









-
Cassava (Manihot esculenta Crantz) is one of the most important tuber crops grown in tropical and subtropical countries[1]. Cassava is the third most important source of carbohydrates for human consumption after rice and maize, and it is a significant source of income in Africa, Asia, and Latin America[2]. In 2022, global cassava farming reached 32.04 million hectares, producing 330.4 million tons with a yield of 10.3 tons per hectare[3]. Cassava varieties are categorized as sweet or bitter, depending on the cyanide content in the tubers. The sweet variety is used as a staple or subsidiary food, and the bitter variety is used to feed livestock and as a raw material for starch-based industries[2]. In addition, cassava is an ideal crop for bioenergy, biomaterials, and animal feed, given its favourable agricultural characteristics and high quantity and quality of starch[4]. Cassava also has medicinal properties. Since 2000, cassava demand has steadily increased, driving the growth in annual production. However, its potential yield remains constrained by susceptibility to biotic and abiotic stresses[5]. In particular, viral, fungal, and bacterial diseases cause severe yield losses.
Potexviruses have a broad geographical distribution, mirroring the range of their hosts and the impact of global commerce[6]. While each Potexvirus species has a naturally limited host range, some can infect multiple plant species under experimental conditions[6]. Cassava (M. esculenta Crantz) has been infected by several potexviruses, including Cassava common mosaic virus (CsCMV)[7], Cassava Caribbean mosaic virus (CsCaMV)[8], Cassava Colombian symptomless virus (CsCSV)[9], and Cassava virus X (CsVX)[10]. CsCaMV was identified in plants exhibiting stunted growth, malformed curled leaves, and a bright yellow mosaic in Colombia's North Coast, where it was associated with yield losses of 35%–39%[11]. Despite displaying similar flexuous particles, symptomatic samples did not react to CsCMV or CsVX antisera[12,13]. CsCSV was found in co-infections with CsVX in plants exhibiting cassava frogskin disease (CFSD) and/or CsCaMV in Colombia. While CsCSV can infect N. benthamiana, it causes only local infections[14], and infected plants do not react to CsVX or CsCMV antisera[13]. CsCaMV and CsCSV may represent earlier detections of what was previously known as cassava new alphaflexivirus (CsNAV)[15]. Consequently, it was recommended that only CsCMV, CsVX, and CsNAV be included in quarantine lists[16]. However, only CsCMV, CsCSV, and CsVX are recognized members of Potexvirus[17]. CsVX was first discovered as flexuous particles in CFSD-symptomatic plants in Colombia[18]. While CsVX preparations have not cross-reacted with CsCMV antiserum, they reacted weakly with Tulip virus X (TuVX) and Potato virus X (PVX) antisera[8]. CsVX was transmitted at a significantly lower frequency than CsCMV and was detected in N. benthamiana using enzyme-linked immunosorbent assay (ELISA), but there has been no evidence of the mechanical transmission of CsNAV[9,15]. Cassava plants infected with CsCMV developed leaf mosaic symptoms under greenhouse conditions, and the virus was mechanically transmitted to N. benthamiana with 90%–100% efficiency in two independent experiments[15,19]. CsVX-infected plants exhibited milder symptoms than those infected with CsCMV[15]. We speculate that CsVX, CsCaMV, and CsCSV may require interactions with other viruses, such as CFSD and CsCMV, to cause disease, as they are often detected alongside other viral infections. Notably, mixed-virus infections in cassava are more common than previously thought and are associated with various leaf symptoms in indicator plants[9].
The growing economic impact of cassava mosaic geminiviruses (CMGs), which cause cassava mosaic disease (CMD) in Africa and Asia, along with the cassava brown streak viruses (CBSVs) responsible for cassava brown streak disease (CBSD) in Africa and CFSD in the Americas[20], renders them the foremost global threats to cassava production[21]. The main viruses compromising cassava production include CMGs and CBSVs[22] in Africa, CsCMV, and Sri Lankan cassava mosaic virus (SLCMV)[23] in Asia; and Cassava vein mosaic virus (CsVMV), CFSD, and CsCMV in the Americas[24]. Recent studies have reported the re-emergence of CsCMV[25], and the potential threat requires further investigation. First identified in cassava in Brazil by Silberschmid (1938)[26], CsCMV can cause yield losses of 30% to 60%[7]. For instance, infection with the Argentinian CsCMV strain negatively affects all growth and yield components, severely reducing tuber size[27]. CsCMV symptoms disrupt photosynthesis and plant development, leading to significant losses in root yield and the quality of propagation material[28]. These findings underscore the impact of CsCMV on cassava crops and the urgent need for its eradication.
Understanding the biology of CsCMV and developing effective management strategies are essential for ensuring sustainable cassava cultivation. This review explores current knowledge on CsCMV and its interaction mechanisms with cassava. It also highlights potential directions for future research and management strategies to address the challenges posed by CsCMV in cassava production. Furthermore, we investigated the evolutionary relationships of reported CsCMV viral proteins and analyzed the geographical distribution of CsCMV.
-
The ICTV classifies potexviruses into species on the basis of host range, the absence of cross-protection in infected plants, serological differences, and sequence identity thresholds of 72% for nucleotides and 80% for amino acids (
https://ictv.global/report/chapter/alphaflexiviridae/alphaflexiviridae/potexvirus ). CsCMV is a potexvirus of the family Alphaflexiviridae (Supplementary Table S1) within the order of Tymovirales[29]. The potexviruses contain a single-stranded (ss) positive-sense (+) RNA genome of 5.5–9 kb with flexuous filamentous virions that are 12–13 nm in diameter and 470–800 nm in length (Fig. 1)[29]. The Potexvirus genome has a 5'-methyl guanosine cap and a 3'-polyadenylated tail flanked by five open reading frames (ORFs)[29]. ORF1 encodes the viral replicase, which includes methyltransferase, helicase, and RNA-directed RNA polymerase domains. Following ORF1 are the triple-gene block (TGB) proteins, a feature common to all plant-infecting members of this family[29]. The TGB proteins are movement proteins (MPs) essential for viruses' cell-to-cell and long-distance movement and are needed to invade the host plant and cause disease successfully[30]. Viruses of the TGB superfamily are categorized into two groups depending on the structural variations of TGB1 and TGB3, as well as the necessity of the coat protein (CP) for their movement[31,32]. The Potexvirus TGB1 protein contains an NTPase/helicase domain involved in hydrolysis of adenosine triphosphate (ATP), RNA binding, homologous interactions, and cell-to-cell movement[16,31,33]. In addition, the TGB1 protein contributes to the variability of RNA silencing suppression in Potexvirus[34]. For instance, the TGB1 protein encoded by Plantago asiatica mosaic virus (PlAMV), which has strong suppressor activity, affected the accumulation of microRNA (miRNA), virus genomic RNA, and virus-derived small interfering RNAs[35]. The TGB2 and TGB3 proteins of some potexviruses have been linked to the formation of perinuclear granular structures and peripheral bodies[36,37]. Several mechanisms for the movement of potexviruses have been proposed, with a particular focus on the roles of TGB2 and TGB3[32]. In a refined model, potexviruses' cell-to-cell movement depends on the virion cargo associating with a TGB2–TGB3 membrane complex, which then recruits TGB1 to the plasmodesmata[38]. TGB2, which contains two transmembrane domains and a hydrophilic core region, is associated with motile granular vesicles derived from the endoplasmic reticulum (ER). These vesicles facilitate viral cell-to-cell transmission and systemic infection[36]. The transport of Potexvirus RNA typically depends on the TGB2–TGB3 complex, which may directly bind viral RNA or interact with other viral components, such as TGB1 or the CP, which possess RNA-binding capabilities, to form virions. These virions are directed to the plasmodesmata via trafficking signals from TGB3 and are transported across the plasmodesmata with the assistance of TGB2-interacting proteins[39]. The CP, which is required for virion assembly and cell-to-cell movement, is encoded by ORF5[40]. In this review, we retrieved CsCMV genome sequences from the National Center for Biotechnology Information (NCBI) database to investigate the evolution of CsCMV viral proteins (Supplementary Table S2). Only complete genome sequences were analyzed. Phylogenetic relationships among CsCMV's viral proteins were subsequently inferred using the MEGA12 program. The analysis revealed that RdRP, CP, and TGB-1, -2, and -3 cluster within a single clade (Group I), distinct from the other potexviruses (Group II), indicating that CsCMV's viral proteins likely evolved from a common ancestor (Supplementary Figs S1−S5). Interestingly, CsCMV TGB2 proteins from Group I are more closely related to the TGB2 proteins of allexiviruses than those of other potexviruses (Group II) (Supplementary Fig. S4), suggesting that they may have originated from a different evolutionary lineage. Notably, all reported CsCMV sequences include the TGB3 protein, highlighting its conserved presence across the Potexvirus population (Supplementary Fig. S5).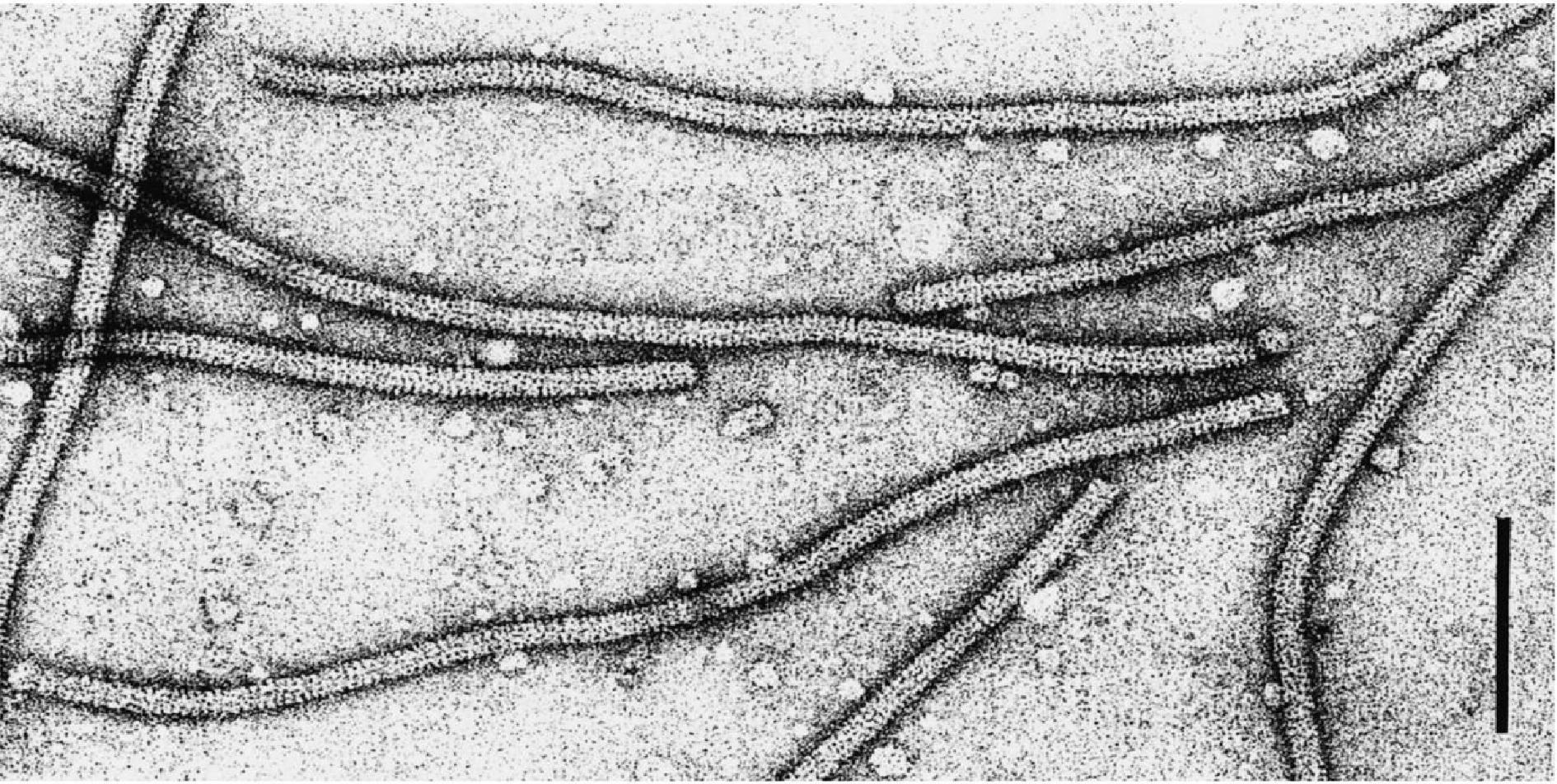
Figure 1.
Potexvirus viral particles. Negative-contrast electron micrograph of particles of an isolate of potato virus X. The bar represents 100 nm (Courtesy of D.-E. Lesemann). The picture was downloaded from the International Committee on Taxonomy of Viruses (ICTV),
https://ictv.global/report/chapter/alphaflexiviridae/alphaflexiviridae/potexvirus . -
Reports of CsCMV infections have primarily been limited to Asia and Latin America, particularly in countries such as Brazil[7,24,26], Argentina[41,42], Venezuela[43], Colombia[7], Paraguay[7,19], Peru[44], and China[25] (Fig. 2). CsCMV has also been identified in chaya (Cnidoscolus chayamansa or Cnidoscolus aconitifolius) in Mexico[45], Venezuela[46], Tuvalu[47], Costa Rica[48], Florida[49], and Colombia[7] (Fig. 2). However, CsCMV's presence in Costa Rica has yet to be confirmed in situ[48]. One African report suggests that CsCMV was introduced to Côte d'Ivoire from Central or South America[50] (Fig. 2). These findings indicate that the virus may be more diverse and widespread in the South American region. Among the affected countries, southern Brazil and Paraguay have reported CsCMV as the most prevalent and significant[51]. Some studies have suggested CsCMV's presence in other hosts, such as Pistacia vera (GenBank: MT334616.1); however, no direct evidence has confirmed its replication in P. vera. CsCMV has a broad host range, infecting species from various dicotyledonous families, including Amaranthaceae, Chenopodiaceae, Compositae, Euphorbiaceae, Malvaceae, and Solanaceae[49,52]. CsCMV causes systemic mosaic symptoms in manually inoculated plants such as chaya, Ricinus communis, cassava, Jatropha spp., Euphorbia spp., Nicotiana benthamiana, and N. occidentalis[41,49], while inducing local lesions in Chenopodium amaranticolor, Cassia occidentalis, Datura stramonium, Chenopodium quinoa, and Gomphrena globose[49]. Notably, CsCMV strains may differ by region. Sequence analysis of the conserved RdRp domain has revealed significant diversity among isolates. Interestingly, isolates from a specific region formed a distinct phylogenetic cluster, sharing only 77.1%–80.3% nucleotide identity with other clusters[42]. However, these genetic differences do not hinder the detection of CsCMV using techniques such as ELISA[42]. CsCMV is primarily transmitted through mechanical means, involving direct contact with infected plant sap or contaminated agricultural tools, and via vegetative propagation when infected stem cuttings are used for planting[53]. Experimentally, the virus can also be transmitted through agroinoculation (Fig. 3). To date, there is no evidence suggesting transmission via seeds or insect vectors[54]. A study speculated that CsCMV, which was primarily found in Brazil exclusively infecting cassava, could spread to other regions, particularly where Brazilian cultivars have been introduced[55]. Cassava colombian symptomless virus (CsCSV) and cassava virus X (CsVX), along with CsCMV, are recognized members of the Potexvirus genus, according to the ICTV. These viruses can be mechanically transmitted and generally cause symptomless infections, except for CsCMV, which is symptomatic[9,42]. Notably, CsCMV strains display considerable diversity, exhibiting varying serological and biological characteristics, highlighting the growing complexity of potexviruses in cassava[9,42]. Environmental factors, such as prolonged cold, intensify its distribution, with southern Brazil and Paraguay showing the highest prevalence[51].
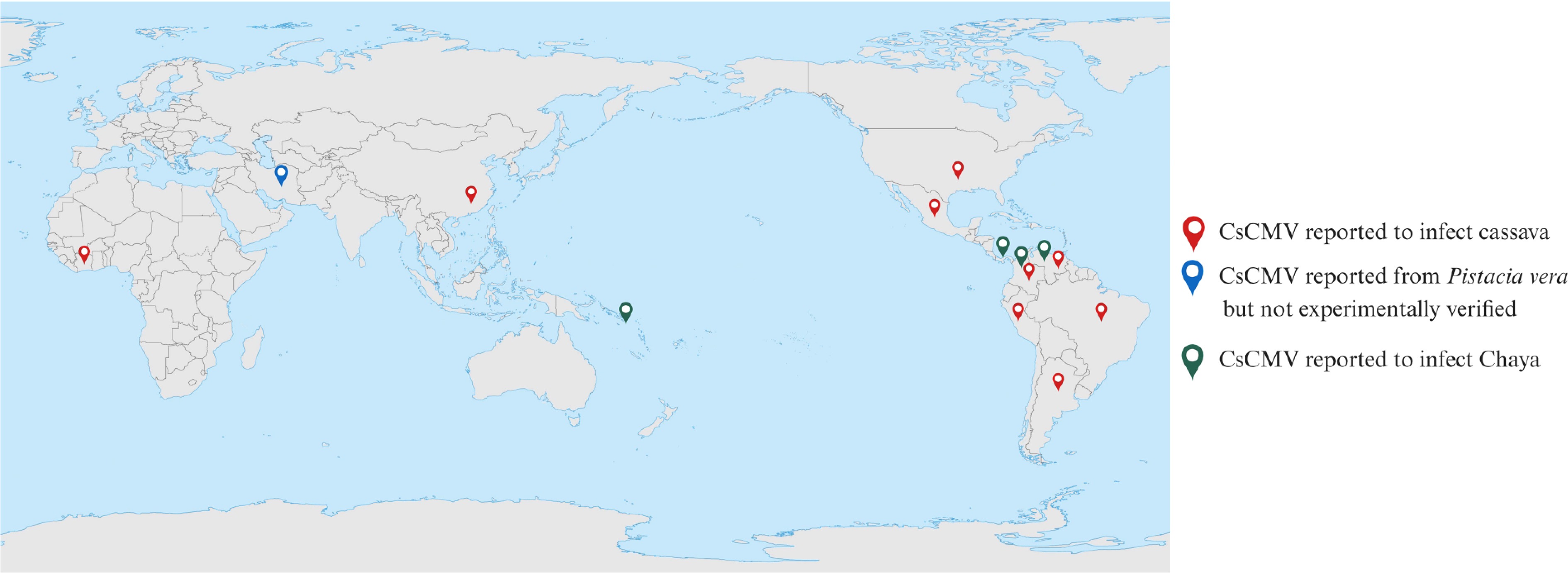
Figure 2.
Geographical distribution of cassava common mosaic virus (CsCMV). CsCMV viral genomes were reported mainly from Latin America and Asia in countries such as Brazil, Argentina, Venezuela, Colombia, Paraguay, Peru, and China. In addition, CsCMV was reported in Côte d'Ivoire and Tuvalu. CsCMV has also been identified in chaya (Cnidoscolus chayamansa or Cnidoscolus aconitifolius) in Mexico, Venezuela, Tuvalu, Florida, Costa Rica, and Colombia.
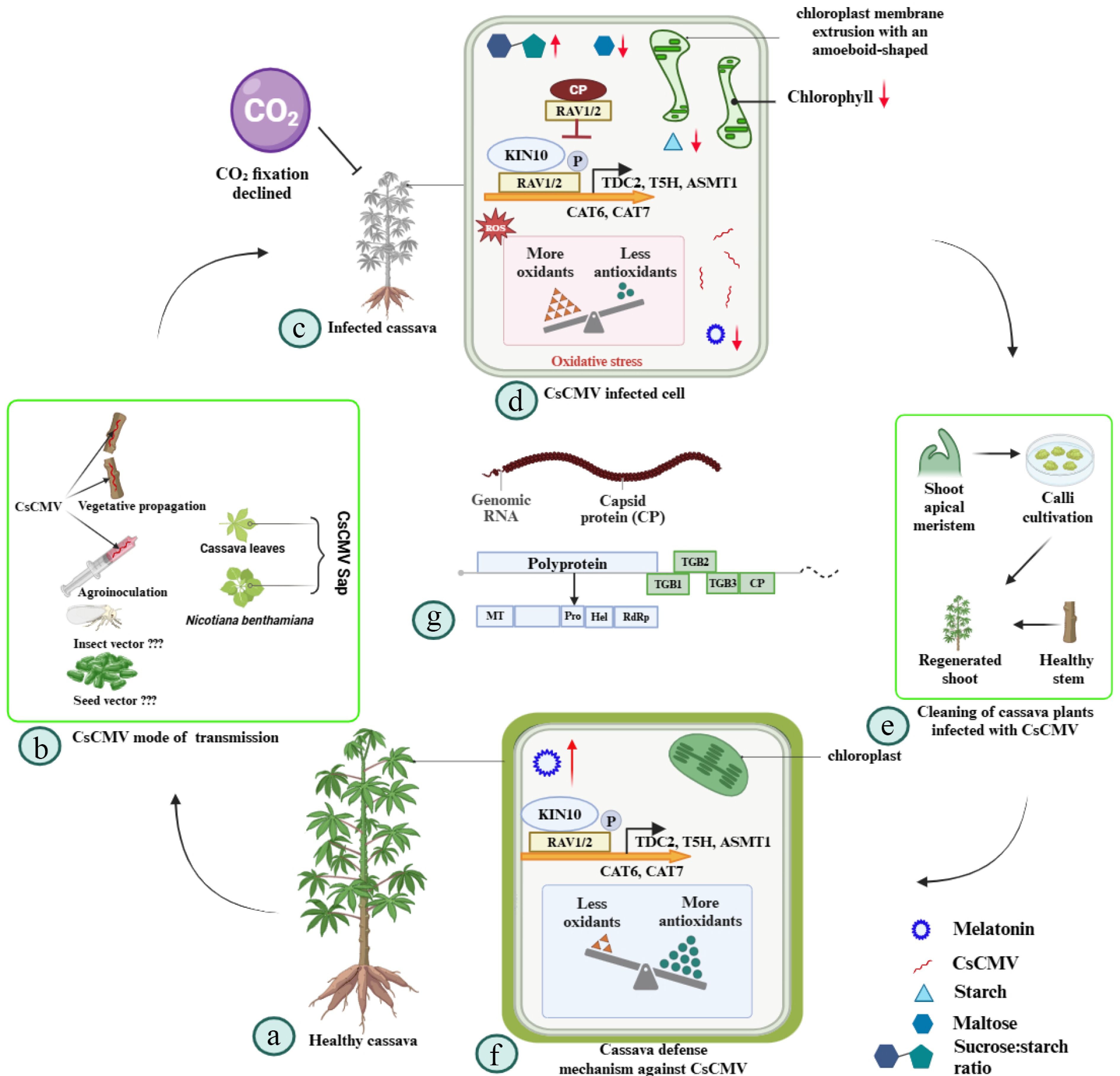
Figure 3.
Advances in research into cassava common mosaic virus (CsCMV). (a) Healthy cassava. (b) Mode of transmission. (c) Infected cassava. (d) Infected cell. (e) Cleaning of cassava-infected plants. (f) Defense mechanism. (g) Genome organization of Potexvirus. RNA-dependent RNA polymerase (RdRp); triple gene block (TGB); coat protein (CP); helicase (Hel); methyltransferase (MT). CsCMV can be transmitted through agroinoculation, vegetative propagation, and sapping, although no seed or insect transmission vectors have been identified. Cassava infected with CsCMV exhibits various physiological effects, including chloroplast membrane extrusion with ameboid-like shapes, oxidative stress, reduced CO2 fixation, and diminished chlorophyll levels, negatively impacting photosynthesis. Additionally, infections lead to decreased maltose and starch content in the plant. Melatonin positively regulates cassava's resistance to CsCMV. CsCMV infections can be addressed by isolating and regenerating the shoots' apical meristem. Furthermore, only healthy plants or stems should be used for vegetative propagation. The cutting tools should be regularly disinfected to minimize the risk of transmission.
-
CsCMV infection causes mosaic leaf patterns, stunted growth, and smaller tubers. A previous study from Florida reported that, in addition to cassava and chaya, other economically significant plants are susceptible to the issue, including Ricinus communis, certain ornamental varieties of Euphorbia, Jatropha podagrica, and Jatropha gossypiifolia[49]. Multiple studies have reported significant yield losses in cassava resulting from CsCMV. For instance, crop production losses in Brazil have been observed to range from 30% to 60%, with the most severe damage occurring in regions experiencing protracted cold periods rather than tropical regions[28]. In Argentina, the incidence of CsCMV reached up to 85.2%[42], while Brazil recorded an incidence of up to 100%[24,56]. Similarly, Colombia reported an incidence as high as 90%[57]. One study evaluated the impact of CsCMV infection on various plant traits, including plant height (PH), shoot fresh weight (SFW), total fresh weight (TFW), tuberous root length (TRL), number of tuberous roots (TRN), fresh weight of individual tuberous roots (FWiTR), tuberous root diameter (TRD), total fresh weight of tuberous roots per plant (FWTR), harvest index (HI), and estimated tuberous root yield (EY). Infected plants exhibited significant yield losses, with EY and FWTR being the most affected traits, both showing a 44.3% reduction. Other severely impacted traits included TFW (37.8% loss), SFW (33.5% loss), TRN (29.9% loss), and FWiTR (24.8% loss)[27]. In addition, CsCMV infection significantly impacts chloroplast function and plant metabolism[58]. It disrupts sugar metabolism and allocation, alters the chloroplast structure, and impairs chloroplast functionality in the source leaves, ultimately reducing cassava tuber yields[59]. For instance, chloroplasts in CsCMV-infected mesophyll cells display membrane extrusion, an ameboid morphology, and disorganized grana stacks, reducing chlorophyll content and impairing CO2 fixation[59] (Fig. 3). In addition, CsCMV-infected plants experience oxidative stress, diminished antioxidant capacity, reduced starch and maltose levels, and an increased sucrose-to-starch ratio, which are all indicative of altered carbon allocation (Fig. 3). These findings underscore the extensive physiological and metabolic disruptions caused by CsCMV, which have profound implications for cassava production and disease management[59]. An optimized quantitative polymerase chain reaction (qPCR) diagnostic method has been established for detecting CsCMV, offering greater sensitivity and accuracy than serological techniques and molecular techniques like double antibody sandwich (DAS)-ELISA and end-point PCR[60]. This enhanced qPCR protocol enables efficient screening of cassava germplasm, ensuring the safe distribution of disease-free planting materials[60].
-
PVX demonstrated that in the absence of TGB3, cell-to-cell and systemic virus movements were inefficient or restricted to the initially infected leaves, where the local symptoms were also less severe[40]. However, some members of the Alphaflexiviridae family (Supplementary Table S1), such as allexivirus, encode a TGB3-like sequence that lacks an AUG start codon[61]. A previous study using bioassays and sequence analysis reported that cassava Colombian symptomless virus (CsCSV) and cassava virus X (CsVX) isolates cause latent infections in various potato landraces[15]. CsCSV and CsVX, which lack the conserved potexviral TGB3, were not efficiently transmitted to N. benthamiana[15]. In contrast, all CsCMV isolates in single infections were efficiently transmitted to N. benthamiana[15]. These findings suggest that the TGB3 protein may play a role in infection and transmissibility. Notably, the absence of TGB3 in CsNAV and CsVX appears to have a more significant impact on cell-to-cell movement than systemic virus movement[15]. It has been hypothesized that TGB3 was lost in CsNAV and CsVX, due to their persistent infection of host plants, as a complete TGB is conserved at the Alpha- and Betaflexiviridae family levels (ICTV, ninth report)[62]. This suggests that TGB3 may not be essential for long-distance transmission in potatoes, as co-infecting viruses could compensate for this function. This hypothesis is supported by in silico reconstructions of viral genomes, which indicate that TGB3 acquisition occurs only after establishing the TGB1–TGB2 block. Consequently, this predicts the existence of infectious viruses that lack TGB3[63]. A recent study found that the CP targets the RAV1 and MeRAV2 TFs in M. esculenta (Me), positively regulating disease resistance against CsCMV (Fig. 3). This regulation occurs through the transcriptional activation of melatonin biosynthetic genes, including tryptophan decarboxylase 2 (MeTDC2), N-aceylserotonin O-methyltransferase 1 (MeASMT1), tryptamine 5-hydroxylase (MeT5H), and MeCatalase6 (MeCAT6) and MeCAT7[64]. The interaction among CP, MeRAV1, and MeRAV2 disrupted the phosphorylation of MeRAV1 and MeRAV2 at Ser45 and Ser44 by the protein kinase 10 (MeKIN10). This interference reduced the transcriptional activation activity of MeRAV1 and MeRAV2 on the melatonin biosynthesis genes MeCAT6 and MeCAT7, which is dependent on the phosphorylation of MeRAV1 and MeRAV2[64]. Additionally, a defective RNA associated with a Potexvirus, containing the 5' and 3' untranslated regions (UTRs) and an ORF, may encode the first 263 amino acids of RdRp and the last 33 amino acids of the CP. However, the role of these defective RNAs in pathogenicity remains unclear[7].
-
Apparent symptoms of CsCMV typically appear on newly developed leaves from infected stem cuttings. Infected plants should be removed at the first stage of infection. If not, they should be destroyed after harvesting the tuberous root[51]. To prevent the transmission of CsCMV, only healthy plants or stems should be used for vegetative propagation. Additionally, cutting tools should be regularly disinfected to minimize the risk of spreading the virus[51]. Careful selection of plant material plays a crucial role in mitigating the economic impact of CsCMV.
-
Recent advances have facilitated the development of virus-mediated protein overexpression (VOX) and virus-induced gene silencing (VIGS) vectors derived from CsCMV. These tools are rapid, cost-effective, and valuable for functional genomic studies, biotechnology applications, and plant genome editing[65,66]. However, a virus-induced genome editing (VIGE) vector has not been reported in cassava. Nevertheless, a VIGE vector was developed in monocots (maize) using foxtail mosaic virus (Potexvirus) vectors[67]. These innovations could extend to genome editing, enhancing the development of resistance.
-
A previous study speculated that cassava-infecting potexviruses may be more diverse[15]. The unavailability of samples from original isolates and the lack of sequence data obstruct the classification of potexviruses, compromising disease diagnostics, evaluation of virus cleaning protocols, cassava germplasm exchange certification, and epidemiological studies[15]. Identifying further incidences of CsCMV across different geographic locations could yield valuable insights regarding the virus's reproduction mechanisms and overall effects. This information would be crucial in developing effective strategies to manage and control CsCMV. A study conducted a metagenomic analysis of a mangrove crab (Aratus pisonii) and discovered the CsCMV genome, most likely due to the cassava plant identified in the data; however, it was recommended that additional research be conducted to determine whether the mangrove crab is an actual sink for CsCMV[68]. The development of a VIGE vector derived from CsCMV is intriguing, since it may facilitate the editing of the cassava genome, which is now challenging due to complex and restricted tools. Breeding for resistance is limited, but biotechnological methods, such as RNA interference (RNAi) and clustered regularly interspaced short palindromic repeat (CRISPR)-based editing, offer the potential for developing resistant varieties. Integrated pest management enhances the efficacy of control. Substantial progress has been made in understanding the effects of CsCMV on chloroplast function. However, the mechanism by which CsCMV induces extrusion of the chloroplast membrane, thereby impairing photosynthesis, remains unexplored. The TGB1 protein was associated with suppressor activity[34,35]; however, its mechanism of action remains largely unexplored. Moreover, the specific roles of TGB2 and TGB3 in CsCMV's pathogenesis remain to be elucidated. Understanding the biological roles of viral proteins and their interaction complexes is critical for identifying their targets, which could help develop effective strategies to mitigate the effects of viral diseases and inspire the creation of novel biotechnological tools. While previous research has highlighted the unique roles of TGB proteins and the CP in the replication and movement of various potexviruses, these functions have rarely been studied in the context of CsCMV infecting cassava. Notably, some potexviruses lack a TGB3 protein and are asymptomatic, making it particularly compelling to investigate the role of TGB3 in CsCMV through approaches such as gene silencing or targeted mutation. Furthermore, identifying and characterizing the proteins involved in the CsCMV-cassava interaction could unravel the molecular mechanisms and key factors driving this interaction. While no vector has been reported as being capable of transmitting CsCMV to noninfected cassava plants, investigating the mechanisms underlying CsCMV's transmission would be of significant interest. Although antioxidants and reactive oxygen species (ROS) have been reported to play essential roles in CsCMV infection, the specific mechanisms and the antioxidants involved remain largely unknown. Although CsCMV was first isolated from cassava in Peru, its evolutionary history remains unclear. Further research is needed to trace the virus's origins and, most importantly, to understand how it spreads across continents.
-
CsCMV significantly impacts cassava production, particularly in Latin America and Asia. Advances in diagnostics, molecular insights, and control strategies are promising, yet challenges like developing resistant varieties and epidemiological understanding persist. Ongoing research and global cooperation are essential to mitigate its effects.
This work was supported by the Natural Science Foundation of China (32260468), the China Agriculture Research System (CARS-11-hncyh), and the International Science & Technology Cooperation Program of Hainan Province (GHYF2024008).
-
The authors confirm their contributions to the paper as follows: conceptualization: Chen Y, Assane Hamidou A; funding acquisition: Chen Yinhua, supervision: Chen Yinhua, Assane Hamidou A, Chen F; validation: Chen Yinhua, Assane Hamidou A, Chen F, Chen Yuhua, Wang H, Abdou Mahaman M; writing–original draft preparation: Assane Hamidou A; writing–review and editing: Chen Yinhua, Assane Hamidou A, Chen F, Chen Yuhua, Wang H, Abdou Mahaman M. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0025)
-
Received 21 January 2025; Accepted 7 July 2025; Published online 28 July 2025
- Supplementary Table S1 Proposed taxonomy of plant viruses (Alphaflexiviridae). International Committee on Taxonomy of Viruses (2024, MSL #40).
- Supplementary Table S2 Cassava common mosaic virus (CsCMV) complete genome sequences used to infer phylogenetics. The complete genome sequences were retrieved from the National Center for Biotechnology Information (NCBI). M. esculenta: Manihot esculenta, N. Benthamiana: Nicotiana benthamiana.
- Supplementary Fig. S1 Phylogenetic analysis of Cassava Common Mosaic Virus (CsCMV) RNA-dependent RNA polymerase (RdRp). MEGA12 was utilized to infer the phylogenetic tree by constructing maximum likelihood phylogenetic trees, using best-fit substitution models LG + G + I, with 1,000 bootstrap replicates. Virus sequences were downloaded from the National Center for Biotechnology Information (NCBI) database. Allexiviruses were used as an outgroup. Viruses names highlighted in red are officially recognized by International Committee on Taxonomy of Viruses (ICTV, 2023).
- Supplementary Fig. S2 Phylogenetic analysis of Cassava Common Mosaic Virus (CsCMV) Coat protein (CP). MEGA12 was utilized to infer the phylogenetic tree by constructing maximum likelihood phylogenetic trees, using best-fit substitution models LG + G + I, with 1,000 bootstrap replicates. Virus sequences were downloaded from the National Center for Biotechnology Information (NCBI) database. Allexiviruses were used as an outgroup.
- Supplementary Fig. S3 Phylogenetic analysis of Cassava Common Mosaic Virus (CsCMV) Triple gene block (TGB)1. MEGA12 was utilized to infer the phylogenetic tree by constructing maximum likelihood phylogenetic trees, using best-fit substitution models LG + G + I, with 1,000 bootstrap replicates. Virus sequences were downloaded from the National Center for Biotechnology Information (NCBI) database. Allexiviruses were used as an outgroup.
- Supplementary Fig. S4 Phylogenetic analysis of Cassava Common Mosaic Virus (CsCMV) Triple gene block (TGB)2. MEGA12 was utilized to infer the phylogenetic tree by constructing maximum likelihood phylogenetic trees, using best-fit substitution models LG + G + I, with 1,000 bootstrap replicates. Virus sequences were downloaded from the National Center for Biotechnology Information (NCBI) database. Allexiviruses were used as an outgroup.
- Supplementary Fig. S5 Phylogenetic analysis of Cassava Common Mosaic Virus (CsCMV) Triple gene block (TGB)3. MEGA12 was utilized to infer the phylogenetic tree by constructing maximum likelihood phylogenetic trees, using best-fit substitution models LG + G + I, with 1,000 bootstrap replicates. Virus sequences were downloaded from the National Center for Biotechnology Information (NCBI) database. Allexiviruses were used as an outgroup.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Assane Hamidou A, Chen Y, Wang H, Abdou Mahaman M, Chen F, et al. 2025. Cassava common mosaic virus (CsCMV): recent advances and future directions. Tropical Plants 4: e027 doi: 10.48130/tp-0025-0025 

Cassava common mosaic virus (CsCMV): recent advances and future directions
- Received: 21 January 2025
- Revised: 24 June 2025
- Accepted: 07 July 2025
- Published online: 28 July 2025
Abstract: Manihot esculenta Crantz (cassava) is a staple food crop in tropical and subtropical regions, supporting over 800 million people globally. However, Cassava common mosaic virus (CsCMV), a potexvirus in the family Alphaflexiviridae, threatens its production, particularly in the Americas and Asia. This review summarizes recent advances in CsCMV research, including its taxonomic classification, epidemiology, diagnostic methods, and interactions with cassava. Key findings reveal that CsCMV possesses a single-stranded RNA genome primarily transmitted mechanically, results in substantial yield reductions in cassava crops and can be detected by novel molecular diagnostic tools. Current challenges in managing CsCMV include limited numbers of resistant cassava varieties and diagnostic accessibility. Future research should prioritise genome editing to develop resistant cassava varieties, investigate the roles of viral proteins in pathogenesis, and promote global collaboration to enhance CsCMV management strategies.