









-
Fungi play crucial roles in nutrient cycling and ecosystem functioning and act as symbionts, pathogens, and decomposers[1−4]. Among them, microfungi comprise a broad range of saprobic, pathogenic, and endophytic taxa that are particularly abundant in terrestrial ecosystems, especially within decaying plant material[5]. Despite their ecological importance and potential biotechnological applications, the diversity of microfungi in tropical regions remains significantly underexplored[6,7]. Despite covering a relatively small area of 6,524,540 hectares, Sri Lanka ranks among the most biologically diverse countries in Asia, with a rich biodiversity resulting from its varied climate and topography across a wide range of ecosystems, including tropical rainforests, montane and sub-montane regions, and dry zones[8,9]. Nevertheless, fungal biodiversity studies in the country have historically been limited, and many microfungal groups remain poorly documented[10,11]. Recent studies have begun to reveal the remarkable diversity of the islandwide mycobiota; however, major gaps remain in the taxonomy, distribution, and ecological functions of many microfungal groups[12−14].
Microfungi are integral to ecosystem functioning due to their highly specialized roles in the decomposition of plant-derived substrates. As primary agents of lignocellulose decay, they secrete a diverse suite of extracellular enzymes, including cellulases, hemicellulases, laccases, and lignin peroxidases, that enable the stepwise breakdown of complex plant polymers and the subsequent mobilization of carbon and nutrients into the soil matrix[2,5,15−18]. Saprobic microfungi exhibit strong substrate specificity, often initiating colonization through hyphal penetration of senescent tissues, where they modify cell-wall architecture and accelerate litter turnover. Increasing evidence also highlights the dual ecological strategies of endophytic fungi: although asymptomatic during the host's lifespan, many taxa undergo a functional shift to saprotrophy following host senescence, rapidly exploiting the nutrient-rich tissues they previously inhabited[19−21]. This endophyte-to-saprobe transition underscores their importance as early colonizers in the decomposition continuum and as key regulators of nutrient fluxes within terrestrial ecosystems.
During a mycological survey conducted within urbanized ecosystems in Sri Lanka, several saprobic fungi were isolated from decaying leaves and plant debris in the premises of the University of Sri Jayewardenepura. Morphological characteristics combined with multi-gene phylogenetic analyses revealed a previously undescribed species, along with two other microfungal species recorded for the first time in Sri Lanka. The present findings contribute to the growing body of knowledge on tropical microfungi, and underscore the importance of local urban ecosystems as reservoirs of undocumented fungal diversity.
-
Woody and plant debris were collected from the University of Sri Jayewardenepura premises (6°51'06" N 79°54'31" E, 25 masl) in November 2024. The collected specimens were placed in eco-friendly high-pressure polyethylene (LDPE) zip-lock bags, and brought to the Genetics and Molecular Biology laboratory, Faculty of Applied Sciences, University of Sri Jayewardenepura, Sri Lanka. The specimens were initially examined using a stereomicroscope (OPTIKA SZO-T, Italy), and all the colony characteristics, including position on the host surface, shape, and color, were recorded. Following the examination, micro-morphological characteristics were photographed using a Nikon ECLIPSE Ni-U upright compound microscope (Nikon, Japan), equipped with an Axiocam 506 color digital camera (ZEISS, Hanover, Germany). All measurements were obtained using the Tarosoft® Image Framework application, while photo plates were prepared with Adobe Photoshop CS3 Extended version 10.0 (Adobe Systems, USA).
The method described by Senanayake et al.[22] was used for the single conidial isolation. After germination, conidia were transferred to potato dextrose agar (PDA) and incubated at 25 °C overnight. Pure cultures were obtained after a subsequent subculturing process. Culture characteristic features were observed and recorded. Type specimens and other dried specimens were prepared and deposited in the Herbarium of the University of Sri Jayewardenepura (USJ-H), Sri Lanka. Isolated cultures were deposited in the fungi culture collection of the University of Sri Jayewardenepura (USJCC) and the culture collection of the Genetics and Molecular Biology Unit, Faculty of Applied Sciences, University of Sri Jayewardenepura (GMBUCC). Nomenclatural data for fungal novelty were deposited in MycoBank (
www.mycobank.org ).Extraction of DNA, PCR amplification, and sequencing
-
Genomic DNA was extracted using fungal strains grown on PDA for 3–4 weeks at 25 °C. Axenic mycelium (80–100 mg) was harvested from actively growing cultures, and genomic DNA was extracted using the Biospin Fungal DNA Extraction Kit (Bioer Technology Co. Ltd., Hangzhou, China) according to the manufacturer's instructions. The DNA products were stored at 4 °C, with replicate samples preserved at −20 °C to ensure long-term stability. Genomic regions amplified included: the internal transcribed spacer (ITS1–5.8S–ITS2), the nuclear ribosomal 28S large subunit rRNA (LSU), the nuclear ribosomal 18S small subunit rRNA (SSU), the partial translation elongation factor 1-alpha (tef1-α), the partial sequence of the beta-tubulin gene (tub2), and the partial second largest subunit of RNA polymerase II (rpb2). The ITS region was amplified using ITS4 and ITS5[23], LSU with LR0R and LR5[24], and SSU with NS1 and NS4[23]. The tef1-α gene was amplified using EF1-983F and EF1-2218R[25], tub2 with Bt2a and Bt2b[26], and rpb2 with fRPB2-5F and fRPB2-7cR[27]. The PCR reaction was performed in a total volume of 25 µl, consisting of 9.5 µl sterilized distilled water, 12.5 µl of 2× Power Taq PCR MasterMix (containing 0.1 U/µl Taq DNA polymerase, and 500 µM of each dNTP: dATP, dCTP, dGTP, dTTP; Bioteke Corporation (Wuxi) Co., Ltd.), 1 µl each of forward and reverse primers (10 pM stock), and 1–2 µl of DNA template. Thermal cycle programs for LSU, SSU, ITS, tef1-α, tub2, and rpb2 genes were followed as mentioned in Thambugala et al.[28]. PCR products were visualized on 1% agarose gels stained with ethidium bromide under UV illumination. Subsequently, PCR product purification and sequencing were performed at Genelabs Medical (Pvt) Ltd, Sri Lanka.
Phylogenetic analyses
-
The resulting sequences (forward and reverse) were initially checked using BioEdit v 7.0.5.3[29] and assembled with SeqMan v. 7.0.0 (DNASTAR, Madison, WI, USA). The nucleotide BLAST search (
https://blast.ncbi.nlm.nih.gov ) was carried out to determine the strains that have high similarities. The reference sequences used in the analyses were obtained from recent publications[30,31] (Supplementary Tables S1−S3). Single-gene datasets were aligned with Bioedit 7.1.3.0[29], and the consensus sequences were further improved with MUSCLE implemented in MEGA11[32]. Alignments were manually checked and adjusted when necessary.Phylogenetic analyses using both maximum likelihood (ML) and Bayesian inference (BI) methods were carried out using concatenated alignments. The best-fit substitution models were chosen using MrModeltest v2.3[33] based on the Akaike Information Criterion (AIC). Maximum Likelihood analysis was conducted with RAxML-HPC2 v8.2.8[34] via the CIPRES Science Gateway[35], employing the GTR + I + G nucleotide substitution model with 1,000 bootstrap replicates. Bayesian Inference was performed in MrBayes v3.2.1[36] with six simultaneous Markov chains run for 1, 5, and 3 million generations, sampling every 100th tree for Paradictyoarthrinium, Torula, and Apiospora, respectively. The first 20% of sampled trees were discarded as burn-in, and the remaining 80% were used to estimate posterior probabilities and produce consensus trees. The resulting phylogenetic trees were visualized and preliminarily edited using MEGA11[32], with final graphical adjustments and layout formatting completed in Microsoft PowerPoint 2016.
-
The phylogenetic trees in this study were constructed using maximum likelihood (ML), and Bayesian inference (BI). The best maximum likelihood trees, with support values from both analyses, are shown at each corresponding node in Figs. 1−3.
Phylogenetic analyses of Paradictyoarthrinium
-
The concatenated LSU (1–867), ITS (868–1,437), and rpb2 (1,438–2,499) sequence datasets were analyzed to identify the phylogenetic position of our newly generated sequence within Paradictyoarthrinium. The dataset consisted of 20 sequences, with Nigrograna obliqua (CBS141477) as the outgroup taxon. The ML and BI analyses yielded similar tree topologies, and therefore, only the best-scoring ML tree is presented (Fig. 1) with the likelihood value of −8,213.564 and the following model parameters: Estimated base frequencies were A = 0.250, C = 0.250, G = 0.250, and T = 0.250; substitution rates were AC = 1.00000, AG = 4.66643, AT = 1.000000, CG = 1.000000, CT = 8.25386, and GT = 1.000000; gamma distribution shape parameter: α = 0.429. The BI analysis ran for one million generations, with the average standard deviation of split frequencies reaching below 0.01 (0.008125). This analysis generated 10,000 trees, from which 8,000 were sampled after discarding 20% as burn-in. The alignment contained a total of 714 distinct patterns. All Paradictyoarthrinium strains formed a strongly supported clade (100% ML, 1.00 PP) sister to Melanomma and Nigrograna. The new isolate USJCC-0274 clustered within the P. diffractum strains (MFLUCC 12-0557, MFLUCC 24-0257, KUMCC 19-0111, UESTCC 24.0187, and MFLUCC 13-0466) with strong statistical support (99% ML/1.00 PP) (Fig. 1).

Figure 1.
Maximum likelihood (ML) tree of combined LSU, ITS, and rpb2 sequences from Paradictyoarthrinium species. Xenomassariosphaeria clematidis MFLUCC 14-0923 and X. rosae MFLU 15-0179 were used as the outgroup. Maximum likelihood bootstrap support values (≥ 50%) and Bayesian posterior probability (≥ 0.90) are indicated at nodes. The newly generated strain is indicated in red, while type strains are shown in bold.
Phylogenetic analyses of Torula
-
The concatenated ITS (1–583), LSU (584–1,445), SSU (1,446–2,471), and tef1-α (2,471–3,367) sequence datasets were analyzed to infer the phylogenetic position of our newly generated sequences within Torula. The dataset consisted of 47 sequences, with Cylindrotorula indica (NFCCI 4836 and NFCCI 4837) as the outgroup taxa. The RAxML and Bayesian analyses produced similar tree topologies, and therefore, only the best-scoring ML tree is presented (Fig. 2) with the likelihood value of -10,479.082 and the following model parameters: Estimated base frequencies were A = 0.250, C = 0.250, G = 0.250, and T = 0.250; substitution rates were AC = 1.68626, AG = 2.49231, AT = 1.68626, CG = 1.000000, CT = 7.12644, and GT = 1.000000; gamma distribution shape parameter: α = 0.557. The BI analysis ran for 5 million generations, with the average standard deviation of split frequencies reaching below 0.01 (0.006276). This analysis generated 50,000 trees, from which 10,000 were sampled after discarding 20% as burn-in. The alignment contained a total of 685 distinct patterns. Torula strains formed a statistically well-supported clade (100% ML/1.00 PP) with five distinct clusters. The newly generated sequences (USJCC-0275 and USJCC-0276) clustered basal to the strains of T. chromolaenae and T. longan with a robust statistical support (100% ML/1.00 PP) (Fig. 2).
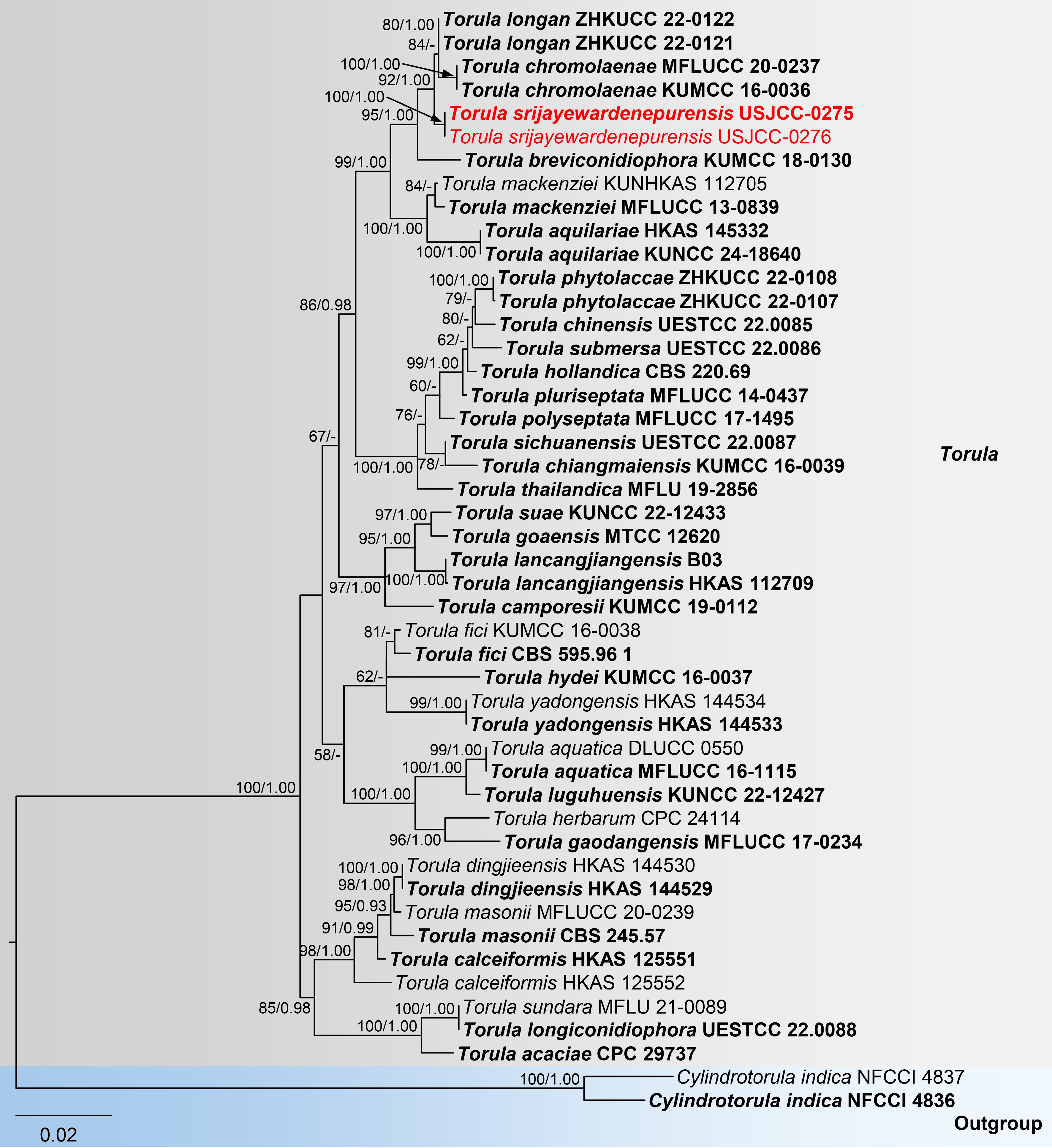
Figure 2.
Maximum likelihood (ML) tree of concatenated ITS, LSU, SSU, and tef1-α sequences from Torula species. Cylindrotorula indica (NFCCI 4836 and NFCCI 4837) was used as the outgroup. Maximum likelihood bootstrap support values (≥ 50%) and Bayesian posterior probability (≥ 0.90) are indicated at nodes. Newly generated strains in this study are highlighted in red, while type strains are shown in bold.
Phylogenetic analyses of Apiospora
-
The concatenated ITS (1–643), LSU (644–1,495), tub2 (1,496–2,082), and tef1-α (2,083–3,137) sequence dataset was analyzed to determine the phylogenetic position of our newly generated sequence within Apiospora. The dataset included 72 sequences, including Arthrinium caricicola (CBS 145127) as the outgroup. The RAxML and Bayesian analyses yielded a similar tree topology, and therefore, only the best-scoring ML tree is presented (Fig. 3) with the likelihood value of −20144.9873 and the following model parameters: Estimated base frequencies were A = 0.250, C = 0.250, G = 0.250, and T = 0.250; substitution rates were AC = 1.0000, AG = 2.5276, AT = 1.0000, CG = 1.0000, CT = 4.1989, and GT = 1.000000; gamma distribution shape parameter: α = 0.732. The BI analysis ran for 3 million generations, with the average standard deviation of split frequencies reaching below 0.01 (0.009897). This analysis generated 30,000 trees, from which 6,000 were sampled after discarding 20% as burn-in. The alignment contained a total of 1,645 distinct patterns. Apiospora strains formed a monophyletic group with seven clusters. The newly generated sequence (USJCC-0277) clustered with the type of A. pseudorasikravindrae (KUMCC 20–0208), forming a sister relationship with A. acutiapica (KUMCC 20-0210) in a well-supported clade (89% ML; Fig. 3).

Figure 3.
Maximum likelihood (ML) tree of concatenated ITS, LSU, tef1-α, and tub2 sequences from Apiospora species. Arthrinium caricicola was used as the outgroup. Maximum likelihood bootstrap support values (≥ 50%) and Bayesian posterior probability (≥ 0.90) are indicated at nodes. The newly generated strain is shown in red, while type strains are shown in bold.
Taxonomy
-
Dothideomycetes O.E. Erikss. & Winka, Myconet 1(1): 5 (1997)
Pleosporales Luttr. ex M.E. Barr, Prodr. Cl. Loculoasc. (Amherst): 67 (1987)
Paradictyoarthriniaceae Doilom, J.K. Liu & K.D. Hyde, Fungal Diversity 72: 133 (2015)
Paradictyoarthrinium Matsush., Matsush. Mycol. Mem. 9: 18 (1996)
Notes: Matsushima[37] introduced Paradictyoarthrinium in Paradictyoarthriniaceae (Pleosporales) as a monotypic genus with P. diffractum. Later studies with multigene phylogeny using LSU, SSU, tef1-α, and rpb2 reconfirmed the placement of Paradictyoarthrinium[38,39]. Paradictyoarthrinium species are characterized by gregarious, black, powdery colonies bearing macronematous conidiophores, which produce asymmetrically and irregularly dictyoseptate, muriform, subglobose to ellipsoidal, dark brown conidia. Currently, the genus Paradictyoarthrinium comprises five species: P. aquatica, P. diffractum, P. hydei, P. salsipaludicola, and P. tectonicola, and they are recorded in different countries, including China, India, South Africa, and Thailand[30]. The members of Paradictyoarthrinium are primarily saprophytes occurring on decaying wood in terrestrial, freshwater, and marine environments[40].
Paradictyoarthrinium diffractum Matsush., Matsushima Mycol. Mem. 9: 18 (1996) Fig. 4

Figure 4.
Paradictyoarthrinium diffractum (USJ-H-255). (a)–(c) Colonies on the host substrate. (d)–(f) Conidiogenous cells and developing conidia. (g)–(m). Conidia. (n) Germinated conidium. (o) Front, and (p) the reverse views of the culture on PDA media. (q) Colonies on PDA media. Scale bars: (d)–(n) = 15 μm.
MycoBank number: MB 415849
Saprobic on a piece of decaying wood. Sexual morph: Undetermined. Asexual morph: Colonies on the woody substrates are scattered, superficial, effuse, gregarious, blackish, and powdery. Conidiophores 15–30 × 1–3 µm (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA are fast-growing, reaching 5–6 cm in diameter after two weeks at 20–25 °C, colonies sparse, circular, raised, surface slightly rough, with an irregular margin, cottony to fairly fluffy with sparse aspects, colony from above, white to cream at the centre, pale brown to dark brown at the margin; from below, dark brown to black at the centre, pale brown at the margin, mycelium white to cream with tufting.
Material examined: Sri Lanka, Western Province, Colombo District, University of Sri Jayewardenepura premises, on a piece of decaying wood, 1 November 2024, Kasun M. Thambugala, KTSL009–2024 (USJ-H 255), culture USJCC-0274 (GMBUCC 24–019).
Notes: The new isolate USJCC-0274 clustered within the Paradictyoarthrinium diffractum clade (99% ML, 1.00 PP) in the combined LSU, ITS, and rpb2 phylogeny. The ITS and rpb2 sequences of isolate USJCC-0274 show 100% similarity to those of P. diffractum isolates GMBCC 2212, KUMCC 21-0336, and MFLUCC 24-0152, with no gaps. The dark brown to black, thick-walled conidia, which arise from branched or unbranched conidiophores and occur singly or occasionally in short chains, are unevenly dictyoseptate, subglobose to broadly ellipsoidal, sometimes irregular or lobed, and verrucose. These features agree with descriptions of P. diffractum in earlier studies[40,41]. P. diffractum has been recorded as saprobic on dead twigs in South Africa[37], dead stems of Tectona grandis in Thailand[42], dead spathes of Cocos nucifera in India[38], dead branches of Pinus taeda in China[40], and decaying wood of Delonix regia in Thailand[41]. Based on morphological and molecular evidence, the isolate is identified as P. diffractum. This is the first report of this species from Sri Lanka.
Torulaceae Corda, Deutschlands Flora, Abt. III. Die Pilze Deutschlands 2: 71 (1829)
Torula Pers., Ann. Bot. (Usteri) 15: 25 (1795)
Persoon[43] first introduced Torula in Torulaceae with T. herbarum as the type species. Later studies with multi-gene phylogeny, Crous et al.[44] confirmed the phylogenetic placement of Torula within Torulaceae (within Pleosporales). The Torula shows a wide host range and is present in both terrestrial and aquatic habitats[45−47]. Torula is characterized by cupulate conidiogenous cells in branched chains, subglobose, verrucose, septate conidia[44,45,48]. Currently, the phylogenetic placement of Torula was well established through multigene phylogeny of ITS, LSU, SSU, tef1-α, and rpb2[30,49].
Torula srijayewardenepurensis Thambugala & Tennakoon, sp. nov. Fig. 5

Figure 5.
Torula srijayewardenepurensis (USJ-H-252, holotype). (a)–(c) Colonies on the host substrate. (d) Conidiophores with conidiogenous cells. (e)–(g) Conidia. (h) Germinated conidium. (i) Front, and (j) the reverse view of the culture on PDA medium. (k) Appearance of colonies on PDA. Scale bars: (d)–(h) = 5 μm.
MycoBank number: MB 860342
Etymology: The specific epithet srijayewardenepurensis refers to the University of Sri Jayewardenepura, Sri Lanka, the location where the holotype was collected.
Saprobic on a decaying unknown twig. Sexual morph: Undetermined. Asexual morph: Colonies effuse, scattered, dark brown to black, powdery, velutinous, dense to moderately dense. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 2–4 μm long × 3–5 μm wide (
$\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA fast-growing, reaching 3–4 cm diameter after two weeks at 20–25 °C, colonies moderately sparse, flat, circular, with a slightly rough surface, entire edge, and well-defined margin, cottony to moderately fluffy with sparse aspects, colony from above, light brown at the centre, cream to yellowish at the margin; from below, light brown to dark brown at centre, yellowish at the margin, mycelium white to cream with tufting.
Material examined: Sri Lanka, Western Province, Colombo District, University of Sri Jayewardenepura premises, on a decaying unknown twig, 1 November 2024, Kasun M. Thambugala, KTSL008–2024 (USJ-H-252, holotype), ex-type culture USJCC-0275 (GMBUCC 24–016); ibid. KTSL008-2–2024 (USJ-H 253), culture USJCC-0276 (GMBUCC 24–017).
Notes: The multi-gene phylogenetic analyses (ITS, LSU, SSU, and tef1-α sequences) place the two isolates USJCC-0275 and USJCC-0276 as an independent lineage basal to Torula chromolaenae and T. longan, with strong support (92% ML, 1.00 BYPP). Torula breviconidiophora KUMCC 18-0130 forms a basal clade to T. chromolaenae, T. longan, and the lineage formed by the two new isolates. The ITS sequence of USJCC-0275 is similar to that of T. chromolaenae (KUMCC 16-0036; 98%, 4/523 gaps) and T. longan (ZHKUCC 22-0121; 99%, 1/505 gaps). The tef1-α sequence is similar to that of T. chromolaenae (KUMCC 16-0036; 98%, 2/874 gaps) and T. mackenziei (MFLUCC 13-0839; 98%, 2/906 gaps). The two isolates differ from T. chromolaenae and T. longan by having conidiophores reduced to conidiogenous cells and by their shorter conidiogenous cells. The conidia of T. breviconidiophora are medium to dark brown, 1–4-septate, with subhyaline or pale brown apical cells, which also distinguishes it from the new isolates[50]. Based on morphological and molecular evidence, the two isolates are identified as a new species, T. srijayewardenepurensis.
Sordariomycetes O.E. Erikss. & Winka, Myconet 1(1): 10 (1997)
Amphisphaeriales D. Hawksw. & O.E. Erikss., Syst. Ascom. 5(1): 177 (1986)
Apiosporaceae K.D. Hyde, J. Fröhl., Joanne E. Taylor & M.E. Barr, Sydowia 50(1): 23 (1998)
Apiospora Sacc., Atti Soc. Veneto-Trent. Sci. Nat., Padova, Sér. 4 4: 85 (1875)
Notes: Saccardo[51] established Apiospora Sacc. with A. montagnei as the type species. However, the taxonomic placement of Apiospora remained unclear with morphologically closer genera Arthrinium and Neoarthrinium due to the presence of arthrinium-like basauxic conidiogenesis[52]. Crous & Groenewald[53] later synonymized Apiospora with Arthrinium based on the one fungus-one name policy. However, later studies with additional genetic data of type species of Apiospora and Arthrinium, Pintos & Alvarado[54] were able to separate Apiospora and Arthrinium into two distinct genera. Pintos & Alvarado[54] revealed that Arthrinium species have variously shaped conidia and inhabit Cyperaceae and Juncaceae in temperate, cold, and alpine habitats, while most Apiospora species have rounded/lenticular conidia, inhabiting mainly Poaceae together with other host plant families distributed in a wide range of habitats from tropical to subtropical regions. Apiospora montagnei, the epitype for the type species of Apiospora, was proposed by Pintos & Alvarado[55]. Apiospora is distinctly characterized by conidia that are globose, subglobose to ellipsoid, oval, or obovoid in face view, appearing lenticular in side view, and by the presence of basauxic conidiogenous cells[54].
Apiospora pseudorasikravindrae (Senan. & Cheew) Thambugala, comb. nov. Fig. 6

Figure 6.
Apiospora pseudorasikravindrae (USJ-H-254). (a)–(c) Colonies on the host substrate. (d), (e) Conidiogenous cells and developing conidia. (f)–(h) Conidia. (i) Germinated conidium. (j) Front, and (k) the reverse views of the culture on PDA media. Scale bars: (d)–(i) 5 μm.
MycoBank number: MB 558505
≡ Arthrinium pseudorasikravindrae Senan. & Cheew., Frontiers Microbiol. 11 (no. 602773): 11 (2020)
Typus: China, Guangdong Province, Shenzhen City, Futian District, northwest of Futian, Bijiashan Park, on sheath of Bambusa dolichoclada (Poaceae), 23 September 2018, IS, SI 73 (HKAS 107669, holotype), ex-type culture, KUMCC 20-0208; ibid October 15, 2018, IS, SI 73-1 (HKAS 107670, paratype), ex-paratype culture KUMCC 20-0211.
Description of the new collection: Saprobic on clumps of Bambusa vulgaris (Poaceae). Sexual morph: undetermined. Asexual morph: Colonies scattered to aggregated, forming numerous black spots, superficial. Conidiophore mother cells are ampulliform, doliiform to subglobose, hyaline, smooth-walled. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 5–10 × 2–5 μm (
$\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA fast-growing, reaching 5–6 cm diameter after two weeks at 20–25 °C, colonies sparse, circular, slightly raised, with a moderately rough surface, entire edge, and well-defined margin, cottony to fairly fluffy with sparse aspects, colony from above, pale brown at the centre, white at the margin; from below, pale brown at centre, white to cream at the margin, mycelium white to cream with tufting.
Material examined: Sri Lanka, Western Province, Colombo District, University of Sri Jayewardenepura premises, on clumps of Bambusa vulgaris Schrad. ex J.C. Wendl., 1 November 2024, Kasun M. Thambugala, KTSL007–2024 (USJ-H 254), culture USJCC-0277 (GMBUCC 24–018).
Notes: Arthrinium pseudorasikravindrae was described from Bambusa dolichoclada in Guangdong Province, China, by Senanayake et al.[56]. Following Pintos & Alvarado[55], Tian et al.[57] proposed a new combination and transferred the species to Apiospora as A. pseudorasikravindrae, based on combined LSU, ITS, tef1-α, and tub2 sequence analyses. However, this name was invalid because the authors did not provide full details of the basionym reference, as required by ICN Art. 41.5. In this study, we provide a valid combination, formally establishing Apiospora pseudorasikravindrae based on comprehensive morpho-molecular evidence.
One of the new isolates (USJCC-0277) forms a sister lineage to A. pseudorasikravindrae (KUMCC 20-0208) in a well-supported clade (89% ML). Isolate USJCC-0277 shares similar morphology with KUMCC 20-0208, including ampulliform, doliiform to subglobose conidiogenous cells (5–10 × 2–5 μm vs 4–10 × 1.2–5 μm), and lenticular conidia in side view, globose in face view, with a pale longitudinal slit. The conidia are thick-walled, brown to dark brown, finely roughened, with one or two concentric pale rings (4.6–9 × 7–12 μm vs 5–10 × 5.5–11 μm). The tub2 and tef1-α sequences of USJCC-0277 show 100% similarity to those of KUMCC 20-0208 with no gaps. The ITS sequence of KUMCC 20-0208, however, contains ambiguous base calls in conserved regions, which may explain the long branch in the phylogeny. Based on protein-coding gene similarity and morphological agreement, the new isolate is identified as A. pseudorasikravindrae. Apiospora pseudorasikravindrae has previously been reported only from Bambusa dolichoclada in China[56]. In this study, A. pseudorasikravindrae was collected on clumps of Bambusa vulgaris in Sri Lanka, representing a new host and a new geographical record.
-
In this study, a novel microfungal species is introduced, Torula srijayewardenepurensis, and the first record of Apiospora pseudorasikravindrae and Paradictyoarthrinium diffractum from Sri Lanka are documented. The discovery of Torula srijayewardenepurensis sp. nov., and the new records of Apiospora pseudorasikravindrae and Paradictyoarthrinium diffractum not only add to the growing account of Sri Lankan mycobiota but also illustrate the hidden richness of microfungi in urban and anthropogenically influenced environments. The isolation of all three taxa from leaf litter collected within an urban setting underscores the immense potential of even highly modified ecosystems in harboring novel and ecologically significant fungal taxa.
This observation suggests that contrary to the widely held assumption that urban environments are uniformly low in biodiversity, certain urban niches may support a greater diversity of microfungi than expected. Rather than making broad claims, the present results indicate that these habitats may harbour previously undocumented or overlooked fungal taxa. However, more systematic and comprehensive sampling across multiple urban settings would be required to substantiate this pattern more broadly. Given that urbanization is rapidly reshaping tropical landscapes, it is essential to include urban ecosystems in fungal biodiversity assessments.
Torula species have a wide host range and occur in diverse habitats, commonly as saprobes in both aquatic and terrestrial environments across temperate to tropical regions[49,58,59]. Although more than 560 species epithets have been recorded under the genus Torula, many of them are considered ambiguous, having been described solely based on morphological characteristics. Although certain morphological traits differentiate each species within Torula, their overall morphology is quite similar, making it difficult to use these features alone for species identification[49].
Accurate delimitation of the new species, as well as verification of the new country records, required the integration of detailed morphology with multi-locus phylogenetic analyses. Neither dataset alone provided sufficient resolution, but together they yielded a consistent and well-supported taxonomic framework.
Although not all gene regions were available for every isolate, the combination of the markers that were successfully sequenced (e.g., ITS, LSU, SSU, and selected protein-coding genes such as tub2, rpb2, or tef1-α, depending on the taxon) provided sufficient phylogenetic resolution to clarify the taxonomic placement and relationships. The multi-locus framework, even when partially represented, strengthened the integrative approach and improved species delimitation within morphologically similar groups such as Torula and related genera.
-
The authors confirm their contributions to the paper as follows: conceptualization, data curation, and formal analysis: Thambugala KM, Tennakoon DS, Samarakoon MC; funding acquisition: Thambugala KM, Hongsanan S; investigation, methodology, and writing − original draft: Thambugala KM, Tennakoon DS, Samarakoon MC, Milanka I, Daranagama DA; supervision: Thambugala KM, Hongsanan S; writing − review and editing: Thambugala KM, Tennakoon DS, Samarakoon MC, Milanka I, Daranagama DA. All authors reviewed the results and approved the final version of the manuscript.
-
All the data generated and analyzed during this study are available in the article. DNA sequence data are available in the GenBank database (www.ncbi.nlm.nih.gov/genbank), and the accession numbers are provided in Supplementary Tables S1–S3.
-
This work was supported by a research grant from the University of Kelaniya Research Council (Grant No. RC/SROG/2025/01). Shaun Pennycook is thanked for checking and suggesting corrections to the Latin names.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Collection details and GenBank accession numbers of the Apiospora isolates included in the multi-gene phylogeny. Strains generated in this study are shown in red, while type strains are indicated in bold.
- Supplementary Table S2 Collection details and GenBank accession numbers of the Torula isolates included in the multi-gene phylogeny. Strains generated in this study are shown in red, while type strains are indicated in bold.
- Supplementary Table S3 Collection details and GenBank accession numbers of the Paradictyoarthrinium isolates included in the multi-gene phylogeny. Strains generated in this study are shown in red, while type strains are indicated in bold.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Thambugala KM, Tennakoon DS, Samarakoon MC, Milanka I, Daranagama DA, et al. 2025. Discovery of Torula srijayewardenepurensis sp. nov. and distribution updates of Apiospora and Paradictyoarthrinium in Sri Lanka. Studies in Fungi 10: e032 doi: 10.48130/sif-0025-0035 

Discovery of Torula srijayewardenepurensis sp. nov. and distribution updates of Apiospora and Paradictyoarthrinium in Sri Lanka
- Received: 24 September 2025
- Revised: 28 November 2025
- Accepted: 29 November 2025
- Published online: 29 December 2025
Abstract: Fungal biodiversity remains an underexplored topic in Sri Lanka, particularly within microfungal communities inhabiting both anthropogenic and natural environments. During the continuous microfungal survey, three fungal isolates were obtained from the premises of the University of Sri Jayewardenepura. Based on morphology and multigene phylogeny (internal transcribed spacer: ITS, nuclear ribosomal 28S large subunit rRNA: LSU, nuclear ribosomal 18S small subunit rRNA: SSU, and translation elongation factor-1α: tef1-α), one of the isolates is introduced as a new species, Torula srijayewardenepurensis. It can be distinguished from the phylogenetically closely related T. chromolaenae and T. longan by having conidiophores reduced to conidiogenous cells and shorter conidiogenous cells. The other two isolates, identified as Apiospora pseudorasikravindrae and Paradictyoarthrinium diffractum, are supported by their morphological traits and phylogenetic relationships and represent new records for Sri Lanka. In addition, Apiospora pseudorasikravindrae comb. nov. (≡ Arthrinium pseudorasikravindrae), previously invalidly published, is validly published in this publication.
-
Key words:
- Ascomycota /
- Diversity /
- Phylogeny /
- Taxonomy /
- Urban habitats