









-
In natural environments, plants are confronted with various stresses such as drought, low temperature, and heat, which disrupt protein homeostasis and significantly impact their growth and development[1]. Recently, the problem of global warming has intensified, and high temperatures have become one of the most serious abiotic stresses affecting plants. Through evolution, plants have established robust self-defense mechanisms to maintain protein functions under stressful conditions[2]. Heat shock proteins (Hsps), universally existing in both prokaryotic and eukaryotic cells, are ubiquitous stress-induced proteins[3,4]. Plants that express Hsps are better able to cope with a variety of unfavorable conditions by preventing protein denaturation[4,5]. Under adversity, heat shock factor (Hsf) recruits the transcriptional machinery to the upstream promoter region of the Hsp by binding to the specific heat shock element (HSE: 5'nGAAnnTTCnnGAAn-3')[6].
On the basis of molecular weight and amino acid sequence homology, Hsps comprise five major families, including Hsp100, Hsp90, Hsp70/Dank, Hsp60/GroE, and small Hsp (sHsp), which are also referred to as Hsp20[7]. Plants possess numerous Hsp20s, with some plants even harbor over 40 Hsp20s[7]. Although the structures of different Hsp20s are distinctive, the α-crystalline C-terminal domain (ACD) is a ubiquitous and highly preserved domain common to Hsp20s[8,9]. Comprising 80–100 conserved amino acids, the ACD domain forms a β-strand structure and is defined by two conserved regions (CRs): CRI (β2-β5) and CRII (β7-β9, and the β6 loop)[10]. Plant Hsp20 proteins are categorized into discrete subfamilies based on key attributes such as sequence homology and function, subcellular localization, and immunological cross-reactivity[11]. Seven subfamilies (CI-CVII) of Hsp20 are localized in the cytosol or nucleus, while the CI gene family constitutes the largest group in plants. Additional Hsp20 subfamilies localize to multiple organelles, including chloroplast (CP), plastid (P), endoplasmic reticulum (ER), mitochondria (M), and peroxisomes (Po)[9,12]. The Hsps thereby function to safeguard cellular proteostasis, mitigating the damage inflicted by environmental stresses[13]. After plants are exposed to heat stress (HS), Hsp20 ensures the normal function of other proteins by safeguarding against heat-denatured aggregation and irreversible denaturation, providing a molecular basis for enhancing the thermotolerance of plant organs[14]. Overexpression of ZmHSP16.9 in tobacco resulted in increased heat tolerance[15]. Heterologous overexpression of TaHsp26 strengthened the thermotolerance of transgenic Arabidopsis[16].
Recognizing the critical role of Hsp20 genes in mediating abiotic stress resistance has prompted genome-wide identification in multiple species, including Arabidopsis, rice, and grape, while the biological functions of crucial Hsp20s have been reported[14,17,18]. The Euphorbiaceae family is both large and geographically widespread, predominantly in tropical and subtropical regions, encompassing life forms such as trees, perennial shrubs, and herbs[19]. Consequently, it is more appropriate to select important species of Euphorbiaceae as objects to analyze the function of Hsp20 in helping plants adapt to environmental changes. Most plants in the Euphorbiaceae originated in the tropics. By contrast, Euphorbia peplus, which belongs to the Euphorbioideae subfamily, originates from the Mediterranean coast in the subtropics, and is now mostly distributed in the subtropics. Ricinus communis (castor bean) and Mercurialis annua, belonging to the Acalyphoideae subfamily, stemmed from the tropics and subtropics and are widely distributed in both tropical and subtropical regions, with castor bean even distributed in temperate regions. Manihot esculenta (cassava), Hevea brasiliensis (rubber tree), Jatropha curcas (physic nut), and Vernicia fordii (tung tree), which belong to the Crotonaceae subfamily, all derive from the tropics. Tung trees, which are also found in small numbers in temperate climates, are the only ones that are widely dispersed in tropical and subtropical regions. Driven by the evolutionary trajectory and trait diversity, Euphorbiaceae species are able to adjust to changing environmental conditions, especially temperature and climate.
The species of Euphorbiaceae are also well known for their wide applications in the industrial and pharmaceutical areas. Cassava can be used to produce environmentally friendly hot-melt adhesives for textile industries and composite films; also, the wastewater generated from its processing industry can be utilized as an economical culture medium for biosurfactant production[20−22]. The rubber tree efficiently dominates the production of natural rubber latex, which can be used for the production of biomass energy, as well as in the construction and furniture industries[23,24]. Moreover, over 60% of natural rubber is utilized in the tire industry[25]. The oil extracted from the seeds of the physic nut can be used to produce biodiesel, aviation fuel, and is also applicable in the pharmaceutical industry[26,27]. The tung tree is an industrial oil tree that can be used to produce biocomposites, rigid polyurethane foam, and tung oil-based oligomer[28−30]. Castor bean is an important industrial crop worldwide, whose oil is extracted from seeds not only be used to produce traditional industrial products, but also to manufacture excellent nanocomposites, new polyurethane adhesives, and waterborne polyurethane composites[31−34]. Furthermore, M. annua is prized for the antimicrobial activity of its extract, and E. peplus has potential as a therapeutic agent for skin cancer[35,36]. Nevertheless, the Hsp20 genes of Euphorbiaceae have not yet been identified at the genome-wide level.
Consequently, this study identified the Hsp20 gene family in seven Euphorbiaceae species to conduct a comparative analysis of their evolution. The characteristics and functions, evolutionary relationships, and expression patterns of Hsp20s were analyzed. 252 Hsp20 genes were identified, among which four genes (MeHsp20-17, EpHsp20-7, MaHsp20-14, and HbHsp20-30) may respond to temperature stress and stem from the same ancestor. The comprehensive identification of Hsp20 genes in Euphorbiaceae species advanced a key hypothesis that Hsp20s mediate environmental adaptation. This premise establishes a conceptual framework for studying the mechanistic role of the Hsp20 gene in influencing the distribution of Euphorbiaceae plants.
-
The protein and CDS sequences for the seven species were sourced from NCBI, Phytozome, and GSA, respectively. The genome version and other information were provided in Supplementary Table S1. The Hsp20s in seven representative Euphorbiaceae species were screened through PF00011 of the Hidden Markov Model profile (HMMER) and BLASTp with an E-value threshold of 1.0 × 10−5. Low-molecular-weight proteins containing an ACD domain, which is composed of approximately 80–100 conserved amino acids, were defined as candidate proteins. Prediction of the conserved domain within the candidate protein sequences was performed via the SMART (
http://smart.embl-Heidelberg.de ) online tool and CDD (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ) database, facilitating the removal of redundant proteins. Finally, the Hsp20 family members with highly conserved feature information were screened, which were named Hsp20s.The key physicochemical parameters, encompassing protein length, molecular weight (MW), isoelectric point (PI), Grand Average of Hydropathicity (GRAVY), and prediction of subcellular localization for the identified Hsp20 proteins were determined via the ExPASy Proteomics Server (
https://web.expasy.org/protparam ) and WoLF PSORT (https://wolfpsort.hgc.jp ), respectively. The Hsp20 protein sequences were aligned with the aid of DNAMAN software.Chromosomal mapping, conserved motifs, and cis-element analysis
-
The chromosome location information of Hsp20 genes was extracted based on genome annotation information, and the chromosome distribution map was drawn using TBtools[37]. To identify conserved motifs, the MEME (
https://meme-suite.org/meme/ ) online tool was employed, with a maximum number of motifs set at 10. TBtools assisted in the visual analysis[37].The 2 kb promoter sequences of Hsp20 from seven Euphorbiaceae species were interrogated for the possible regulatory elements. Location-based matrix, consensus, and specific promoter sequences on a single site of various regulatory elements were predicted through the online website PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ).Phylogenetic, whole-genome duplication, and synteny analysis
-
The Hsp20 protein sequences of M. esculenta, R. communis, J. curcas, M. annua, V. fordii, H. brasiliensis, E. peplus, O. sativa, and A. thaliana were stored in FASTA format in the same file. The ClustalW command was used to perform protein sequence alignment using the MEGA7.1 tool. Guided by the multiple sequence alignment results, an interspecific phylogenetic tree was inferred via the neighbor-joining (NJ) method, employing 1,000 bootstrap replicates for support. The Evolview (
https://evolgenius.info/helpsite/qst1.html ) online website was utilized to edit the evolutionary tree.To compare whole-genome duplication events and identify homologous blocks in different species, whole-genome duplication (WGD) analysis was performed using whole-genome duplication integrated (WGDI) in Python. The results of the comparison were presented in a dot plot. MCScanX inferred collinear regions within the intraspecific and interspecific genomes, and the results were visualized using TBtools.
Homology modeling and conservation position analysis
-
Arabidopsis sHsp (PDB ID: 7BZW) was used as a model[38]. The model was visualized and modified using PyMOL software. The protein sequences of Arabidopsis and 252 Euphorbiaceae Hsp20s were input on the ConSurf server (
https://consurf.tau.ac.il/consurf_index.php ), and the evolutionary conservation and variation scores of each residue were calculated.Prediction of transcription factor binding Hsp20s, protein-protein interaction network, and KEGG pathway
-
To investigate the transcription factors that bind to the Hsp20s of seven Euphorbiaceae, the coding sequences of Hsp20 genes were extracted. The Plant Transcriptional Regulatory Map (PlantRegMap: Plant Regulation Data and Analysis Platform @ CBI, PKU [gao-lab.org]) was leveraged to predict transcription factors that bind to the promoter of Hsp20s.
The STRING online service (
https://cn.string-db.org/ ) with a confidence parameter of 0.15 was used to detect the functional protein-protein interaction network (PPI). Protein association networks for R. communis, M. esculenta, and J. curcas were developed using their species-specific protein sequences, whereas the PPI for the other species were generated based on the homology to Arabidopsis Hsp20 proteins. The KEGG pathway database (www.kegg.jp/kegg/pathway.html ) was employed to predict the functional pathway of Hsp20 proteins.Expression pattern profiling of cassava Hsp20s
-
Gene expression data, encompassing responses to drought and cold treatments, as well as data from different tissues, were sourced from the NCBI database. Gene expression levels were quantified as FPKM (Fragments Per Kilobase of exon model per Million mapped fragments) value, followed by visualization with TBtools[37].
-
Seven typical Euphorbiaceae species were selected for analysis and classified into three different genera (Supplementary Fig. S1). Of these, four species belong to Crotonaceae, two to Acalyphoideae, and one to Euphorbioideae (Supplementary Fig. S1). Using 19 A. thaliana Hsp20s (AtHsp20s) and 39 Oryza sativa Hsp20s (OsHsp20s) as queries, the potential Hsp20 homologs were identified in seven representative Euphorbiaceae species, namely M. esculenta (cassava), R. communis (castor bean), J. curcas (physic nut), H. brasiliensis (rubber tree), V. fordii (tung tree), M. annua, and E. peplus. A total of 252 Hsp20 genes constituted the Hsp20 gene family identified in this study, comprising 23 in R. communis, 24 in J. curcas, 50 in H. brasiliensis, 50 in M. esculenta, 56 in V. fordii, 32 in M. annua, and 17 in E. peplus (Supplementary Fig. S2). The Hsp20 proteins from seven Euphorbiaceae species were named according to species, chromosome number, and gene position on the chromosome. The prefixes 'Me', 'Hb', 'Rc', 'Jc', 'Ep', 'Ma', and 'Vf' were used to denote cassava, rubber, castor bean, physic nut, E. peplus, M. annua, and V. fordii, respectively. As demonstrated in Table 1, although the physic nut, whose genome size was the smallest, contained a greater number of Hsp20s than E. peplus. Interestingly, except for physic nut, species in the Crotonaceae subfamily contained more Hsp20 genes than the other two subfamilies (Supplementary Fig. S1; Table 1).
Table 1. The Hsp20 gene family in Euphorbiaceae.
Family Subfamily Genus Species Gene number Genome size Euphorbiaceae Crotonaceae Jatropha Jatropha curcas 24 266.8 Mb Hevea Hevea brasiliensis 50 1.9 Gb Manihot Manihot esculenta 50 582.28 Mb Vernicia Vernicia fordii 56 1.12 Gb Acalyphoideae Ricinus Ricinus communis 23 315.6 Mb Mercurialis Mercurialis annua 32 453.2 Mb Euphorbioideae Euphorbia Euphorbia peplus 17 267.2 Mb A comparative analysis of physicochemical properties, encompassing protein lengths, protein molecular weight (MW), isoelectric point (pI), and subcellular localization, was undertaken for the Hsp20 proteins from seven Euphorbiaceae species. The molecular weights of Hsp20s ranged from 9,944.53 kDa (MeHsp20-22) to 76,130.34 kDa (HbHsp20-22) within the seven species, while the peptide lengths of Hsp20s varied from 80 (MeHsp20-22) to 692 (HbHsp20-22) amino acids (Supplementary Table S2). The isoelectric points of the Hsp20 protein varied significantly, with the lowest being 4.69 (EpHsp20-14), and the highest being 9.84 (JcHsp20-24). Among them, 158 Hsp20s possessed values lower than 7, indicating that most were acidic. Except for VfHsp20-52, the other Hsp20 proteins showed negative GRAVY values, indicating their hydrophilic nature. The prediction of subcellular localization suggested that Hsp20 proteins of the Euphorbiaceae were widely distributed in the cytoplasm and different organelles, suggesting that they may perform multiple functions (Supplementary Table S2). Only VfHsp20-11 was distributed in the cytoskeleton, suggesting that it may play a special role.
Phylogenetic analysis of Hsp20s
-
To elucidate the evolutionary relationships, an unrooted phylogenetic tree of 252 Euphorbiaceae Hsp20s, 39 rice Hsp20s (OsHsp20s), and 19 A. thaliana Hsp20s (AtHsp20s) was constructed (Fig. 1). All of the Hsp20s in this study were categorized into 13 subfamilies, including two subfamilies in mitochondria (M), seven subfamilies in the cytoplasm or nucleus (CI-CIV), one subfamily in the endoplasmic reticulum (ER), chloroplast (P), and peroxisomal (Po), and 57 Hsp20s in the unclassified group. Most Hsp20s were classified into the CI subfamily, and they were predicted to be located in the cytoplasm or nucleus, including 21 MeHsp20s, 17 HbHsp20s, seven JcHsp20s, 10 MaHsp20s, 28 VfHsp20s, and eight RcHsp20s. Additionally, none of the EpHsp20s were assigned to the CI subfamily, indicating that the Hsp20s of E. peplus, a species that is extensively found in subtropical areas, varied functionally from Hsp20 proteins of other species. Subfamilies ER and M contained all seven species, while subfamily Po contained the Hsp20s from five species except R. communis and E. peplus, and subfamily P contained the Hsp20s except RcHsp20s and JcHsp20s. The results indicated similar evolutionary patterns of Euphorbiaceae and Arabidopsis (Fig. 1). Additionally, the phylogenetic reconstruction identified the ER and Po subfamilies, which diverged later in the evolution of the Hsp20 gene family (Fig. 1), implying that these two subfamilies might have particular functions in the Euphorbiaceae. However, only two Hsp20s (MaHsp20-23 and JcHsp20-8) in Euphorbiaceae were grouped at the same end of the branch as AtHsp20s; while 84 Hsp20s in Euphorbiaceae were classified together, such as MeHsp20-28/HbHsp20-32, RcHsp20-21/HbHsp20-7, and JcHsp20-12/VfHsp20-48 (Fig. 1). These 84 genes were assigned to all subfamilies, excluding CII and MII. In particular, 18 MeHsp20s and HbHsp20s, respectively, clustered at the end of the same branch.
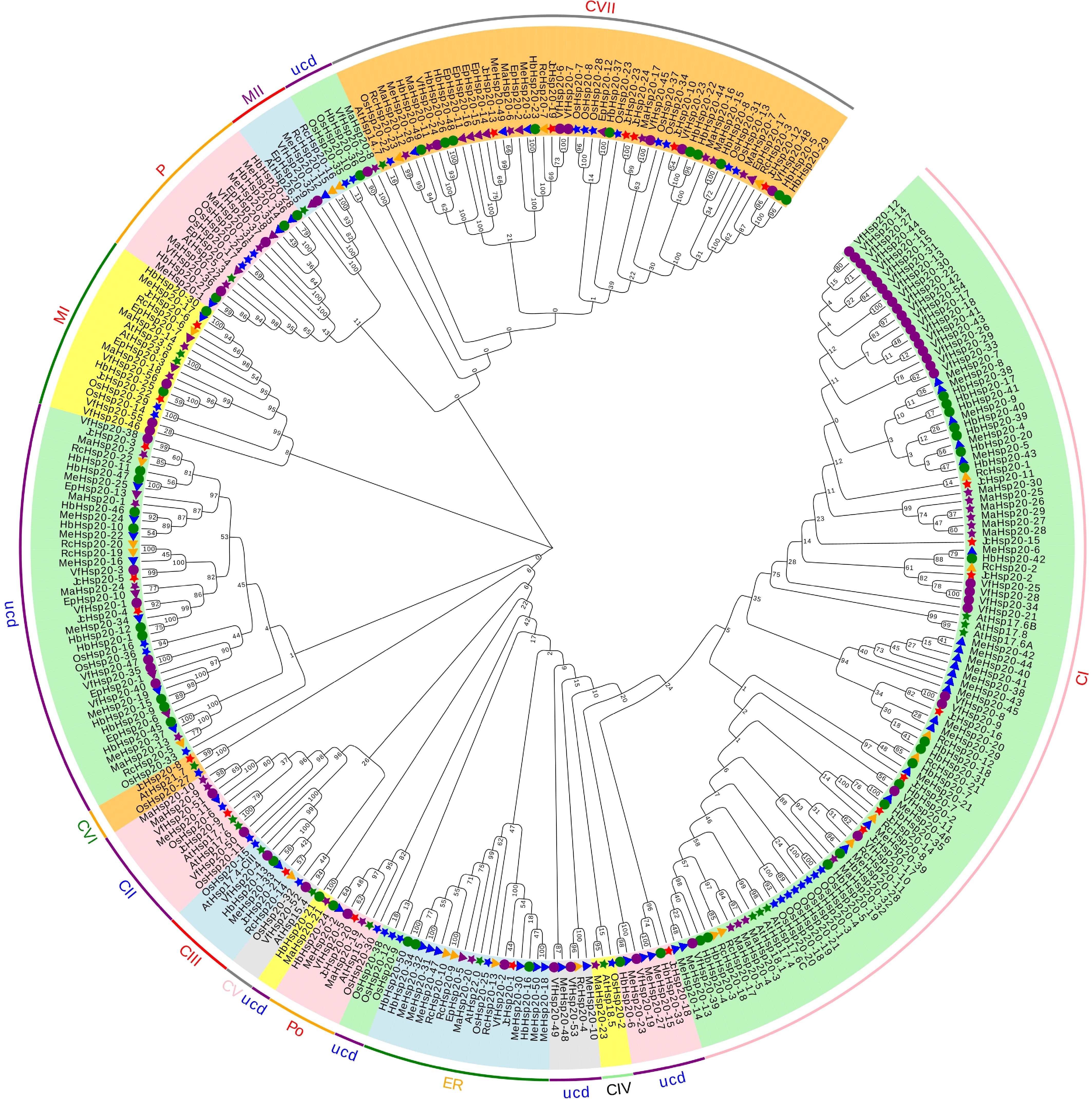
Figure 1.
Unrooted phylogenetic tree of the Hsp20s in seven Euphorbiaceae, rice, and A. thaliana. The Neighbor-joining (NJ) phylogenetic tree was inferred using MEGA 7.1. Different colors represent different clades. Me, M. esculenta; At, A. thaliana; Rc, R. communis; Jc, J. curcas; Hb, H. brasiliensis; Ep, E. peplus; Ma, M. annua; Vf, V. fordii; Os, O. sativa. Green star, blue star, blue triangle, yellow triangle, red star, green circle, purple triangle, purple star, and purple circle indicate A. thaliana, O. sativa, M. esculenta, R. communis, J. curcas, H. brasiliensis, E. peplus; M. annua; and V. fordii, respectively. CI-CIV: cytoplasm or nucleus; ER: endoplasmic reticulum; Po and P: peroxisomal; MI-MII: mitochondria; ucd: unclassified.
Whole-genome duplications analysis
-
Whole-genome duplication (WGD) is a major evolutionary mechanism that leads to the simultaneous duplication of all chromosomal material. Five Euphorbiaceae species with genomes assembled to the chromosomal level were selected, and genome collinear dot plots were drawn with Arabidopsis separately (Supplementary Fig. S3). The density of dots in the dot plots of different Euphorbiaceae species was similar, indicating that the number of highly comparable fragments in the genomes of different Euphorbiaceae species and A. thaliana was similar. In highly similar segments, those containing Hsp20s were labeled. Comparative analysis of syntenic regions revealed a pronounced disparity in Hsp20 tandem array size, which varied from two (castor bean) to nine (rubber tree) across the examined species (Fig. 2). Six HbHsp20s were located in collinear segments and were collinear with seven AtHsp20s. A total of eight AtHsp20s intersected with Hsp20s of five Euphorbiaceae in collinear long segments, among which the genes that intersected most with other Hsp20s was AtHsp23.5 and AtHsp23.6, which were shared by all the other four species except castor bean (Fig. 2). However, AtHsp25.3 only intersected HbHsp20-27, which was divided into the P subfamily, and HbHsp20-27 clustered with MeHsp20-1 at the end of the same branch (Fig. 2d, e). Significantly, MeHsp20-1 was not located in the collinear segments. According to the above results, it was deduced that some Hsp20s in Euphorbiaceae were derived from the original Hsp20s, but more Hsp20s may have been acquired during the evolution of the species itself, and environmental adaptation.
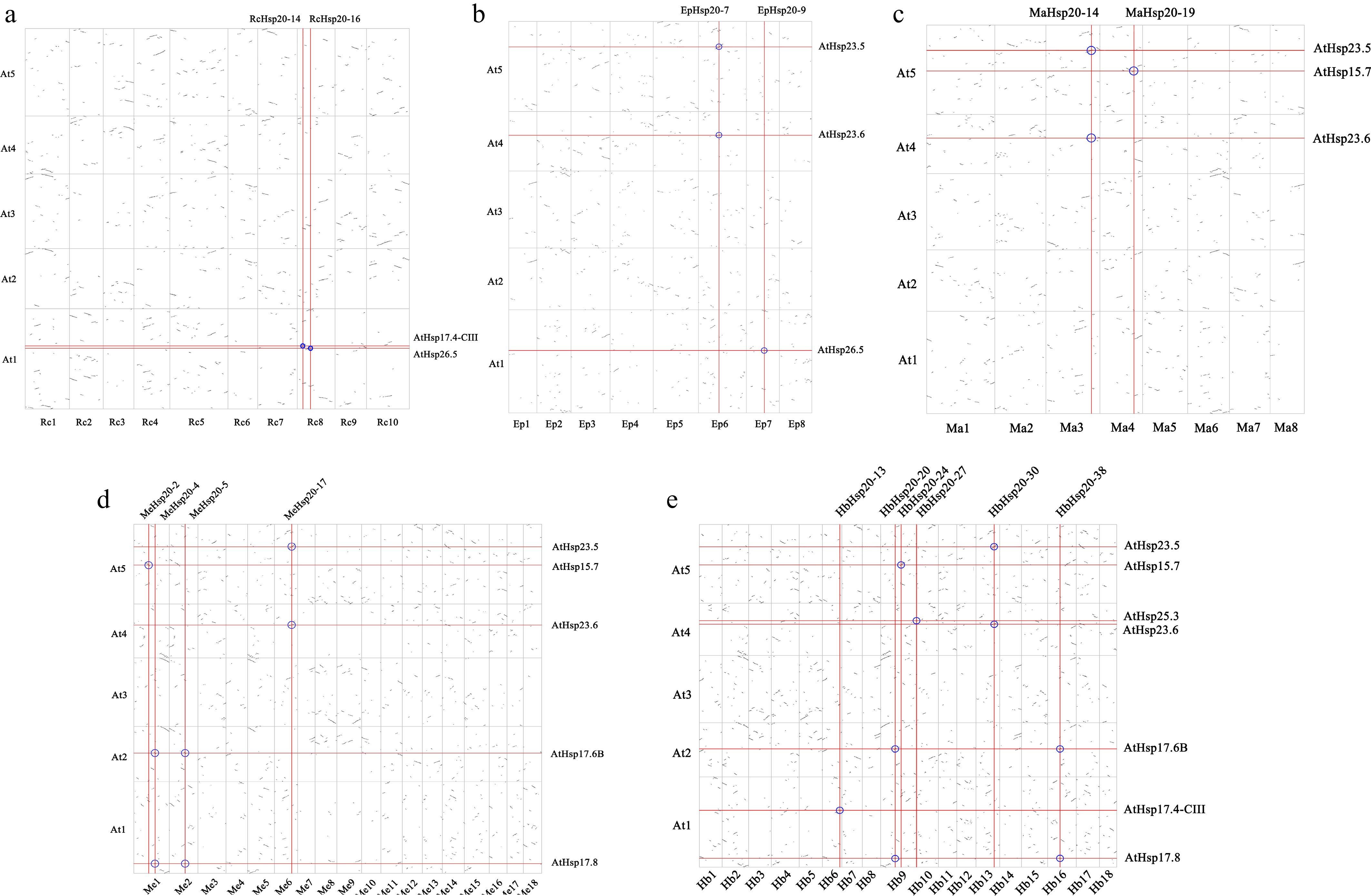
Figure 2.
Genome-wide duplication events between Arabidopsis and Euphorbiaceae. Gene dot plot between At and (a) Rc, (b) Ep, (c) Ma, (d) Me, and (e) Hb. At, A. thaliana; Me, M. esculanta; Rc, R. communis; Hb, H. brasiliensis; Ep, E. peplus; Ma, M. annua.
To analyze the evolution and function of intraspecific genes, a genome collinear dot plot of five species of Euphorbiaceae was drawn and analyzed (Supplementary Fig. S4). Different colored dots and lines represent different degrees of similarity. The most similar segments are red, followed by yellow, and gray. Each chromosome has distinct, intense red spots that allow each fragment to be regarded as highly comparable. In addition, many unfocused yellow dots were observed, caused by ancient γ-WGD events. However, there were a few gray fragments in the five Euphorbiaceae species, suggesting that older duplications are not apparent. In five Euphorbiaceae species, a diagonal line of red dots appeared on the genome dot plot, indicating the collinearity of each gene within the genome. Particularly, different numbers of collinear long fragments can be observed in the dot plots of different species. More obvious fragments were observed in the genome dot plots of cassava and rubber trees, implying that the traces were retained after large-scale genome duplication (Supplementary Fig. S4).
Chromosomal mapping and synteny analysis of Hsp20s
-
In order to explore the replication pattern and evolutionary mechanism of the Hsp20s in Euphorbiaceae, chromosome localization analysis was conducted on the seven Euphorbiaceae (Supplementary Fig. S5). Notably, an uneven distribution of Hsp20 genes was discovered throughout the chromosomes of each species. The result of MeHsp20s showed that Me14 contained the largest number (nine) of MeHsp20 genes, followed by Me2, and Me10. For RcHsp20s, most of the RcHsp20 genes were presented on Rc8 (eight genes), followed by Rc5, which contained four genes. The number of HbHsp20 genes across chromosomes ranged considerably, with Hb9 and Hb16 harboring eight genes. The remaining HbHsp20 genes were randomly distributed across the other rubber tree chromosomes and scaffolds. EpHsp20s were distributed on six E. peplus chromosomes. In contrast to the mere one to three genes found on other chromosomes, Ep8 harbored seven EpHsp20 genes. The 32 MaHsp20s were unevenly distributed across eight chromosomes. Ma1 and Ma7 harbored the greatest number of genes (seven members each), followed by Ma4. Due to the incomplete genome assembly of the physic nut and tung tree, all the JcHsp20 and VfHsp20 genes were mapped only at the scaffold level. The JcHsp20 and VfHsp20 genes were randomly located on the 16 and 36 scaffolds, respectively.
A comparative analysis of genomic syntenic blocks in A. thaliana and seven Euphorbiaceae was employed to elucidate the evolutionary history of the Hsp20 gene family (Fig. 3). A. thaliana had a collinear relationship with Euphorbiaceae, and homologous gene pairs were identified in seven studied species, including four pairs in castor bean, 26 pairs in rubber tree, 10 in E. peplus, 11 in M. annua, 15 pairs in cassava, two in physic nut and eight in tung tree (Fig. 3). Among them, castor bean, rubber tree, E. peplus, M. annua, cassava, physic nut, and tung tree respectively, had 2, 9, 3, 4, 11, 2, and 6 collinear relationships with the sHsp genes of Arabidopsis (Fig. 3; Supplementary Table S3). Among all the gene pairs, a pair of collinear genes (VfHsp20-52, AtHsp15.4) was classified into the same branch end in the phylogenetic tree (Fig. 1). The results of synteny analysis pointed to a potential relationship between the number of gene pairs and both genome size and gene family size. Furthermore, AtHsp17.4-CIII was collinear with Hsp20s of the other five species except for E. peplus and M. annua. Of all the Euphorbiaceae collinear Hsp20 genes, eight genes (JcHsp20-21 and RcHsp20-14, HbHsp20-13 and MeHsp20-33, HbHsp20-20 and MeHsp20-4, HbHsp20-30 and MeHsp20-17) were clustered at the branching end of the evolutionary tree (Fig. 1). Whereas, AtHsp25.3, AtHsp15.4, and AtHsp14.7 only formed collinear gene pairs with HbHsp20-27, VfHsp20-52, and VfHsp20-55, respectively. The fact that Ks for 12 gene pairs was negative, however, cannot be ignored; this suggests that the sequence divergence of these gene pairs is significant and evolved over time. In addition to these genes, the remaining paralogous genes underwent purifying selection, and the proteins encoded by these genes were likely to have eliminated harmful mutations and preserved the functions of the ancient Hsp20 protein (Supplementary Table S4).
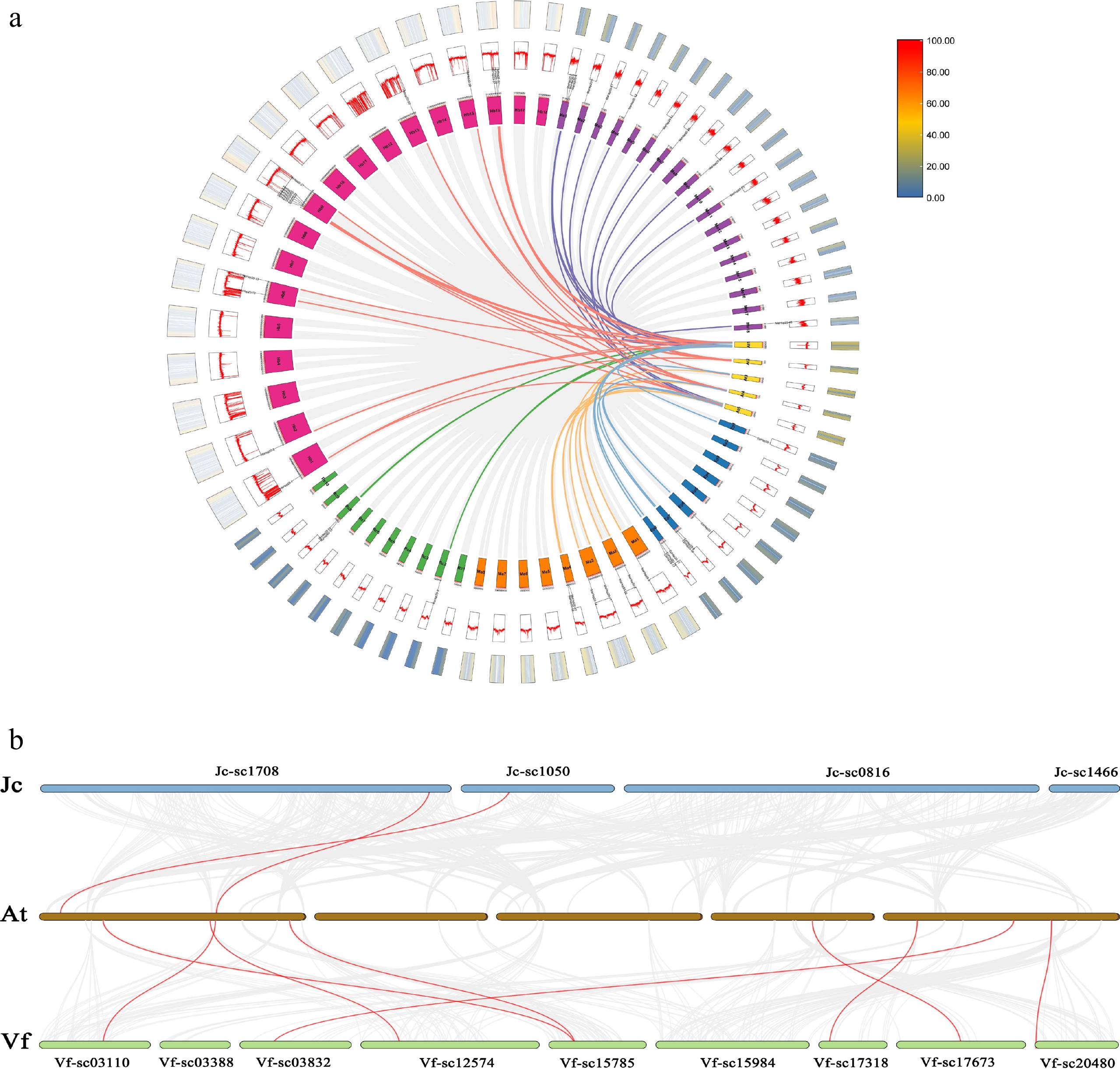
Figure 3.
Synteny relationships of Hsp20 genes between A. thaliana and Euphorbiaceae. (a) Synteny analysis of Hsp20 genes between Rc, Hb, Me, Ep, Ma, and A. thaliana. (b) Synteny analysis of Hsp20 genes between A. thaliana and physic nut, tung tree. Species chromosomal-level genome assemblies were co-mapped. Jc and Vf, with genome assembly at the scaffold level, were analyzed and drawn using scaffolds containing the Hsp20s. The A. thaliana, castor bean, rubber tree, cassava, E. peplus, and M. annua are labeled At, Rc, Hb, Me, Ep, and Ma in a circle diagram. At, Rc, Hb, Me, Ep, and Ma are shown in yellow, green, pink, purple, blue, and orange. The blue, green, and brown scaffolds and chromosomes represent physic nut, tung tree, and A. thaliana.
Conserved motifs, cis-regulatory elements, and transcription factors analysis of Hsp20s
-
Analysis of conserved motifs based on protein sequences was utilized to investigate the conservation of Hsp20 gene family members in Euphorbiaceae. Among all Hsp20 members, 234 Hsp20s (92.9%) contained Motif 2, 153 (60.7%) contained Motif 1, and 111 (44.0%) contained Motif 3 (Supplementary Table S5). In particular, the Hsp20s with comparable motifs were categorized within the same subfamily (Fig. 4a, b). Additionally, Motif 2 was a universal feature of all Hsp20s. Each subfamily was defined by specific motif signatures: Subfamily I contained Motif 1, 2, 3, and 5; Subfamily Po contained Motif 1, 2, and 4; Subfamily ER contained Motif 1, 2, 4, 5, and 10; Subfamily M contained only Motif 1 and 2. Motif 2 included the β2, β3, and β4 of the ACD structure; Motif 3 included β5 to β7; Motif 4 and Motif 9 included β8 and β9, respectively (Fig. 4c).
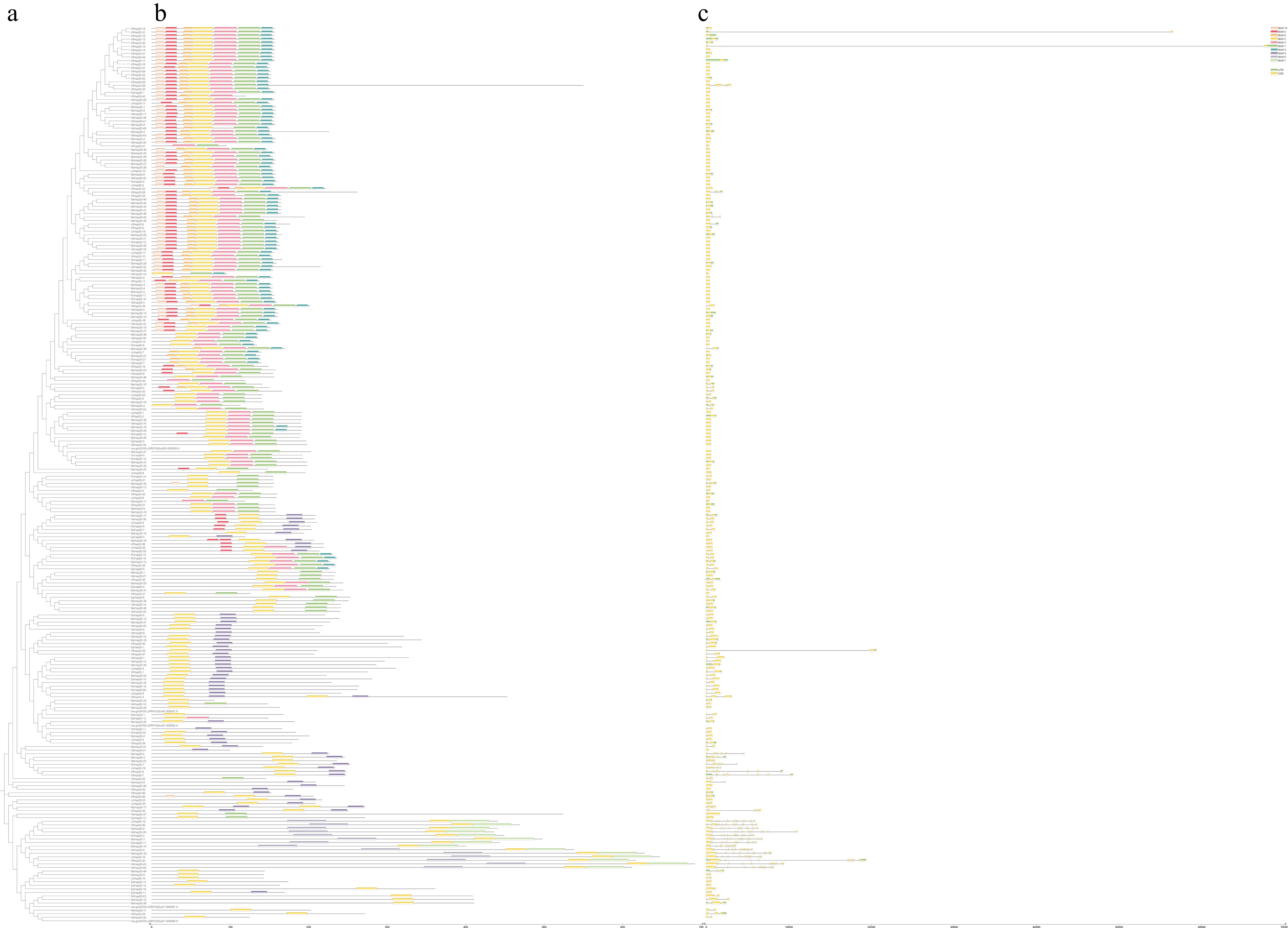
Figure 4.
Phylogenetic tree and conserved motifs of Hsp20 genes of Euphorbiaceae. (a) The phylogenetic tree of Hsp20 genes of seven Euphorbiaceae species; (b) the motif compositions of Hsp20s; (c) the conserved motif contained an ACD domain. Numbers on the X-axis represent the position of the amino acids. The relative frequency of amino acids in the motif is expressed in font size. The Motif logos are Motif 2, 3, 4, and 9 from top to bottom. The color boxes in Motif 2 represent the β2, β3, and β4; Motif 3 represents the β5, β6, and β7; Motif 4 represents the β5; and Motif 9 represent the β9 of the ACD structure.
The extraction of promoter regions from 252 genes and predictive analysis of their cis-acting elements were instrumental in illuminating the transcriptional regulation of Hsp20s in the examined species. Three categories of cis-elements were identified, including plant growth and development, phytohormone-related, and external stress cis-elements (Supplementary Fig. S6). All Hsp20 genes had light-responsive elements whose proportion was approximately 46.63% to 58.39%. Interestingly, E. peplus, which originated in subtropics, has the lowest proportion of light-responsive elements compared to other species (Supplementary Table S6). Among the elements of the phytohormone-responsive category, most of the Hsp20 genes possessed abscisic acid- and MeJA-responsive elements (Fig. 5), which implied that the function of Hsp20s might be regulated by abscisic acid and MeJA. Notably, the number of elements in different species varied greatly (Fig. 5; Supplementary Table S6). MeHsp20s contained 144 MeJA-responsiveness elements, while RcHsp20s had only 32. Compared with other species, MeHsp20s consisted of many individual response elements (Supplementary Table S6). The abundance of light response and hormone response factors demonstrated that Hsp20 gene expression in Euphorbiaceae was governed by light and hormones. More diverse roles of Hsp20 genes in stress responses were implied by the identification of diverse stress-related cis-acting elements (e.g., defense, drought-inducible, and anaerobic-inducible). Additionally, cis-elements involved in growth and development, such as circadian control, meristem expression regulatory, and endosperm expression regulatory elements, were also found in all studied species (Supplementary Table S6). The results pointed to a potential role of Hsp20 genes in regulating the growth and development of Euphorbiaceae plants.
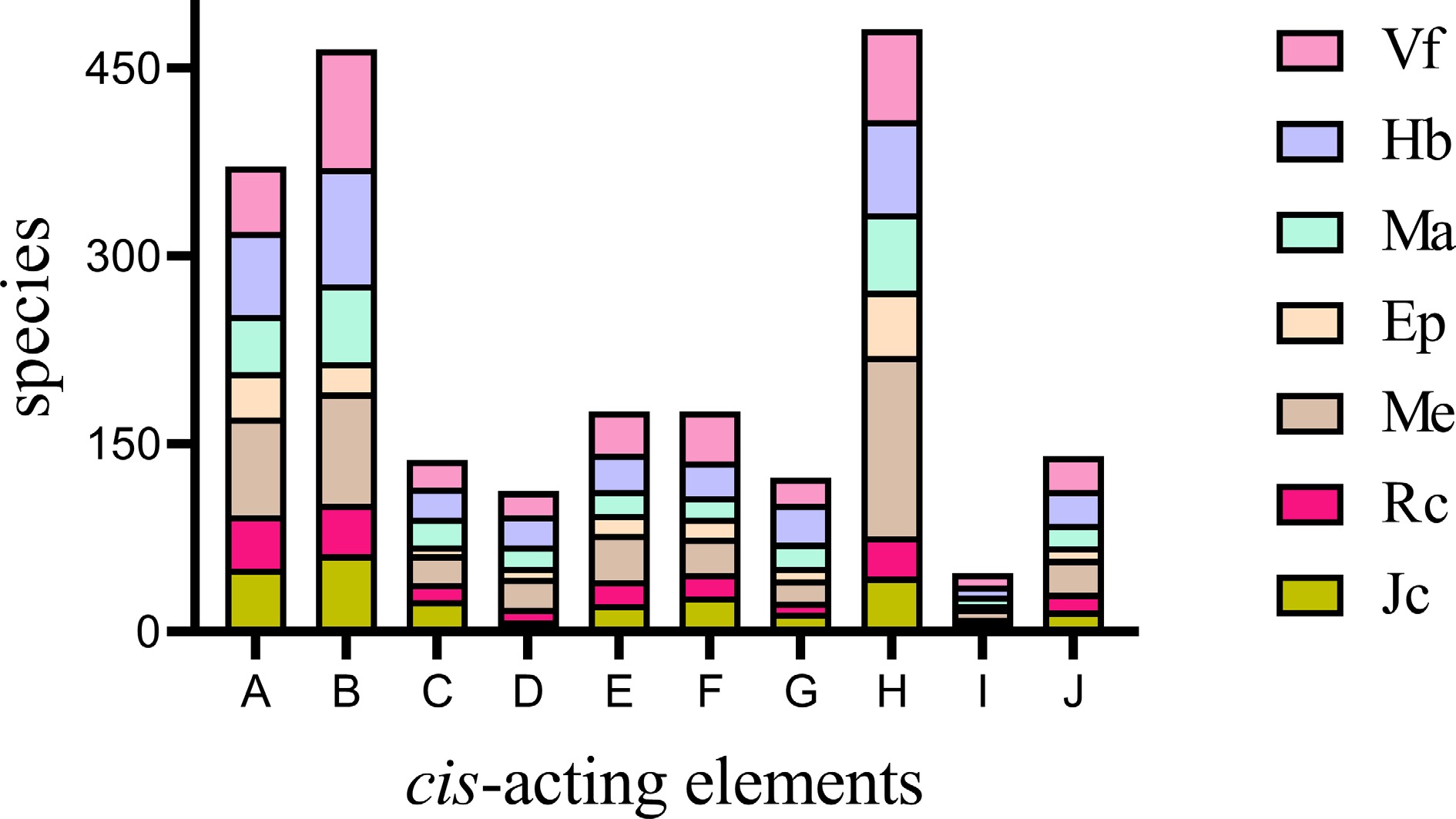
Figure 5.
Cis-acting element distribution in the promoter of Hsp20s. The number of cis-acting elements in different species is shown in different colors. A–J represent different cis-acting elements. A: abscisic acid responsiveness; B: anaerobic induction; C: auxin responsiveness; D: defense and stress responsiveness; E: drought inducibility; F: gibberellin responsiveness; G: low-temperature responsiveness; H: MeJA-responsiveness; I: MYB binding site; J: salicylic acid responsiveness.
Hsp20s of different Euphorbiaceae species contained different types and quantities of transcription factor binding sites (Fig. 6; Supplementary Table S7). The Hsp20s of the remaining species all had 43 distinct transcription factor binding sites, while JcHsp20s and MeHsp20s had 39, and 40 distinct binding sites, respectively (Supplementary Table S7). Notably, the number of the top 10 transcription factor binding sites contained by Hsp20s was similar across species. JcHsp20s and RcHsp20s contained the largest number of Dof (DNA binding with one finger) binding sites, while the others contained ERF (ethylene-responsive factor) binding sites. In particular, the transcription factors ERF, Dof, and MYB (v-myb avian myeloblastosis viral oncogene homolog) were the most abundant binding sites across all studied species except cassava.
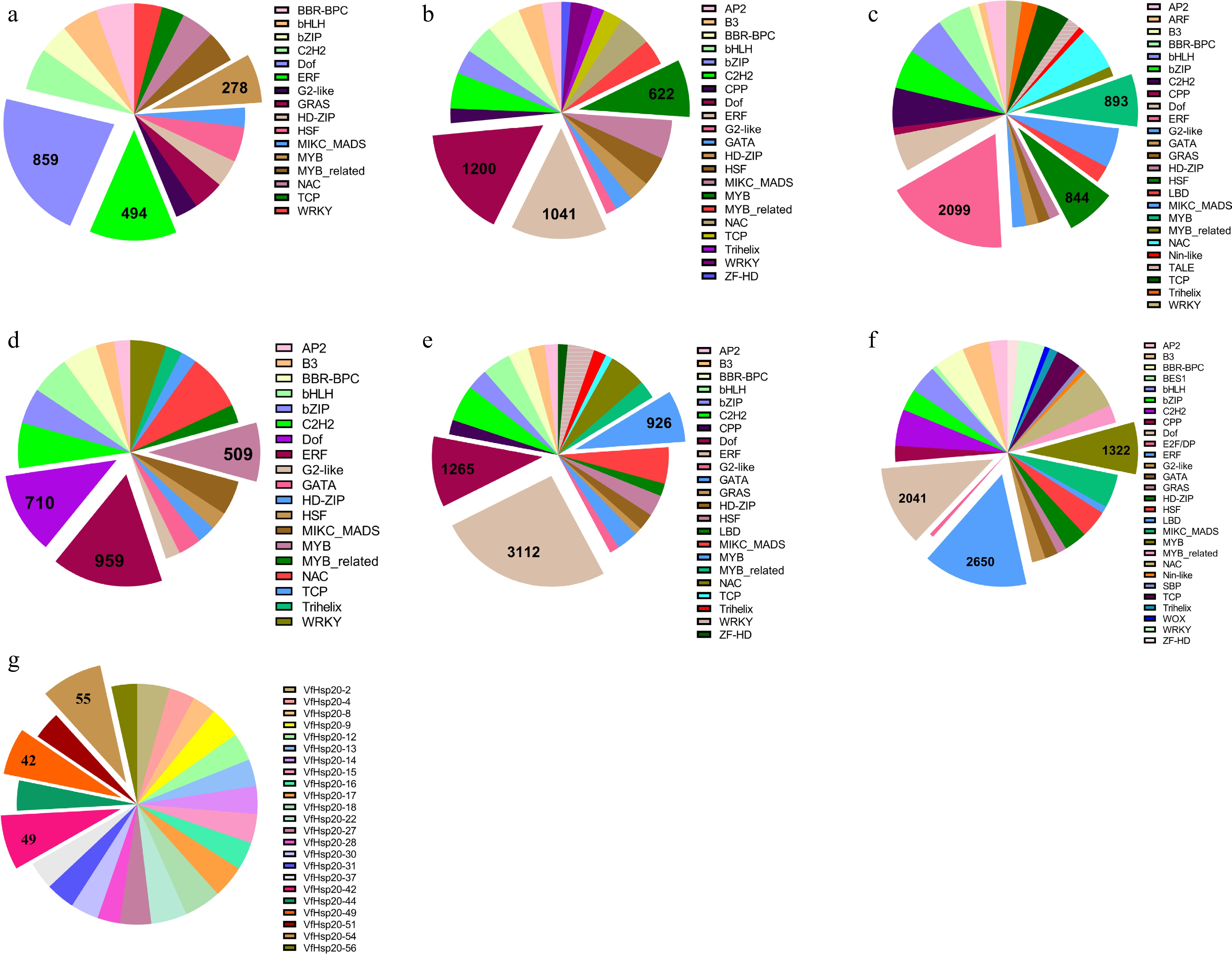
Figure 6.
Composition of predicted transcription factor binding sites in Hsp20s. The abundance of different transcription factors is displayed on the pie chart. (a) J. curcas; (b) R. communis; (c) M. esculenta; (d) E. peplus; (e) M. annua; (f) H. brasiliensis; and (g) V. fordii.
It is well known that Hsf binds to the upstream region of Hsp to regulate expression[6]. All cassava Hsp20s contained 844 Hsf binding sites, the largest number of Hsf binding sites among all studied species (Fig. 6; Supplementary Table S7). The number of Hsf binding sites contained in RcHsp20, JcHsp20, MaHsp20, EpHsp20, HbHsp20, and VfHsp20 was 318, 200, 378, 155, 700, and 836, respectively (Supplementary Fig. S7; Supplementary Table S7). Among the studied species, E. peplus had the lowest proportion of the Hsp20 genes containing the Hsf binding site (Supplementary Table S8). Although Hsf binding sites were ubiquitous, being identified in 90% of MeHsp20s, their distribution was markedly skewed, as the three most abundant genes collectively constituted merely 14.95% of the total sites. This indicated that Hsf binding sites were widely distributed in MeHsp20s and might be involved in controlling Hsp20 transcription (Supplementary Fig. S7; Supplementary Table S8). Of the top three Hsp20s containing the most Hsf binding sites in the same species, four genes (JcHsp20-1, JcHsp20-11, MeHsp20-9, VfHsp20-49) each phylogenetically clustered at the branch terminus with Hsp20s from other Euphorbiaceae (Fig. 1). Except for Hsf binding sites, the number of ERF binding sites was the largest (Supplementary Table S8). The proportion of Hsp20s containing Hsf and ERF binding sites differed in different species (Fig. 6). For the Hsf binding site, 45 MeHsp20s (90%) contained the Hsf binding sites, while only 25 of 32 MaHsp20s contained the Hsf binding sites. For the ERF binding site, 959 were found in 16 of 17 EpHsp20s, and 1,908 ERF binding sites were found in 27 of 50 EpHsp20s. Only four genes (RcHsp20-6, MeHsp20-11, EpHsp20-3, and EpHsp20-8) contained many Hsf and ERF binding sites.
Analysis of key amino acid variation sites and tertiary structure
-
To determine the functional sites, the conservation and scores of each site of the Hsp20 protein were calculated based on the homology model of Arabidopsis sHsp (PDBID: 7BZW). The results of variation sites showed that E113, D123, W156, L168, and A181 were highly conserved, and these amino acids were respectively located in β2, β3, β6, β7, and β8, which constituted the ACD domain (Supplementary Fig. S2; Supplementary Table S8). In the studied Euphorbiaceae, most of the L168 residues mutated into F residues. K133 and I134 were the conserved sites located in β4, while L188, P193, and K194 were the conserved sites located in β9. I143, G145, and E146 were all highly conserved sites in β5, among which I143 was a buried structural residue. Except for I143, the other highly conserved residues were all exposed functional residues. The Hsp20 protein consisted of 12 peptide chains, with different types and numbers of secondary structures that made up each chain (Supplementary Fig. S8). The 12 peptide chains that make up the Hsp20 protein are composed of various secondary structures, including a comparatively high percentage of random coils.
Protein–protein interaction (PPI) network and KEGG pathway of Hsp20
-
The highest homologous STRING proteins were identified by the Hsp20 protein sequences of seven Euphorbiaceae species, respectively, to predict the protein–protein interaction (PPI) network. The homologous proteins were matched based on the highest bit score by default, which led to the identification of 25 MeHsp20, 14 RcHsp20, 11 JcHsp20, 15 EpHsp20, 25 MaHsp20, 32 HbHsp20, and 49 VfHsp20 proteins within the PPI networks (Fig. 7). As shown in Fig. 7, the interacting partners of Hsp20 proteins were not only proteins from the Hsp family, but also proteins from other family members. Except for Hsp gene family members, the majority of Hsp20 protein interaction partners were BAG (Bcl-2 associated athanogene) and CLPB (casein lytic proteinase B), which implied that Hsp20 proteins might be regulated by other proteins in Euphorbiaceae. In cassava, MeHsp20-1 had 16 interaction partners, followed by MeHsp20-17 (13), MeHsp20-12 (10), and MeHsp20-3 (10). Nevertheless, more than 10 HbHsp20s had more interaction partners. Five HbHsp20s (HbHsp20-19/24/25/30/31) had 19 interaction partners.
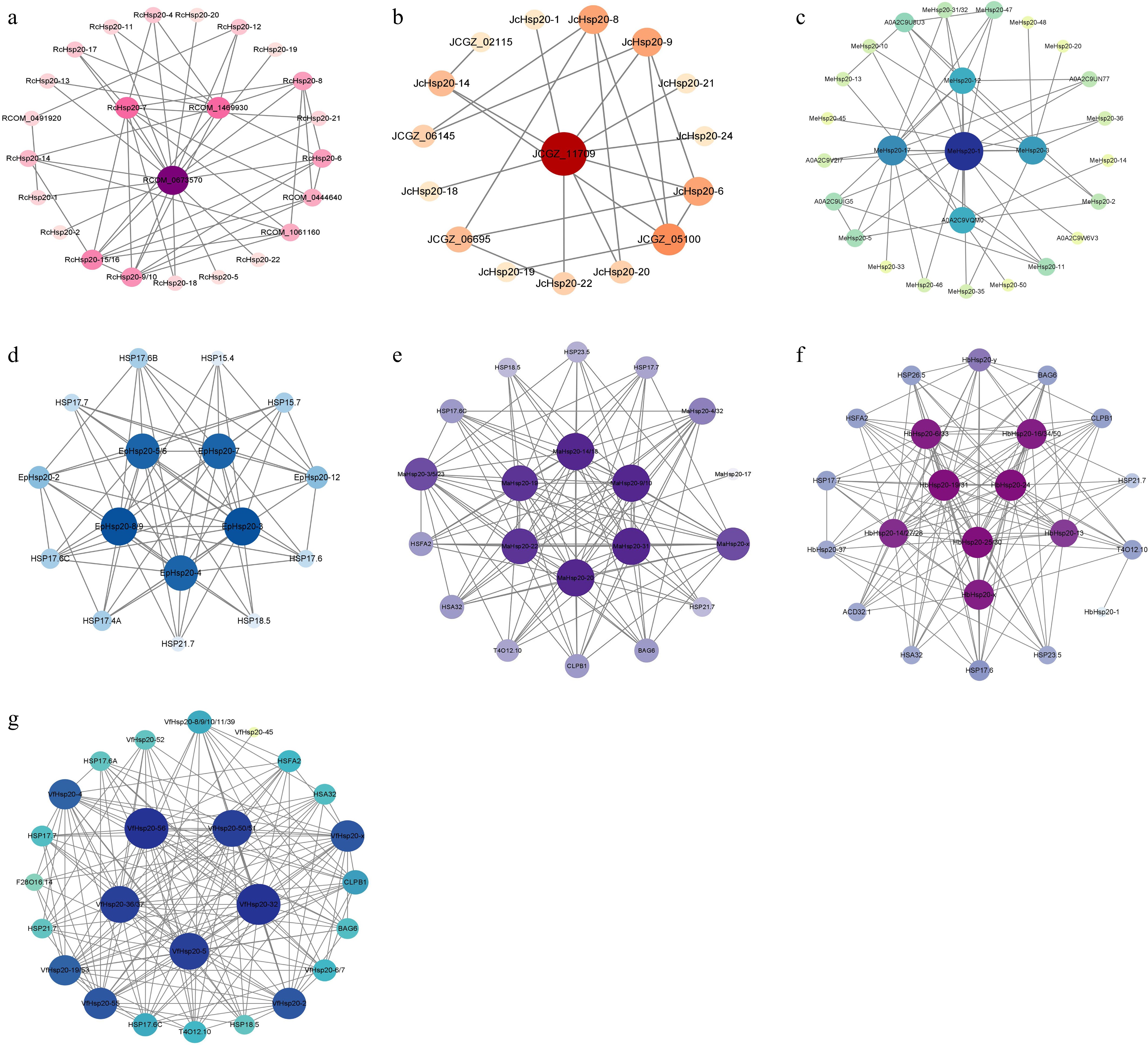
Figure 7.
PPI network of Hsp20 proteins. Circle size and color depth correspond to the number of interacting proteins. (a) R. communis; (b) J. curcas; (c) M. esculenta; (d) E. peplus; (e) M. annua; (f) H. brasiliensis; and (g) V. fordii.
Heat shock proteins, as molecular chaperones, participate in the formation and degradation of proteins. In cells, proteins fold with the help of lumenal chaperones in the endoplasmic reticulum (ER). The synthesized peptide chains were transferred into the ER and were glycosylated. The correctly folded peptide chains were further transported to the Golgi complex. In contrast, the misfolded peptide chains remained in the ER and bound to the BiP, then were degraded by the proteasome during ER-associated degradation (ERAD) (Supplementary Fig. S9). The ERAD process requires the involvement of the Hsp protein.
Expression pattern profiling of MeHsp20s
-
The transcriptome expression data of Euphorbiaceae were downloaded from the GEO database to determine the expression patterns of Hsp20s in Euphorbiaceae. Among the seven Euphorbiaceae species, cassava is the main source of energy for more than 800 million people worldwide and can be used in a variety of industries, including bioenergy, industry, and medicine. Therefore, cassava was selected as a representative species for transcriptome analysis of Hsp20 genes.
The expression profile of 50 MeHsp20 genes was visualized by the method of hierarchical clustering (Supplementary Fig. S10). The Hsp20 genes of cassava showed significant tissue specificity, with the mean expression values of MeHsp20 genes maintained at high levels in root tubers (SR), leaves, and midveins. In contrast, the 50 MeHsp20s displayed noticeably low expression in somatic embryos (OES), with relative expression levels of only about 6. Compared with other genes, MeHsp20-29 had the highest expression level in SR, while MeHsp20-1 and MeHsp20-23 were the most highly expressed genes in leaves and midveins, respectively. Notably, the MeHsp20 genes displayed distinctive tissue-specific expression patterns. The relative expression of MeHsp20-3 was high in leaves and midveins; however, it was barely expressed in SR. The relative expression of MeHsp20-20 was the opposite. Other genes, such as MeHsp20-14 showed a stable and relatively high expression in the three tissues. Among the 50 MeHsp20 genes, MeHsp20-29 was highly expressed in all tissues, especially in SR.
Under drought stress, the results showed that MeHsp20 genes displayed different expression. More importantly, a clustering analysis demonstrated that the MeHsp20 genes could be grouped into two clusters (group I and group II) (Fig. 8a). The expression of genes in group 1 was generally lower than that in group 2. The low-expression group (group I) and high-expression group (group II) included 29, and 24 genes, respectively. In group II, there were eight genes with relatively high expression, among which one gene (MeHsp20-27) had the highest expression in different varieties, and 2 genes (MeHsp20-5 and MeHsp20-20) had more stable expression (Fig. 8a). However, under cold stress, only two genes (MeHsp20-15 and MeHsp20-26) had higher expression levels than other genes. The expression levels of most genes were below 1 or even undetectable (Fig. 8b). Ultimately, MeHsp20-26 was upregulated in response to escalating cold stress (Fig. 8b).
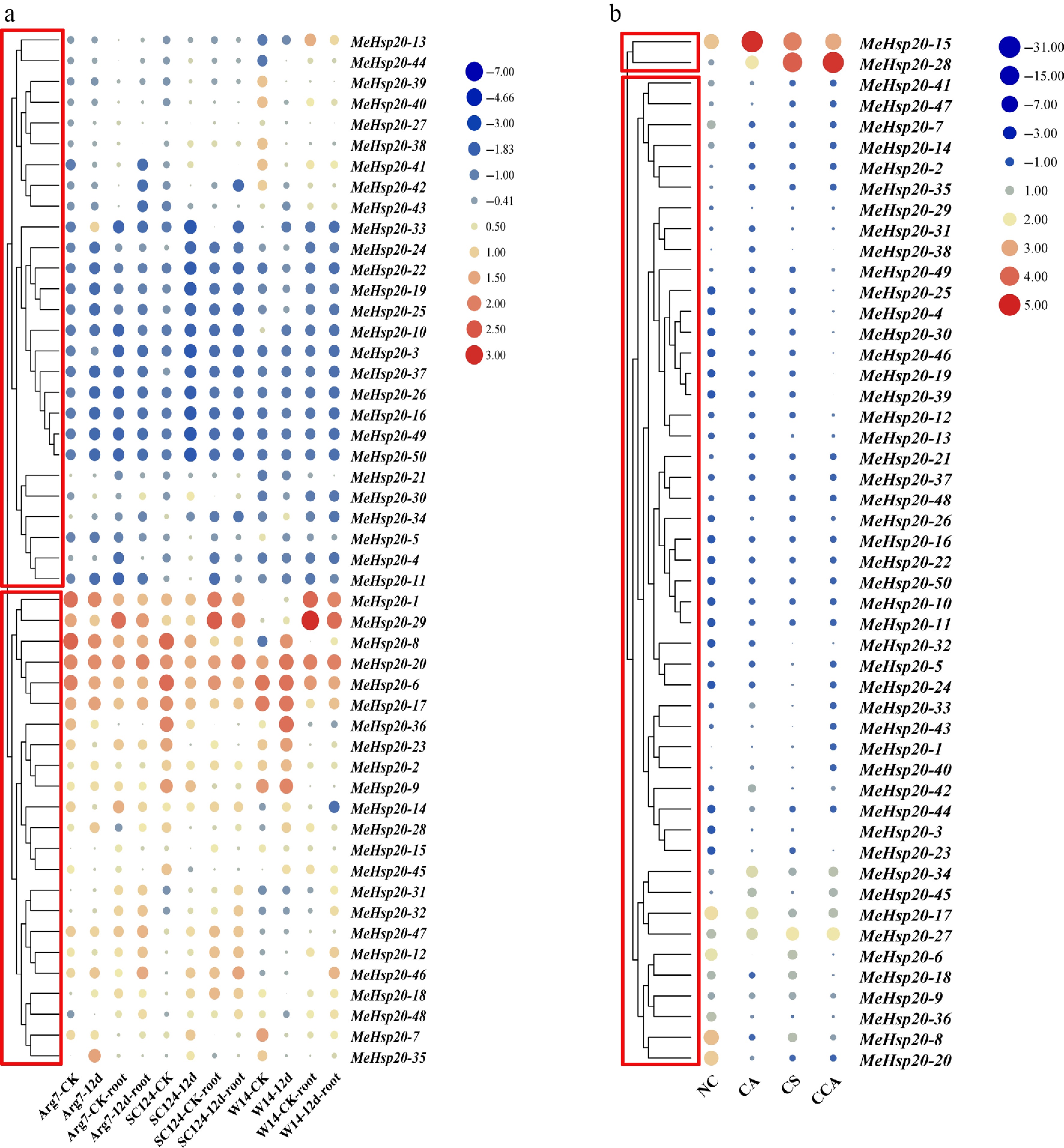
Figure 8.
The expression profile of MeHsp20 genes under (a) drought, and (b) cold stress. NC: control group; CA: gradual chilling acclimation; CS: chilling shock; CCA: chilling stress after chilling acclimation. Control group; color scale denotes relative expression values: blue (values < 0) for downregulated genes; red (values > 0) for upregulated genes.
-
Euphorbiaceae plants and their products have demonstrated high economic and practical value, and are widely used in industries such as food processing, manufacturing, aerospace, and medicine. The Euphorbiaceae species, such as M. esculenta, E. pepluss, and H. brasiliensis, are important economic plants or food sources worldwide. In this paper, a comprehensive analysis of the Hsp20 gene family in seven representative Euphorbiaceae plants was conducted, encompassing biochemical characteristics, functions, and evolutionary relationships.
The genome-wide analysis established the presence of 252 Hsp20 genes across the seven Euphorbiaceae genomes. A greater number of Hsp20s were identified in species from the Crotonaceae and Acalyphoideae subfamilies than in the Euphorbioideae subfamily, which evolved in the subtropics (Supplementary Fig. S1; Table 1). Within the Euphorbiaceae family, the species with the smallest genome size was not the one with the fewest copies of the Hsp20s. Except for the Euphorbiaceae family, Dendrobium catenatum, whose genome size is 1.01 Gb, contains only 18 Hsp20s[39,40]. The phenomenon indicated that the genome size cannot explain the variation in the Hsp20 gene copy number. The conserved motif, chromosome localization, and phylogenetic analysis results verified the classification and conservation of the Hsp20 gene family in Euphorbiaceae (Figs 2, 4). As observed in rice and tomato, the CI subfamily contained the most Euphorbiaceae Hsp20s, and the members within each subfamily shared similar conserved motifs (Fig. 1)[6,41]. In addition to confirming the conservation of coding sequences and conserved motifs of Hsp20s in Euphorbiaceae, variations were also found in their conserved motifs. Though the components of the ACD domain were distributed in four conserved motifs, mutations or deletions existed in the components of the ACD domain of several Hsp20s in Euphorbiaceae, suggesting potential functional divergence (Fig. 4c; Supplementary Fig. S2). Although there are variations of the Hsp20 gene within the Euphorbiaceae family, the results of the phylogenetic analysis suggested that Hsp20s in the Euphorbiaceae and A. thaliana have gradually shown a tendency to diverge during evolution. Moreover, it is speculated that gene pairs divided into the end of the same branch and from different species may have the same function (Fig. 1). Interestingly, the distribution of the Hsp20s on chromosomes in Euphorbiaceae was similar to that in tomato and coix, being mainly located at both ends, particularly at the distal ends of the short arms (Supplementary Fig. S5)[41,42].
Whole-genome duplication (WGD) plays a pivotal role in angiosperm morphological and physiological diversity, and the evolution of plant stress resistance[43]. Arabidopsis thaliana, a model species for the eudicot clade, has undergone two rounds of WGDs (β WGD and α WGD)[44]. The ancient γ-whole genome triplication event approximately 117 million years ago (Mya) was shared by all core-eudicots, including Arabidopsis, poplar, and the four studied species (cassava, castor bean, rubber tree, and physic nut)[45]. This might explain why the AtHsp20s found through the WGD events were basically the same (Fig. 2). Subsequently, around 39 to 47 Mya, cassava experienced a genome-wide replication after diverging from physic nut and castor bean. A parallel WGD was also observed in the rubber tree[46]. The analysis results of the WGD events within the genome were consistent with these findings (Supplementary Fig. S4).
To develop greater awareness of the evolutionary relationship between Hsp20s in Arabidopsis and Euphorbiaceae, interspecific collinearity and WGD analyses were performed. Thirty seven collinear Hsp20 gene pairs between Euphorbiaceae and Arabidopsis were observed, of which M. esculenta accounted for the highest number (11 pairs) (Fig. 2; Supplementary Table S3). The low prevalence of synteny (14.68%) between Euphorbiaceae and Arabidopsis Hsp20s indicated substantial evolutionary divergence, notwithstanding their notable conservation within the Euphorbiaceae family. Notably, the location of 24 out of the 37 gene pairs in chromosomal syntenic blocks provided evidence that they stemmed from ρ WGD (Fig. 2). Among the 24 Hsp20s, 23 Hsp20s were classified into CI, CIII, Po, and M subfamilies, while HbHsp20-27 belonged to the P subfamily (Fig. 1). Interestingly, 24 Hsp20s in Euphorbiaceae were co-located with eight of the 19 AtHsp20s in syntenic blocks, which may illustrate that they were derived from sHsps in Arabidopsis and had similar functions. The nearly identical conserved motifs of genes in the same subfamily further supported this hypothesis (Fig. 4). With the exception of R. communis, most Euphorbiaceae Hsp20s shared collinear gene pairs with AtHsp23.5 and AtHsp23.6 (Fig. 3). Given the significant upregulation of AtHsp23.5 under heat stress, it is plausible that its homologous genes are also involved in high-temperature response (Fig. 3)[47]. Four genes (MeHsp20-17, EpHsp20-7, MaHsp20-14, and HbHsp20-30) located in collinear blocks with the AtHsp25.3-P, a HS-responsive gene, may also be involved in responding to HS[48]. HbHsp20-27, as the sole gene in the AtHsp25.3 collinear block, was speculated to be induced by HS. In addition to these 24 Hsp20s, the origin of other Hsp20 genes in Euphorbiaceae may be ascribed to intraspecific duplication events, primarily tandem and segmental duplication.
Under various stress conditions, heat shock proteins are produced and are activated by upstream genes to perform specific functions. Analysis of the promoter regions of the Euphorbiaceae Hsp20 genes revealed that they mainly contained 11 cis-acting elements, predominated by light responsiveness elements and then by MeJA-responsiveness elements (Supplementary Fig. S6; Supplementary Table S6). A parallel cis-acting element analysis of the genes in Dendrobium officinale was consistent with the result, with the analysis in Coix also yielding an abscisic acid responsiveness element[42,49]. Additionally, analysis of wheat sHsp26 promoter elucidated that it is responsive to multiple abiotic stresses, encompassing not only heat but also salt, cold, and drought conditions[16,50]. It can be inferred that the expression of Hsp20s in Euphorbiaceae may be regulated by hormones to resist adverse environments.
Many studies have shown that Hsp20s respond to temperature stress. The expression of over half of the CrHsp20s in the C. rosea seedlings increased significantly after 2 h of heat shock[51]. In Dendrobium catenatum, all 18 DcHSP20 genes were upregulated under high temperature, while six genes were significantly upregulated under low-temperature conditions, with overexpression of DcHSP20-12 conferred thermotolerance[40]. Cold stress stimulated the upregulation of TaHSP16.9H-CI and TaHSP23.5B-MTI in bread wheat[52]. Five out of 51 GmHsp20s in soybean responded to cold stress[53]. Transcriptome expression analysis revealed that only MeHsp20-15 and MeHsp20-28 displayed an upregulation tendency under the cold stress (Fig. 8b), implying that the majority of Hsp20s in Euphorbiaceae may respond to HS rather than cold stress. Commonly, Hsf regulates the transcription of Hsp genes by recognizing and binding to the heat shock elements within the Hsp promoters when plants experience HS. HSP17 from wheat was identified as the target gene of HSFA2h, and their expression levels were positively correlated under HS in transgenic Arabidopsis[54]. Using gel shift and LUC-reporter experiments, Reddy et al. demonstrated that HvHsfB2c may regulate the expression of HvHsp17.5-CI under HS[55]. Likewise, out of the 252 Hsp20s from the Euphorbiaceae, 207 had Hsf binding sites, suggesting that they were engaged in reacting to HS (Supplementary Fig. S7; Supplementary Table S7).
Furthermore, it is frequently reported that Hsp20s react to other kinds of stress. Stronger resistance to drought and salt was conferred by overexpressing Arabidopsis thaliana Hsp17.6A.[56]. Pepper CaHSP16.4 enhanced the elimination of reactive oxygen species in response to heat and drought stress[57]. A rapid and significant upregulation of multiple MeHsp20s was observed after drought stress across different varieties, especially MeHsp20-35, MeHsp20-28 and MeHsp20-9, suggesting that they could be involved in drought stress (Fig. 8a). In Dendrobium catenatum, the germination rate, fresh weight, and root length of the DcHSP20-12 overexpressing transgenic Arabidopsis lines were significantly higher than those of the wild-type (WT) plants, indicating that this Hsp20 participated in plant growth and development[40]. Similarly, the consistently stable expression of MeHsp20-17 across 11 tissues suggested that it may also contribute to this process (Fig. 6; Supplementary Fig. S10). The remaining Hsp20s might have different functional roles (Supplementary Tables S7, S8). Transcriptome analysis provides support for the inference. In cassava, some Hsp20s showed high expression levels in specific tissues, while others were not, which indicated that different genes play specialized roles in specific tissues (Supplementary Fig. S10).
Nonetheless, it can't be ignored that Hsp20s may function with other proteins or transcription factors to regulate plant resistance to HS. In Arabidopsis, HSP21 abundance could be regulated by FtsH6 (filamentation temperature sensitive)[58]. The enrichment of ERF transcription factor binding sites in the promoters of Euphorbiaceae Hsp20s implicates a functional link to the ethylene signaling pathway (Supplementary Tables S7, S8). The results of the PPI network also indicated that the Hsp20 protein of the Euphorbiaceae could form interaction relationships with other proteins, such as Bcl-2-associated athanogene. Bcl-2-associated athanogene (BAG), belonging to the NEF chaperone family, is known to mediate interactions between the Hsp chaperone system and its substrates[59]. In tomato, BAG9 was highly induced and interacted with Hsps in the cytoplasm, promoting the stability of Hsps under HS[59]. In the rubber tree, 12 HbHsp20s were predicted to interact with BAG, implying that they may be protected by BAG (Fig. 7f). The results strongly demonstrated that the Hsp20 gene can play discrepant functions across different signaling pathways. Future work will further elucidate the functions of these stress-responsive Hsp20 genes in abiotic stress resistance.
-
Hsp20 genes have been identified and characterized in several species; however, limited attention has been given to plants within the Euphorbiaceae family. This study conducted a genome-wide identification of 252 putative Hsp20 genes across seven Euphorbiaceae species, followed by a comprehensive analysis of their functions, phylogeny, and evolutionary relationships. Moreover, the genome comparison indicated that all syntenic gene pairs underwent segmental duplication and were subjected to purifying selection, with 24 Hsp20s in Euphorbiaceae potentially originating from β WGD. Additionally, MeHsp20-17, EpHsp20-7, MaHsp20-14, and HbHsp20-30 may play a role in environmental adaptation. The observed differences in tissue-specific expression and the expression levels of different genes under diverse stress conditions highlight the functional diversity acquired by Hsp20s during evolution. In conclusion, these results provide a foundation for further exploration of the functions of the Hsp20 gene family in Euphorbiaceae plants, particularly under adverse conditions, and offer new insights into the regulatory mechanisms governing the Hsp20 gene family.
-
The authors confirm their contributions to the paper as follows: study conception and design: Chen Yinhua, Yu X; data collection: Zheng L, Chen Yuhua, Zhang Z, Wang H; analysis and interpretation of results: Zheng L, Ambuyoc MKA; draft manuscript preparation: Zheng L, Jin J, Hamidou Abdoulaye A. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in the NCBI repository. These data were derived from the following resources available in the public domain: PRJNA324539; PRJNA227109; PRJNA246428.
-
This work was supported by the National Natural Science Foundation of China (32260468) and the earmarked fund for the China Agriculture Research System (CARS-11-HNCYH).
-
The authors declare that they have no conflict of interest
-
accompanies this paper online at: https://doi.org/10.48130/tp-0026-0017.
-
Received 6 February 2026; Accepted 17 April 2026; Published online 12 June 2026
-
# Authors contributed equally: Linling Zheng, Maria-Kristina Abello Ambuyoc
- Supplementary Table S1 Genome information of seven Euphorbiaceae.
- Supplementary Table S2 Hsp20 genes identified from Euphorbiaceae and their physicochemical properties information.
- Supplementary Table S3 Synteny gene pairs between Euphorbiaceae and Arabidopsis.
- Supplementary Table S4 Selection pressure analysis of Euphorbiaceae and Arabidopsis.
- Supplementary Table S5 The conserved motifs information of Hsp20s in Euphorbiaceae.
- Supplementary Table S6 The number of important cis-acting elements in different species
- Supplementary Fig. S4
- Supplementary Table S7 The number of different transcription factor binding sites of Hsp20s.
- Supplementary Table S8 The number of ERF and Hsf binding sites of Hsp20s.
- Supplementary Fig. S1 Genome evolutionary tree of seven Euphorbiaceae.
- Supplementary Fig. S2 Multiple sequence alignment of Hsp20.
- Supplementary Fig. S3 Genome dot plot within A. thaliana and 5 Euphorbiaceae species.
- Supplementary Fig. S4 Genome-wide duplication events within species.
- Supplementary Fig. S5 Chromosome distribution of Hsp20s from seven Euphorbiaceae species.
- Supplementary Fig. S6 Cis- acting element distribution in putative promoters of Hsp20 genes.
- Supplementary Fig. S7 Prediction of Hsf transcription factor binding sites of Hsp20s.
- Supplementary Fig. S8 Sequence conservation and variation among seven Hsp20 Euphorbiaceae and Arabidopsis.
- Supplementary Fig. S9 S9 KEGG pathway of Hsp20.
- Supplementary Fig. S10 Heatmap representation and hierarchical clustering of the MeHsp20 genes in various cassava tissues.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng L, Ambuyoc MKA, Jin J, Chen Y, Hamidou Abdoulaye A, et al. 2026. Heat shock protein 20 gene family involved in the temperature adaptation of typical Euphorbiaceae. Tropical Plants 5: e019 doi: 10.48130/tp-0026-0017 

Heat shock protein 20 gene family involved in the temperature adaptation of typical Euphorbiaceae
- Received: 06 February 2026
- Revised: 04 April 2026
- Accepted: 17 April 2026
- Published online: 12 June 2026
Abstract: In higher plants, heat shock protein 20 (Hsp20) is integral to growth, development, and temperature stress adaptation. While most Euphorbiaceae species originated in the tropics and subtropics, they have since proliferated across both tropical and temperate regions. Investigating Hsp20 function and evolution in Euphorbiaceae clarifies the role in temperature adaptation. This work performed a genome-wide analysis of 252 Hsp20 genes across seven representative Euphorbiaceae species, enabling phylogenetic delineation of 13 distinct subfamilies. Among the 37 paralogous gene pairs shared with Arabidopsis, 24 Hsp20s were anchored within conserved syntenic blocks. Castor bean presented only two syntenic Hsp20s with AtHsp20s, whereas cassava and rubber tree possessed four and six Hsp20s, respectively. Transcriptome expression analysis of MeHsp20s unveiled that at least 16 Hsp20s across various tissues were involved in modulating cassava growth and development. Additionally, 25 MeHsp20s were drought-responsive, while only two MeHsp20s were induced by cold stress, which implies that Hsp20s in Euphorbiaceae may play a role in helping plants adapt to high temperatures rather than cold stress. These findings thereby establish a foundation for future investigations into the molecular mechanisms by which Hsp20s underlie growth and thermal responses in Euphorbiaceae plants.
-
Key words:
- Euphorbiaceae /
- Hsp20 /
- Gene evolution /
- Abiotic stress