









-
RNA modifications, including N6-methyladenosine (m6A), N7-methylguanosine (m7G), 5-methylcytosine (m5C), N1-methyladenosine (m1A), pseudouridine (Ψ), and N4-acetylcytidine (ac4C), play vital roles in regulating fundamental RNA metabolism, such as mRNA's stability and translation efficiency, thus playing essential roles in modulating various plant development and stress responses[1,2]. Among these, m5C is a conserved RNA methylation modification formed by the addition of a methyl (-CH3) group to the fifth carbon position of cytosine residues. It is widespread in transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), and messenger RNAs (mRNAs) across eukaryotes, archaea, and some bacteria and is generally regulated by three key classes of proteins: Writers (methyltransferases), erasers (demethylases), and reader proteins[3,4]. In contrast to m6A modification, whose writers, erasers, and reader proteins have been well characterized and proven to play pivotal regulatory roles in various physiological processes[2,5−7], the molecular components underlying m5C methylation remain largely elusive in plants. To date, only a limited number of m5C methyltransferases have been identified in plants, including transfer RNA aspartic acid methyltransferase 1 (AtTRDMT1) and tRNA-specific methyltransferase 4A/B (AtTRM4A/B) in Arabidopsis thaliana (orthologs of human DNMT2 and NSUN2, respectively), as well as the m5C methyltransferase OsNSUN2 in rice (Oryza sativa)[8,9]. The A. thaliana Alkylation B (ALKB) domain-containing protein AtALKBH6 has been shown to directly bind to m5C-modified RNAs in vitro, thus qualifying as a putative m5C demethylase[10]. Additionally, two Aly/REF export factor (ALYREF) family proteins in A. thaliana, ALY2 and ALY4, have been identified as m5C reader proteins[11,12]. It is distinctly different from m6A modification, which is recognized by YT521-B homology (YTH)-domain proteins, such as YTHDF1-3[13].
Emerging evidence indicates that m5C methylation participates in plant development and plants' abiotic stress responses. For instance, AtTRM4B in A. thaliana regulates root development by modulating the stability and abundance of m5C-modified transcripts of key root-development genes, including Short hypocotyl 2 (SHY2) and Indoleacetic acid-induced protein 16 (IAA16)[14]. The OsNSUN2 mutation led to shortened root length in rice, further supporting a role for m5C in developmental regulation, although the underlying molecular mechanism remains elusive[9]. Moreover, attrm4b mutants of A. thaliana exhibit enhanced sensitivity to oxidative stress[8], whereas osnsun2 mutants of rice display a pronounced heat-sensitive phenotype, associated with reactive oxygen species (ROS) accumulation caused by photosystem defects[9]. Collectively, these studies demonstrate that m5C methylation possesses broad regulatory functions comparable with m6A modifications, particularly in modulating plants' development and abiotic stress responses. However, the role of m5C in plants' responses to biotic stress remains largely unclear.
Tomato (Solanum lycopersicum) is one of the most economically important horticultural crops worldwide, valued for its essential nutritional components and strong adaptability to diverse growth environments. However, postharvest decay caused by fungal pathogens remains a major constraint in tomato production and storage, leading to an annual global yield loss of 15%–25%[15]. Among these, B. cinerea, a ubiquitous necrotrophic fungus, causes gray mold disease, characterized by water-soaked lesions and rapid fruit decay, severely compromising fruit quality and commercial value[16]. To defend against B. cinerea infection, tomato fruits have evolved complex defense mechanisms, including transcriptional activation of defense-related genes[17], protein kinase-mediated signal transduction[18], synthesis of antimicrobial compounds[19], and modulation of hormone signaling pathways, particularly those involving jasmonic acid (JA) and ethylene[20−22].
The activation of defense-related genes is tightly controlled by multiple layers of gene expression regulation. Several key transcription factors, such as the helix–loop–helix (bHLH) transcription factor SlMYC2[23], the R2R3 MYB transcription factor SlMYB75[24], and the B-box (BBX) family transcription factor SlBBX20[25], have been demonstrated to directly bind to and activate defense-related genes at the transcriptional level. Additionally, epigenetic modifications, such as DNA methylation[26], noncoding RNAs[27], and microRNAs[28], have also been proven to function in upregulating the expression of defense-related genes. In recent, m6A methylation was shown to enhance resistance to B. cinerea in leaves of A. thaliana by stabilizing defense-related genes at the post-transcriptional level[29]. These findings highlight RNA modification as a critical regulatory layer in plants' defense responses and raise the possibility of other RNA modifications. However, the role of m5C modification in B. cinerea invasion, particularly during the postharvest stage of tomato fruits, remains unknown.
Traditional methods for detecting m5C modifications, such as bisulfite sequencing, suffer from limited resolution and an inability to capture full-length RNA molecules. To overcome these limitations, Nanopore direct RNA sequencing (DRS) has emerged as a powerful technology that enables single-molecule, full-length RNA sequencing and direct detection of RNA modifications based on characteristic current signals, without chemical treatment[30]. In this study, we used Nanopore DRS technology to systematically profile the m5C methylomes in B. cinerea-infected and mock-inoculated tomato fruits. We aimed to characterize the global landscape of m5C modification during B. cinerea infection, identify dynamic changes in the m5C sites associated with hormone- and defense-related genes, and uncover key m5C-modified transcripts in tomato's resistance to B. cinerea. This study provides new insights into the RNA epigenetic regulatory networks underlying tomato fruit's defense against B. cinerea and establish a foundation for improving postharvest resistance in tomato.
-
Tomato seedlings (Solanum lycopersicum cv. 'Micro-Tom') were planted in a greenhouse under standard cultivation conditions (23 °C, 16 h light/8 h dark photoperiod, and 60% relative humidity). Mature green tomato fruits at approximately 40 days postanthesis (DPA) were harvested. The fruits were surface-sterilized with a NaClO solution (2%, v/v) for 2 min, rinsed with sterile water three times, and surface-dried for subsequent use.
B. cinerea culture and inoculation
-
B. cinerea (B05.10) was cultured on solid potato dextrose agar (PDA) medium for 7–14 days. The culture condition was set at 25 °C under a 16 h light/8 h dark photoperiod. Conidia were collected in a half-strength liquid potato dextrose broth (PDB) medium, and the conidial concentration was adjusted to 5 × 105 spores per mL. For inoculation, the conidial suspension (5 μL) was dropped onto the surface of tomato fruits. Mock-inoculated fruits (control group) were treated with 5 μL of half-strength PDB medium without conidia. All inoculated fruits were placed in a growth chamber with a conditions of 25 °C and 80% relative humidity. At 48 h postinoculation, pericarp tissues with an area approximately 2–3 mm thick around the lesions were collected for RNA extraction.
Total RNA extraction
-
Total RNAs were extracted from the pericarp tissues of B. cinerea-infected and control fruits using the hot phenol method as described by Moore et al[31]. Briefly, 0.5 g of the powdered sample was mixed with 1 mL of an RNA extraction buffer (100 mM Tris-HCl, pH 8.0, 10 mM ethylenediaminetetraacetic acid [EDTA], 100 mM LiCl, 1% (w/v) sodium dodecyl sulfate [SDS], and 50% (v/v) water-saturated phenol) in a 2-mL centrifugation tube, followed by vortexing at room temperature for 10 min. After the addition of 500 µL of chloroform, the mixture was vortexed at room temperature for another 10 min, followed by centrifugation at 15,000×g for 20 min. The supernatant was transferred to a new 1.5-mL centrifugation tube and extracted once again with 500 µL of chloroform. Subsequently, total RNAs were precipitated with 2 M LiCl, washed twice with 75% (v/v) ethanol, and dissolved in Diethyl pyrocarbonate (DEPC)-treated water. The purity and integrity of total RNAs were evaluated using a NanoDrop 2000 spectrophotometer and a 1.5% agarose gel electrophoresis, respectively.
Nanopore direct RNA sequencing
-
Nanopore direct RNA sequencing (DRS) was performed by Benagen Technology Co., Ltd. (Wuhan, China), following the method described by Song et al.[32]. In brief, total RNA was used to isolate mRNAs using the mRNA Dynabeads Kit (Thermo Fisher, 61006). The enriched mRNAs were then ligated to Nanopore RT adapters using T4 DNA ligase (NEB, M0202M). The resulting ligation products were purified using Agencourt RNACleanTM XP beads (Beckman Coulter, California, USA). Subsequently, after the addition of the RNA adapter, the obtained products were purified again with Agencourt RNACleanTM XP beads. The prepared library was loaded into an R9.4 sequencing flow cell and then sequenced on a PromethION sequencer (Nanopore Technologies, Oxford, UK). Three independent biological replicates were subjected to generate real-time single-molecule sequencing data.
m5C data analysis
-
Raw sequencing data in FAST5 format were converted to FASTQ format following base calling by GUPPY software (version 3.2.6). Raw reads were filtered using NanoFilt (version 2.8.0) to remove adapter sequences and low-quality reads (quality score < 10). The filtered clean reads were then mapped to the ITAG3.2_release reference annotation (
https://ftp.solgenomics.net/tomato_genome/ ) using Minimap2 software (version 2.2.17)[33], with the tomato genome build SL3.0 as the reference genome. Statistical comparisons of the mapping results were conducted using Samtools (version 1.11)[34]. Subsequently, m5C modification sites were identified using Modkit software (version 0.4.1) with the 'detect_modifications' module, which detects RNA modifications on the basis of current signal deviations from the reference sequence. m5C sites with a fraction score > 0.7 were defined as high-confidence sites. Differential m5C sites were identified using the R package 'DESeq2' with the criterion of p < 0.05. Motif analysis was performed using MEME software (version 5.5.1)[35]. Gene Ontology (GO) enrichment analysis was carried out on the Gene Ontology Consortium platform (https://www.geneontology.org/ ). GO terms that with a Fisher's exact false discovery rate (FDR) < 0.05 were statistically significant.Transcriptome-wide gene expression analysis
-
The raw DRS sequencing data were concurrently used to perform gene expression analysis. The transcript abundance of gene transcripts was calculated as transcripts per million (TPM) using Salmon software (version 1.9.0)[36]. Differential expression analysis was conducted using the R package 'DESeq2' with the criteria of log2(fold change) ≥ 1 and an adjusted p-value < 0.05.
Phylogenetic analysis
-
The protein sequence from A. thaliana AtALKBH6 were downloaded from the Arabidopsis Information Resource (TAIR;
www.arabidopsis.org ). Using this sequence as a query, putative homologous proteins across multiple plant species were identified in the Phytozome database (https://phytozome-next.jgi.doe.gov ). Multiple protein sequence alignments were performed using Clustal X (version 2.1) with the default settings. These resulting alignment data were imported into MEGA (version 5.2) for phylogenetic tree construction via the neighbor-joining algorithm, with 1,000 bootstrap replicates for assessing the reliability.Statistical analysis
-
All experiments in this study were carried out with three independently biological replicates. Microsoft Excel and GraphPad Prism 8.0 software were used to process and visualize the experimental data. GraphPad Prism 8.0 was used to analyze statistical significance (two-tailed Student's t-test).
-
To characterize the m5C methylome landscapes in tomato fruits in response to B. cinerea infection, Micro-Tom fruits at the mature green stage were harvested and inoculated with B. cinerea. Pericarp tissues approximately 2–3 mm thick surrounding the disease lesions were collected at 48 h postinoculation (Fig. 1a). Total RNA was extracted from three biological replicates of both mock-inoculated (control) and B. cinerea-infected fruit pericarps, followed by mRNA enrichment and library preparation. The constructed RNA libraries were subjected to Nanopore DRS, and the raw data were filtered and aligned to the tomato reference genome SL3.0. The mapped reads were of high quality, with an average length of approximately 1,000 nt and a mapping rate above 90% (Supplementary Table S1), supporting the reliability of the sequencing data.
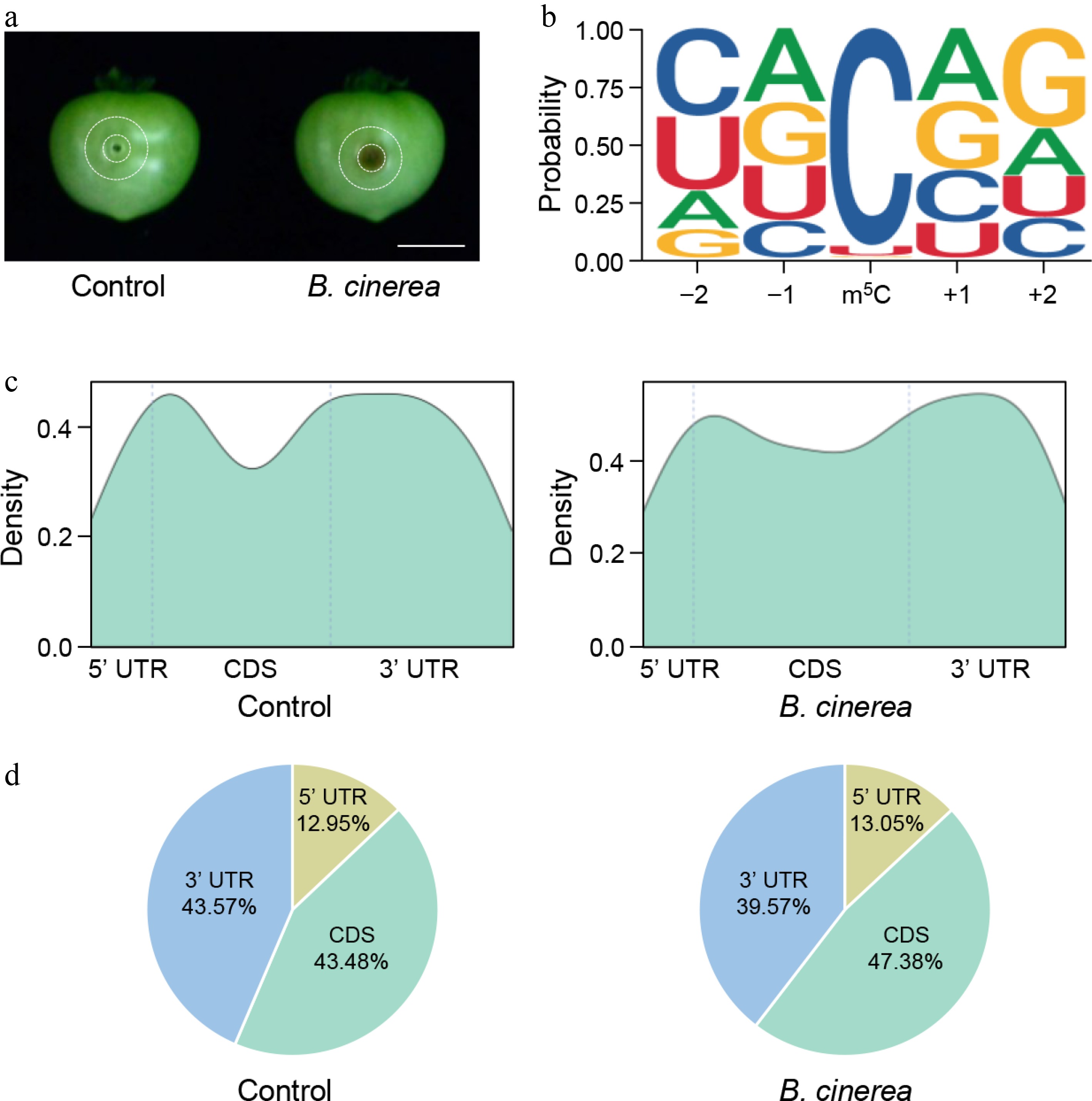
Figure 1.
m5C methylome profiles of tomato fruits under Botrytis cinerea infection. (a) Representative photographs of mock-inoculated and B. cinerea-infected tomato fruits at 48 h postinoculation. Micro-Tom fruits at the mature green stage were inoculated with B. cinerea, and pericarp tissues between the two indicated circles were collected as experimental materials. Scale bar = 1 cm. (b) Sequence motif identified around m5C sites using MEME software. All m5C sites with a fraction score > 0.7 were used for motif prediction analysis. (c) Metagenomic profiles of m5C site distributions in the 5' untranslated region (5' UTR), coding sequence (CDS), and 3' UTR region. (d) Pie charts showing the proportion of m5C sites in the 5' UTR, CDS, and 3' UTR.
The m5C modification sites were identified on the basis of their fraction scores, which represent the modification ratio and reflect the relative m5C methylation level. Sites with a fraction score > 0.7 were regarded as high-confidence m5C sites, and a total of 221, 233, and 242 high-confidence m5C sites were identified in the three control replicates, whereas 209, 177, and 201 sites were identified in the B. cinerea-infected tomato fruits. To clarify the nucleotide preference of m5C modifications, motif analysis was performed on these high-confidence sites. No obvious nucleotide preference was found at the –1 and +1 positions flanking the m5C site (Fig. 1b). By contrast, the –2 position was predominantly enriched in cytosine (C) and uracil (U), and the +2 position showed a significant enrichment of guanine (G) (Fig. 1b). This conserved motif pattern was comparable between the control and B. cinerea-infected groups, suggesting that the sequence preference of m5C modification in tomato fruits is an intrinsic feature and is not substantially altered by B. cinerea infection.
To further assess the positional preference of m5C sites across gene transcripts, each transcript was divided into three functional regions: the 5' untranscribed region (UTR), the coding sequence (CDS), and the 3' UTR. The distribution pattern of m5C sites was highly similar between control and B. cinerea-infected samples (Fig. 1c, d). In both groups, m5C sites were mainly distributed in the CDS and 3' UTR regions, each accounting for approximately 40% of the total m5C sites. In comparison, only almost 13% of m5C sites were distributed in the 5' UTR (Fig. 1c, d). This distribution pattern may be attributed to the shorter length of the 5' UTR and thus fewer potential modification sites. Collectively, these results indicate that B. cinerea infection does not alter the regional distribution pattern of m5C sites in tomato fruit mRNAs, and m5C modification has no obvious regional preference along gene transcripts.
Analysis of differential m5C sites upon B. cinerea infection
-
To investigate the effect of B. cinerea infection on m5C modification levels, differential m5C sites were identified by comparing the control and infected groups. m5C sites with a fraction score ≥ 0.1 in at least one group were defined as reliable m5C sites. In total, 39,610 reliable m5C sites were identified, which were distributed across 8,580 gene transcripts. Among these, 354 m5C sites were significantly upregulated and 341 were significantly downregulated in B. cinerea-infected fruits relative to the control group (p < 0.05; Fig. 2a), corresponding to 340 and 324 distinct gene transcripts, respectively (Supplementary Tables S2 and S3). These results suggest that B. cinerea infection triggers specific changes in m5C methylation levels across the tomato fruit transcriptome.
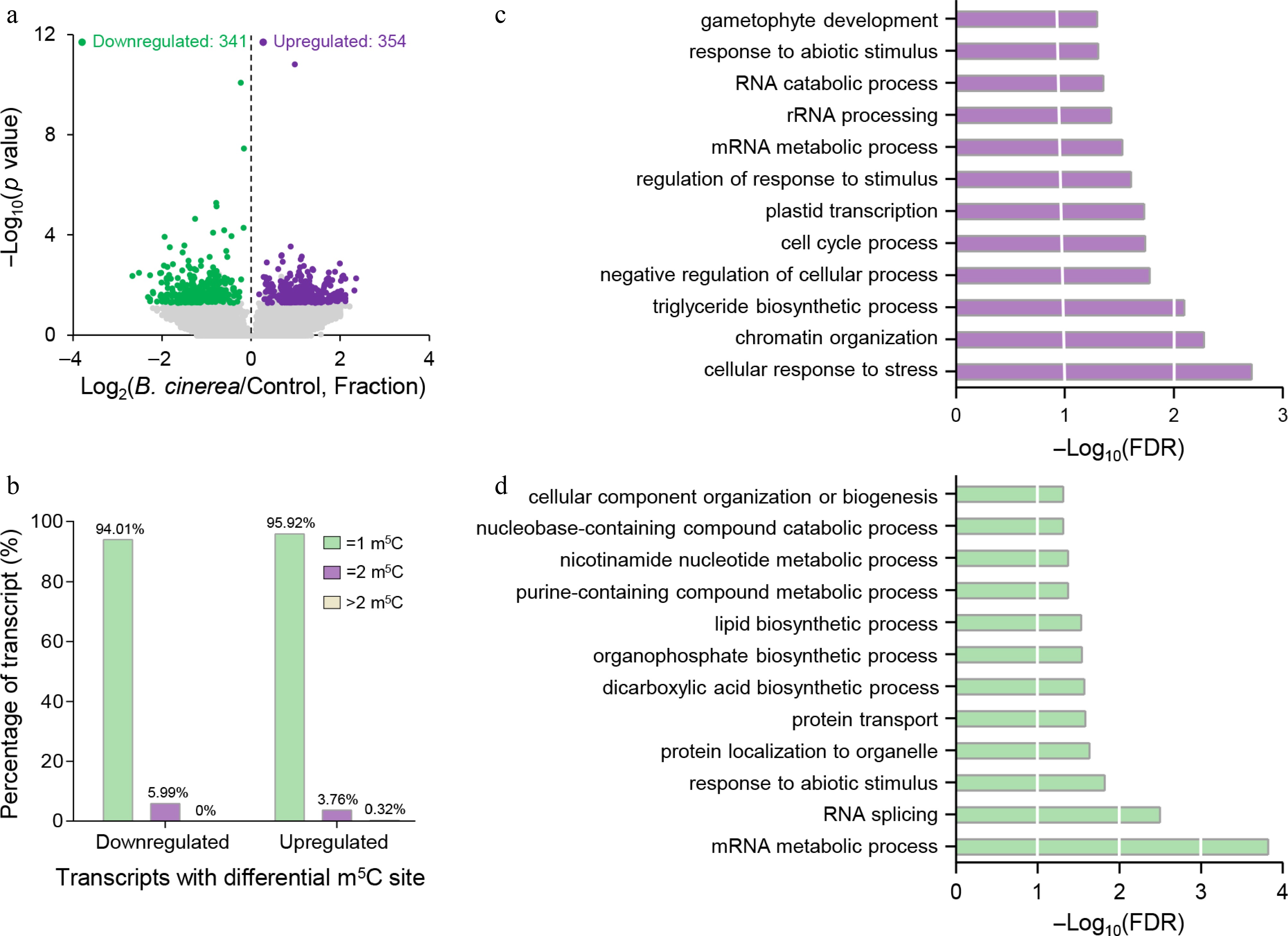
Figure 2.
Identification of differential m5C sites in B. cinerea-infected tomato fruits. (a) Volcano plot showing significantly unregulated (purple) and downregulated (green) m5C sites in B. cinerea-infected tomato fruits compared with the control group. (b) Proportions of the differential m5C-modified transcripts containing different m5C site numbers. (c) Gene Ontology (GO) enrichment analysis of biological processes for gene transcripts harboring significantly upregulated m5C sites. (d) GO enrichment analysis of biological processes for gene transcripts harboring significantly downregulated m5C sites. FDR, false discovery rate.
Further statistical analysis of transcripts harboring differential m5C sites revealed that the vast majority (approximately 95%) contained only one differential m5C site, whereas only a small proportion (around 5%) harbored two or more differential m5C sites (Fig. 2b). This observation indicates that the regulatory effect of B. cinerea infection on m5C modification in tomato fruits occurs primarily through site-specific local regulation, rather than coordinated modification changes across multiple sites within the same transcript.
To uncover the biological processes regulated by m5C modification in response to B. cinerea infection, GO enrichment analysis was performed on genes exhibiting differential m5C methylation. The results showed that genes harboring upregulated m5C sites were significantly enriched in several biological processes, including RNA metabolism (e.g., RNA catabolic process, rRNA processing, and mRNA metabolic process), responses to external stimuli (e.g., response to abiotic stimulus, regulation of responses to stimuli, and cellular response to stress), chromatin organization, and plastid transcription (Fig. 2c). By contrast, genes with downregulated m5C sites were not only enriched in RNA metabolism (e.g., RNA splicing and mRNA metabolic process) and responses to external stimuli (e.g., responses to abiotic stimuli) but also significantly enriched in protein localization and transport (e.g., protein transport and protein localization to organelles), as well as the biosynthesis and metabolism of diverse organic compounds including lipids, organophosphates, and dicarboxylic acids (Fig. 2d). These results suggest that m5C modification is broadly implicated in the regulation of multiple biological processes in tomato fruits during B. cinerea infection, implying multifaceted roles for m5C modification in mediating tomato fruit's resistance to B. cinerea.
m5C methylation is generally negatively correlated with gene expression upon B. cinerea infection
-
To explore the putative regulatory role of m5C modification in gene expression, we next analyzed the correlation between m5C modification levels and transcript expression abundance. A parallel transcriptome-wide gene expression analysis (RNA-seq) was performed in conjunction with Nanopore DRS analysis, and we identified 7,509 upregulated and 7,255 downregulated transcripts in the B. cinerea-infected group compared with the control group, respectively (Supplementary Table S4). Subsequently, an integrated analysis was conducted to associate transcripts harboring differential m5C sites with those exhibiting significantly altered expression levels. The results showed that among transcripts with upregulated m5C sites, 113 transcripts exhibited significantly decreased expression levels, whereas only 38 transcripts showed significantly increased expression levels (log2(fold change) ≥ 1; adjusted p-value < 0.05) in B. cinerea-infected fruits (Fig. 3a; Supplementary Table S5). By contrast, among transcripts with downregulated m5C modification, 48 transcripts had significantly decreased expression levels and 97 transcripts displayed significantly increased expression levels (Fig. 3b; Supplementary Table S6). Furthermore, when this analysis was extended to the whole transcriptome, cumulative fraction analysis revealed that gene transcripts harboring downregulated m5C sites had an obvious higher proportion of increased gene expression (relative to the control group) compared with those containing upregulated m5C sites (Fig. 3c). Collectively, these results indicate an overall negative correlation between m5C modification and gene expression, suggesting that m5C modification may predominantly act as a negative regulator of gene expression in tomato fruits during B. cinerea infection.
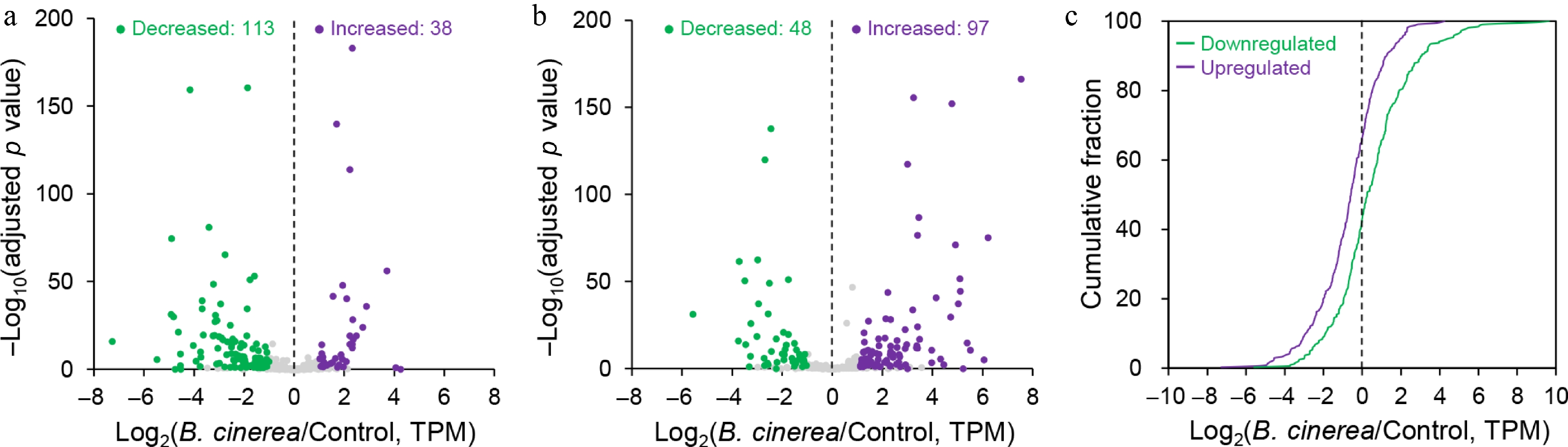
Figure 3.
m5C methylation is globally negatively correlated with transcript abundance. (a) and (b) Volcano plots displaying the expression ratios of transcripts containing upregulated and downregulated m5C sites in B. cinerea-infected tomato fruits compared with the control group, respectively. Transcripts with significantly increased and decreased mRNA levels (log2(fold change) ≥ 1; adjusted p-value < 0.05) in B. cinerea-infected tomato fruits are highlighted in purple and green, respectively. (c) Cumulative distribution of gene expression changes in transcripts harboring upregulated and downregulated m5C sites in B. cinerea-infected tomato fruits.
Ethylene signaling pathway genes exhibit differential m5C modification and transcript level during B. cinerea infection
-
The phytohormone ethylene plays a pivotal role in regulating plant growth and development, as well as in mediating responses to biotic and abiotic stresses[37,38]. Previous studies have demonstrated that the ethylene signaling pathway is involved in defending tomato leaves and fruits against B. cinerea infection by regulating the expression of downstream defense-related genes[21,37,38]. In the present study, we analyzed changes in m5C modification and expression levels of ethylene signaling pathway genes, and identified three such genes that exhibited coordinated changes in m5C modification and transcript abundance. Compared with the control group, one ethylene response factor (ERF) domain-containing protein gene (Solyc03g093610) and one ethylene insensitive 3 (EIN3)-like protein gene (Solyc01g014480) harbored downregulated m5C sites and significantly increased transcript levels in the B. cinerea-infected tomato fruits (Fig. 4). In addition, another ERF domain-containing protein gene (Solyc03g093560) exhibited upregulated m5C modification accompanied by decreased gene expression (Fig. 4). These findings indicate that m5C modification may participate in modulating the ethylene signaling pathway to regulate tomato fruit's resistance against B. cinerea infection.
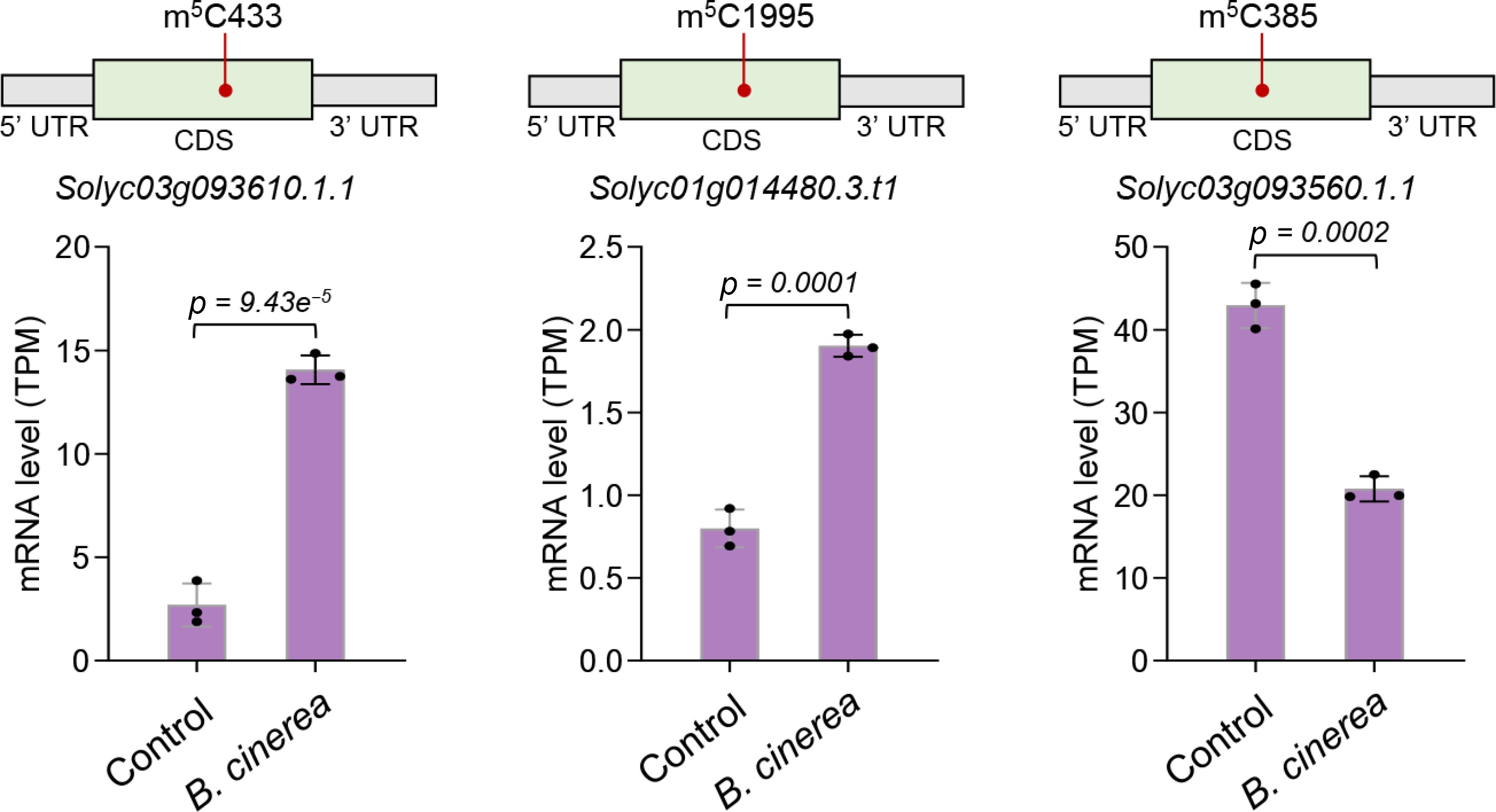
Figure 4.
Ethylene signaling pathway genes exhibited differential m5C methylation and expression abundance under B. cinerea infection. The transcript levels of three ethylene signaling pathway genes, including two ethylene response factor genes (Solyc03g093610 and Solyc01g014480) and one ethylene insensitive 3-like DNA-binding domain-containing protein gene (Solyc03g093560) were determined by RNA-seq analysis. Error bars represent the standard deviation of three independent experiments. Significance analysis was performed using Student's t-test, with p-values indicated. For each gene transcript, the position of differential m5C site is shown.
Defense-related genes exhibit differential m5C modification and transcript levels during B. cinerea infection
-
By integrating differential m5C modification and gene expression analyses, we found that the transcripts of numerous defense-related genes harbor downregulated m5C sites and significantly increased expression abundance in B. cinerea-infected fruits compared with the control group, which further verified the negative regulatory relationship between m5C modification and gene expression. Among these genes, protein kinase genes were the most prominent category. In total, 12 protein kinase genes showed decreased m5C modification and elevated transcript levels (Supplementary Table S6). Three of these protein kinase genes displayed a dramatic increase in expression abundance (at least a 10-fold increase; Fig. 5a), including two protein kinase domain-containing protein genes (Solyc03g115610 and Solyc04g075000) and one nonspecific serine/threonine protein kinase gene (Solyc03g078520). Previous studies have demonstrated that protein kinase genes mediate signal transduction activated by fungal pathogen infection and regulate fruit's resistance to pathogenic fungi[18,39,40]. Thus, these genes may be direct targets of m5C methylation and play a vital role in defending against B. cinerea infection as key defense-related genes. In addition, four redox-related genes, including two peroxidase genes (Solyc02g092580 and Solyc04g071890) involved in ROS metabolism, one procollagen-proline 4-dioxygenase gene (Solyc02g067530), and one MsrB domain-containing protein gene (Solyc02g083360), as well as one toxin-resistant protein gene (Solyc03g118970), exhibited decreased m5C modification and elevated transcript levels (Fig. 5b, c). It is possible that under B. cinerea infection, tomato fruits enhance their resistance by reducing the m5C modification levels of these potentially targeted defense-related genes, thereby promoting their expression.
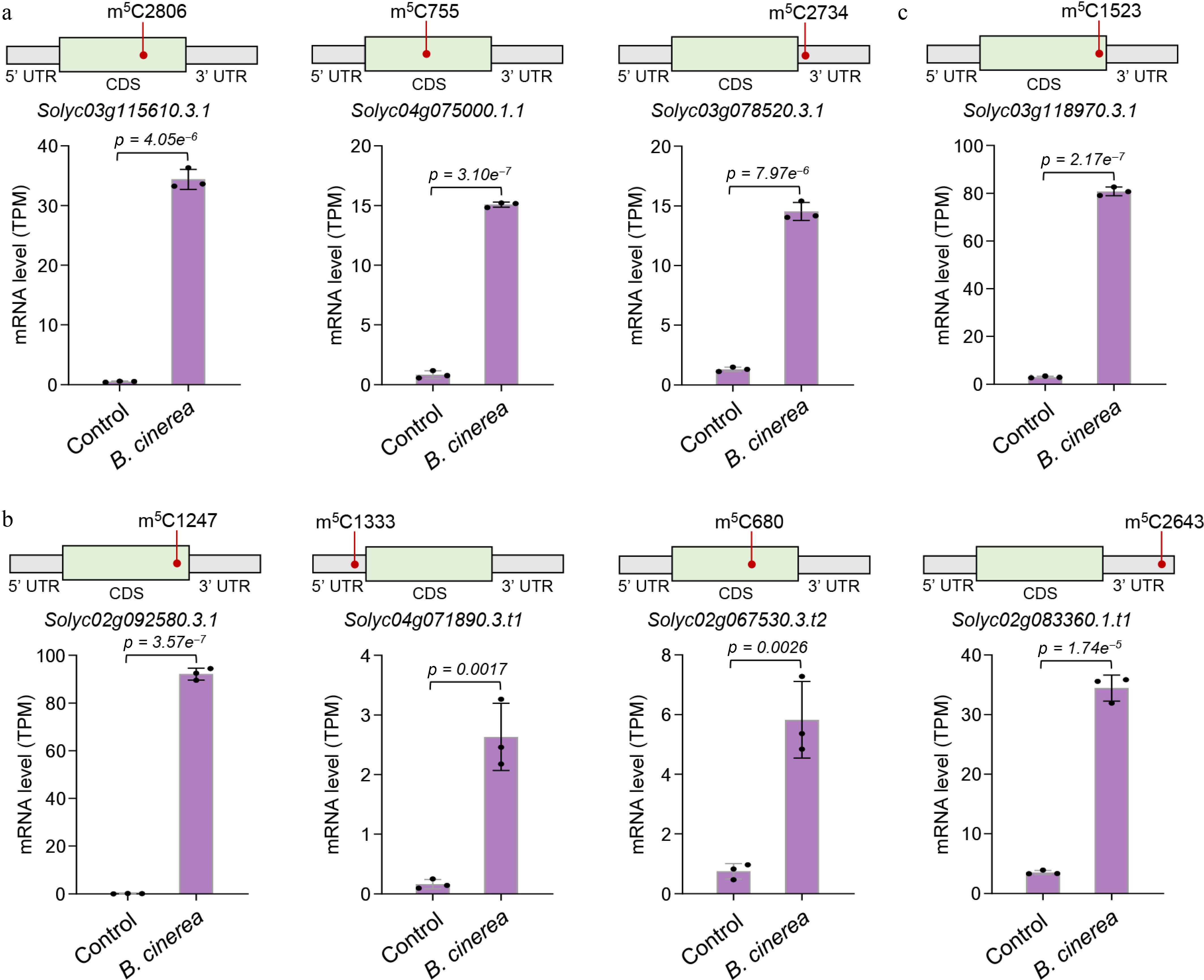
Figure 5.
Defense-related genes exhibited downregulated m5C modification and increased expression abundance under B. cinerea infection. (a) Changes in the transcript levels of three protein kinase genes. (b) Changes in the transcript levels of four redox-related genes. (c) Changes in the transcript levels of one toxin-resistant protein gene. Gene expression levels were determined by RNA-seq analysis. Error bars represent the standard deviation of three independent experiments. Significance analysis was performed using Student's t-test, with p-values indicated. For each gene transcript, the position of downregulated m5C site is shown.
Expression of m5C methyltransferase, demethylase, and reader protein genes in response to B. cinerea infection
-
The dynamic changes in and molecular functions of m5C methylation in vivo depend on the coordinated regulation of m5C methyltransferases, demethylases, and reader proteins[41]. However, in plants, only a limited number of m5C methyltransferase genes have been identified, and the coding genes of m5C demethylases and reader proteins remain largely uncharacterized. In A. thaliana, the reported m5C methyltransferase genes include AtTRDMT, AtTRM4A, and AtTRM4B[14,41]. Their homologous genes in tomato were identified as SlTRDMT, SlTRM4A, and SlTRM4B through BLASTP analysis[42,43]. Gene expression analysis revealed that the mRNA levels of SlTRDMT, SlTRM4A, and SlTRM4B in B. cinerea-infected fruits were not significantly different from those in the control group (Fig. 6a), indicating that these three putative m5C methyltransferase genes do not respond significantly to B. cinerea infection at the transcriptional level.
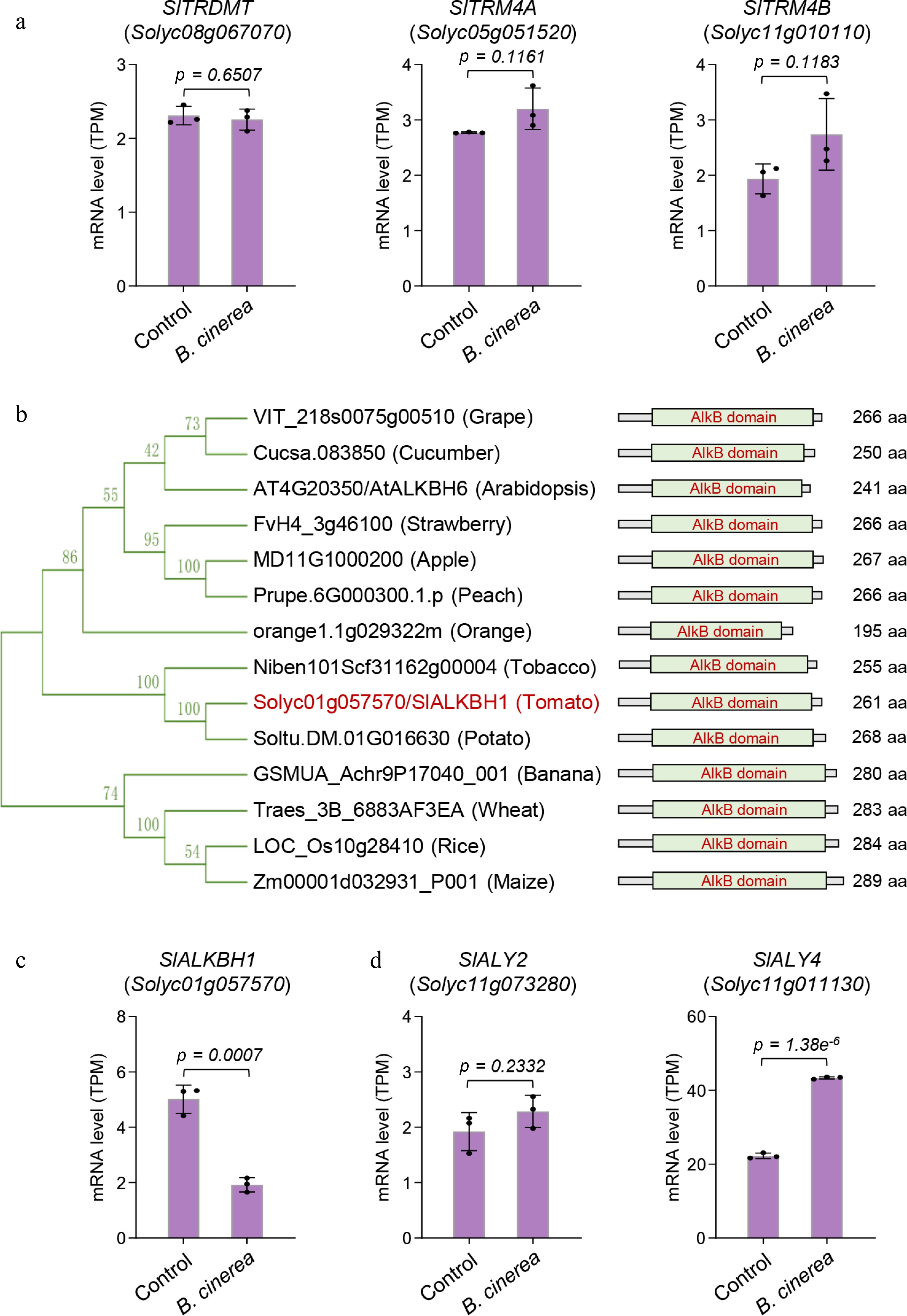
Figure 6.
Changes in the transcript levels of putative tomato m5C methyltransferase, demethylase, and reader protein genes. (a) The transcript levels of three putative m5C methyltransferase genes (SlTRDMT, SlTRM4A, and SlTRM4B) did not significantly alter upon B. cinerea infection. (b) Phylogenetic analysis of the putative m5C demethylase gene among various plant species. The phylogenetic tree was generated using MEGA 5.2 software. Bootstrap values from 1,000 replications are shown for each branch. aa, amino acid. (c) The transcript level of the putative m5C demethylase gene SlALKBH1 significantly decreased upon B. cinerea infection. (d) The transcript level of the putative m5C reader protein gene SlALY4, but not that of the SlALY2, significantly increased upon B. cinerea infection. Error bars represent the standard deviation of three independent experiments. Significance analysis was performed using Student's t-test, with p-values indicated.
In A. thaliana, an ALKB domain-containing protein encoded by AtALKBH6 has been shown to directly bind to m5C-modified RNAs, thereby qualifying as a putative m5C demethylase[10]. Given the amino acid sequence of AtALKBH6, we searched for its homologous proteins across multiple plant species, and identified a ALKB domain-containing protein with the ID Solyc01g057570 in tomato, which we previously named SlALKBH1 and characterized as a putative m6A demethylase[44]. Phylogenetic analysis demonstrated that SlALKBH1exhibits high similarity to those from other Solanaceous crops including potato (Solanum tuberosum) and tobacco (Nicotiana benthamiana) (Fig. 6b). Furthermore, the expression level of SlALKBH1 was significantly decreased under B. cinerea infection (Fig. 6c), implying that it may play a critical regulatory role in mediating tomato's resistance to B. cinerea through m5C demethylation. Additionally, on the basis of the amino acid sequences of the reported m5C reader proteins ALY2 and ALY4 in A. thaliana[11,12], we identified two putative tomato m5C reader proteins via sequence homology analysis, designated SlALY2 and SlALY4. Gene expression analysis showed that SlALY4 was significantly upregulated upon B. cinerea infection, whereas SlALY2 was expressed at low and stable levels (Fig. 6d), suggesting that SlALY4 may be functionally important for the defense response of tomato fruits to B. cinerea.
-
m5C methylation is a conserved epigenetic modification present in various RNA species, playing crucial roles in regulating gene expression and RNA metabolism[45,46]. In this study, we used Nanopore DRS to profile the m5C methylomes of tomato fruits in response to B. cinerea infection for the first time, thereby revealing the dynamic changes in m5C modification and its regulatory roles in tomato fruit's reaction to B. cinerea. Our findings provide new insights into the RNA epigenetic mechanisms underlying plant–pathogen interactions, particularly highlighting the involvement of m5C methylation in mediating the defense responses of postharvest tomato fruits.
The distribution of m5C sites across mRNA regions is closely associated with their regulatory functions. For instance, the m5C modification localized in the 3' UTR and stop codon regions could coordinate with m6A methylation to regulate translation efficiency and mRNA stability[47]. In the present study, m5C methylome profiling revealed that the m5C sites in tomato fruits are preferentially distributed within the CDS and 3' UTR, with only a small proportion located in the 5' UTR (Fig. 1c, d). This distribution pattern is consistent with that reported in A. thaliana seedlings and Ailsa Craig tomato fruits[8,47], as revealed by bisulfite RNA sequencing (bsRNA-seq) and m5C-RNA immunoprecipitation sequencing (RIP-seq) analysis, respectively, but differs from that in tomato leaves as revealed by m5C-RIP-seq analysis, where m5C is mainly distributed in the CDS region[42]. The low proportion of m5C sites in the 5' UTR may be attributed to the short length of the 5' UTR and the limited number of modifiable cytosine residues. Notably, B. cinerea infection did not alter the distribution pattern of m5C sites, which is distinct from the effect of tomato spotted wilt virus in tomato leaves, whose infection induces significant alterations in the distribution preference of m5C peaks[42]. This observation indicates that the regional distribution of m5C modification in tomato fruits is stable and not affected by B. cinerea infection, which may be associated with the fundamental regulatory roles of m5C modification in RNA metabolism.
Differential m5C modification analysis revealed that B. cinerea infection induces specific changes in the m5C modification levels of tomato fruit mRNAs, with 354 upregulated and 341 downregulated m5C sites (Fig. 2a). The similar number of increased and decreased m5C sites suggests that B. cinerea infection does not cause an overall change in m5C modification levels, but rather elicits site-specific local changes. The correlation between m5C modification and gene expression is a key issue for understanding the regulatory function of m5C methylation. In this study, we identified a negative correlation between m5C modification levels and gene expression levels (Fig. 3a–c). This finding is inconsistent with previous studies in A. thaliana roots and tomato leaves, where m5C modification was positively correlated with transcript levels, as mutation of the m5C methyltransferase gene AtTRM4B and SlTRM4B led to reduced m5C modification and decreased expression of m5C-targeted genes[8,14,42]. This discrepancy implies that the regulatory role of m5C modification in gene expression may be organ-specific in plants. In fact, the causal mechanism by which m5C modification negatively regulates gene expression remains largely uncharacterized. Previous studies have demonstrated that m6A modification can recruit the CCR4-NOT RNA degradation complex to specifically mediate the decay of target mRNAs within cytoplasmic P-bodies[48]. In addition, m6A triggers ribosome stalling and collision, induces premature termination of translation, and promotes mRNA degradation, thereby reducing overall mRNA stability and translational efficiency[49]. Whether m5C represses gene expression through a similar mechanism or distinct regulatory pathways warrants further investigation.
GO enrichment analysis demonstrated that genes harboring differential m5C sites were significantly enriched in multiple biological processes, including RNA metabolism and responses to external stimuli (Fig. 2c, d). The enrichment of genes associated with the response to external stimuli highlights the involvement of m5C modification in the defense response of tomato fruits against B. cinerea infection. Specifically, we found that several defense-related genes, encompassing redox-related genes, toxin-resistant protein genes, and protein kinase genes, exhibited reduced m5C modification and elevated expression levels following B. cinerea infection (Fig. 5a–c). Previous studies have demonstrated that ROS and reactive nitrogen species act as both cytotoxic agents and key signaling molecules in plants' responses to environmental cues[50]. Redox-related genes targeted by m5C modification include multiple peroxidase genes responsible for ROS scavenging. This process is critical for plants' defense against pathogen invasion by alleviating oxidative damage[51−53]. Notably, A. thaliana peroxidases PRX33 and PRX34 are essential for H2O2 accumulation during defense responses against pathogens[50,54]. Accordingly, we propose that m5C may modulate tomato fruit's resistance to B. cinerea by regulating H2O2 content. Toxin-resistant protein genes are responsible for the degradation and clearance of mycotoxins produced by B. cinerea[55], whereas protein kinase genes can sense pathogen-associated molecular patterns (PAMPs) and trigger downstream defense signaling cascades[56]. The increased expression of these genes is likely to enhance the ability of tomato fruits to resist B. cinerea infection.
In addition, our results revealed that m5C modification potentially targets and modulates the expression of ethylene signaling pathway genes, including those encoding ethylene response factors and EIN3-like protein genes (Fig. 4). Previous studies have demonstrated that the ethylene signaling pathway contributes to the defense of tomato leaves and fruits against B. cinerea infection by regulating the transcription of downstream defense-related genes[24,38,41]. To our knowledge, this study is the first to document that m5C modification is involved in the regulation of the ethylene signaling pathway in B. cinerea-infected tomato fruits, thereby expanding our understanding of the regulatory mechanisms underlying the ethylene-mediated defense response.
Notably, no significant differentially methylated m5C sites were detected in transcripts of the jasmonic acid (JA) signaling pathway, an indispensable hormonal cascade regulating tomato's resistance to B. cinerea. Additionally, auxins have been implicated in activating defense mechanisms against pathogens by reinforcing the cell walls' integrity and are widely recognized as crucial hormones for plants' defense responses[57]. However, no expressed auxin-related gene transcripts (TPM ≥ 1) exhibited significant differential m5C modification during B. cinerea infection (Supplementary Tables S5 and S6). These results indicate that transcripts associated with the JA and auxin signaling pathways may not be direct targets of m5C modification in tomato fruits upon B. cinerea challenge. It would be intriguing to further explore whether these hormone-related transcripts are regulated by other RNA modifications, such as m6A and m1A, during tomato fruit's defense against fungal infection, which warrants further investigation.
The dynamics of m5C modification is tightly orchestrated by three classes of key regulators: Methyltransferases, demethylases, and reader proteins. In A. thaliana, AtTRDMT, AtTRM4A, and AtTRM4B have been characterized as functional m5C methyltransferases[8,17,44]. In the present study, we identified their homologous genes in tomato, designated as SlTRDMT, SlTRM4A, and SlTRM4B. However, B. cinerea infection did not significantly alter the transcript levels of these three genes, suggesting that these methyltransferases may not participate in the regulation of m5C modification during B. cinerea infection. Alternatively, their regulatory effects might be mediated at the post-transcriptional or post-translational level, which requires further investigation.
It should be noted that this study has several inherent limitations. First, our analysis of the m5C methylome was restricted to a single time point (48 h postinoculation), leaving the dynamic changes in the m5C methylome across different stages of B. cinerea infection largely unexplored. Second, the regulatory mechanisms through which m5C modification modulates gene expression, such as its potential effects on mRNA stability or translation efficiency, have not been validated using in vitro experimental approaches Third, how ethylene-mediated hormone signaling is regulated by m5C methylation and participates in the defense response remains to be further investigated. Fourth, the specific roles of m5C methyltransferases, demethylases, and reader proteins in mediating tomato fruit's resistance to B. cinerea infection remain elusive. In future research, we will focus on addressing these issues to further delineate the regulatory mechanisms by which m5C modification governs tomato fruit's defense responses against B. cinerea, which may provide novel targets for improving tomato fruit's postharvest resistance to gray mold disease.
-
In this study, we profiled the m5C methylomes of B. cinerea-infected and mock-inoculated tomato fruits via Nanopore DRS. Our results demonstrated that m5C modification in tomato fruit's mRNAs exhibits a specific nucleotide preference and is predominantly distributed in the CDS region and 3' UTR. Notably, B. cinerea infection induces site-specific changes in m5C modification but does not alter the global m5C modification level. Globally, B. cinerea-triggered m5C modification negatively regulates gene expression, and genes with differential m5C modifications are involved in multiple biological processes, including RNA metabolism and responses to external stimuli. Importantly, a large number of ethylene signaling pathway and defense-related genes are directly targeted by m5C modification, implying its critical involvement in mediating tomato fruit's resistance to B. cinerea. Additionally, the putative tomato m5C demethylase gene SlALKBH1 and the reader protein gene SlALY4, but not the three m5C methyltransferase genes, respond significantly to B. cinerea infection at the transcriptional level. Collectively, this study provides the first comprehensive m5C methylome profile of tomato fruits during B. cinerea infection, advancing our understanding of RNA-based epigenetic regulation in plant–pathogen interactions and offering novel targets for improving postharvest resistance in tomato fruit.
We thank Professor Shiping Tian from the Institute of Botany, Chinese Academy of Sciences, for providing the B. cinerea strain B05.10. This work was supported by the Fundamental Research Funds for the Central Universities of Chongqing University (Grant No. 2025CDJ-IAISYB-065) and the National Natural Science Foundation of China (Grant No. 32472403).
-
The authors confirm their contributions to this study as follows: conceived and designed the experiments: Zhou L, Gao Y; performed the experiment and analyzed the data: Cai X, Zhou L; provided critical discussions: Qin G, Cassan-Wang H, Cai J, Huang B, Cheng Y; wrote the manuscript: Zhou L; contributed to the writing: Gao Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Information of Nanopore direct RNA sequencing in both the B. cinerea-infected and the control tomato fruits. Rep, replicate.
- Supplementary Table S2 Transcripts containing upregulated m5C sites in B. cinerea-infected tomato fruits compared to the control group.
- Supplementary Table S3 Transcripts containing downregulated m5C sites in B. cinerea-infected tomato fruits compared to the control group.
- Supplementary Table S4 Differential expressed gene transcripts in the B. cinerea-infected tomato fruits compared to the control group.
- Supplementary Table S5 The expression abundance of transcripts containing upregulated m5C sites in B. cinerea-infected tomato fruits compared to the control group.
- Supplementary Table S6 The expression abundance of transcripts containing downregulated m5C sites in B. cinerea-infected tomato fruits compared to the control group.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cai X, Huang B, Cheng Y, Cai J, Cassan-Wang H, et al. 2026. Nanopore direct RNA sequencing reveals the m5C methylomes of tomato fruits in response to Botrytis cinerea. Plant Hormones 2: e011 doi: 10.48130/ph-0026-0010 

Nanopore direct RNA sequencing reveals the m5C methylomes of tomato fruits in response to Botrytis cinerea
- Received: 20 March 2026
- Revised: 20 April 2026
- Accepted: 24 April 2026
- Published online: 14 May 2026
Abstract: RNA methylation, particularly 5-methylcytosine (m5C) modification, has emerged as a pivotal regulatory layer governing plant development and stress responses. However, the role of m5C in mediating the resistance of tomato (Solanum lycopersicum) fruit to the fungal pathogen Botrytis cinerea remains unclear. In this study, we utilized Nanopore direct RNA sequencing to generate the transcriptome-wide m5C methylomes in B. cinerea-infected tomato fruits. Our results revealed that m5C sites are preferentially enriched in coding sequences (CDSs) and 3' untranslated regions (3' UTRs). Notably, B. cinerea infection induced site-specific m5C changes without altering the global modification level. Integrated omics analysis identified a transcriptome-wide negative correlation between m5C modification and gene expression. Importantly, a large number of ethylene signaling pathway and defense-related genes exhibited reduced m5C levels, accompanied by significantly increased transcript abundance, implying that m5C may act as a novel regulator of fruit's resistance to B. cinerea by coordinating hormone signaling and defense response pathways. Furthermore, the putative m5C demethylase gene SlALKBH1 and the reader protein gene SlALY4 were transcriptionally downregulated and upregulated by B. cinerea infection, respectively. Collectively, this study establishes the first comprehensive m5C methylomes of tomato fruits upon B. cinerea infection and provides new insights into RNA-based epigenetic regulation of hormone signaling during plant–pathogen interactions.
-
Key words:
- Tomato fruit /
- m5C /
- Ethylene /
- Defense-related gene /
- Botrytis cinerea