









-
Fritillaria unibracteata, as one of the traditional basal source plants of Fritillaria cirrhos, has a long history of medicinal use. The earliest record of its medicinal use dates to the Han Dynasty (220 BC.) in China[1]. Its therapeutic properties have been useful in treating common illnesses, such as pneumonia, owing to its ability to reduce lung moisture, dissolve phlegm, and relieve cough[2]. F. unibracteata contains alkaloids, nucleosides, saponins, steroids, and terpenoids[3]. The nucleoside components of the majority of it are water-soluble and have been documented to exhibit properties such as anti-inflammatory effects, platelet coagulation inhibition, blood pressure reduction, and smooth muscle relaxation[4]. The medicinal effects of F. unibracteata were determined based on the concentrations of its nucleoside components. This concentration significantly influences its potency and serves as a crucial benchmark for quality evaluation.
F. unibracteata is mainly distributed on the Qinghai-Tibetan Plateau at altitudes of 3,000–5,000 m. This species grows slowly and requires a specific environment for growth. However, over-excavation has severely damaged the wild resources of F. unibracteata, leading to scarcity, and subsequently, a significant price increase. Between 2002 and 2017, the price of F. unibracteata increased from USD
${\$} $ ${\$} $ The Tibetan Plateau, where F. unibracteata grows, is an important pasture area in China. Cows and sheep are the main livestock grazers in this region. They excrete nitrogen annually through their feces, with cows excreting 9.0−23.0 × 104 t and sheep excreting 0.2−3.0 × 104 t[6,7]. Cow manure and sheep manure are produced in large quantities in the Tibetan Plateau, and their full use to develop cow manure organic fertilizer (CF) and sheep manure organic fertilizer (SF) not only reduces the use of chemical fertilizers and protects the ecological environment, but also helps to improve soil properties and achieve ecological restoration of farmland soils. For example, Honorato et al. found that adding organic cow manure to thyme (Thymus vulgaris L.) instead of chemical fertilizers increased dry matter accumulation and thymol content by 10-fold to 65.42%[8]. Du et al. found that the addition of organic fertilizer increased soil fertility, walnut growth, and enrichment of soil organisms with beneficial microorganisms (Bacillus spp. and Solicoccozyma spp.) and reduced the number of harmful microorganisms Fusarium spp.[9]. Previous studies have shown that the application of organic fertilizers can improve soil pH, influence soil microbial diversity, effectively regulate the structure of soil microbial communities, increase plant root vigor and nutrient uptake[10], and reduce the incidence of plant diseases[11,12]. However, the effects of applying cow and sheep manure as organic fertilizers on F. unibracteata yield quality, soil microbial community, and soil ecosystem function are unknown.
The objectives of this study were: (1) to elucidate the differences in soil physicochemical properties and soil microbial communities by applying cow and sheep manure organic fertilizers to F. unibracteata; and (2) to analyze and elucidate the effects of cow and sheep manure application on F. unibracteata yield and quality, soil microbial network structure, and the variation in key microorganisms and functional taxa. In this study, we aimed to assess the feasibility of using organic manure to improve F. unibracteata yield quality and beneficial soil microbial communities on the Tibetan Plateau, with potential implications for sustainable development within the F. unibracteata cultivation industry.
-
The experiments were conducted at a Fritillaria plantation base in Haidong City, Qinghai, China. The plantation base is located at 101°59' N, 36°59' E, and has an altitude of 3,050 m. The average annual temperature is 0 °C with an average daily difference of 13 °C. The annual precipitation was 466 mm. The warmest month has a maximum temperature of 18 °C while the coldest month has a minimum temperature of −17 °C. The study involved transplanting 3-year-old healthy F. unibracteata plants of similar size into the soil in October 2022. The total mass of F. unibracteata was recorded for each plot before planting. The basic characteristics of the original soil were determined before fertilizer application and are listed in Supplementary Table S1. Organic fertilizers from Hebei Rundong Fertilizer Co., Ltd. (Hebei, China), including well-rotted sheep and cow manure, were used in this study, and the specific physicochemical information is listed in Supplementary Table S1.
Experimental design
-
The field experiment consisted of three treatments: blank control (CK) treatment, sheep manure organic fertilizer (SF) treatment, and cow manure organic fertilizer (CF) treatment. Each treatment and control group had three replicates, totaling 33 plots with 162 plants per plot. The area of each plot was 1 m × 1 m, in Fig. 1a. Figure 1b and c present a photographic documentation of the experimental field and the surrounding environment. Before planting F. unibracteata bulbs in October 2022, all organic fertilizers were applied simultaneously, mixed well with the soil, and applied to a depth of 15−20 cm. The plots were then mulched to a height of 5−7 cm above the ground. Other field management measures were similar to conventional local practices. Before fertilization, the physicochemical properties of CF and SF were determined (Supplementary Table S1).
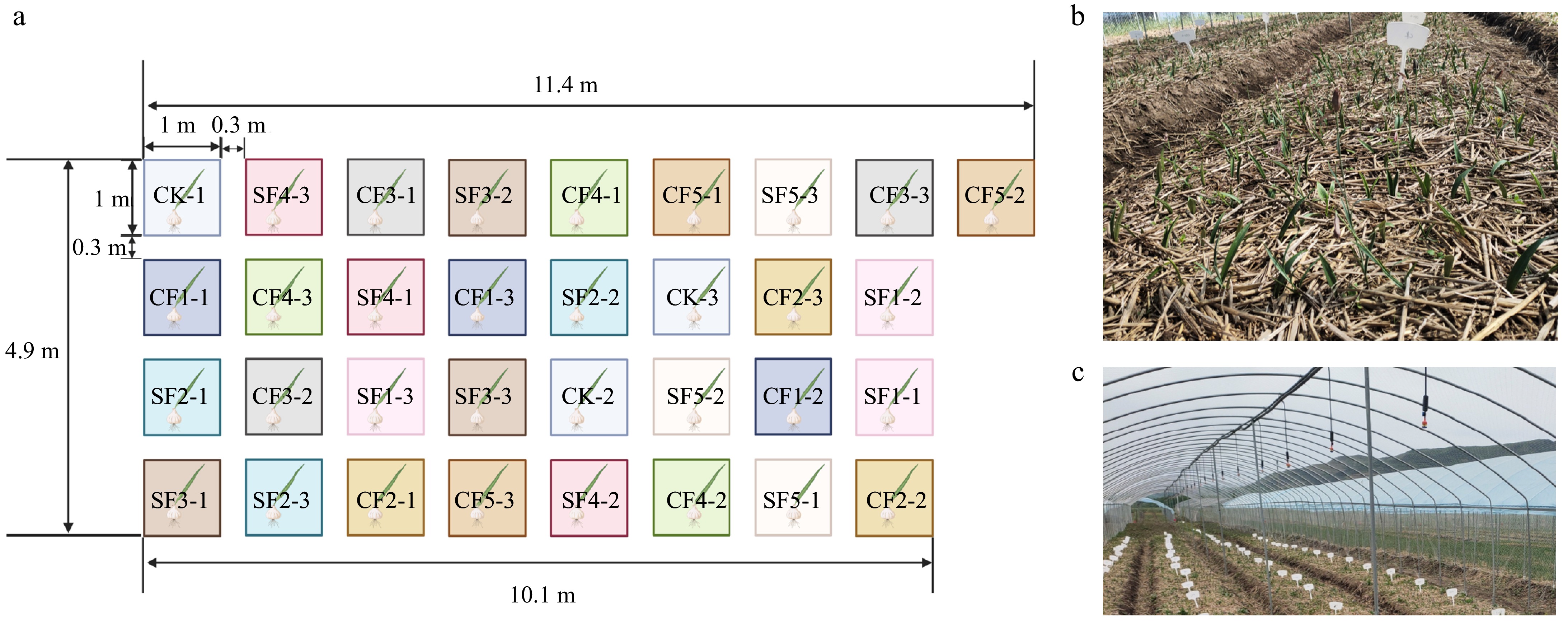
Figure 1.
Overview of the experimental design and layout. In the (a) experimental distribution map, each square's position corresponds to the plot position in the field, with every three squares of the same color representing three replicate treatment plots. The experiment was designed using a randomized distribution. (b) Actual photograph of the experimental plot. (c) Photograph of the overall environment of the experimental facility.
The germination rate was recorded on day 30 of F. unibracteata growth. On day 45, during the leaf development stage, leaf area was measured using a portable leaf area meter. On day 90 of F. unibracteata growth, soil samples were collected from the root zone on day 90 of the F. unibracteata growth. Finally, on day 120, during the bulb harvesting stage, the bulb yield was accurately measured using an electronic balance. Soil samples were collected from two treatment groups, CF 24.3 t·ha−1 and SF 43.7 t·ha−1, which had the most significant effect on F. unibracteata yield increase, and from the CK group, the no-fertilization blank control, for subsequent analyses. For soil sample collection, the soil was collected from 15 points at a depth of 10−20 cm within each treatment plot using the 'S' shaped sampling method. The soil collected from each plot was mixed thoroughly to ensure homogeneity. Subsequently, the soil was passed through a 20 mesh sieve, snap frozen in liquid nitrogen, transferred to 1.5 mL sterile EP tubes, and stored at −80 °C for subsequent experimental analyses. Following the above sampling, additional soil samples were dried in the shade, ground and passed through 80- and 20-mesh sieves for measurement of soil physicochemical properties and enzyme activities.
Soil physicochemical analysis and measurement of rhizosphere soil enzyme activities
-
Soil pH was analyzed after shaking the soil in a water-to-soil ratio of 2.5:1 (v/w) and was determined using a pH meter, according to the method of Chen et al.[13]. Soil total organic carbon (TOC) and total nitrogen (TN) were determined using an elemental analyzer according to the method of Liu et al.[14]. Total phosphorus (TP) and total potassium (TK) were determined using sodium hydroxide melting flame spectrophotometry according to the method of Ren et al.[15]. Available phosphorus (AP) was determined using the NaHCO3 method and available potassium (AK) was determined using ammonium acetate extraction–flame spectrophotometry. Available nitrogen (AN) was measured by diffusion dish method, according to the method of Wu et al.[16]. Soil urease (S-UE), soil catalase (S-CAT), soil neutral phosphatase (S-NP), soil polyphenol oxidase (S-PPO), soil dehydrogenase (S-DHA), soil sucrase (S-SC), and soil β-glucosidase (S-β-GC) were determined using 96-well microplate fluorimetric assay, following the method described by DeForest[17].
Analysis of plant growth characteristics, yield, and quality of F. unibracteata
-
At the emergence stage of F. unibracteata (30th day of growth), the emergence rate of F. unibracteata was recorded by observing and recording the emergence rate of F. unibracteata from each sample plot. At the leaf unfolding stage of F. unibracteata (on the 50th day of growth), 10 plants were randomly selected from each plot, and the leaf area was measured using a portable leaf area meter. At harvest time (120th day of growth) F. unibracteata, 20 plants were randomly selected from each sample plot, and the fresh weight of the clean plant bulbs was measured using a precision electronic balance. The nucleoside content of F. unibracteata bulbs was evaluated using high-performance liquid chromatography (HPLC) after harvesting[18]. The HPLC chromatographic conditions were as follows: the mobile phase consisted of deionized water (A) and acetonitrile (B): 0.00–10.00 min, 1.00%–5.00% B; 10.00%–15.00 min, 5.00%–15.00% B; 15.00–20.00 min, 15.00%–20.00% B; 20.00–30.00 min, 20%–30% B. The DAD detector wavelength was set at 260 nm.
DNA extraction and PCR amplification
-
Total DNA was extracted from a 0.50 g soil sample using the FastDNA® Spin Kit for Soil (MP Biomedicals, Inc., Santa Ana, CA, USA). The primers ITS1F (5'-CTTGGTCATTTAGAGGAAGTAA-3') and ITS4-2409R (5'-TCCTCCGCTTATTGATATGC-3') were used to amplify ITS2 sequences of fungi. The 27F (5'-AGAGTTTGATCCTGGCTCAG-3') and 1492R (5'-TACGACTTAACCCCAATCGC-3') primers were used to target the V3–V4 region of the 16S rRNA gene of bacteria. PCR was performed with an initial denaturation of 5 min at 95 °C, followed by 28 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s. The amplicons were sequenced using an Illumina NovaSeq6000 platform (Illumina, USA). The raw sequences were submitted to the Sequence Read Archive of the National Center for Biotechnology Information, and the accession number for the SRA data was PRJNA1094691.
Bioinformatics analysis
-
The QIIME software package (version 1.8.0) was used to analyze raw Illumina HiSeq sequencing data. Reads were trimmed by discarding those with quality scores < 20 and sequence lengths < 400 bp. Unique sequences among the remaining reads were used to define operational taxonomic units (OTUs) using Usearch (version 7.1,
http://drive5.com/uparse/ ), with a threshold of 97% similarity. The taxonomic identities of the phylotypes were determined using the Ribosomal Database Project (RDP) Classifier (version 2.2,https://sourceforge.net/projects/rdp-classifier/ ) at a confidence threshold of 70%. Alpha diversity, which included the Chao1, Shannon, and ACE indices, was analyzed using Mothur (v. 1.30.1)[19]. Mantel tests and correlation analyses were conducted using RStudio (4.2) and various R packages to examine the relationships between microbial communities, soil enzymes, and bioactive compounds. Furthermore, RStudio (4.2) was utilized to analyze the fungal and bacterial symbiotic networks and to compute their topological properties through pipeline processing. The results were subsequently visualized using the Gephi (0.10) software. Structural equation modeling (SEM) analysis was performed using RStudio (4.2) to investigate the effects of organic fertilizer addition on soil and plant variables, as well as microbial diversity.Statistical analysis
-
Data processing, analysis, and plotting were performed using Excel 2017, IBM SPSS v.20.0, and GraphPad Prism 9. Significant differences between samples (p < 0.05) were determined using one-way ANOVA. Analytical mapping and functional enrichment of the soil microbiome were performed using the Lynn BioCloud platform (
www.cloud.biomicroclass.com ). Principal coordinate analysis (PCoA) was used to classify the differences in microbial structure among the organic fertilizer samples. Redundancy analysis (RDA) was employed to determine the degree of correlation between the soil physicochemical properties and microbial communities. -
ANOVA revealed that the application of CF and SF significantly increased soil-available phosphorus (AP), soil-available potassium (AK), and soil-available nitrogen (AN) contents in F. unibracteata compared to the CK group (p < 0.05, Table 1). Comprehensive analysis showed that compared with the CK group, the application of CF and SF significantly improved soil fertility and promoted the quality and yield of F. unibracteata (Table 1).
Table 1. Soil chemical properties under different fertilization treatments.
Soil physicochemical
propertiesCK CF (24.3 t·ha−1) SF (43.7 t·ha−1) pH 7.94a ± 0.40 8.04b ± 0.40 8.08c ± 0.40 TOC (g·kg−1) 24.55b ± 1.31 24.70a ± 0.80 24.19b ± 0.69 TN (g·kg−1) 2.34a ± 0.07 2.33a ± 0.02 2.38a ± 0.03 TP (g·kg−1) 0.93b ± 0.01 1.10a ± 0.01 1.11a ± 0.03 TK (g·kg−1) 21.34ab ± 0.37 21.57ab ± 0.30 22.11b ± 0.66 AN (mg·kg−1) 141.06c ± 7.05 152.76a ± 7.64 146.57b ± 7.33 AK (mg·kg−1) 134.01c ± 6.70 215.03a ± 1.73 170.33b ± 8.52 AP (mg·kg−1) 19.21c ± 0.96 48.12a ± 2.40 41.41b ± 2.07 CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment. pH stands for soil pH; TOC represents soil total organic carbon content, g·kg−1; TN stands for soil total nitrogen content, g·kg−1; TP represents soil total phosphorus content, g·kg−1; TK represents soil total potassium content, g·kg−1; AN stands for soil available nitrogen content, mg·kg−1; AK stands for soil available potassium content, mg·kg−1; AP stands for soil available phosphorus content, mg·kg−1. Different letters (a, b, c) on the same row indicate values that are significantly different (p < 0.05) based on one-way ANOVA and LSD post-hoc test, the numbers after '±' represent the standard error values. ANOVA revealed significant effects of CF and SF application on the soil urease content, with increases of 44.07% and 61.02%, respectively, compared with the CK group (p < 0.05, Table 2). Notably, the application of CF did not significantly affect soil dehydrogenase content, whereas the application of SF led to a remarkable increase of 82.19% compared with the CK group. Additionally, the application of CF and SF resulted in a reduction in soil polyphenol oxidase content by 14.94% and 16.09%, respectively, compared to the CK group. No significant differences were observed in the contents of soil sucrase, soil neutral phosphatase, and soil β-glucosidase in the soil following the application of CF and SF, as compared to the CK group (p < 0.05).
Table 2. Soil enzyme activity content under different treatments.
Soil enzyme
activitiesCK CF (24.3 t·ha−1) SF (43.7 t·ha−1) S-CAT (mL·g−1) 3.57b ± 0.18 3.33ab ± 0.17 3.88a ± 0.19 S-SC (mg·d−1·g−1) 88.81a ± 4.44 89.39a ± 4.47 95.62a ± 4.78 S-UE (mg·d−1·g−1) 0.59c ± 0.03 0.85b ± 0.04 0.95a ± 0.05 S-NP (mg·d−1·g−1) 2.89a ± 0.14 3.09a ± 0.15 3.05a ± 0.15 S-PPO (mg·g−1·2h−1) 0.87a ± 0.04 0.74c ± 0.04 0.73b ± 0.04 S-β-GC (μg·g−1·h−1) 139.56a ± 6.98 133.15a ± 6.66 143.31a ± 7.17 S-DHA (μg·g−1·d−1) 90.58a ± 4.53 88.67a ± 4.43 165.03b ± 8.25 S-CAT represents soil catalase content, mL·g−1; S-SC represents soil sucrase content, mg·d−1·g−1; S-UE represents soil urease content, mg·d−1·g−1; S-NP represents soil neutral phosphatase content, mg·d−1·g−1; S-PPO represents soil polyphenol oxidase content, mg·g−1·2h−1; S-β-GC represents soil β-glucosidase content, μg·g−1·h−1; S-DHA represents soil dehydrogenase content, μg·g−1·d−1. Different letters (a, b, c,) on the same row indicate values that are significantly different (p < 0.05) based on one-way ANOVA and LSD post-hoc test, the numbers after '±' represent the standard error values. Optimized application of CF and SF increases F. unibracteata yield, emergence, and nucleoside content, while influencing uracil and guanine levels
-
The results suggest that the judicious application of CF and SF can have a significant impact on the growth and development of F. unibracteata. Examination of the influence of different fertilization treatments on the yield of F. unibracteata bulbs revealed a non-linear effect, where the yield initially increased and then decreased with increasing concentrations of CF and SF. Specifically, the highest F. unibracteata yield was achieved with CF application at an application rate of 24.3 t·ha−1, resulting in a statistically significant 119.45% increase over the CK group (p < 0.05). However, exceeding an application rate of 78.7 t·ha−1 of CF significantly decreased bulb yield. Likewise, SF application at 43.7 t·ha−1 resulted in the peak bulb yield for F. unibracteata, showing a significant 97.22% increase compared to the CK group (p < 0.05, Table 3).
Table 3. Yield of F. unibracteata under different fertilization treatments.
Treatment
groupWeighing before fertilization (g) Weight at
harvest (g)Yield increasing rate (%) CK (0 t·ha−1) 164.62 ± 8.23 383.61 ± 19.18 0.00% CF1 (7.5 t·ha−1) 156.73 ± 7.84 425.75 ± 21.29* 38.59%* CF2 (13.5 t·ha−1) 164.04 ± 8.2 427.22 ± 21.36* 27.41%* CF3 (24.3 t·ha−1) 151.9 ± 7.59 535.53 ± 26.78* 119.45%* CF4 (43.7 t·ha−1) 178.25 ± 8.91 507.44 ± 25.37* 51.67%* CF5 (78.7 t·ha−1) 178.21 ± 8.91 418.3 ± 20.92 1.63% SF1 (7.5 t·ha−1) 165.02 ± 8.25 501.31 ± 25.07* 70.72%* SF2 (13.5 t·ha−1) 170.48 ± 8.52 492.94 ± 24.65* 56.18%* SF3 (24.3 t·ha−1) 171.55 ± 8.58 521.85 ± 26.09* 71.17%* SF4 (43.7 t·ha−1) 163.05 ± 8.15 538.44 ± 26.92* 97.22%* SF5 (78.7 t·ha−1) 160.96 ± 8.05 412.36 ± 20.62 23.19%* CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment. * indicates statistically significant comparisons with the CK on the same columns (p < 0.05) based on one-way ANOVA and LSD post-hoc test, the numbers after '±' represent the standard error values. A similar trend was observed in the analysis of F. unibracteata emergence rates under various fertilization treatments, with an initial increase followed by a decrease as CF and SF concentrations increased. Although the highest emergence rates occurred with CF at 24.3 t·ha−1 (93.83 ± 2.01%) and SF at 43.7 t·ha−1 (94.33 ± 3.51%), compared to the CK group (Supplementary Table S2), these increases were not significant statistically. Furthermore, the application of SF at 43.7 t·ha−1 significantly increased the leaf area of F. unibracteata to 8.84 ± 1.38 cm2, which is approximately 22.00% increase compared to the CK group (p < 0.05, Supplementary Table S2).
The results of F. unibracteata quality analysis showed that uracil and guanine concentrations decreased with increasing organic fertilizer concentrations. In particular, the highest uracil and guanine contents in F. unibracteata bulbs were observed with SF application at 7.5 t·ha−1, resulting in statistically significant increases of 154.29% for uracil and 135.98% for guanine, compared to the CK group (p < 0.05). However, the application of SF at an application rate of 24.3 t·ha−1 resulted in significant reductions of 14.66% in uracil content (p < 0.05), respectively, compared to the CK treatment group (Fig. 2c).
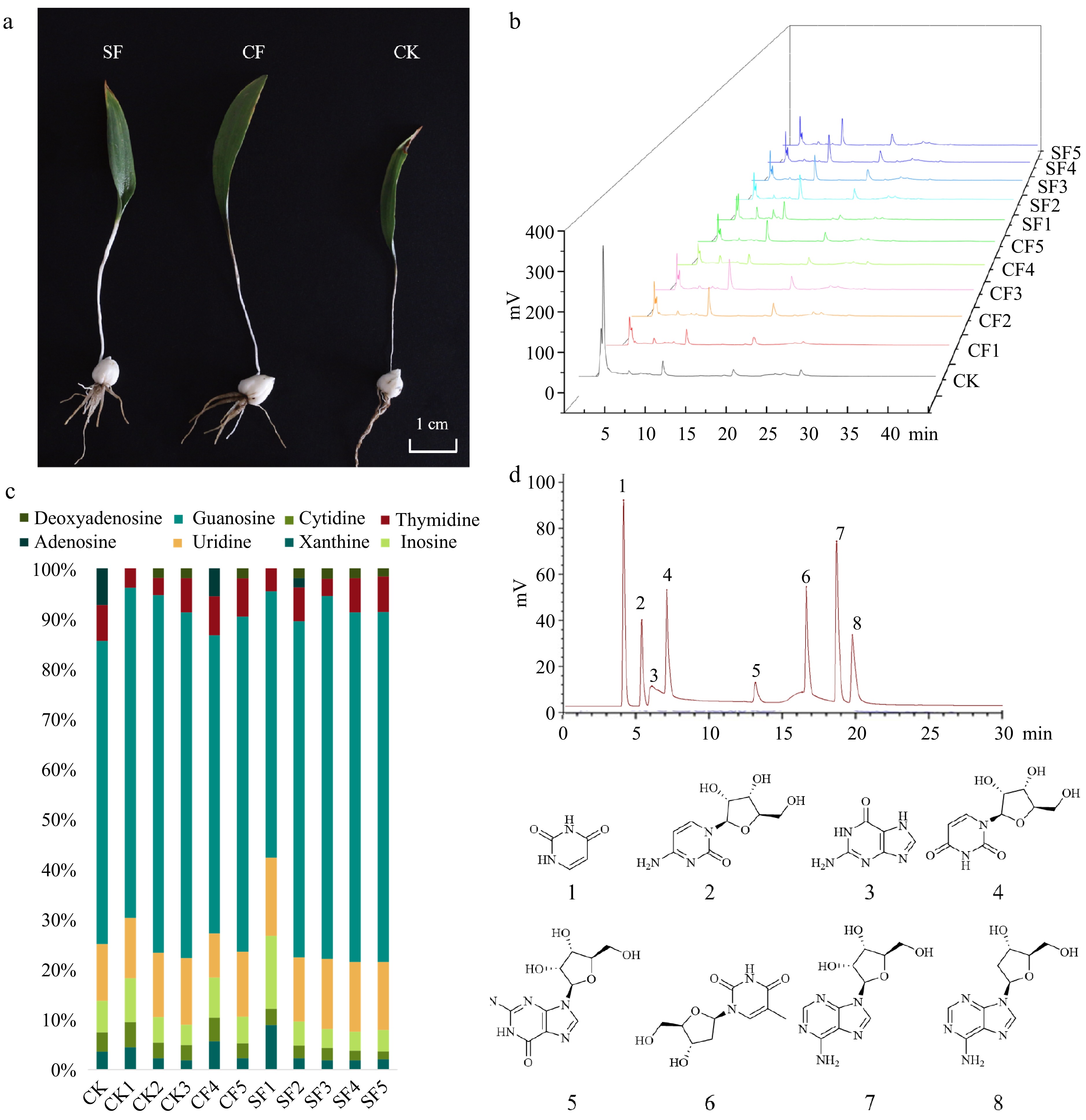
Figure 2.
(a) Plot of F. unibracteata growth under each treatment. (b) HPLC fingerprints of F. unibracteata bulbs from different fertilization treatments. (c) Accumulation plot of nucleoside content of F. unibracteata under different fertilization treatments. (d) Fingerprints of nucleoside active ingredient standards. Peak identification: 1. xanthine; 2. cytidine; 3. inosine; 4. uridine; 5. guanosine; 6. thymidine; 7. adenosine; 8. deoxyadenosine.
The nucleoside levels of uridine, guanosine, and adenosine showed a similar trend, with an initial increase, followed by a decrease, with increasing organic fertilizer concentration. Specifically, the application of CF at 24.3 t·ha−1 significantly increased uridine, guanosine, and adenosine levels in the bulbs of F. unibracteata by 100.01%, 48.37%, and 38.32% (p < 0.05), respectively, compared to the CK group. However, when the CF concentration was increased to 43.7 t·ha−1, the uridine level decreased by 16.40%, while the guanosine and adenosine levels increased by only 5.09% and 15.15%, respectively, compared to the CK group (Fig. 2c).
CF and SF shift relative abundance of dominant bacterial and fungal phyla without affecting overall diversity
-
According to dilution curve analysis, the curve tended to be flat, indicating that the amount of sequencing data in this study was sufficient to reflect the vast majority of the microbial information in the samples (Supplementary Fig. S1).
The investigation included an examination of the α-diversity of soil microbial communities in F. unibracteata by examining the Chao1, ACE, and Shannon indices (Fig. 3), allowing for a comprehensive assessment of the effects of different fertilization treatments. Analysis using these diversity indices using box plots revealed no significant deviations in microbial diversity and richness levels (p < 0.05), indicating that the exclusive application of CF over a one-year period did not significantly affect the α-diversity of soil microbial communities. Interesting trends were observed, indicating a tendency for increased soil bacterial diversity following the CF application, along with a similar trend for increased fungal diversity following the SF treatment. The PCoA plot showed clear segregation between the soil groups of F. unibracteata subjected to the three fertilization treatments, indicating notable differences in the microbial community structure of F. unibracteata soil among the different fertilization practices (Fig. 4a & b).
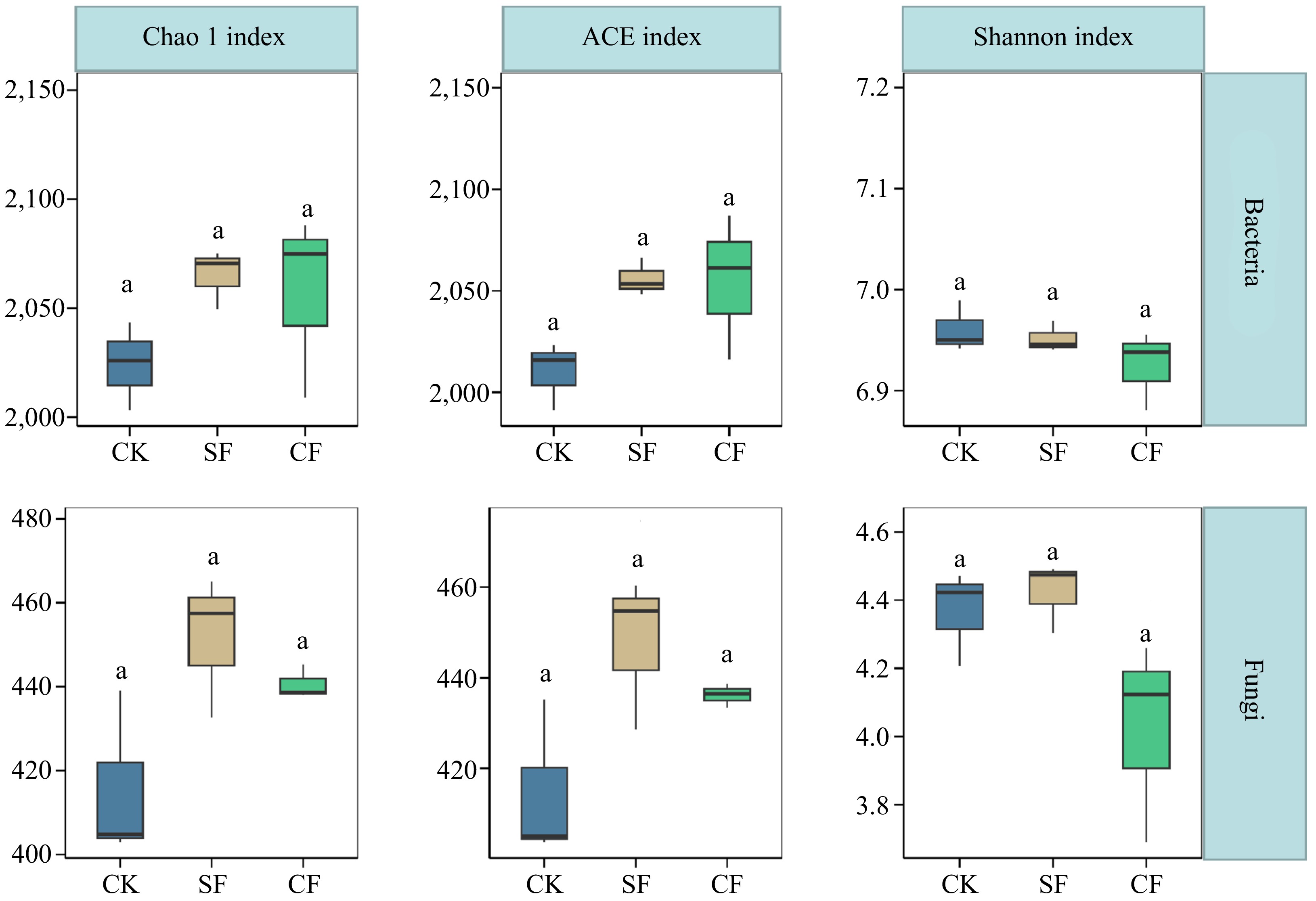
Figure 3.
Effect of different treatments on Alpha diversity indices (ACE, Chao1, and Shannon) of soil bacterial and fungal communities. The letter 'a' indicates that there is no significant difference between the treatment groups (p < 0.05; multiple comparison with ANOVA tests). CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment.
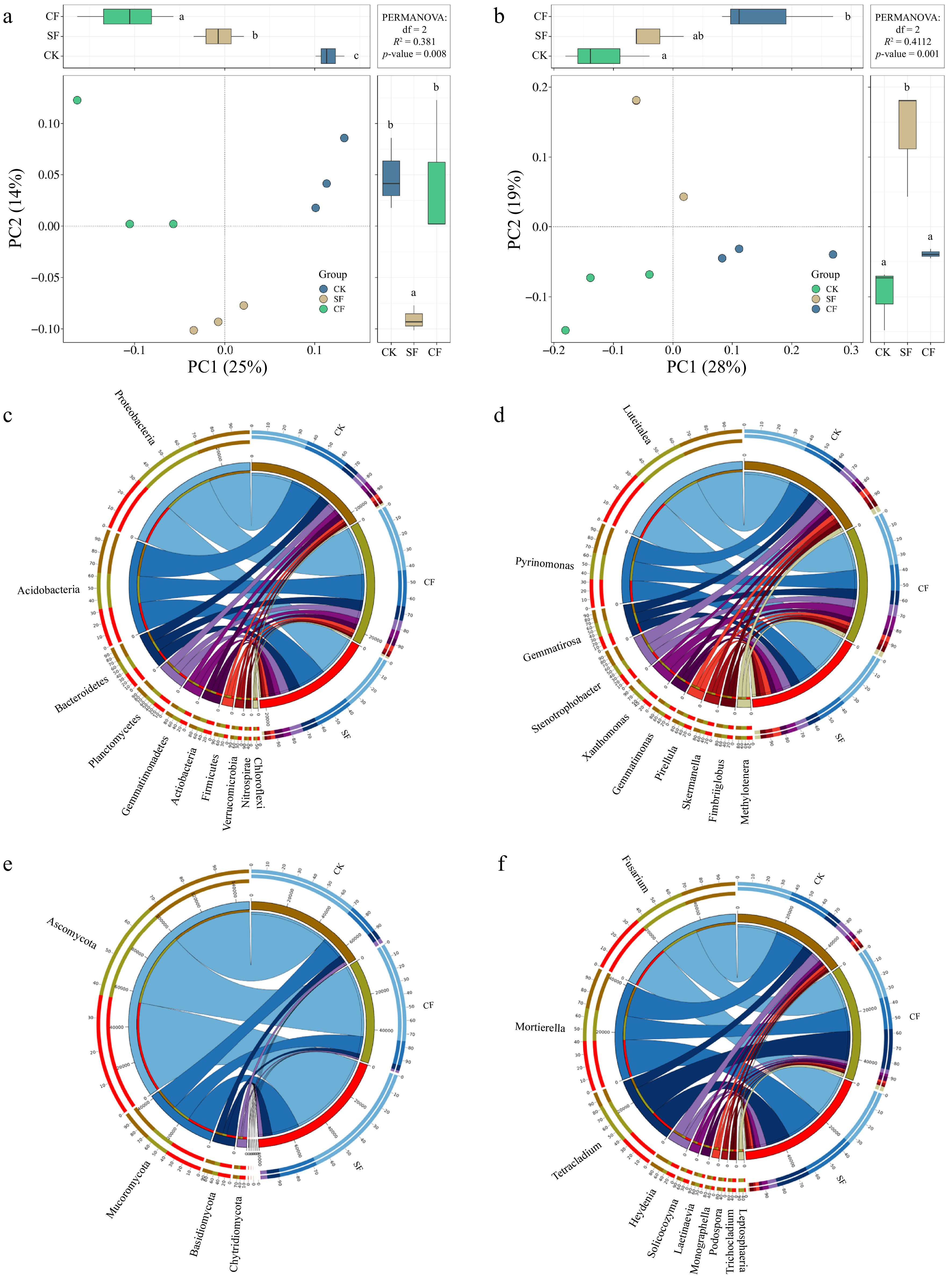
Figure 4.
PCoA results of (a) bacterial, and (b) fungal communities found with different fertilization types. PCoA plot of first two principal components based on operational taxonomic units in F. unibracteata with different fertilization types. Letters a, b, c indicate significant differences between treatments (p < 0.05; multiple comparison with ANOVA tests). CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment. Composition of the soil microbial community under different fertilization regimes. (c) Composition of the bacterial community at the phylum level. (d) Composition of the fungal community at the phylum level. (e) Composition of the bacterial community at the genus level. (f) Composition of the fungal community at the genus level.
Bacterial OTUs were assigned to 21 phyla and 365 genera, whereas fungal OTUs were assigned to seven phyla and 121 genera. Within the bacterial communities of all fertilization treatment groups (with a relative sequence abundance greater than 1.00%), the predominant bacterial phyla were Proteobacteria (38.36%), Acidobacteria (27.01%), Bacteroidetes (9.80%), Planctomycetes (5.62%), Gemmatimonadetes (5.46%), Actinobacteria (4.37%), Firmicutes (2.75%), Verrucomicrobia (1.82%), and Cyanobacteria (1.04%), which together accounted for 96.23% of the bacterial microbial sequences (Fig. 4c). However, the fungal communities of all fertilization treatment groups (with a relative sequence abundance greater than 1.00%) were characterized by the presence of a dominant fungal phyla such as Ascomycota (71.65%), Mucoromycota (20.05%), Basidiomycota (5.69%), Chytridiomycota (2.51%), Zoopagomycota (0.05%), Olpidiomycota (0.03%), and Microsporidia (0.02%), encompassing the entirety of the fungal microbial sequences (Fig. 4d). In addition, the proportion of some phytopathogenic fungi decreased after CF application at the fungal genus level compared with CK; for example, Heydenia abundance decreased by 63.10%, Podospora abundance decreased by 40.00%, and Solicoccozyma abundance decreased by 19.40% (p < 0.05; Fig. 4f).
Analysis of bacterial community composition at the phylum level revealed significant treatment-induced variations (Fig. 5a). Compared to the CK control, CF application significantly reduced the relative abundances of Acidobacteria and Cyanobacteria (p < 0.05). In contrast, SF application significantly increased Planctomycetes abundance (p < 0.05), while showing non-significant increases in Actinobacteria and Bacteroidetes. CF treatment significantly enhanced Proteobacteria abundance (p < 0.05), with marginal, non-significant increases in Firmicutes and Gemmatimonadetes.
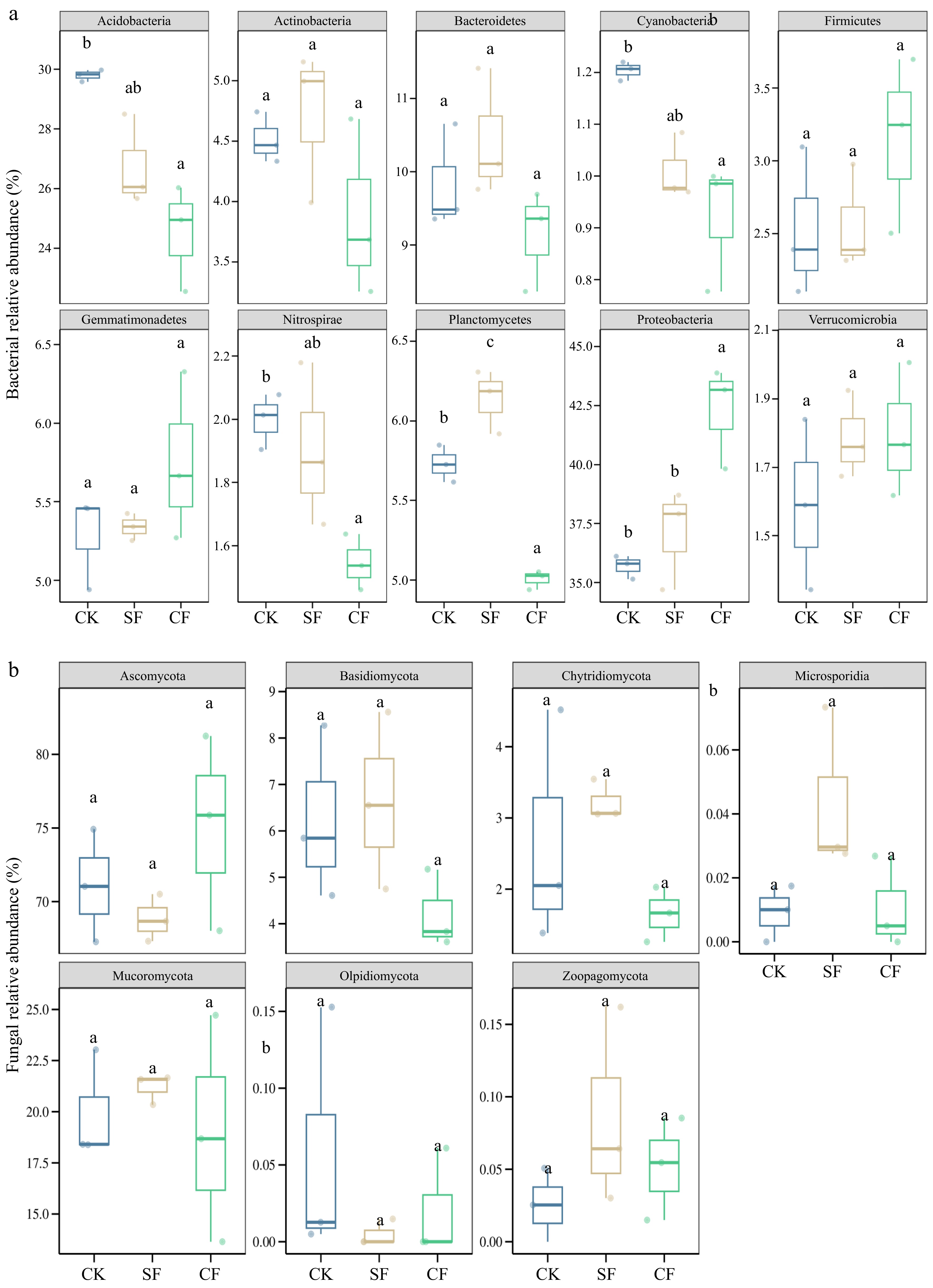
Figure 5.
Sectional boxplots of group differences in the relative abundance of the top 10 species in soil samples from the comparative study of soil (a) bacterial and (b) fungal phyla levels under different fertilization treatments. Treatment groups with different lowercase letters indicate significant differences. Different letters (a, b, c) indicate significant differences among treatments (p < 0.05; multiple comparison with ANOVA tests). CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment.
Fungal community analysis at the phylum level revealed distinct patterns (Fig. 5b). The CF amendment resulted in increased Ascomycota abundance compared to CK, although this change was not statistically significant (p < 0.05). Both the CF and SF treatments showed similar trends of increased Zoopagomycota abundance relative to the control, but these differences were not statistically significant (p < 0.05).
CF and SF increase co-occurrence network connectivity and modularity in soil microbial communities
-
When analyzing the topological indices of the co-occurrence network within the bacterial community, the application of CF resulted in the highest number of edges and an average degree in the co-occurrence network, surpassing other fertilizer treatments (Supplementary Table S3). Furthermore, the modularity index of the co-occurrence network was the most pronounced in the CF treatment group, with the second-highest value observed in the SF treatment group. This observation implies that the application of CF and SF may enhance connectivity within the soil bacterial community, thereby promoting high modularity (Fig. 6a). However, the results of the robustness analysis of the bacterial network showed greater robustness in the SF treatment group than in the CF and CK groups (Fig. 6c).
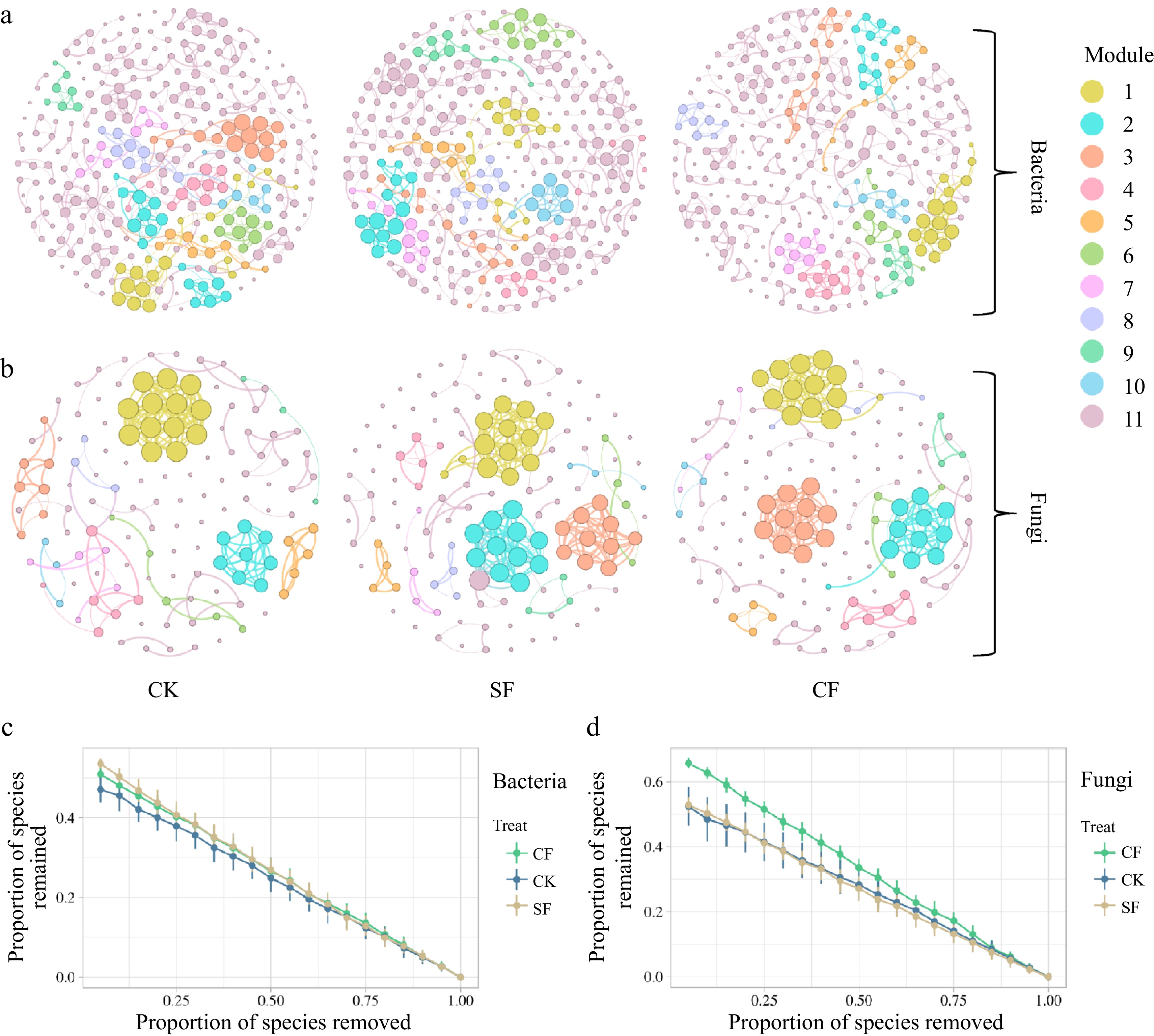
Figure 6.
Co-occurrence networks of the soil microbial communities at the genus level in (a), (b) different fertilization treatment groups; and (c) the robustness of bacterial network, and (d) fungal network. The node size is proportional to the taxon abundance, and the nodes represent bacterial or fungi taxa at the genus level (genera with relative abundances greater than 0.10%). The node colors represent different bacterial and fungal phyla. CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment.
Similarly, examination of the topological indices of the fungal community network revealed an increase in the total number of edges, average clustering coefficient, graph density, and average degree of co-occurrence in the CF and SF treatment groups, compared to the CK treatment group (Supplementary Table S3). In addition, the modularity index was most pronounced in the CF treatment group, indicating that the application of CF enhanced interconnectedness within the fungal community, resulting in a more compact fungal community structure compared to the other treatment groups (Fig. 6b). Furthermore, the results of the fungal network robustness analysis highlighted the superior robustness of the CF treatment group compared to the CK and SF treatment groups, indicating an improvement in the stability of the fungal community structure (Fig. 6d).
CF and SF improve microbial functions related to metabolism and organic matter decomposition
-
FAPROTAX functional analysis revealed that chemoheterotrophy, aerobic heterotrophy, urea degradation, xylan degradation, and nitrate reduction were dominant bacterial community functions in each fertilizer treatment group (Fig. 7a). The functional abundances of chemoheterotrophic and aerobic heterotrophic bacteria were significantly higher in the CF and SF treatment groups than in the CK treatment group. This observation suggests that the application of CF and SF increased the relative abundance of chemoheterotrophic and aerobic heterotrophic bacteria in the soil, thereby enhancing the decomposition and transformation of organic matter.
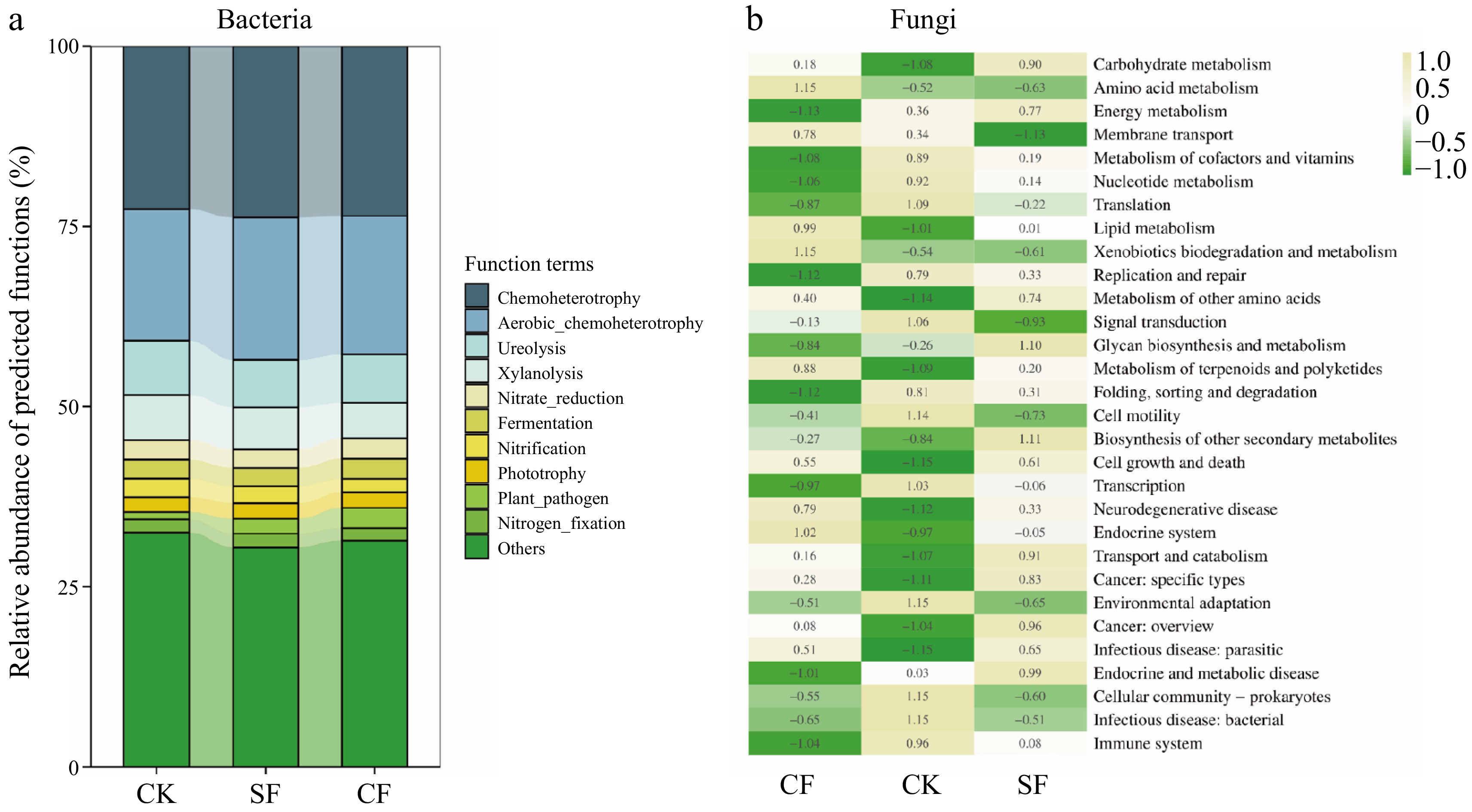
Figure 7.
Analysis of soil microbial community function in different treatment groups. (a) Abundance barplot of FAPROTAX functional analysis of bacterial community and (b) the heat map of PICRUSt2 functional analysis of fungal community were performed. CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment.
According to PICRUSt2 functional analysis, metabolism emerged as the primary functional classification of the fungal community in each fertilizer treatment group. Among the top 30 functions, the SF treatment significantly increased the abundance of carbohydrate metabolism, partial amino acid metabolism, energy metabolism, sugar biosynthesis metabolism, and secondary metabolite biosynthesis compared with the other treatment groups. In contrast, the CF treatment significantly improved amino acid metabolism. Notably, the biodegradation and metabolism of xenobiotics, metabolism of terpenoids and polyketides, and other functional relationships were also affected (Fig. 7b). These results indicate that the application of CF and SF enriches the fungi associated with organic matter metabolic functions, thereby promoting the decomposition and transformation of organic fertilizers.
Key soil factors selectively shape bacterial and fungal communities that influence F. unibracteata yield and quality
-
Linear regression analyses revealed a positive correlation between the soil environmental factors and soil microorganisms (Fig. 8a & b). The RDA showed that the selected environmental factors accounted for 40.35% of the total variation in the bacterial community (Fig. 8c) and 47.43% of the total variation in the fungal community (Fig. 8d). The RDA plots identified pH (ANOVA, p = 0.027), AP (ANOVA, p = 0.017), and TP (ANOVA, p = 0.022) as the key environmental factors shaping the bacterial community composition, whereas pH (ANOVA, p = 0.002), TP (ANOVA, p = 0.009), and AN (ANOVA, p = 0.008) were identified as the primary drivers of fungal community composition. Random forest (RF) analysis confirmed that soil pH and TP, AP, AK, and AN contents were the predominant influencers of soil microbial community variation (Fig. 8e & f).
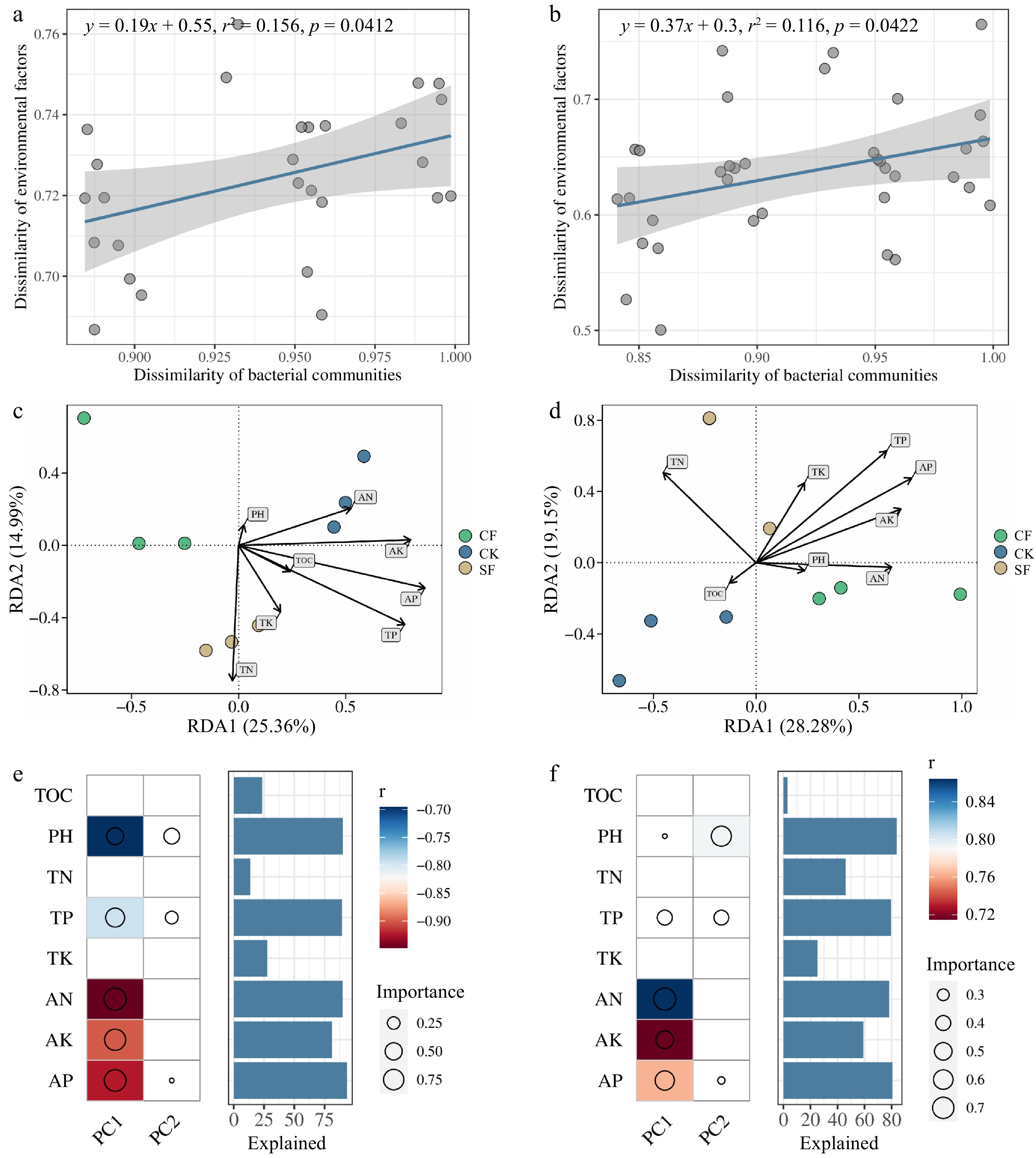
Figure 8.
Plot of the results of the ordinal regression analysis of environmental factors. Linear regression to assess the correlation of Euclidean distances of environmental factors with Bray-Curtis distances of (a) bacterial and (b) fungal microbial communities. Distance-based redundancy analysis (RDA) among (c) bacterial, or (d) fungal communities and environmental factors. Heat map of correlations between environmental factors and changes in (e) bacterial and (f) fungal microbial communities and their contribution. CK: no fertilizer treatment, CF: cow manure organic fertilizer treatment, SF: sheep manure organic fertilizer treatment.
The Mantel test revealed a significant positive correlation (r = 0.52) between the bacterial community and cytosine levels in the F. unibracteata bulb (Fig. 9a). Similarly, a positive correlation (r = 0.36) was observed between the fungal community and cytosine content in the F. unibracteata bulb. Pearson's analysis was used to examine the relationship between the relative abundance of bacterial and fungal phyla and F. unibracteata bulb yield and quality (Fig. 9b & c). The results indicated significant positive correlations between the relative abundances of Proteobacteria (r = 0.78), Deinococcus-Thermus (r = 0.75), and Candidatus Sumerlaeota (r = 0.73), and F. unibracteata bulb yield (p < 0.05). Conversely, significant negative correlations were observed between the relative abundances of Cyanobacteria (r = −0.86), Acidobacteria (r = −0.70), and F. unibracteata bulb yield (p < 0.05). Furthermore, the microbial phyla Verrucomicrobia, Rhodothermaeota, Proteobacteria, Ignavibacteriae, Deinococcus-Thermus, and Candidatus Sumerlaeota showed significant positive correlation with F. unibracteata bulb quality (p < 0.05). Conversely, Planctomycetes, Nitrospirae, Cyanobacteria, Basidiomycota, Chytridiomycota, and Olpidiomycota were significantly and negatively correlated with F. unibracteata bulb quality (p < 0.05).
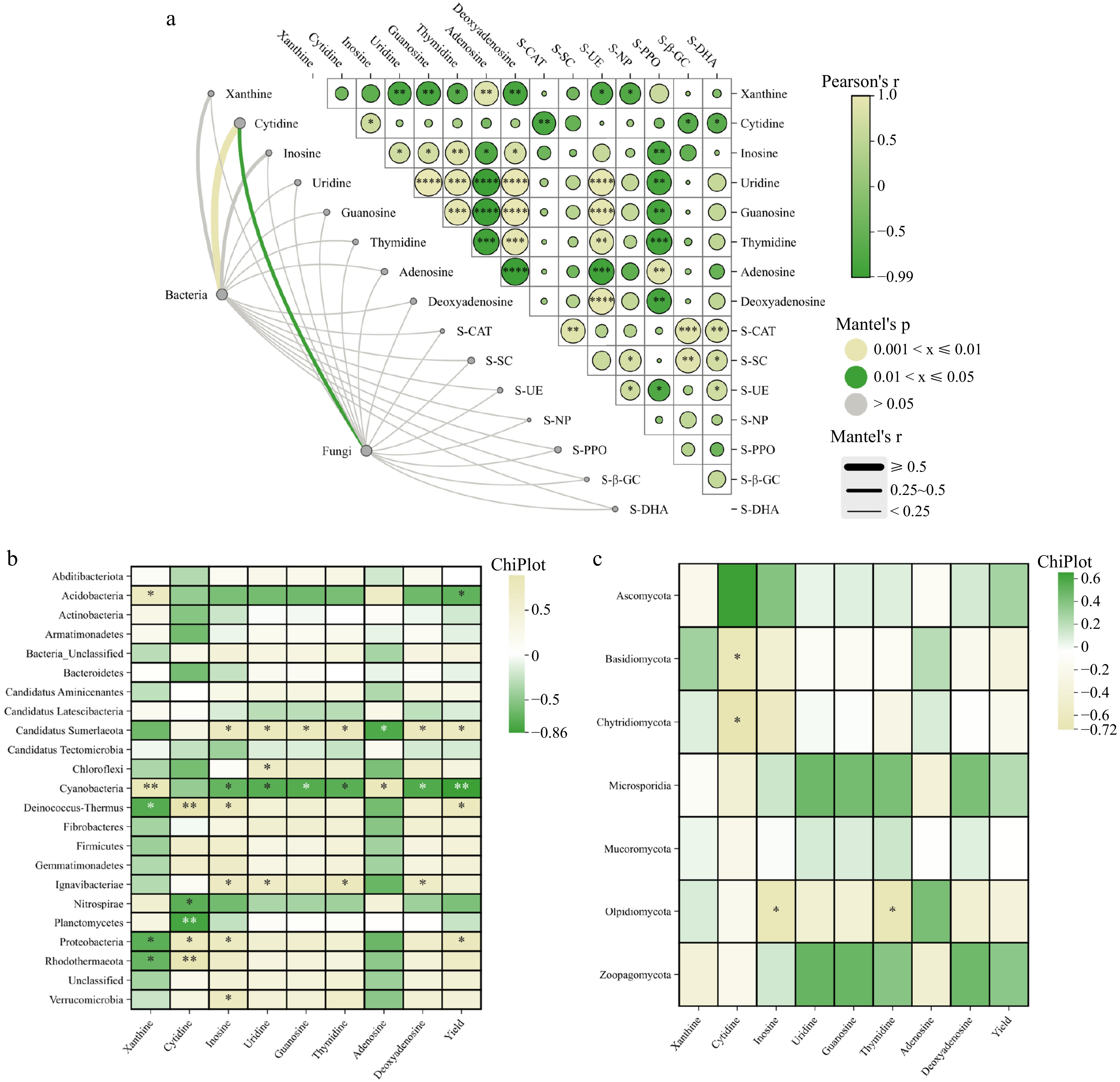
Figure 9.
(a) Mantel test analysis among soil enzymes, nucleosides, and microbial communities. Correlation analysis between (b) bacterial and (c) fungal phylum and the yield and quality of bulbs (*, p < 0.05, **, p < 0.01).
SEM was used to further elucidate the complex relationships between soil physicochemical properties, bacterial community diversity, fungal diversity, F. unibracteata yield, and quality. The model showed a high degree of fit to the data (χ2/df = 17.676, GFI = 1, SRMR = 0.064), indicating substantial agreement between the hypothesized model and the empirical observations. The results demonstrated that soil physicochemical properties, enzyme activity, and microbial diversity accounted for 99.8% of the variance in F. unibracteata yield. The TP, AP, and F. unibracteata quality showed significant positive correlations, whereas soil enzyme activity and bacterial diversity were negatively correlated. TP, soil pH, and soil enzyme activity were positively correlated with bacterial diversity, and these factors combined accounted for 70.5% of the variance in bacterial diversity. TP, soil pH, and soil enzyme activity were also significantly positively correlated with fungal diversity, accounting for 60.1% of the variance in fungal diversity. A significant positive correlation was also found among soil TP, soil enzyme activity, and F. unibracteata quality (Fig. 10).
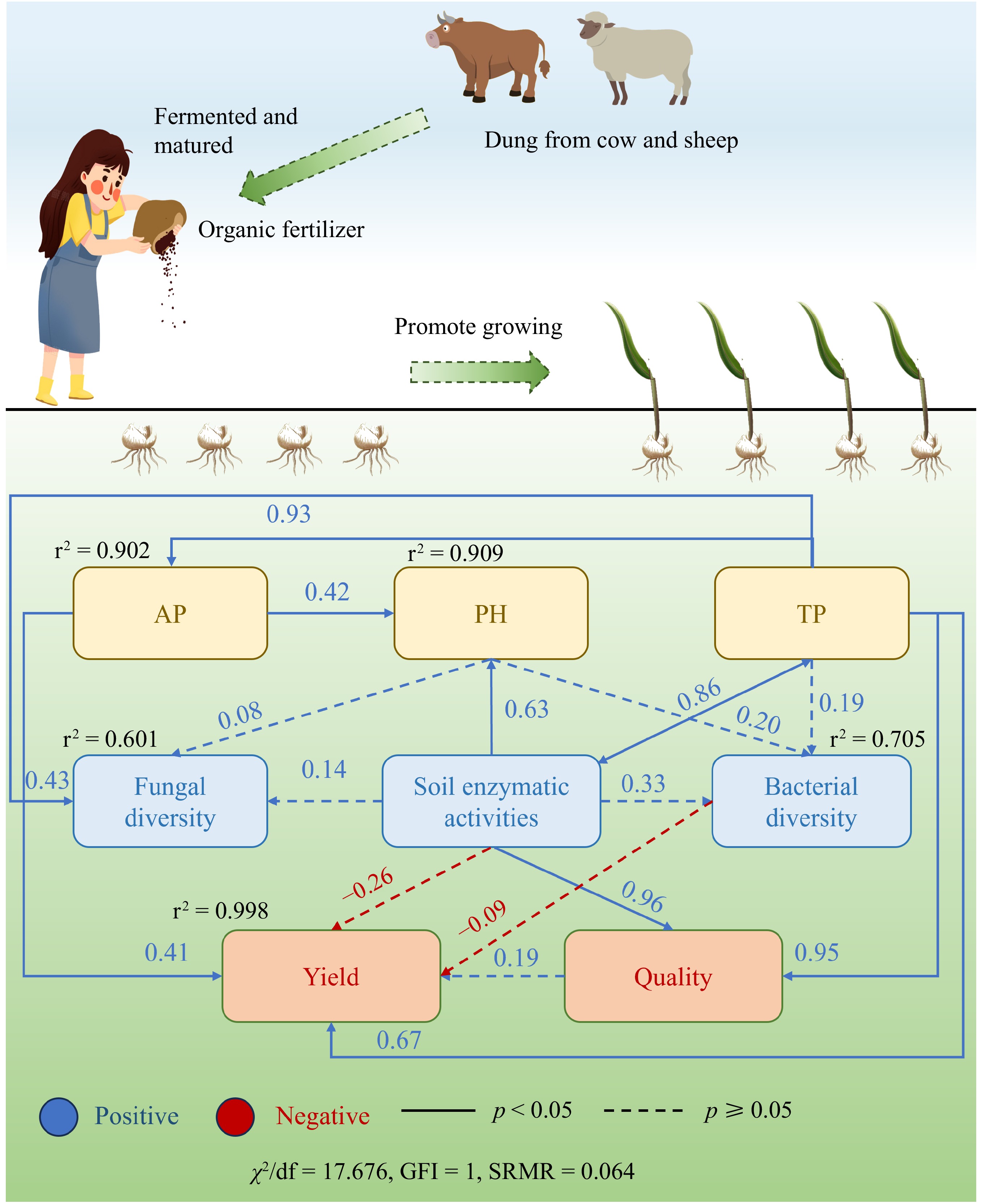
Figure 10.
Structural equation modeling results describing the relationship among soil nutrients, microbial diversity, and F. unibracteata yield and quality. The relationship among soil pH value, total soil phosphorus content (TP), available phosphorus (AP), F. unibracteata production, and F. unibracteata quality with bacterial diversity, fungal diversity, and soil enzymatic activities. Blue lines: positive correlation; full line: p < 0.05; dotted line: p ≥ 0.05. The numbers above the arrows indicate correlation strength. r2 denotes explained variance score. χ2, Chi-square; df, degrees of freedom; SRMR, standardized root mean square residual.
-
Organic fertilizers are primarily derived from animal manure, animal and plant residues, and other organic-matter-rich byproducts. These materials are transformed into green fertilizers through a series of processes, including fermentation and decomposition. Extensive research has demonstrated that the application of organic fertilizers can enhance plant yield and quality, increase soil nutrient enzyme activity, and bolster soil microbial communities[20−22].
The present study showed that the application of organic manure derived from CF and SF significantly increased the AP, AK, and AN contents in the soil (Table 1). This observation is consistent with previous findings regarding fertilizer application in soybean[23] and rice crops[24]. This finding confirms previous research on the remediation of acidic soils through long-term application of organic fertilizers[25]. In the present study, the application of both CF and SF resulted in a noticeable increase in the soil S-UE and S-NP content (Table 2). S-UE and S-NP play crucial roles in plant growth because they are involved in urea degradation and organic phosphorus conversion[26]. Furthermore, our study revealed that CF and SF significantly reduced the S-PPO content in the soil. Zhou et al. also reported a decrease in S-PPO content with the application of organic fertilizers in a wheat-maize rotation system[22], which is consistent with our findings. By reducing the S-PPO content in the soil, enzymatic browning can be effectively inhibited, thereby enhancing the quality and economic value of plants.
In this study, the application of appropriate concentrations of CF and SF significantly increased the yield and quality of F. unibracteata. Application of CF at 24.3 t·ha−1 and SF at 43.7 t·ha−1 significantly increased F. unibracteata yield and quality, resulting in 119.45% and 97.22% improvements over the CK group, respectively (p < 0.05, Table 3). This observation is consistent with previous findings regarding the positive effects of organic fertilizers on the yield and quality of medicinal plants, such as Perilla frutescens[27], Chrysanthemum[28], and Sophora tonkinensis[29]. Organic fertilizers enhance crop yield and quality by improving soil conditions and modulating soil microbial communities, thereby promoting soil metabolism and nutrient cycling.
Changes in microbial community diversity and composition due to organic fertilization
-
Soil microorganisms are indispensable components of agricultural ecosystems, enhance soil fertility through mineralization and nitrogen fixation, and serve as crucial indicators of soil quality. In this study, the application of CF and SF for one year did not significantly alter the soil microbial diversity of F. unibracteata (Fig. 3). These results are consistent with those of previous studies, suggesting that soil microbial diversity and abundance are relatively stable. Although common agricultural practices can influence soil physicochemical properties and microbial communities to some extent, they often have a limited impact on soil microbial diversity and richness.
Fertilizer management is an important factor influencing soil microbial diversity and is associated with changes in soil organic matter content, nutrient availability, and pH. Based on the species-level differential analysis at the bacterial phylum level, the application of CF and SF reduced the relative abundances of Cyanobacteria and Acidobacteria. Cyanobacteria, which are self-photosynthetic nutrient producers, and Acidobacteria, which are commonly found in nutrient-poor soils, are important microbial indicators of soil fertility. This reduced abundance suggests that the CF and SF treatments improved soil fertility. Generally, the behavior and function of microorganisms in the environment can be assessed in terms of their beneficial or harmful effects. For example, beneficial microorganisms promote plant growth, whereas harmful microorganisms can cause plant diseases[30−32].
In addition, CF increased the population of beneficial bacteria within the Proteobacteria phylum compared to that in the control group (Fig. 5a). For example, CF increased the abundance of Comamonadaceae, Hyphomicrobiaceae, and Sphingomonadaceae. Comamonadaceae produce urease, which degrades urea, thereby inducing cadmium mineralization, and are as well as siderophores, indole-3-acetic acid (IAA), and other plant growth-promoting substances[33]. Hyphomicrobiaceae produce antimicrobial compounds that combat invading plant pathogens[34] (Fig. 4e).
In addition, the proportion of some phytopathogenic fungi decreased after CF application at the fungal genus level compared with that of CK; for example, Heydenia abundance decreased by 63.1%, Podospora abundance decreased by 40.0%, and Solicoccozyma abundance decreased by 19.40% (Fig. 4f).
In conclusion, the use of organic fertilizers has the potential to enhance the relative abundance of beneficial bacterial and fungal communities in F. unibracteata soils. Specifically, the application of organic fertilizers has been shown to increase the relative abundances of several bacterial communities, including those belonging to Proteobacteria, Hyphomicrobiaceae, and Sphingomonadaceae. The application of organic fertilizers resulted in an increase in the relative abundance of these beneficial microbial communities, which contributed to an improvement in soil microbial diversity, promoted the growth and development of F. unibracteata, and enhanced its yield and quality of F. unibracteata.
When applied over the long term, organic fertilizers can provide adequate nutrient sources for soil microorganisms, thereby increasing soil microbial diversity. This led to an increase in the abundance of microorganisms involved in soil nutrient cycling. These microorganisms play essential roles in the soil, such as nitrogen fixation, phosphorus solubilization, and potassium mineralization, which in turn improve soil fertility. In addition, the long-term application of organic fertilizers can increase the organic matter content in the soil, which helps improve soil structure and increases soil water-holding capacity and aeration, thus promoting plant growth[35−37].
SF treatment and CF treatment increased the connectivity and structural stability of soil microbial communities
-
Microbial networks are commonly used in soil microbiological analyses of a wide range of plants, including rice[38] and ginseng[39]. These intricate networks of microbial communities function in concert to maintain an intricate balance in the soil ecosystem, thereby influencing larger soil communities. These interactions are objective and serve as important indicators of the soil balance. Changes in fertilizer management practices can result in shifts in the soil microbial co-occurrence network[40]. Following the application of CF and SF, the modularity index of the soil microbial community was higher than that in the CK group. This suggests that the application of these organic fertilizers increased the interconnectedness of soil microbial communities and promoted closer interactions between them (Fig. 6a & b; Supplementary Table S3), which is consistent with previous studies on this topic[41−43].
Communities with complex, co-occurring microbial networks are more resilient in their environments than simpler, less-interconnected networks. These networks comprise numerous interconnected nodes that interact in diverse and complex ways. This extensive connectivity results in a decentralized system structure that reduces the effect of a failure or perturbation in one node on the functioning of the entire network. By contrast, simpler networks tend to have a more centralized structure owing to less connectivity, making them vulnerable to the failure of a critical node, which can lead to the collapse of the entire system[44]. The application of CF in this study significantly enhanced the network connectivity of the soil bacterial and fungal communities compared to the CK group. This suggests that organic fertilizers enhanced the resilience of soil microorganisms to perturbations, promoted closer interactions between bacteria and fungi, and thus enriched network system complexity (Supplementary Table S3). Increasing the network system complexity of soil microbial communities has the potential to improve the degradation efficiency of soil organic matter and the adaptability of soil microorganisms to environmental changes. In addition, robustness analyses revealed that the application of organic manure enhanced the stability of bacterial and fungal network structures (Fig. 6c & d). This may be due to the richness of nutrients in cow and sheep manure, which can provide sufficient energy and nutrients to soil microorganisms after application, thereby improving the compactness and stability of the microbial community structure. For example, Li et al.[45] found that soil microbial community network connectivity could be improved and soil nutrient cycling could be promoted by adding sheep manure to a wheat-maize rotation system.
Relationship among soil physicochemical properties, soil microbial community, and F. unibracteata yield and quality
-
This study revealed a positive association between the modified soil microbial community following the application of organic fertilizers and the accumulation of the active ingredient cytosine in the F. unibracteata bulbs (Fig. 9a). Notably, Nitrospirae and Planctomycetes showed a strong positive association with cytosine accumulation in F. unibracteata bulbs (Fig. 9b). Similarly, a considerable positive association was observed between the Cyanobacteria microbial community and the accumulation of multiple active ingredients, including inosine, uridine, guanosine, thymidine, and deoxyadenosine, as well as the yield of F. unibracteata bulbs. These findings suggest a potential link between alterations in the soil microbial community following organic fertilization and the accumulation of active ingredients and yield in F. unibracteata bulbs. Furthermore, the study identified a positive association between the microbial communities Planctomycetes, Nitrospirae, Cyanobacteria, Basidiomycota, Chytridiomycota, and Olpidiomycota and the yield of F. unibracteata (Fig. 9b & c). These findings suggest a possible role for these microorganisms may be involved in the growth and development of F. unibracteata. Further research is required to elucidate the specific mechanisms by which these microbial communities influence the accumulation and yield of active ingredients.
The RDA results highlighted TP and AP as the most influential factors affecting both the bacterial and fungal communities (Fig. 8). Once introduced into the soil, a fraction of the fertilizer is assimilated by plants to meet their nutritional needs, whereas the remaining fraction can alter the soil environment. This dynamic can lead to shifts in the composition of bacterial[46] and fungal[47] communities within the soil and consequently affect the diversity and functionality of the soil microbial community. Different fertilization treatments can affect the composition of the soil microbial community by modifying the physicochemical properties of F. unibracteata soils, with a notable effect originating from the levels of AP and TP.
SEM analysis revealed significant positive correlations between TP and soil enzyme activity and the quality (nucleoside content) of F. unibracteata plants (Fig. 10), indicating that increasing TP and enzyme activity could improve plant quality. Furthermore, the yield of F. unibracteata showed a direct and significant positive correlation with AP, indicating the effect of AP content on plant yield, with increased AP content potentially leading to improved yield. In addition, the diversity of soil bacteria and fungi showed significant positive correlations with TP, soil pH, and soil enzyme activity. It has been found that increasing soil phosphorus content can improve crop yield and quality[48]. Similarly, increasing soil AP content can increase soil pH and soil enzyme activity and strengthen soil microbiota, which is consistent with the results of our study[49]. Notably, this study found a negative correlation between soil bacterial diversity and F. unibracteata yield, which is in contrast to previous long-term fertilization studies[50−53]. However, in this short-term study, the application of organic fertilizer significantly increased the bulb yield of F. unibracteata without a significant increase in bacterial diversity (Chao1 index) (Fig. 3). This discrepancy could be attributed to the fact that organic manure application may stimulate the rapid growth of specific dominant beneficial bacteria in the soil over a short period. At the same time, it may reduce the abundance of oligotrophic microorganisms that are not accustomed to organic fertilizers. The application of organic manure has the potential to improve soil physicochemical properties, particularly TP and AP levels, and effectively improve the quality and yield of F. unibracteata.
-
The findings of this study indicate that the application of optimal concentrations of CF (24.3 t·ha−1) and SF (43.7 t·ha−1) resulted in enhanced synergism and stability of the soil microbial community structure, an increased proportion of chemoenergetic heterotrophic microorganisms that facilitate nutrient cycling, and a reduction in the proportion of phytopathogenic fungi. This resulted in a notable enhancement in enzyme activities in the S-UE, S-NP, and S-SC treatments, as well as an increase in the efficacy of soil nitrogen, phosphorus, and potassium. This, in turn, led to significant improvements in the yield and quality of F. unibracteata, including bulb quality, seedling emergence, and bulb nucleoside content.
Thus, it can be concluded that the judicious application of organic fertilizers in the Tibetan Plateau region can effectively alter the composition of the soil bacterial community, enhance soil fertility, and optimize the yield and quality of F. unibracteata plants. This study elucidated the mechanism by which organic fertilizers enhance the yield and quality of F. unibracteata plants, thereby providing a theoretical foundation for standardized cultivation.
This research was financially supported by the Beijing Natural Science Foundation (No. 7254508), Fundamental Research Funds for the Central public welfare research institutes (Grant/award Nos L2023022, Z2023014, ZZ18-YQ-036, ZZ16-XRZ-067, ZXKT22061 and ZXKT25005), and the Scientific and technological innovation project of China Academy of Chinese Medical Sciences (CI2024C003YN). In addition, we thank the anonymous reviewers for their valuable comments regarding the manuscript.
-
The authors confirm contribution to the paper as follows: study conception and design: Wu Y, Yang Z; data collection: Wu Y, Yang Z, Zhang H, Gao X, Gao D ; analysis and interpretation of results: Wu Y, Yang Z, Zhang H, Gao X, Gao D, Li X; draft manuscript preparation: Wu Y, Gao D, Li X. All authors reviewed the results and approved the final version of the manuscript.
-
The 16S and ITS rRNA sequences of soil microorganisms used in this manuscript have been submitted to NCBI under sequence number PRJNA1094691. Most of the data generated and analyzed during the course of the study are included in this paper and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yuhan Wu, Zemin Yang
- Supplementary Table S1 The physical and chemical properties of the cow manure organic fertilizer, sheep manure organic fertilizer, and the soil at the experimental site used in the trial.
- Supplementary Table S2 Emergence rate and leaf area of F. cirrhosa under different treatments.
- Supplementary Table S3 Topological indices of each co-occurrence network in Fig. 5.
- Supplementary Fig. S1 The dilution curve in different treatment. (a: bacterial dilution curve; b: fungal dilution curve).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wu Y, Yang Z, Gao D, Zhang H, Gao X, et al. 2025. Integrative microbiomics and co-occurrence network analyses decipher the mechanisms underlying organic fertilizer-driven yield and nucleoside content enhancement in Fritillaria unibracteata. Technology in Horticulture 5: e018 doi: 10.48130/tihort-0025-0013 

Integrative microbiomics and co-occurrence network analyses decipher the mechanisms underlying organic fertilizer-driven yield and nucleoside content enhancement in Fritillaria unibracteata
- Received: 22 November 2024
- Revised: 20 February 2025
- Accepted: 10 March 2025
- Published online: 07 May 2025
Abstract: Fritillaria unibracteata is a medicinal plant with high economic and medicinal value. The mechanistic understanding of how varying fertilization strategies influence its yield and phytochemical quality however remains largely unexplored. In this study, we aimed to comprehensively evaluate the effects of cow and sheep manures on the soil nutrient enzyme activities, microbiology, yield, and nucleoside content of F. unibracteata. A comprehensive evaluation of the physicochemical properties of the soil, enzyme activities, microbiomics, and co-occurrence networks was conducted at the time of F. unibracteata harvest with the application of cow and sheep manure. Our findings demonstrated that the application of cow manure (24.3 t·ha−1), and sheep manure (43.7 t·ha−1) resulted in remarkable yield increases of 119.45% and 97.22%, respectively. Organic fertilizer treatment significantly increased the concentrations of critical nucleosides—uridine, guanosine, and thymidine—by 119.65%, 105.26%, and 70.20%, respectively. Microbiome profiling and co-occurrence network analyses revealed that organic fertilizer enhanced the functional synergy and structural resilience of soil microbial communities, with notable proliferation of nutrient-cycling taxa, such as Comamonadaceae and Sphingomonadaceae significantly increasing by 31.7% and 27.0%, respectively. Pathogenic fungal taxa, including Heydenia and Solicoccozyma, showed significant reductions of 63.1% and 19.4%, respectively. Structural equation modeling identified the enhancement of soil enzyme activity and soil phosphorus as the principal factors driving the observed increases in F. unibracteata yield and nucleoside accumulation. The application of organic fertilizers enhances the yield and quality of F. unibracteata by recruiting beneficial microorganisms that promote soil nutrient cycling and improve the soil microbiological environment.