






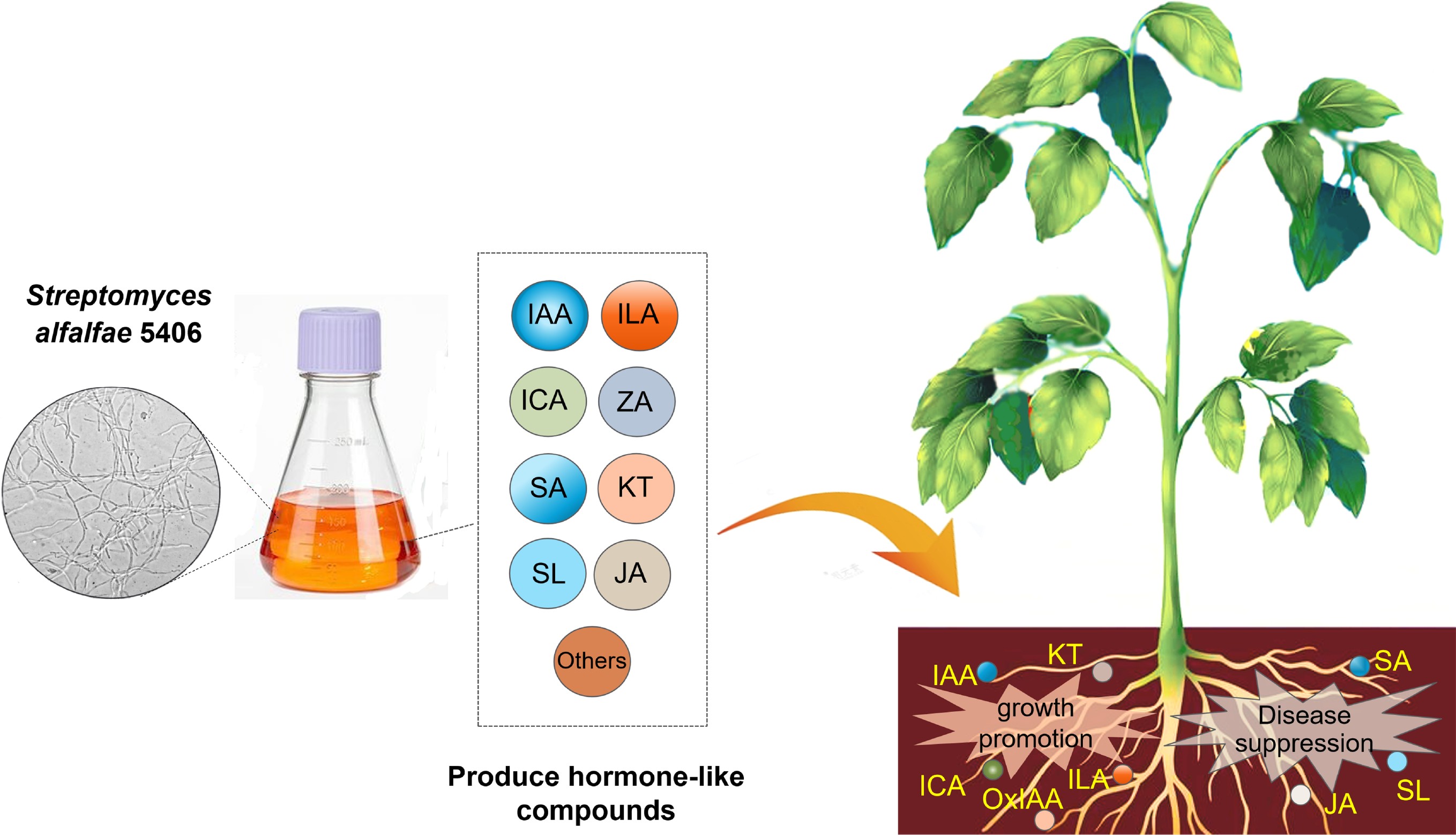



-
Actinomycetes are an important component of the plant rhizosphere microbial community[1]. They are commonly found in soil and are considered plant growth-promoting rhizobacteria (PGPR) due to their ability to produce a wide range of biologically active metabolites that contribute to enhanced plant growth and stress resistance[2]. Streptomyces, the largest and most promising genus among actinomycetes, can utilize precursor compounds from primary metabolism to synthesize diverse secondary metabolites with biological activity. Among these secondary metabolites are phytohormones. Some Streptomyces species secrete hormones such as indole-3-acetic acid (IAA) and salicylic acid (SA), which have the capacity to enhance plant growth and confer resistance to pathogen invasion[3,4].
Streptomyces alfalfa is an excellent example of plant growth-promoting streptomyces species. It was initially identified as Streptomyces microflavus and is commonly known in China as Jingyang streptomyces or Actinomycete 5406, reflecting both the geographic origin of the strain and its associated strain codes[5–7]. This bacterium (hereafter referred to as S. alfalfa 5406) was first isolated in 1953 from the rhizosphere of alfalfa grown in Jingyang County, Shanxi Province of China[5]. The strain name 5406 denotes its origin in northwest China and indicates that it was the sixth strain isolated from the alfalfa rhizosphere[4]. In 2018, the bacterium was re-classified as S. alfalfa based on 16S rRNA gene phylogeny, whole-genome sequencing, and morphological, physiological, and chemotaxonomic analysis conducted by Liang et al.[8,9].
It was found that the metabolites of S. alfalfa 5406 could promote the growth of bean sprouts. Later, its secreted stimulants were shown to be more effective than IAA and gibberellin[10]. Additionally, these stimulants were found to induce earlier sprouting in dormant potatoes and to promote the germination of wheat, sweet potatoes, and other crops[10]. Subsequent research revealed that S. alfalfa 5406 could produce antibiotics in the fermentation broth of a soybean medium, which was effective in preventing and controlling crop rot, as well as diseases such as cotton wilt and yellow wilt[11]. Preliminary extraction confirmed that S. alfalfa 5406 produces two types of antibiotics: one antibacterial and the other antifungal[11]. Notably, the antibacterial compounds were found to be heat-resistant[11]. In 1965, Yin et al. examined the effects of S. alfalfa 5406 on nitrogen and phosphorus transformation and its growth ability in 25 different soil types, including black soil, sandy soil, and cabbage soil. as well as its to elements. They found that S. alfalfa 5406 could increase the levels of available nitrogen and phosphorus in soil[12].
In the 1970s, due to shortages of grain, cotton, and oil in China, there was a high demand for growth-promoting and disease-suppressive agents. As a result, strain 5406 was produced on a large scale across various regions of the country[13,14]. However, most of the manufacturing relied on traditional methods and rudimentary equipment. This led to widespread failures in quality control, with contamination becoming a common issue. Consequently, the agricultural application of S. alfalfa 5406 was discontinued due to the lack of expected economic benefits[14].
S. alfalfa 5406 has recently regained interest due to its promising capabilities in potassium and phosphorus solubilization, disease resistance, and growth promotion, making it a potential biological control agent and biofertilizer[6]. It has been shown to increase the yield of crops such as cucumber, chilli, and tomato[15]. Application of strain 5406 has been documented to promote seed germination and seedling growth in plants including watermelon, eggplant, tomato, and wheat[6,16]. Moreover, its fermentation broth exhibits an inhibitory effect against several Gram-positive bacteria (Bacillus subtilis, Streptococcus lactis, Staphylococcus aureus) and plant pathogens such as Verticillium wilt, Fusarium oxysporum, and F. graminearum[7,17]. Strain 5406 can also improve the yield and quality of peppers, increase the iron content in fruits, and enhance soil enzyme activities such as catalase, peroxidase, urease, and sucrase[18].
Previous work found that S. alfalfa 5406 produces effects similar to those of auxin and cytokinin, which can stimulate both transverse and longitudinal cell growth, promote cell proliferation, and delay the degradation of nucleic acids and proteins in cells. These effects ultimately delay crop aging and increase yield[19]. It was also discovered that metabolites produced by strain 5406 could stimulate chlorophyll synthesis, accelerate plant rooting and germination, and contribute to phosphorus solubilization, potassium solubilization, and nitrogen fixation[20]. In tomato plants, strain 5406 has been shown to promote root elongation and tillering and to improve the photosynthetic capacity of leaves[6]. However, the specific biologically active compounds produced by S. alfalfa 5406 in vivo have not yet been identified.
This study aims to unravel the molecular mechanisms underlying the well-documented plant growth-promoting activities of S. alfalfa 5406. To this end, LC-MS/MS analysis was employed to identify the hormone-like compounds produced by S. alfalfa 5406. Next, experiments were designed to verify the plant growth promotion and disease suppression activities, using both the fermentation broth of strain 5406 and a selected panel of its derived hormonal compounds. The resulting data will allow for discussions about the potential of S. alfalfa 5406 and its associated metabolites for agricultural applications.
-
Banana seedlings with two leaves were pre-cultured for 2–3 d in tubes, each containing 200 mL of nutrient solution. The fermentation broth of S. alfalfa 5406 was added at the final concentrations of 0.1%, 1%, 2%, and 10%. A treatment without fermentation broth was included as a control. Each treatment included 20 replicate plants. The effects of strain 5406 inoculation were assessed based on the fresh weight of the roots and aboveground parts of the banana plants after 30 d of treatment.
Sample preparation for hormonomics analysis
-
Strain 5406 used in this study was obtained from China General Microbiological Culture Collection Center (CGMCC) under the strain number CGMCC4.0891. Single colonies were picked and cultured in PBS medium (potatoes 200 g·L−1, millet 10 g·L−1, soybean meal 10 g·L−1, sucrose 30 g·L−1, pH 7.0; sterilized at 121 °C for 20 min) at 200 rpm in a 28 °C shaker for one week. Tomato plantlets were grown in a 28 °C growth room under a 16 h/8 h light/dark cycle with a light intensity of 5,000 lux. The plantlets were cultured in bottles containing ¼ MS salts medium. For treatment, the roots of the plantlets were immersed in the broth of strain 5406 and incubated on a shaker at 70 rpm and 28 °C for 2 h. Two control treatments were included: strain 5406 cultured in PBS medium without plant exposure and PBS medium without any bacterial inoculation. For each sample, 100 mL of liquid was harvested, filtered through a 0.22 μm membrane, flash-frozen in liquid nitrogen, and stored at −80 °C until use.
The reagents used in this work are listed as follows: MilliQ water (Millipore, Bradford, USA) was used in all experiments. HPLC-grade acetonitrile (ACN) and methanol (MeOH) were obtained from Merck (Darmstadt, Germany). HPLC-grade formic acid was purchased from Aladdin. All analytical standards were obtained from Olchemim Ltd. (Olomouc, Czech Republic) and isoReag (Shanghai, China). Acetic acid and formic acid were from Sigma-Aldrich (St Louis, MO, USA). Stock solutions of the standards were prepared at a concentration of 1 mg/mL in MeOH and stored at −20 °C. Before analysis, the stock solutions were diluted with MeOH to the appropriate working concentrations.
Sample extraction
-
At the time of harvest, the samples were immediately frozen in liquid nitrogen and stored at −80 °C until use. A 50 μL aliquot of each sample was mixed with 1 mL methanol/water/formic acid (15:4:1, V/V/V). Then, 10 μL of the internal standard mixture (100 ng·mL−1) was added to the extract for quantification. The mixture was vortexed for 10 min, followed by centrifugation at 12,000 rpm for 5 min at 4 °C. The supernatant was transferred to clean plastic microtubes, evaporated to dryness, and re-dissolved in 100 μl of 80% methanol (V/V). The resulting solution was filtered through a 0.22 um membrane filter for subsequent LC-MS/MS analysis[21,22].
LC-MS/MS analysis
-
The sample extracts were analyzed using a UPLC-ESI-MS/MS system (UPLC: ExionLCTM AD,
https://sciex.com.cn ; MS: Applied Biosystems 6500 Triple Quadrupole,https://sciex.com.cn ). HPLC conditions: Column, Waters ACQUITY UPLC HSS T3 C18 (100 mm × 2.1 mm i.d., 1.8 µm); solvent system, water with 0.04% acetic acid (A), acetonitrile with 0.04% acetic acid (B); gradient program: 5% B (0–1 min), increased to 95% B (1–8 min), held at 95% B (8–9 min), then returned to 5% B (9.1–12 min); Flow rate: 0.35 mL/min; Column temperature: 40 °C; Injection volume: 2 μL[23−25].ESI-MS/MS conditions
-
Linear ion trap (LIT) and triple quadrupole (QQQ) scans were acquired using a triple quadrupole-linear ion trap mass spectrometer (QTRAP), the QTRAP®6500+ LC-MS/MS system, equipped with an ESI Turbo IonSpray interface. The system operated in both positive and negative ion modes and was controlled by software Analyst 1.6.3 (Sciex). ESI-MS/MS conditions: ion source, ESI±; source temperature 550 °C; ion spray voltage (IS) 5,500 V (positive), –4,500 V (negative); curtain gas (CUR), 35 psi.
Phytohormones were analyzed using scheduled multiple reaction monitoring (MRM). Data acquisition was performed with Analyst 1.6.3 software, and quantification of all metabolites was conducted using MultiQuant 3.0.3 software (Sciex). Mass spectrometer parameters, including the declustering potentials (DP) and collision energies (CE) for individual MRM transitions, were optimized for each MRM transition. A specific set of MRM transitions was monitored during each period according to the retention time of eluting metabolites[26,27].
Standard curve preparation for hormone quantification
-
Standard solutions were prepared at the following concentrations: 0.01, 0.05, 0.1, 0.5, 1.0, 5.0, 10, 50, 100, 200, and 500 ng·mL−1. For TRP and SAG, the concentrations were 20 times higher than those listed above, resulting in a standard curve concentration range of 0.2–10,000 ng·mL−1. The linear equation and correlation coefficient of the standard curve for a total of 88 tested compounds are provided in Supplementary Table S1.
KEGG annotation and enrichment analysis
-
Identified metabolites were annotated using the KEGG Compound Database (
www.kegg.jp/kegg/compound ), and the annotated metabolites were subsequently mapped to the KEGG Pathway Database (www.kegg.jp/kegg/pathway.html )[28]. Pathways containing significantly regulated metabolites were then subjected to metabolite sets enrichment analysis (MSEA), and their significance was determined using p-values from the hypergeometric test.Antibacterial test
-
To prepare the pathogenic bacterial plates, Ralstonia solanacearum colonies were picked and cultured in the Luria-Bertani (LB) broth at 28 °C with shaking at 180 rpm for 12–16 h. Then, 100 μL of the bacterial solution was evenly spread onto LB agar plates. The antibacterial test was performed using the hole-punching method. Wells (Φ = 8 mm) were created using 1 mL pipette tips, and 100 μL of the sterile supernatant of strain 5406, PBS medium, and the salicylic acid (SA, 20 mg·mL−1), each filtered through a 0.22 μm membrane, was added to the wells after removing the excised agar. The plates were incubated at 30 °C for 48 h, and the diameter of the antibacterial zone was measured using the crossover method.
Effects of hormone substances on plant growth
-
Tomato seedlings with two leaves were selected and placed into tubes with 45 mL of plant culture medium (Hoagland Nutrient Solution) for hydroponic cultivation. After approximately two weeks, once the seedlings were stably growing, hormone substances produced by S. alfalfa 5406 were added to the culture solution to a final concentration of 5 μg·mL−1. Each treatment included 20 replicates. The water-treated plants were used as the control. After 30 d of cultivation, the tomato seedlings were evaluated for root length, aboveground weight, fresh weight, dry weight, and the number of lateral roots.
Statistical analysis
-
A multiple comparison test (Tukey's HSD) was conducted to determine the significant differences among treatments using SPSS version 18.0 (SPSS Inc., Chicago, IL, USA).
-
In previous studies, it was found that strain 5406 could promote the growth of tomato and that 1% of the fermentation broth significantly enhanced both root and shoot development in tomato seedlings[6]. To further confirm the growth-promoting effects of strain 5406, another plant species, banana, was selected for similar experiments. The results indicated that 0.1%, 1%, and 2% of the fermentation broth all significantly promoted the growth of banana seedlings (Fig. 1). Among these, the 1% concentration showed the most pronounced growth-promoting effect. In contrast, a 10% concentration of the fermentation broth inhibited the growth of banana seedlings.

Figure 1.
Growth effect of S. alfalfae 5406 fermentation broth on banana seedlings. (a) Soil extracts. (b) 0.1% concentration of the fermentation broth. (c) 1% of the fermentation broth. (d) 2% of the fermentation broth. (e) 10% of the fermentation broth. (f) Root and shoot weight of the banana seedlings. Different letters above the bars indicate significant differences between samples at the level of p < 0.05.
Hormonomics analysis of the culture filtrates of strain 5406
-
Phytohormones are typically unstable, easily decomposed, and sensitive to temperature and environmental conditions, so the phytohormones from strain 5406 were analyzed separately. A total of 88 hormone substances were quantitatively analyzed, of which 66 were detected across all samples. Specifically, 60, 48, and 48 hormone substances were identified in the media, strain 5406, and the treatment samples, respectively. The identities and concentrations of these hormones are listed in Supplementary Table S2. The numbers of hormone substances showing statistically significant changes between sample groups are illustrated in the Venn diagrams (Fig. 2). There were 57 significantly altered hormone substances in both the media vs strain 5406 and media vs treatment comparisons, and 28 in the strain 5406 vs treatment comparison. Overall, relatively few substances were found to be up-regulated or down-regulated in the strain 5406 vs treatment comparison, whereas a larger number showed altered levels in the media vs strain 5406 and media vs treatment comparisons.
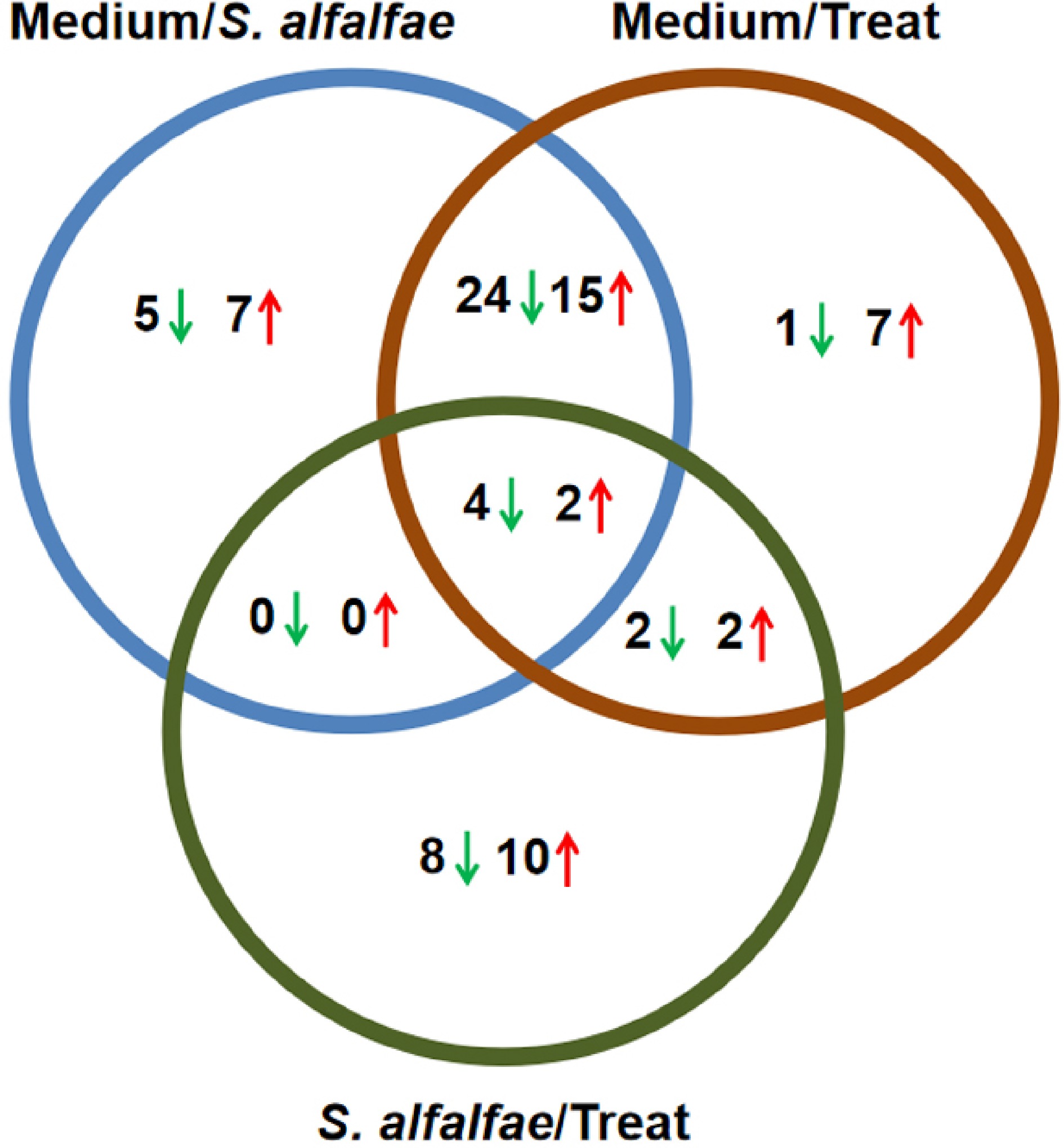
Figure 2.
Venn diagram for differentially expressed hormone-like substances. The numbers represent differentially expressed substances identified from comparisons among the medium, S. alfalfae 5406, and the treatment group. Green arrows denote down-regulated substances, while red arrows indicate up-regulated ones.
Next, cluster analysis of metabolites and samples was performed. As shown in Fig. 3, the sample from S. alfalfa 5406 and the treated group clustered together, while the media sample formed a separate cluster. The metabolite contents of JA-Phe, OPC-6, BAP, GA15, TRA, cZROG, tZ, 2MeSiPR, IPA, GA3, MEJA, cZ, IAA-Leu-Me, pTR, iP7G, oT, DHZROG, DHZ7G, IPR, tZOG, OPDA, SAG, TRP, tZR, GA4, ACC, cZ9G, ICAld, GA24, DHZR, IAA-GI, JA-ILE, and ABA were high in the media but very low in strain 5406 and the treated samples. This suggests that these metabolites were either utilized by strain 5406 or absorbed by the tomato plants. Conversely, certain hormones that were not detected or present at low levels in the media were found at high levels in strain 5406 and the treated group., indicating that these metabolites were produced by the bacterial strain 5406. Additionally, some hormones were detected at high levels in strain 5406 but at low levels in the treated group, suggesting that these metabolites were synthesized by the strain but utilized by the tomato plants.
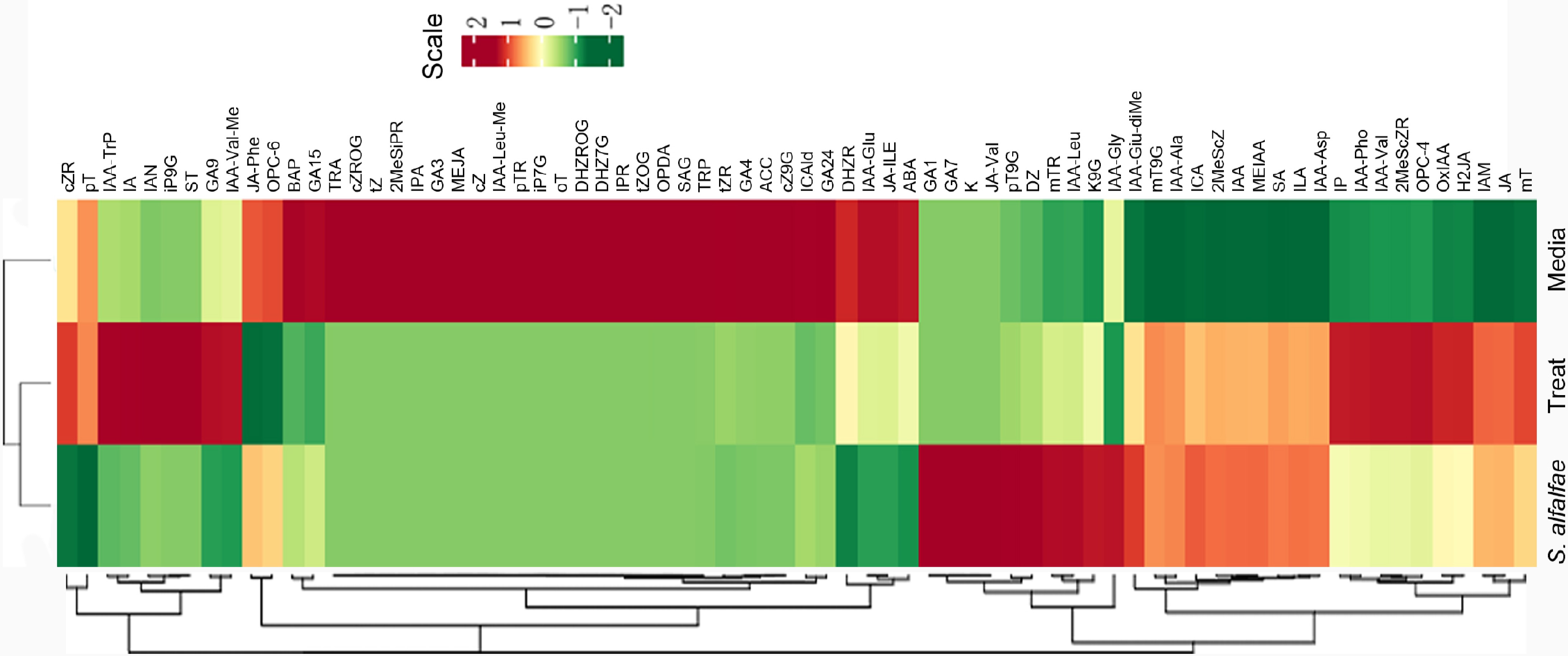
Figure 3.
Cluster analysis of metabolites and samples. The horizontal axis represents the sample name, while the vertical axis shows metabolite information. Colors indicate standardized relative abundance values, with red representing high content and green representing low content. The dendrogram on the left displays metabolite clustering, and the dendrogram at the top shows sample clustering.
Based on KEGG enrichment analysis of the differential metabolites between samples (Fig. 4), in the comparison between S. alfalfa 5406 broth and the treated group, the most significantly enriched pathway was the biosynthesis of secondary metabolites, followed by the general metabolic pathway and then the plant hormone signal transduction pathway. Overall, very similar enrichment patterns of differential metabolites were observed between the media vs strain 5406 and media vs treated groups. This suggests that the metabolite profiles of strain 5406 and the treated samples were quite similar.
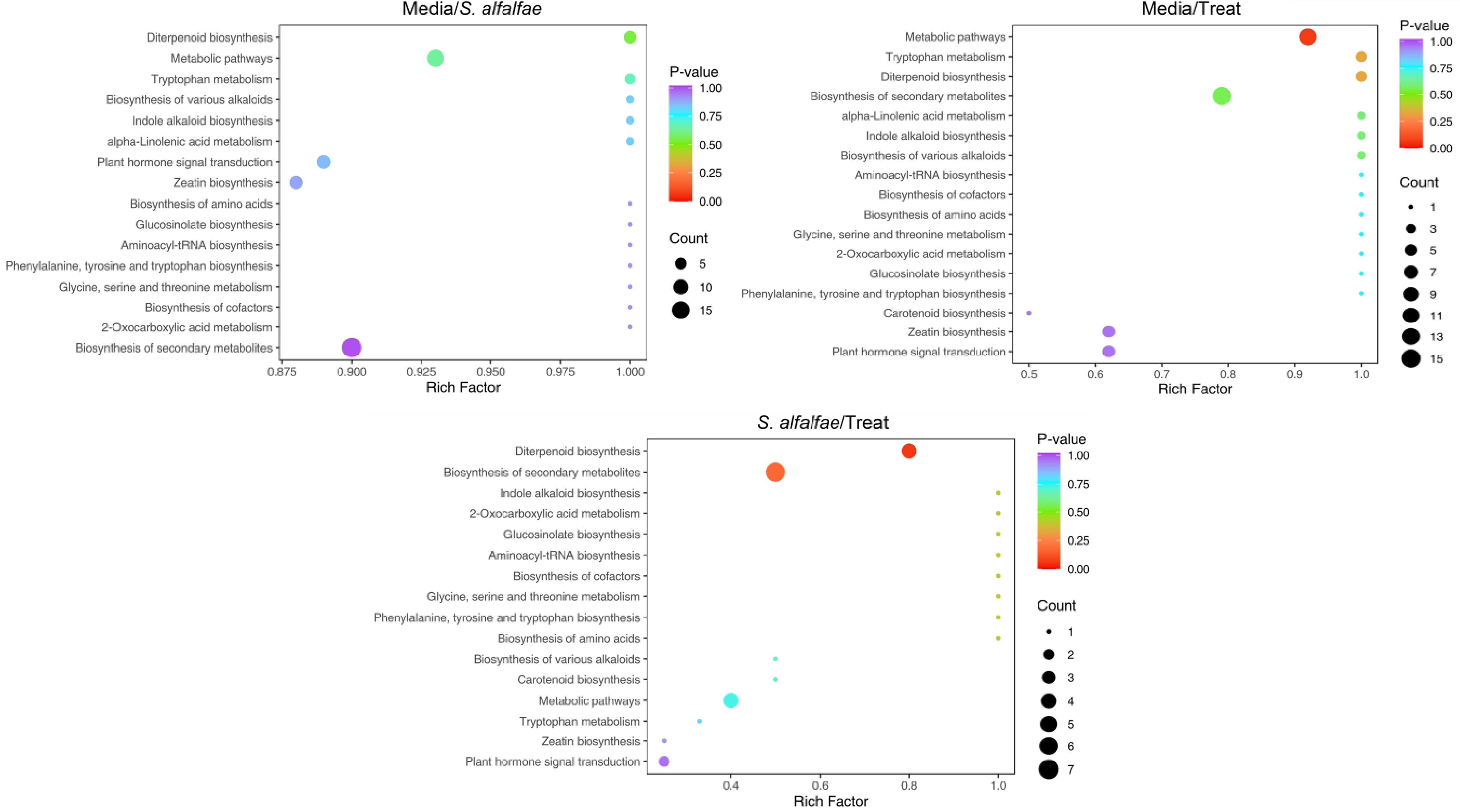
Figure 4.
KEGG enrichment analysis of differential metabolites between samples. The x-axis represents the enrichment factor to each pathway, and the y-axis lists the pathway names. The color of each dot indicates the p-value, with redder colors representing more significant enrichment. Dot size reflects the number of enriched differential metabolites.
Though analysis of the KEGG metabolic pathway (Fig. 5), compared with media, auxin, salicylic acid (SA), and jasmonic acid (JA) were up-regulated in strain 5406 in the biosynthesis of secondary metabolites pathway (Fig. 5a). In the tryptophan metabolism pathway (Fig. 5b), IAA and IAM were up-regulated, while TRP and TRA downregulated (Fig. 5b). Salicyiate was up-regulated in the biosynthesis of various alkaloids pathway (Fig. 5c). JA was up-regulated, while MEJA was down-regulated in the α-linolenic acid metabolic pathway (Fig. 5d). Most zeatin compounds were down-regulated, while dihydrozeatin was up-regulated in the zeatin biosynthesis pathway (Fig. 5e). GA1 and GA7 were up-regulated, whereas GA3, GA4, GA9, and GA24 were down-regulated in the diteroenoid biosynthesis pathway (Fig. 5f).
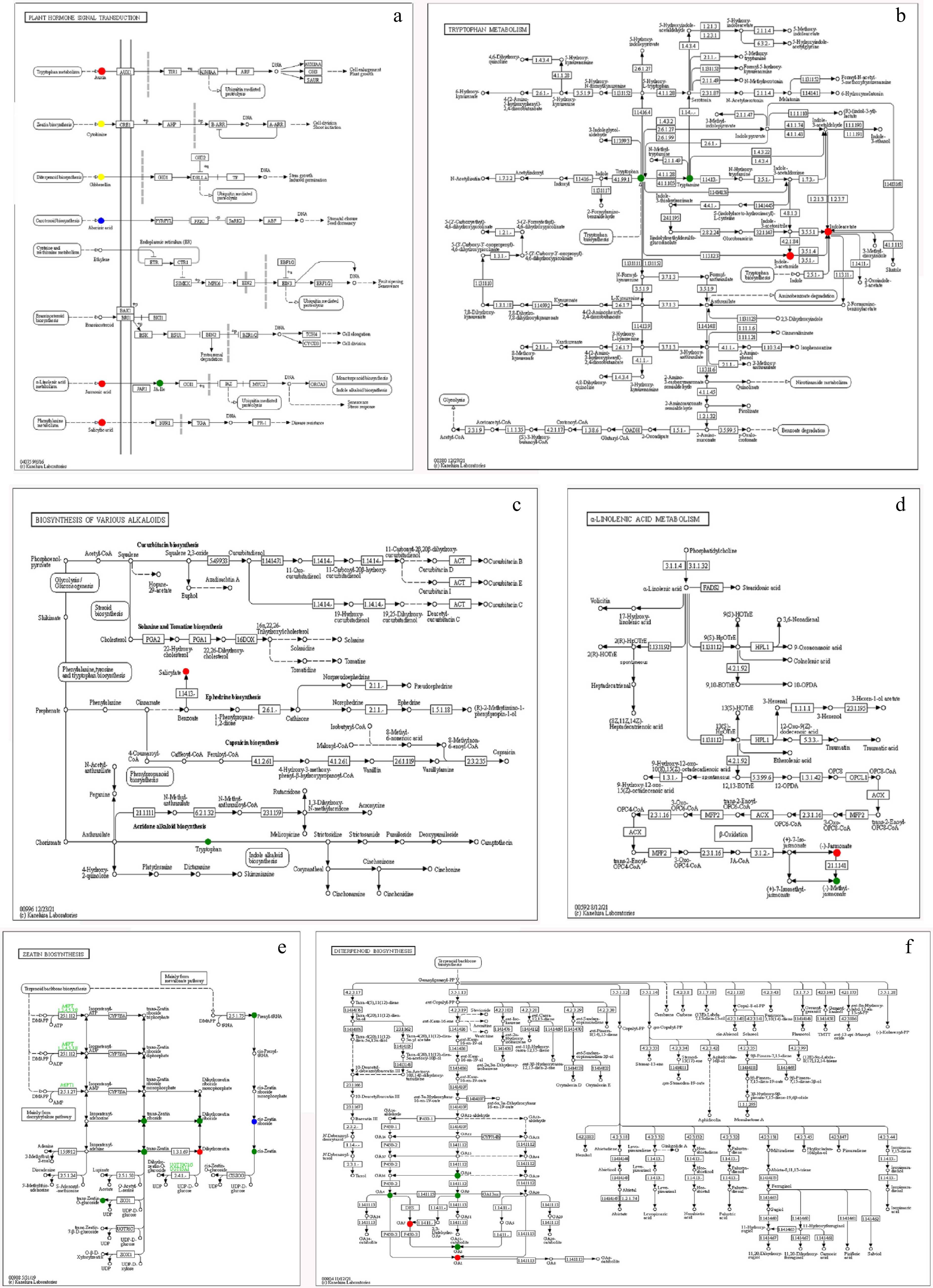
Figure 5.
Metabolite KEGG pathway map of metabolites differentially detected in medium vs S. alfalfae 5406 treatments. Significantly affected pathways include biosynthesis of (a) secondary metabolites, (b) tryptophan metabolism, (c) biosynthesis of various alkaloids, (d) α-linolenic acid metabolism, (e) zeatin biosynthesis, and (f) diterpenoid biosynthesis. Red indicates metabolites that were significantly up-regulated in the S. alfalfae 5406 sample; blue indicates metabolites that were detected but not significantly changed; and green indicates significantly down-regulated metabolites in the S. alfalfae sample.
The main hormone substances produced by S. alfalfae 5406
-
The differential hormone substances produced by strain 5406 are listed in Table 1. Compared with the media, the differential hormones included auxins, cytokinins, SA, JA, ethylene, and strigolactones. The main up-regulated substances associated with growth promotion were auxins, cytokinins, and strigolactones. Among the auxins, IAA-Ala and IAA-Glu-diMe were not detected in the media but were present in both the strain 5406 broth and the treated samples (Table 2), indicating that these substances were produced by strain 5406. IAM, ILA, and ICA were found at levels more than 10-fold higher in the strain 5406 broth and the treated samples compared to the media. IAA levels were also over 5-fold higher, reaching 153.83 ng·mL−1 in the strain 5406 broth and 134.50 ng·mL−1 in the treated samples, confirming that strain 5406 produced these auxins in substantial amounts. However, tryptophan and tryptamine, precursors for IAA synthesis, were abundant in the media but greatly reduced in both the strain 5406 broth and the treated samples, suggesting that strain 5406 utilized these two compounds for auxin production.
Table 1. Summary of selected differential hormone substances.
Compounds Index Medium (ng·mL−1) S. alfalfae (ng·mL−1) Treat (ng·mL−1) Auxin L-tryptophan TRP 11,509.8 60.0 169.4 Tryptamine TRA 1.94 N/A N/A N-(3-Indolylacetyl)-L-alanine IAA-Ala N/A 0.94 0.91 Indole-3-acetyl-L-glutamic acid dimethyl ester IAA-Glu-
diMeN/A 0.73 0.48 2-oxindole-3-acetic acid OxIAA 53.06 95.49 132.37 Methyl indole-3-acetate MEIAA 0.06 0.74 0.64 Indole-3-acetic acid IAA 20.55 153.83 134.50 3-Indole acetamide IAM 0.20 2.60 3.07 Indole-3-carboxaldehyde ICAld 6.90 1.53 0.77 Indole-3-lactic acid ILA 7.33 81.41 73.07 Indole-3-carboxylic acid ICA 6.04 84.60 68.53 Cytokinin Kinetin-9-glucoside K9G N/A 1.79 0.77 N6-Isopentenyl-adenine-9-glucoside iP9G N/A N/A 0.69 cis-Zeatin-O-glucoside riboside cZROG 0.26 N/A N/A N6-isopentenyladenosine IPR 0.40 N/A N/A N6-Isopentenyl-adenine-7-glucoside iP7G 3.47 N/A N/A trans-Zeatin-O-glucoside tZOG 1.84 N/A N/A 2-Methylthio-cis-zeatin riboside 2MeScZR 0.07 3.84 9.28 Dihydrozeatin-7-glucoside DHZ7G 0.12 N/A N/A 4-[(9-beta-D-glucopyranosyl-9H-purin-6-yl)amino]methyl]phenol pT9G 0.07 0.15 0.08 cis-Zeatin riboside cZR 0.28 0.21 0.32 6-Benzyladenine BAP 0.30 0.08 0.03 meta-Topolin riboside mTR 0.01 0.16 0.06 trans-Zeatin riboside tZR 0.12 0.02 0.03 2-Methylthio-cis-zeatin 2MeScZ N/A 0.29 0.25 SA Salicylic acid SA 65.90 317.23 298.24 Salicylic acid 2-O-β-glucoside SAG 229.79 0.65 0.33 JA Dihydrojasmonic acid H2JA 0.68 0.76 0.84 Jasmonic acid JA 0.75 1.69 1.86 Methyl jasmonate MEJA 1.26 N/A N/A Jasmonoyl-L-isoleucine JA-ILE 0.30 0.12 0.18 cis(+)-12-Oxophytodienoic acid OPDA 15.90 0.07 0.03 3-oxo-2-(2-(Z)-Pentenyl) cyclopentane-1-butyric acid OPC-4 N/A 5.45 14.41 3-oxo-2-(2-(Z)-Pentenyl)cyclopentane-1-hexanoic acid OPC-6 14.78 11.73 3.81 GA Gibberellin A3 GA3 0.48 N/A N/A Gibberellin A4 GA4 3.52 N/A 0.16 Gibberellin A1 GA1 N/A 1.82 N/A Gibberellin A15 GA15 0.26 0.07 N/A Gibberellin A24 GA24 0.49 0.21 0.19 SL Strigol ST N/A N/A 0.58 ETH 1-Aminocyclopropanecarboxylic acid ACC 273.16 8.12 17.59 The numbers in bold represent the changes that are 5-fold or higher. 'N/A' indicates that the substance is not detected in this sample. Table 2. Antibacterial effects of S. alfalfae 5406 and SA treatments against R. solanacearum.
Strain/standard substance Diameter of inhibition zone (cm) S. alfalfae cakes 2.65 ± 0.30 S. alfalfae sterile supernatant 2.08 ± 0.13 SA 2.77 ± 0.22 PBS medium 0 The diameter of the inhibition zone are mean ± standard deviation. In terms of cytokinins, substances such as K9G and 2MeScZ were not detected in the media but were present in both the strain 5406 broth and treated samples. Additionally, 2MeScZR was more than 10-fold higher in the broth and treated samples compared to the media, indicating these were also synthesized by strain 5406. Interestingly, iP9G was not detected in the media or the strain 5406 broth but appeared in the treated samples, suggesting that plant interaction may be required to induce its production by strain 5406. The cytokinin mTR was present at low levels in the media but accumulated in strain 5406 broth and was nearly absent in the treated samples, suggesting that mTR was produced by the strain but utilized by the plant.
Furthermore, ACC (1-aminocyclopropane-1-carboxylic acid), a precursor of ethylene, was abundant in the media (273.16 ng·mL−1) but significantly lower in the strain 5406 broth (8.12 ng·mL−1) and the treated samples (17.59 ng·mL−1). This suggests that strain 5406 may reduce ethylene levels by degrading ACC, thereby promoting plant growth.
It was also found that SA, JA, and strigol (hormones associated with plant resistance) were produced by strain 5406 (Table 1). The content of SA was significantly higher in the strain 5406 broth (317.23 ng·mL−1) and in the treated samples (298.24 ng·mL−1) compared to the media (65.9 ng·mL−1). This suggests that the strain may use salicylic acid 2-O-β-glucoside (SAG) and cis-(+)-12-oxophytodienoic acid (OPDA) as precursors to synthesize SA and JA, thereby enhancing plant stress resistance.
Effects of several hormone substances on the growth of tomato seedlings
-
To verify the effects of hormone-related substances produced by S. alfalfae on tomato growth, the compounds mTR, IAA-Ala, and ILA were selected to treat tomato seedlings. The results showed that ILA significantly promoted plant growth (Fig. 6). The number of lateral roots (Fig. 6j), root length, and shoot length (Fig. 6h), as well as the fresh and dry weights (Fig. 6i) of plants treated with ILA, were all greater than those of the control (Fig. 6d). In contrast, plants treated with mTR (Fig. 6a, e), IAA-Ala (Fig. 6b, f) exhibited slower growth compared to the control (Fig. 6). The inhibitory effects of IAA-Ala on plant growth may be due to its role as a conjugated form of IAA. Compounds such as IAA-Ala, IAA-Asp,and IAA-Glu are typically considered storage or inactive forms of IAA and are involved in regulating free IAA levels in plants[29].
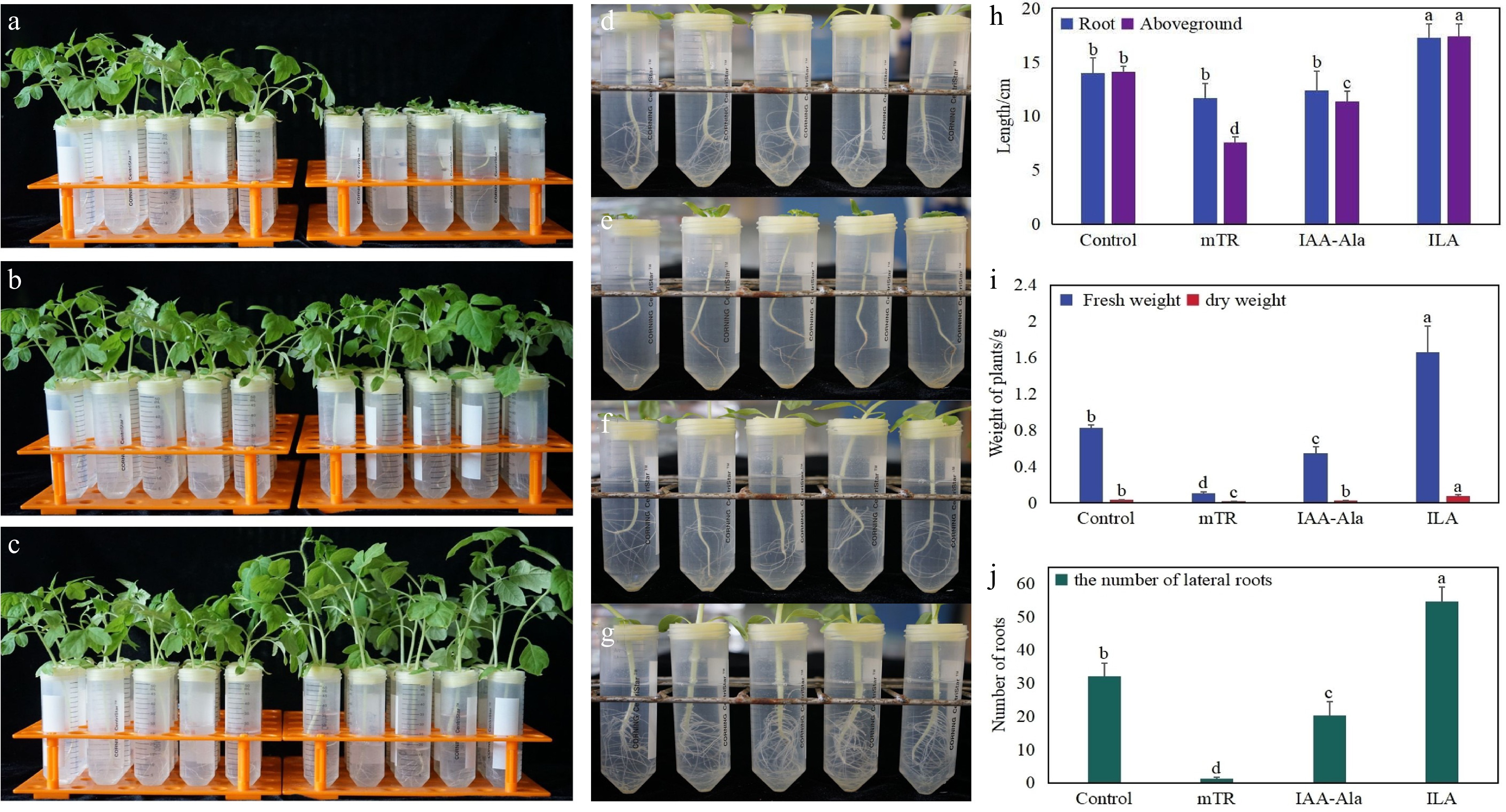
Figure 6.
Effect of three candidate metabolites on the growth of tomato seedlings. (a) Growth of seedlings treated with culture medium (left) and mTR (right). (b) Growth of seedlings treated with culture medium (left) and IAA-Ala (right). (c) Growth of seedlings treated with culture medium (left) and ILA (right). (d) Growth of roots in culture medium. (e) Growth of roots treated with mTR. (f) Growth of roots treated with IAA-Ala. (g) Growth of roots treated with ILA. (h) Statistical analysis of root length and shoot weight. (i) Statistical analysis of fresh weight and dry weight. (j) Statistical analysis of lateral root number. Different letters in the figure indicate significant differences between treatments at p < 0.05.
Antibacterial effects of S. alfalfae 5406 on R. solanacearum
-
To confirm the effects of S. alfalfae against pathogenic bacteria, an antagonistic experiment was conducted to evaluate its activity against R. solanacearum, the causative agent of tomato bacterial wilt. Additionally, the effect of SA, a compound known to be produced by strain 5406, was also accessed. The results are shown in Fig. 7 and Table 2. strain 5406 (Fig. 7a), its sterile supernatant (Fig. 7b), and SA (Fig. 7d) all significantly inhibited the propagation of R. solanacearum. The diameters of the inhibition zones were 2.65 ± 0.30, 2.08 ± 0.13, and 2.77 ± 0.22 cm, respectively (Table 2). In contrast, the controls (R. solanacearum treated with PBS broth [Fig. 7c] or with no additive) showed robust growth. These results indicated that S. alfalfae inhibits the propagation of R. solanacearum, likely through the production of antibacterial substances such as SA, which appears to play a key role.
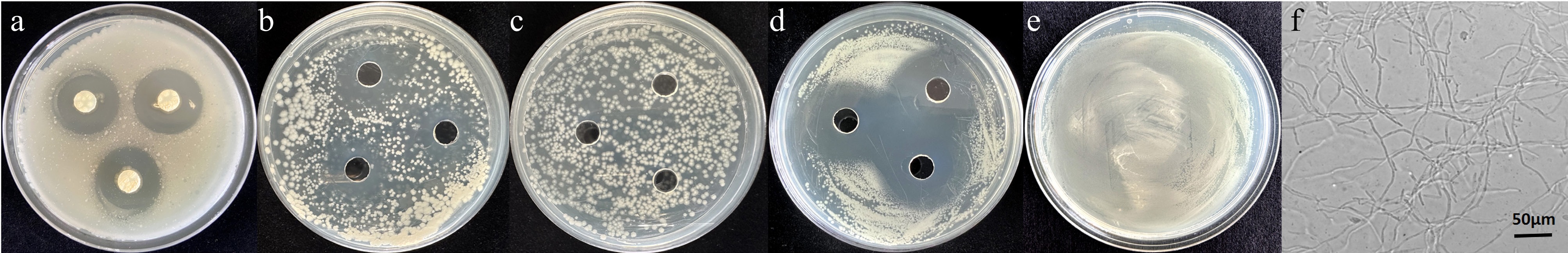
Figure 7.
Antibacterial effects of S. alfalfae 5406 and salicylic acid against R. solanacearum. (a) S. alfalfae 5406 mycelial discs placed on LB agar inoculated with R. solanacearum. (b) Sterile supernatant of S. alfalfae 5406 added to wells in LB agar inoculated with R. solanacearum. (c) Control of R. solanacearum with PBS broth added to the wells. (d) SA added to wells in LB agar with R. solanacearum. (e) Control of R. solanacearum only, without additives. (f) Microscopic morphology of S. alfalfae 5406.
-
In this study, it is demonstrated that the plant growth-promoting bacterium S. alfalfae 5406 can produce IAA, a compound known to stimulate plant cell division and elongation[30,31]. The genome of strain 5406 was sequenced by She et al.[9]. It is appropriately 8 Mb in size with a G + C content of 71.8%. The assembled genome contained 68 contigs, three repeat sequences, 85 tRNA genes, 7,174 CDS, one tmRNA, and 626 signal peptides, as revealed through gene annotation. The genome harbors a large number of genes coding for enzymes involved in the biosynthesis of bioactive substances. Genes associated with plant growth promotion were identified in S. alfalfae 5406, including those encoding indole-3-glycerol phosphatese and tryptophan metabolism-related enzymes. The empirical data presented in this study provide supporting evidence for the genomic predictions.
ILA is an indole metabolite produced through the decomposition of tryptophan. ILA can enhance the expression of key enzymes involved in tryptophan metabolism and promote the synthesis of other indole derivatives, such as indole-3-propionic acid and indole-3-acetic acid[32]. It has been reported to possess anti-inflammatory and potential antiviral properties[33,34]. In this study, it was also found that S. alfalfae 5406 can produce ILA and confirmed that ILA significantly promotes the growth of tomato seedlings. However, in Pisum sativum L., ILA exhibits little to no auxin-like activity within physiological concentration ranges[35]. Indole-3-carboxylic acid (ICA) was also found to be produced by S. alfalfae 5406. ICA has demonstrated antifungal activity, such as against Leptosphaeria biglobosa[36].
In addition, the study found that S. alfalfae 5406 could produce the substances 2MeScZ, K9G, 2MeScZR, and mTR. These compounds can influence plant growth by regulating the levels of zeatin or kinetin[37].
SA functions not only as a signaling molecule but also as an active compound involved in disease suppression and stress resistance in plants. In addition to R. solanacearum, SA has been shown to inhibit many other pathogens, including Staphylococcus aureus, Escherichia coli, Bacillus subtilis, Fusarium oxysporum f. sp. Vasinfectum, Verticillium dahliae, F. oxysporum f. sp. Cubense, Colletotrichum gloeosporioides[4,38]. As is well known, JA and strigol, also produced by S. alfalfae strain 5406, are associated with plant defense responses. These findings enhance our understanding of the mechanisms behind the plant growth-promoting and disease-resistance effects of S. alfalfae, and provide a foundation for its future application in agriculture.
-
Hydroponic experiments confirmed that Streptomyces alfalfae 5406 can significantly influence plant growth. Specifically, a 1% concentration of the strain’s broth markedly promoted banana seedling development, while a higher concentration (10%) exhibited inhibitory effects. Hormonomics analysis using UPLC-MS/MS further revealed that S. alfalfae 5406 produces a diverse array of plant hormones, including auxins such as IAA, ILA, and ICA; cytokinins like kinetin and zeatin, as well as salicylic acid (SA) and jasmonic acid (JA). Antagonistic plate assays demonstrated that both the live strain and its sterile supernatant strongly inhibited the growth of Ralstonia solanacearum, with SA specifically showing potent inhibitory activity—suggesting its involvement in disease resistance. Additionally, ILA produced by the strain was found to enhance root branching, elongation, and shoot growth, further supporting its role in promoting plant development. Collectively, these findings shed light on the dual functional role of S. alfalfae 5406 in stimulating plant growth and suppressing pathogenic threats, offering promising potential for its application in sustainable agriculture.
This work was supported by the Hainan Provincial Natural Science Foundation of China. (Grant Nos 325RC820 and 423MS116) and Central Public-interest Scientific Institution Basal Research Fund. (Grant No. 1630042025014).
-
The authors confirm their contributions to the paper as follows: study conception and design: Li C, Zhang L; data collection: Zheng Y, Zhang Q, Sun J, Wang H, Tan Y, Chen H; analysis and interpretation of results: Zheng Y, Li C, Peng M; draft manuscript preparation: Zheng Y, Li C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed in this study are included in the article or its supplementary information.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0022)
-
Received 3 March 2025; Accepted 27 May 2025; Published online 4 September 2025
-
S. alfalfae 5406 can influence plant growth.
S. alfalfae 5406 produces various plant hormones, including auxin (e.g., IAA, ILA, ICA), cytokinin (kinetin and zeatin), SA, JA, and others.
Both S. alfalfae 5406 and its sterile supernatant significantly inhibited the growth of the R. solanacearum pathogen. SA may contribute to resistance against R. solanacearum.
ILA produced by S. alfalfae 5406 significantly promoted plant growth by enhancing root branching, root elongation, and shoot development.
- Supplemental Table S1 The linear equation and correlation coefficient of the standard curve for detected substances.
- Supplemental Table S2 The compounds identified in samples.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng Y, Zhang Q, Sun J, Wang H, Tan Y, et al. 2025. Streptomyces alfalfae 5406 promotes plant growth and enhances disease resistance by producing hormone production. Tropical Plants 4: e031 doi: 10.48130/tp-0025-0022 

Streptomyces alfalfae 5406 promotes plant growth and enhances disease resistance by producing hormone production
- Received: 03 March 2025
- Revised: 21 May 2025
- Accepted: 27 May 2025
- Published online: 04 September 2025
Abstract: Streptomyces alfalfae 5406 is a plant growth-promoting rhizobacterium widely used as a microbial fertilizer in agricultural production. However, the mechanisms of action and the detailed composition of its bioactive compounds remain unclear. This study analyzed the hormone-like substances produced by S. alfalfae 5406 and confirmed its plant growth-promoting effects using banana seedlings. The results showed that 1% S. alfalfae 5406 fermentation broth significantly promoted banana seedling growth, while 10% inhibited growth. Hormonomics analysis using UPLC-MS-MS revealed the presence of various plant hormones, including auxin, cytokinin, salicylic acid (SA), and jasmonic acid (JA). Plate antagonism assays demonstrated that S. alfalfae 5406 and its sterile supernatant could strongly inhibit the growth of Ralstonia solanacearum, a causal agent of bacterial wilt. SA produced by S. alfalfae 5406 also showed significant inhibition of R. solanacearum, suggesting that SA plays a potential role in disease resistance mediated by strain 5406. In addition to the well-known indole-3-acetic acid (IAA), this study is the first to report that S. alfalfae 5406 produces indole-3-lactic acid (ILA), which significantly enhances plant growth by promoting root branching, root elongation, and shoot development. Together, these findings improve our understanding of the mechanisms by which S. alfalfae 5406 promotes plant growth and confers disease resistance, laying a foundation for its broader application in sustainable agriculture.