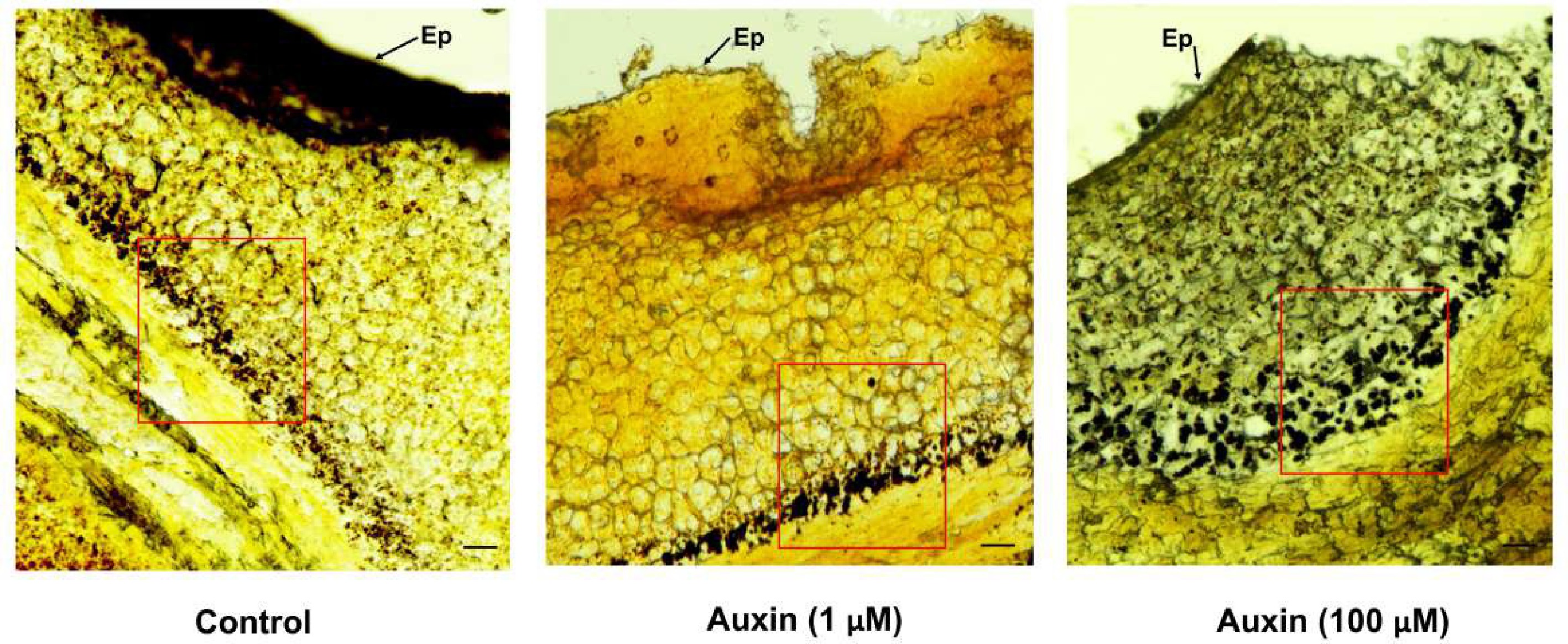
Figure 1.
Light microscopic images of 1 μm thick sections of P. grandiflorus nectaries stained with lugol reagent. Starch granules are shown in the control, and auxin treatments in pre-secretory parenchymal cells. Ep-Epidermal layer; Bar = 100 μm.
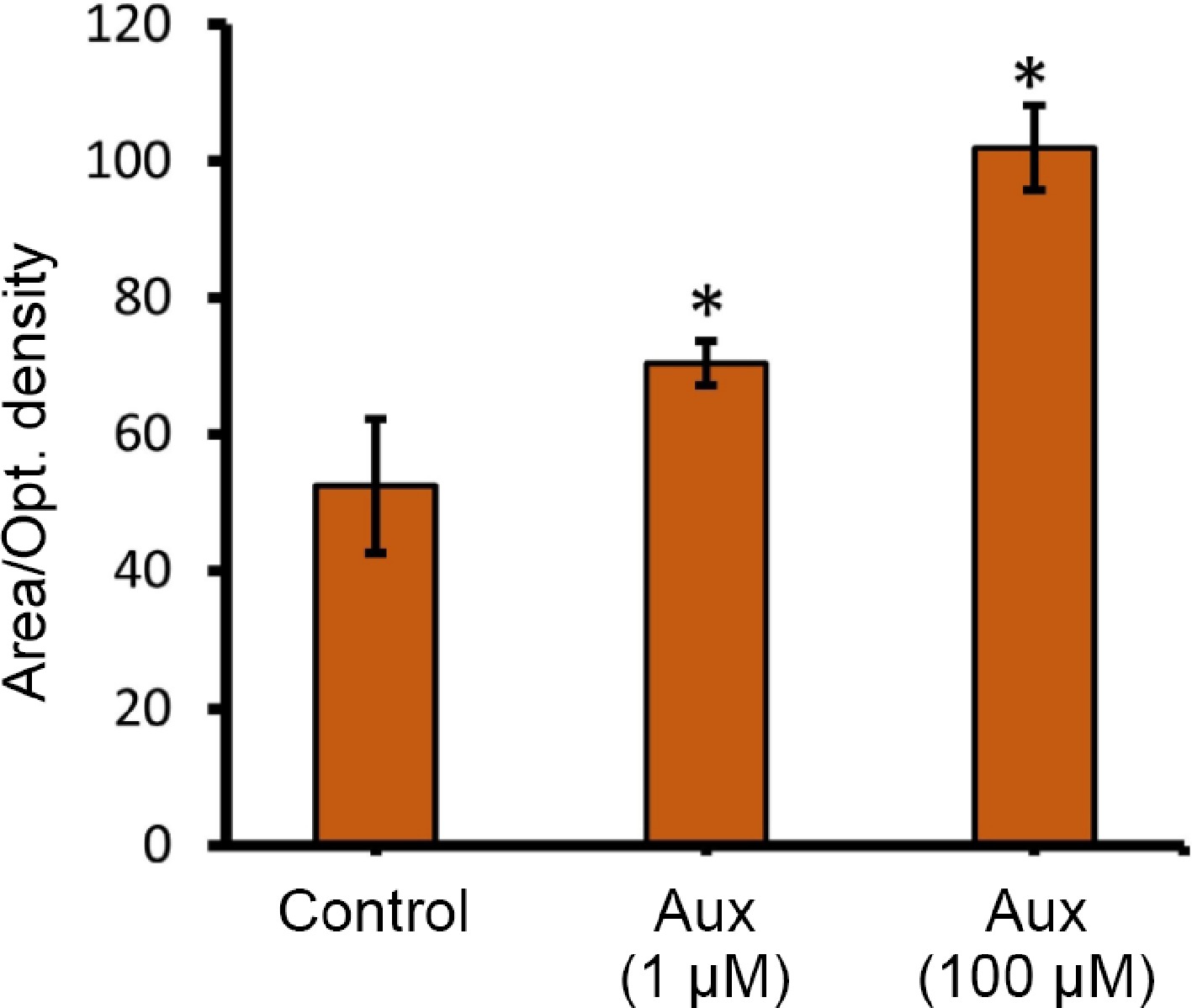
Figure 2.
Quantification of starch accumulation differences among control, and auxin treatments via ImageJ software (area/optical density). The Image J online browser was used. Image > Type (8-bit) > Adjust > Threshold (set 70 for iodine stain) > select rectangle tool to select the region > Measure. Use the tracing tool to select similar regions in treatment groups and measure. Quantification was carried out in three different spots unanimously, and the readings were taken. * Represents statistical significance between the treatments.
To determine the molecular mechanisms, the floral buds treated with 100 μM auxin, and control (water) after 48 h treatments were frozen in liquid nitrogen, and stored at −80 °C, for further analysis. Stored samples were ground using liquid nitrogen, and the total RNA was extracted. High-throughput RNA sequencing was performed. Transcriptome analysis was conducted to identify the most highly Differentially Expressed Genes (DEGs). From the present results, 91 genes were identified as being upregulated, and 41 genes downregulated (auxin vs control). Gene ontology biological process (GOBP) analysis showed many of the upregulated genes were from the sugar metabolic pathway. Multiple sucrose transport genes, like STP1, STP2, and STP8 were found to be highly upregulated (Fig. 3, Supplementary Table S2). Plasma membrane localized sugar transporters have been found to transport sucrose in the guard cells of Arabidopsis thaliana[23]. The transported sugar from phloem is stored as starch in the pre-secretory cells[7]. Interestingly, genes involved in the conversion of glucose to starch, such as G6P, FEH, and UGP2, were found to show increased expression, compared to the control treatments (Supplementary Table S2). An interesting transcription factor MYB104, was also found to be highly upregulated. It has been previously reported that CiMYB5 and CiMYB3 transcription factors regulate the expression of 1-FEH genes in chicory[24]. Similarly, another MYB transcription factor (SlMYB) acts as a transcriptional activator that directly controls the expression of genes involved in sucrose uptake, and vacuolar storage[25]. From the present results, as well as from previous reports, it is highly plausible that MYB104 acts as a key regulator of sugar allocation. The most parsimonious initial hypothesis is that MYB104 directly regulates genes involved in starch conversion (like amylases, BAM, DBE., etc), rather than directing STP family genes. However, a role in regulating specific STP genes for specific tissue-level sugar partitioning cannot be ruled out, and needs further investigation. In addition, several genes playing crucial roles in regulating flowering time, such as BRR2A, LDL3, GA20Ox2, and BBX28 were found to be upregulated. At the same time, two negative regulators of flowering, such as CNGC2 and BKK1, were found to be downregulated (Supplementary Table S2). An ATP-binding cassette (ABCG1), which plays a major role in auxin distribution, was seen to be upregulated[26]. In cucumber, CsSWEET7 was involved in phloem unloading, and sugar partitioning in pre-secretory tissues[4]. The present transcriptome data were further validated using the qRT-PCR gene expression analysis, with genes showing differential expression (Fig. 4). The sectioning results, and transcriptome analysis mainly indicate one thing in common, rapid inflow of sucrose from phloem, and the conversion of the abundant sucrose to starch, for the carbohydrate uploading mediated by exogenous auxin (100 μM). To confirm whether this enormous carbohydrate uploading has resulted in enhanced nectar secretion, the nectar volume, and nectar composition were assessed on the second day after flower opening.
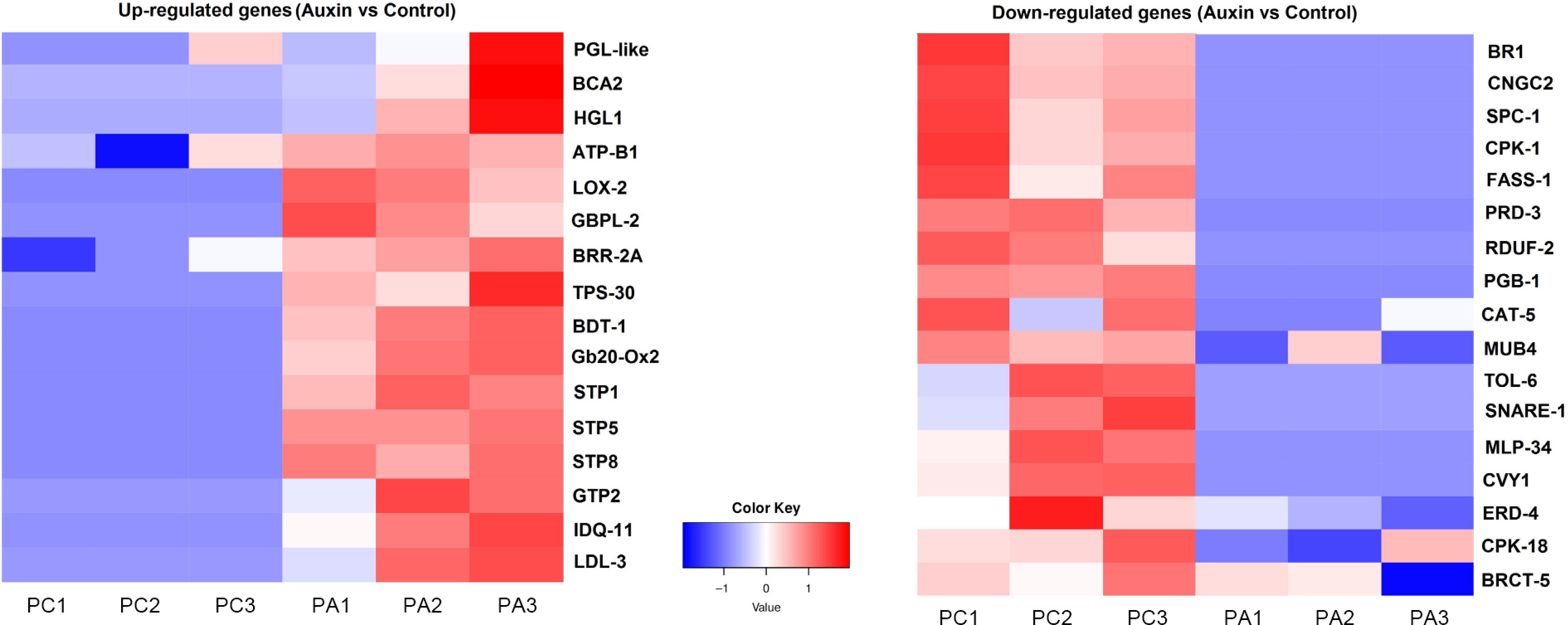
Figure 3.
Heat map showing highly upregulated and downregulated genes (auxin vs control). Sucrose transported genes STP1 and STP2 are found to be highly upregulated under auxin treatment.
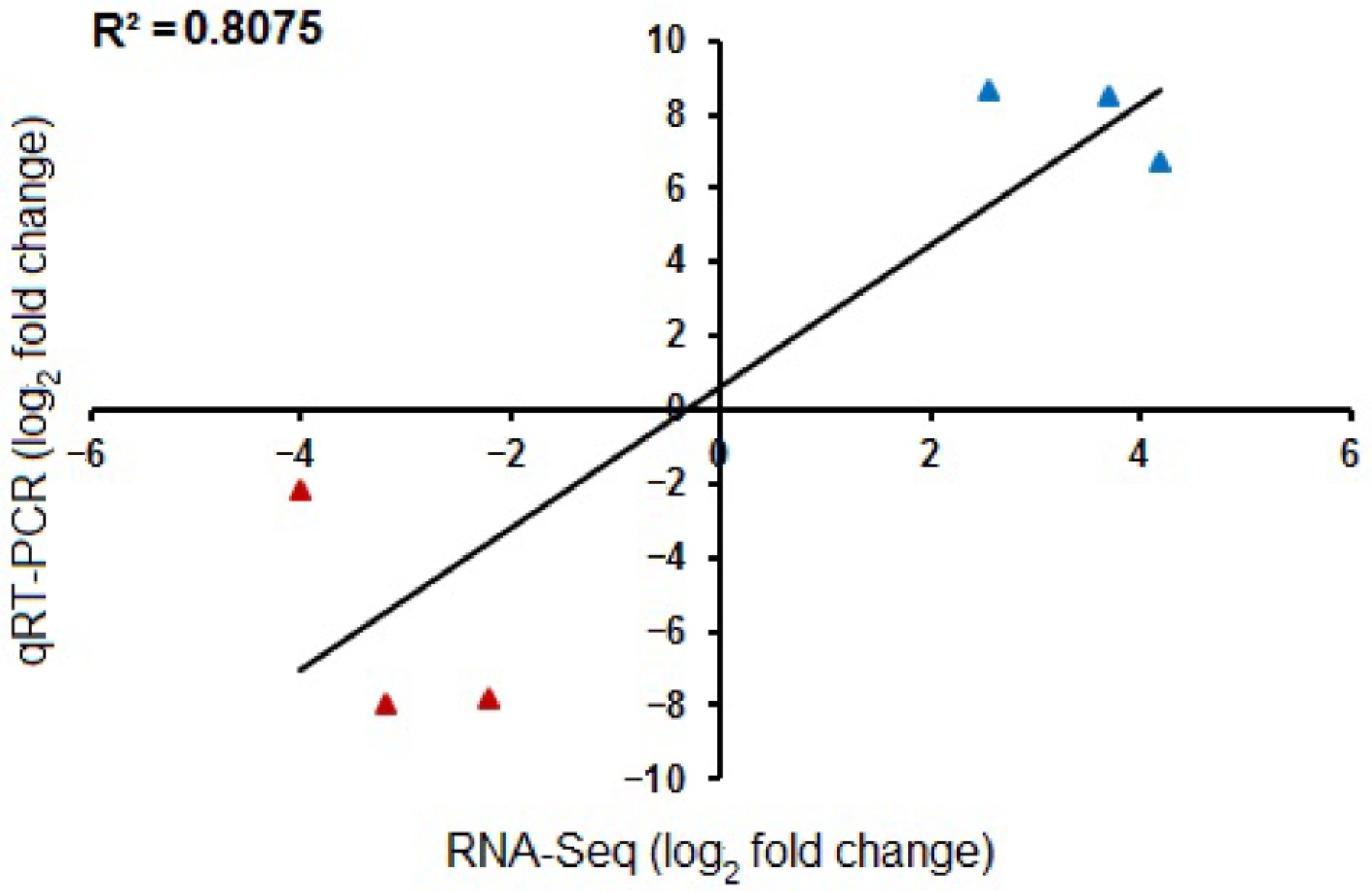
Figure 4.
Validation of −log2FC RNA-Seq values with the −log2FC qRT-PCR gene expression values. The qRT-PCR results (both upregulated and downregulated genes) show high correlation with RNA-Seq data, as denoted by an R2 value of 0.8075. Genes used for qRT-PCR include, upregulation-STP1, STP2, and ABCG1; downregulation-SPC1, CNGC2, and BKK1.
Auxin treatment significantly increased nectar sugar production, with auxin-treated floral buds yielding more than twice the glucose and fructose content compared to the control (Figs. 5a, b, 6). Such differences are possibly related to the excess amounts of sucrose being transported from the phloem, with the help of sugar transporters, and converted to starch, mediated by auxin. It has to be noted that the carbohydrate buildup at the pre-secretory cells may have been split to glucose and fructose by various cleavage enzymes like invertases or sucrose synthases during the post-secretory stage[27]. This dramatic increase suggests auxin plays a key role in regulating nectar secretion, potentially influencing pollinator attraction. Starch degradation in nectaries has also been reported in other day-flowering species[22,28]. More importantly, a key function of GA is to promote the breakdown of starch into soluble sugars (like glucose and fructose)[29]. The nectaries of many plants accumulate starch before flowering. GA signaling triggers the enzymatic conversion of this starch reserve into osmotically active sugars, which drives the osmotic flow of water into the nectary, and ultimately increases nectar volume[30]. The present results show that auxin treatment significantly enriched the expression of GIBBERELLIN 20 OXIDASE 2 (GA20ox2) (Supplementary Table S2). Given the established cross-talk between phytohormones, it is proposed that exogenous auxin promotes starch degradation, a key driver of nectar volume, via the induction of GA20ox2[31]. Future research on GA20ox2 should clarify its role in mediating auxin-GA cross-talk during nectar secretion.
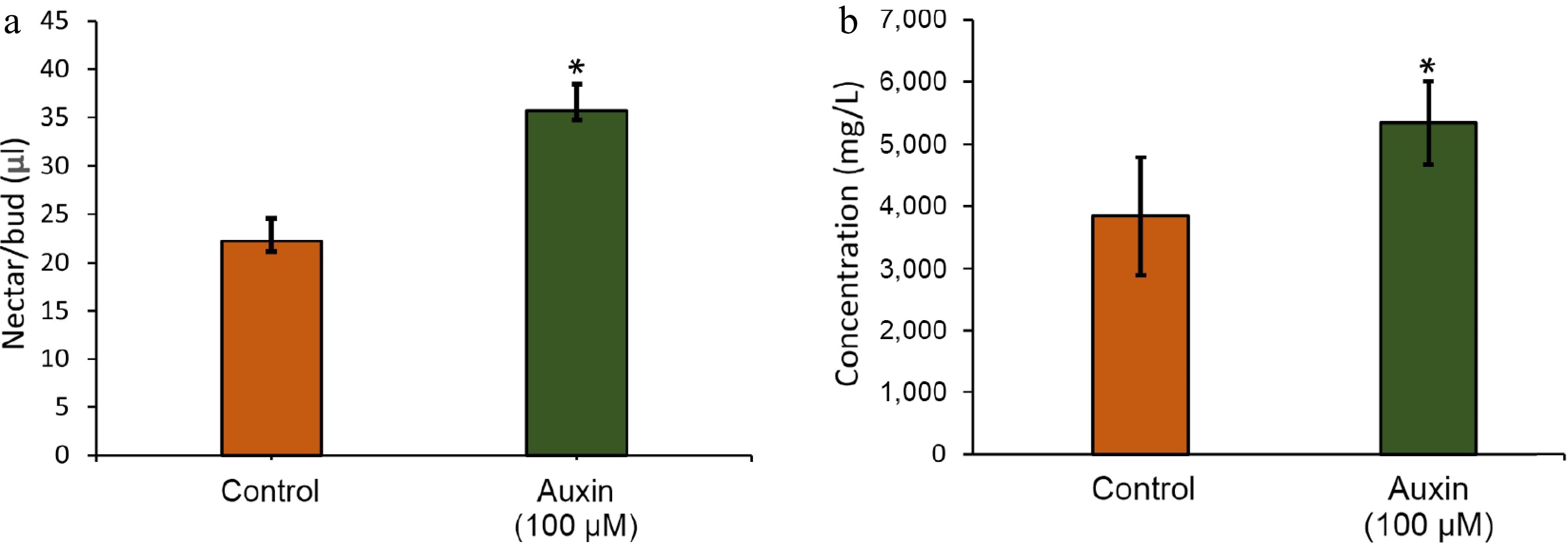
Figure 5.
(a) Volume of nectar as physically measured using a micropipette. (b) Total sugar concentration in the nectaries treated with control and auxin treatment.
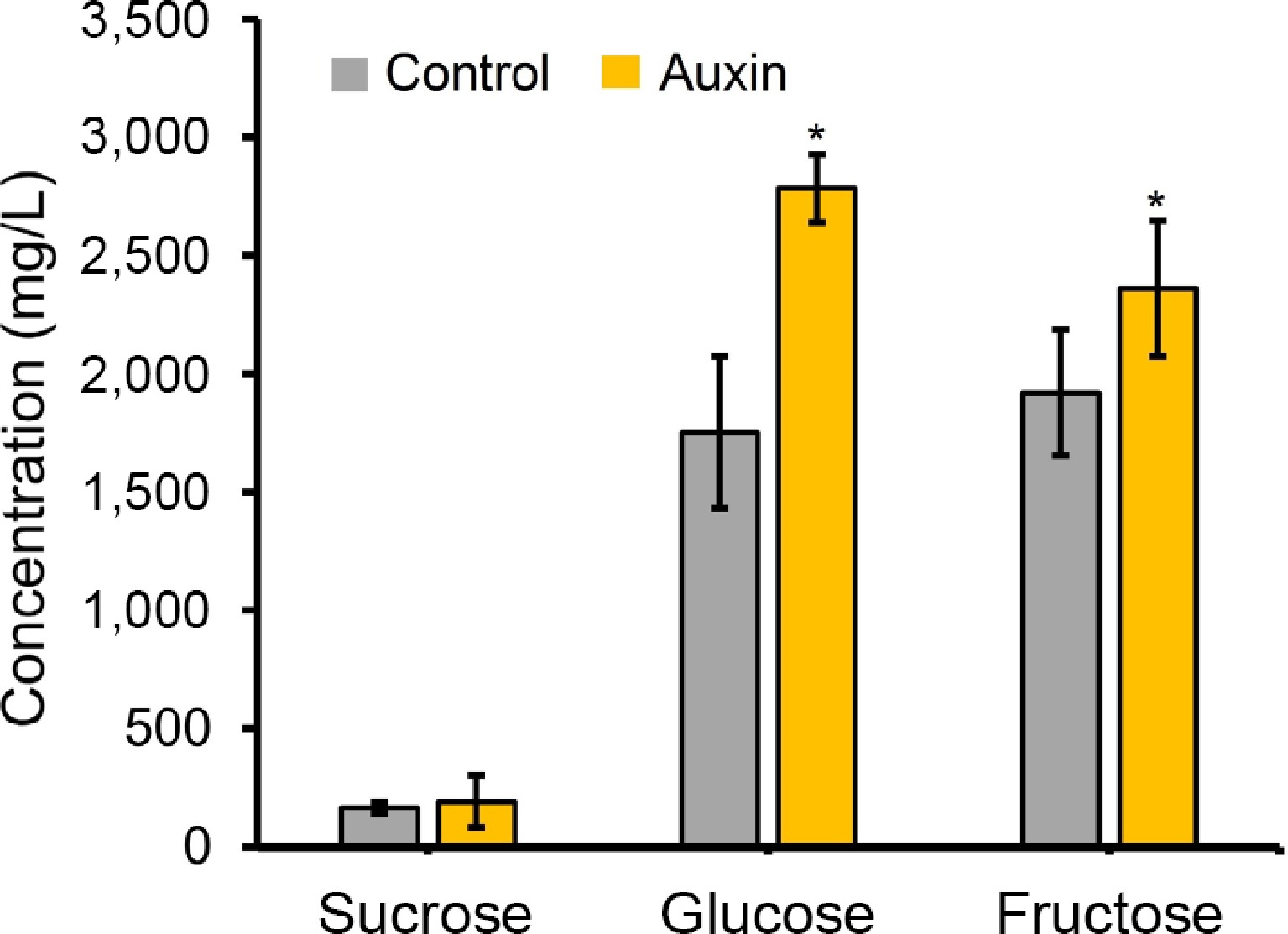
Figure 6.
Sugar component analysis from the nectar obtained (control and auxin [100 uM] treatments). Sugar components were detected using the HPLC method. * Represents statistical significance between the treatments.
This can be hypothesized as early auxin maxima not only specifiying nectary fate, but also upregulating genes involved in starch biosynthesis and storage within the nectary parenchyma. This would ensure the nectary accumulates sufficient starch reserves, the raw material for future nectar secretion. In this early phase, auxin and GA may act synergistically, as both are promoters of growth and sink strength, potentially coordinating the flow of photo-assimilates into the developing nectary[31]. Crucially, the present findings that auxin upregulates GA20ox2 suggests a mechanism by which auxin pre-programs the tissue for this switch, priming the GA pathway for later activation.
Further, the present transcriptome data revealed the significant enrichment of lipoxygenase (LOX2), a key gene encoding the first committed enzyme in the JA biosynthetic pathway (Supplementary Table S2). This finding provides direct molecular evidence that JA biosynthesis is activated during exogenous auxin treatment, positioning JA not merely as a signal, but as an actively synthesized regulator within the present system. This aligns with established models where a local burst of JA is essential for initiating nectary function[31]. The identification of the JA biosynthetic gene LOX, allows for the proposal of a more detailed model for the hormonal cross-talk regulating nectar accumulation in P. grandiflorus. It is speculated that the early developmental signal, provided by auxin, primes the parenchymal tissue, which is followed by a critical activation of JA biosynthesis, as evidenced by the upregulation of LOX. This newly synthesized JA then acts as the key trigger to initiate the secretory program, potentially by activating transcription factors that prepare the nectary for sugar export. Subsequently, the GA-mediated pathway, potentially pre-induced by auxin, drives the final step of starch mobilization to determine nectar volume. The co-occurrence of LOX upregulation, with our previous finding of auxin-induced GA20ox2, suggests a sophisticated, sequential hormone cascade[29]. A plausible hypothesis is that auxin signaling creates a competent state that permits, or even stimulates JA biosynthesis (via LOX), and this JA burst then modulates the GA pathway to execute the final physiological output. Future work using hormone mutants or inhibitors will be crucial to disentangle this sequence of events.
Collectively, the present results confirm that the auxin establishes the nectary secretion, and upregulates GA20ox2, priming the GA pathway which might create a permissive environment for JA biosynthesis (upregulation of LOX2). Eventually, this JA signal might have initiated the secretory program, promoting the expression of sucrose transporters (STPs), and activating enzymes for cell wall modification to facilitate nectar transport (Fig. 7). CRISPR-based gene editing of critical genes like GA20ox2 and LOX2 might further confirm this tri-hormonal cross-talk in regulating nectar synthesis and secretion. Thus, beyond its developmental roles, auxin orchestrates the temporal alignment of nectar sugar production with floral anthesis. This ensures that pollinator visitation is met with a maximized energy reward, highlighting a key hormonal link in the pollination ecology of P. grandiflorus.
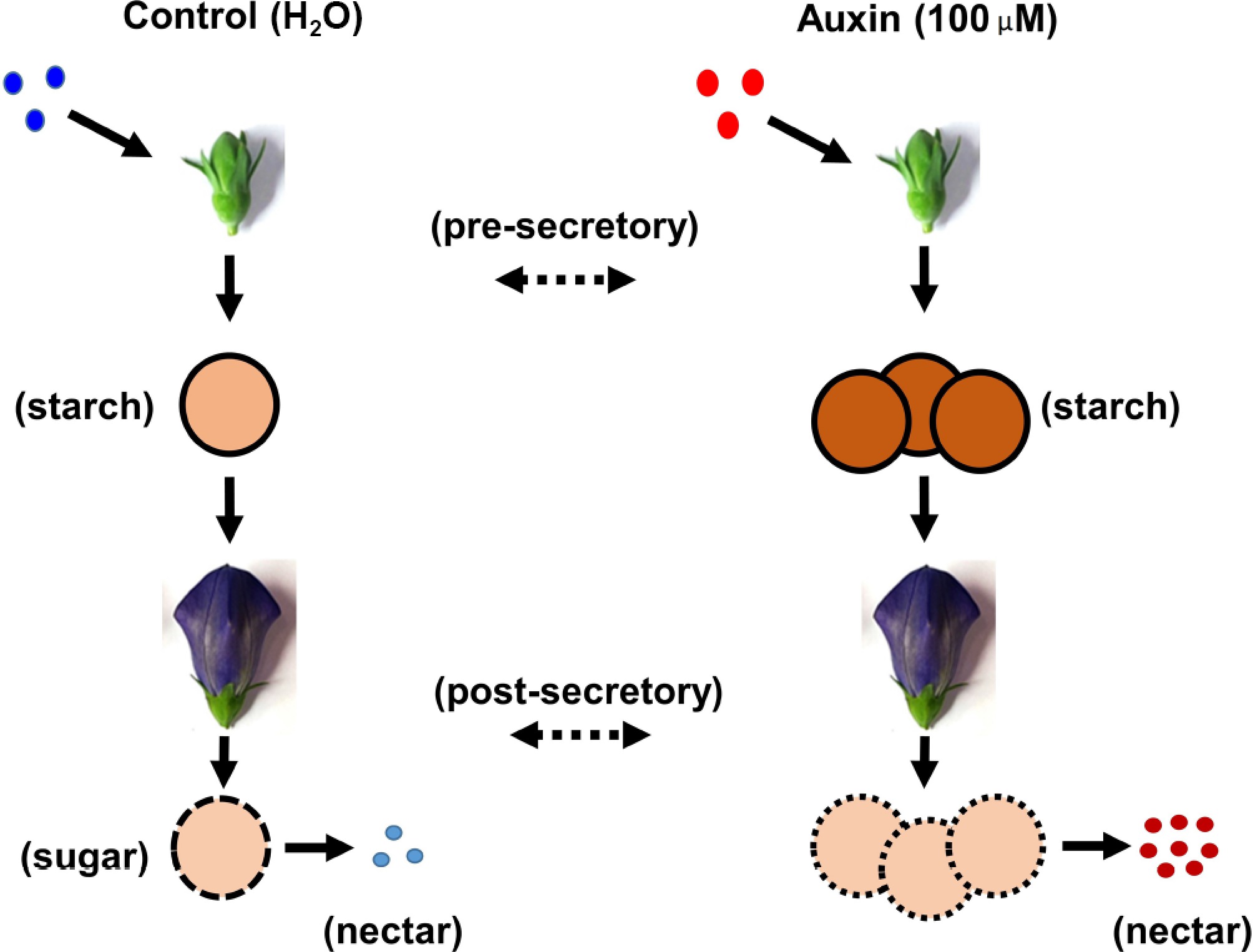
Figure 7.
Graphical illustration of the auxin treatment in the nectaries of P. grandiflorus.
This work was supported by a basic science research program grant through National Research Foundation (NRF) funded by the Ministry of Education (2021R111A2044159), Republic of Korea.
-
The authors confirm their contributions to the paper as follows: designed the study: Chandrasekaran U, Kim HS; performed the field, laboratory experiments, and wrote the manuscript: Chandrasekaran U; assisted with laboratory experiments: Kim J; assisted with green house setting, and field experiments: Oh NH, Choe H. All authors reviewed the results and approved the final version of the manuscript.
-
All the raw read sequences were deposited in the NCBI sequence read archive (SRA) under Accession No. PRJNA1348704.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 List of primers used in this study.
- Supplementary Table S2 List of genes differentially expressed after exogenous auxin (100 µM) treatment.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chandrasekaran U, Kim J, Oh NH, Choe H, Kim HS. 2026. Genetic basis for auxin induced pre-secretory starch accumulation in Platycodon grandiflorus. Plant Hormones 2: e003 doi: 10.48130/ph-0026-0001 

Genetic basis for auxin induced pre-secretory starch accumulation in Platycodon grandiflorus
- Received: 27 October 2025
- Revised: 31 December 2025
- Accepted: 06 January 2026
- Published online: 03 February 2026
Abstract: Nectar serves as the primary carbohydrate source for pollinators and defensive mutualists. Despite advancements in research, the mechanisms regulating pre-nectar carbohydrate transport from the phloem remain poorly understood. Could an exogenous factor, such as a phytohormone, influence this transport during the pre-secretory stage of nectar synthesis? This study aims to uncover the genetic basis for auxin-induced pre-secretory starch accumulation in the herbal plant Platycodon grandiflorus. It is demonstrated that exogenous application of auxin (100 µM) enhances starch accumulation in pre-secretory cells. Transcriptomic analysis of pre-secretory tissues revealed significant upregulation of multiple sucrose transporters (STP1, STP2, and STP3), glucose-starch conversion genes (G6P, UDP, and FRUH), GAOx2, and LOX2 in auxin-treated floral buds. These genes play a critical role in sucrose transport and its subsequent conversion to starch. The degradation of pre-secretory starch was confirmed by an increase in nectar volume and sugar (glucose and fructose) composition during the post-secretory stage, compared to the control. Collectively, the present findings suggest that exogenous auxin activates sucrose transporters, facilitating rapid sugar transport from the phloem to pre-secretory cells, through a possible GA × JA × auxin phytohormone cross-talk. This sugar is then converted into starch granules, ultimately driving a rapid sugar influx during the post-secretory stage.
-
Key words:
- Floral bud /
- Auxin /
- Pre-secretory stage /
- Starch accumulation /
- Sugar conversion