









-
The disjunct distribution of plant lineages across the Northern Hemisphere represents a classic biogeographic pattern that has long motivated research into its underlying mechanisms and spatiotemporal history[1]. Three major hypotheses have been advanced to explain these patterns. The Boreotropical hypothesis posits that widespread forests across northern mid- to low-latitudes during the Late Cretaceous to Early Eocene[2] underwent contraction and fragmentation due to subsequent global cooling, leading to modern relictual distributions—a scenario supported by studies of groups such as Pachygoneae[3] and Juglandaceae[4]. In contrast, the land-bridge hypothesis emphasizes the role of former terrestrial connections, particularly the Beringian and North Atlantic routes, which facilitated floristic interchange during periods of low sea level[5,6]. The eventual submergence of these corridors led to the isolation of once-continuous populations, as evidenced in taxa such as Cladrastis[7] and Nyssa. A third model, the Out-of-Xizang hypothesis, proposes that late Miocene cooling—intensified by the strengthening of the Asian monsoon—enabled cold-adapted lineages to disperse from the Qinghai-Xizang Plateau into temperate regions of East Asia and North America[8,9], with empirical support from groups including Berberidaceae[10] and Polygonatum[11]. Among these disjunctions, the Eastern Asian–North American pattern has been the most intensively studied across a wide range of plant groups using phylogenetic, paleobotanical, and biogeographic approaches[12,13]. By comparison, disjunctions spanning Asia, Europe, and North America have received far less attention, leaving key aspects of their biogeographic origins and evolutionary trajectories unresolved.
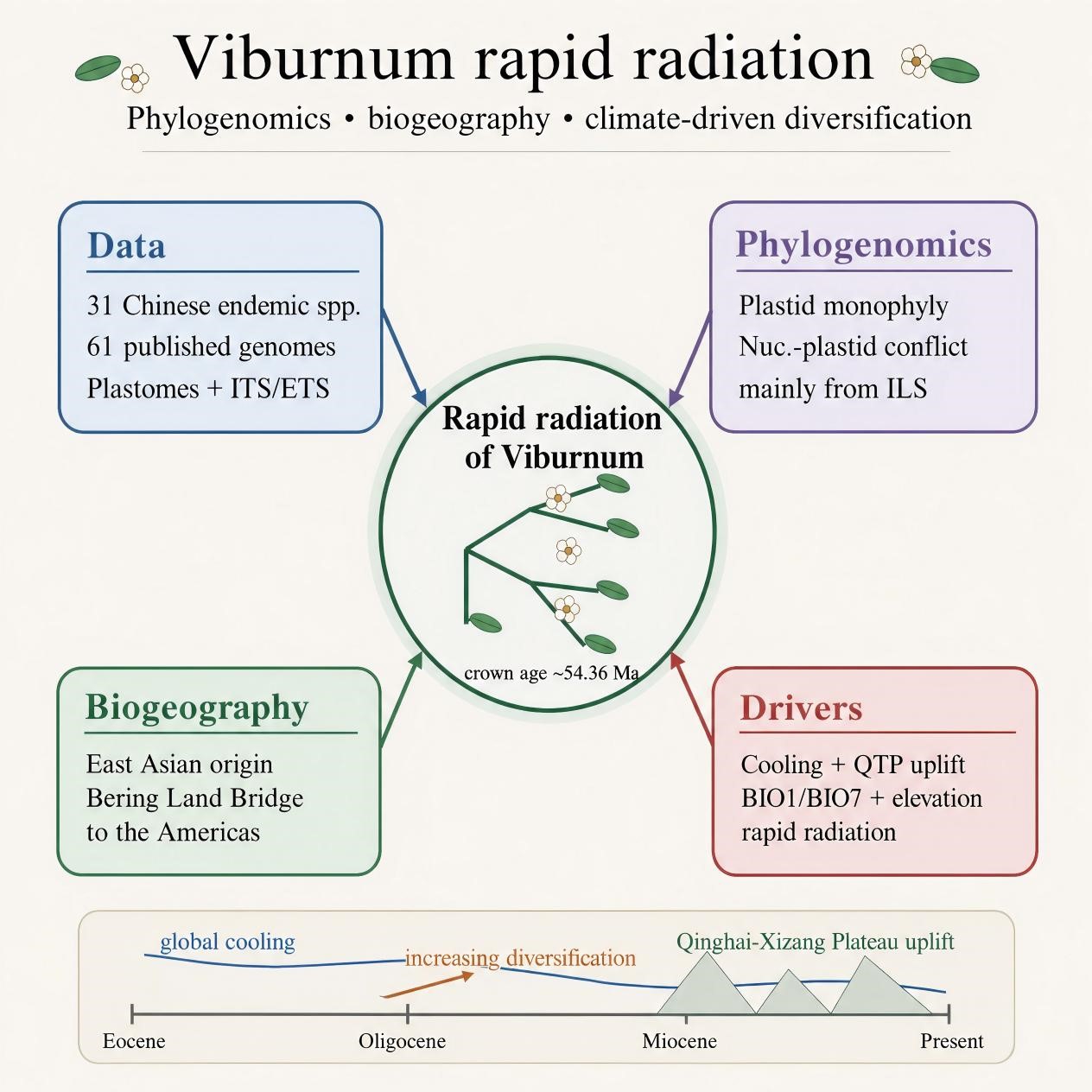
The disjunct distribution of Viburnum across Eurasia and North America establishes it as an ideal model system for investigating the biogeographic history of Northern Hemisphere intercontinental disjunctions[14−16]. Nevertheless, substantial gaps remain in our understanding of the distribution patterns, which represent a critical geographic area for the group. Landis et al.[15] categorized the global distribution of Viburnum into six major regions (Southeast Asia, East Asia, Europe, North America, Central America, and South America) and proposed a Beringian migration route into North America from East Asia. However, the principal drivers underlying the global radiation and distribution of Viburnum remain poorly resolved. China serves as a central diversity hotspot for Viburnum, hosting approximately 62% of all recognized species worldwide[17]. The genus is widely distributed across most Chinese provinces, with only a few northern regions lacking native representatives, underscoring China's role as a major center of diversity and diversification in Asia. Recent phylogeographic analyses suggest that temperate forests in Southwestern China functioned as primary centers of speciation, from which lineages subsequently dispersed to eastern and southeastern regions[17]. The nine sections of Viburnum exhibit distinct morphological traits that reflect their ecological adaptations (Fig. 1). Section Platyphylla bears laterally arranged flowers and large leaves (Fig. 1a), while section Odontotinus is characterized by toothed leaf margins (Fig. 1b). Section Megalotinus features exceptionally large leaves (Fig. 1c), and section Pseudopulus has densely pubescent leaves (Fig. 1d). Section Pseudotinus displays sympodial branching (Fig. 1e), and section Opulus is notable for palmately lobed leaves (Fig. 1f). The core section Viburnum has naked buds (Fig. 1g), section Tinus is distinguished by persistent, leathery leaves (Fig. 1h), and section Thyrsosma features thyrsoid inflorescences (Fig. 1i). This complex history of range expansions, coupled with repeated ecological transitions between colder deciduous and warmer evergreen temperate forests, likely promoted frequent niche shifts and triggered multiple evolutionary radiations within the genus[16,18]. Elucidating the biogeographic history of Viburnum not only enhances our understanding of Northern Hemisphere floristic evolution but also provides a critical theoretical foundation for guiding biodiversity conservation, sustainable agricultural production, and the rational utilization of genetic resources[19]. In summary, Viburnum serves as an ideal system for examining competing biogeographic hypotheses. Leveraging phylogenetic discordance between plastid and nuclear genomes allows us to refine historical inferences, thereby providing a robust phylogenetic framework for the conservation and utilization of Viburnum species.

Figure 1.
Morphological characterization of flower and leaf blades of Viburnum fruits, Sect. Platyphylla: (a) Viburnum laterale; Sect. Odontotinus: (b) Viburnum erosum; Sect. Megalotinus: (c)Viburnum lutescens; Sect. Pseudopulus: (d) Viburnum hanceanum; Sect. Pseudotinus: (e) Viburnum sympodiale; Sect. Opulus: (f) Viburnum koreanum; Sect. Viburnum: (g) Viburnum keteleeri 'Sterile'; Sect. Tinus: (h) Viburnum atrocyaneum; Sect. Thyrsosma: (i) Viburnum erubescens.
The phylogenetic relationships within Viburnum have been the subject of multiple studies, yet substantial challenges to the complete resolution of the group persist. Previous studies have predominantly relied on chloroplast DNA fragments[18−22]. While these studies established a foundational understanding, their limited genomic sampling has resulted in several persistently unresolved nodes. For instance, analyses based on chloroplast markers failed to confirm the monophyly of sections Megalotinus and Odontotinus[23].
Early phylogenetic studies based on chloroplast DNA fragments established the foundational framework of Viburnum relationships, but often suffered from limited resolution at deep nodes[18,20]. Winkworth & Donoghue[21] employed the duplicated nuclear gene GBSSI (granule-bound starch synthase I) to reconstruct Viburnum phylogeny, recovering several major clades consistent with chloroplast-based trees while providing independent support for the monophyly of sections Opulus, Tinus, and Viburnum. Their study highlighted the utility of low-copy nuclear genes for resolving relationships in a genus with a history of rapid diversification. More recently, Ran et al.[23] assembled complete chloroplast genomes of 17 Chinese Viburnum species and conducted comparative genomic analyses, revealing highly conserved plastome structure and identifying several mutational hotspots. Their phylogenetic analyses supported the monophyly of most sections but failed to resolve the relationships among sections Megalotinus, Odontotinus, and Opulus, suggesting that even whole chloroplast genomes may have limitations in fully disentangling the evolutionary history of rapidly radiating lineages. Taken together, these studies underscore the need for expanded taxonomic sampling and integration of multiple genomic data sources.
Studies incorporating broader taxonomic sampling and multi-locus datasets, such as Spriggs et al.[16]—which analysed 138 species under multiple partition schemes for nuclear and chloroplast data—have provided support for many major clades. Similarly, Clement et al.[20] employed maximum likelihood and Bayesian inference to reconstruct relationships across 113 species, yet key uncertainties remain, particularly concerning the phylogenetic placement of the Pseudotinus and Opulus clades. In summary, while significant progress has been made in clarifying the broad phylogenetic framework of Viburnum, existing studies indicate that limited sampling and a reliance on chloroplast and single-locus nuclear data are insufficient to fully resolve its evolutionary history.
Numerous studies have employed nuclear and organellar genomic data to estimate divergence times within Viburnum. Among these, Winkworth & Donoghue[21] used the GBSSI nuclear gene to estimate divergence times within Viburnum, recovering a crown age of approximately 30 Ma (Oligocene) and supporting an Asian origin with subsequent dispersals to North America. Their time estimates, however, were constrained by limited fossil calibrations and relatively sparse taxon sampling compared to later studies. Ran et al.[23] further contributed to divergence time estimation by analyzing complete chloroplast genomes of Chinese Viburnum species, recovering a crown age of approximately 48 Ma (Eocene) and highlighting the role of the Qinghai-Xizang Plateau uplift in driving diversification of East Asian lineages (see Supplementary Materials, Supplementary Fig. S1).
Spriggs et al.[16] inferred the divergence rate of Viburnum and explored the potential morphological and ecological drivers underlying its diversification. Their analyses support an Eocene origin of Viburnum in the tropical forests of Southeast Asia, followed by three independent radiations of temperate lineages during the Miocene. By incorporating fossil pollen evidence and expanding molecular datasets, they further proposed a crown age of 55 Ma (95% HPD: 46–73 Ma), notably older than previous estimates. Landis et al.[15] suggested that two distinct lineages of Viburnum dispersed from Asia to North America during the Eocene, around 40 Ma. Among these, the lineage corresponding to the clade Oreinotinus underwent southward expansion from Mexico starting in the late Miocene (8–12 Ma), followed by a rapid diversification phase, as further supported by Clement et al.[20].
Although previous studies have investigated the phylogeny of Viburnum, research that integrates chloroplast and nuclear gene data and combines ecological niche modeling with diversification rate and morphological evolution analyses to explore its biogeographic origins remains limited. In this study, we investigated the potential drivers of rapid radiation within the genus Viburnum by sequencing the chloroplast genomes and ITS/ETS regions of 31 Chinese endemic species, while integrating previously published chloroplast genome data. First, we reconstructed the phylogenetic tree and framework for this genus based on the chloroplast genome, ITS region, and ETS region, respectively. We then estimated species divergence times based on the chloroplast phylogenetic tree. To evaluate biogeographic scenarios underlying the current distribution and diversification of Viburnum, we analyzed the influence of environmental variables on the global diversity distribution patterns of Viburnum species and combined the projections with phylogenetic and divergence-time evidence to assess three prevailing hypotheses: the Northern Tropical Origin Hypothesis, the Land Bridge Hypothesis, and the Out-of-Xizang Hypothesis. This integrative approach provides a comprehensive basis for understanding the evolutionary and biogeographic mechanisms that shaped the diversification history of Viburnum. At the same time, we acknowledge that our phylogenetic inferences are primarily based on maternally inherited chloroplast genomes, supplemented by nuclear ribosomal ITS/ETS data, and thus represent a plastid-based perspective on Viburnum evolution. Future studies incorporating genome-wide nuclear data will be necessary to fully resolve cytonuclear discordance and test for potential hybridization.
-
Genomic DNA was extracted from 31 Viburnum samples using a cetyltrimethylammonium bromide (CTAB) protocol[24,25]. The sampled taxa represent all nine recognized sections of the genus, encompassing major morphological and phylogenetic diversity: the bare bud group (sect. Viburnum), the sympodial group (sect. Pseudotinus), the bulbous kernel group (sect. Tinus), the conical group (sect. Thyrsosma), the pteridophyte group (sect. Pseudopulus), the lateral flower group (sect. Platyphylla), the large leaf group (sect. Megalotinus), the toothed leaf group (sect. Odontotinus), and the cleft leaf group (sect. Opulus). In addition to these newly sequenced samples, we retrieved 61 published chloroplast genomes of Viburnum and eight outgroup taxa from GenBank for comparative analysis (Supplementary Table S1).
Prior to whole-genome sequencing, genomic DNA quality was assessed using a bioanalyzer. Only samples exhibiting a minimum mass of 0.8 µg were selected for sequencing, which was performed on the BGISEQ-500 platform at BGI Genomics (Shenzhen, China). Sequencing was performed using double-ended sequencing libraries with insert fragments of 300–500 bp. Raw sequencing reads were processed using SOAPfilter v2.2 to remove adapter sequences.
Chloroplast genomes were assembled de novo using GetOrganelle v1.7.5.1[26] under default parameters. To evaluate assembly quality and structural integrity, all reconstructed genomes were visualized in Bandage v0.8.1[27]. Each assembly was inspected for circularization and tetrameric structure to confirm completeness and avoid chimeric contigs.
Data analysis
-
To ensure structural accuracy and proper gene orientation, a global alignment of all chloroplast genome sequences was conducted using the Mauve Genome plugin implemented in Geneious Prime 2023.0.4[28]. Annotation of assembled genomes was performed by referencing the published chloroplast genome of Viburnum schensianum (NC_056104) within the same software environment.
A comparative genomic analysis was carried out across 92 Viburnum species and eight outgroup taxa to assess sequence variability and structural conservation. The organization of genes at partition boundaries was initially visualized using Geneious Prime. Subsequently, mVISTA[29] was employed to quantify GC content and total gene counts across taxa, while also facilitating the identification of regions exhibiting high sequence divergence. Finally, multi-genome alignments were generated using Mauve[30] to evaluate large-scale structural conservation and conformational similarity among chloroplast genomes.
Phylogenetic reconstruction was performed based on chloroplast genome sequences from 92 Viburnum taxa and eight outgroup species. Multiple sequence alignment was conducted with the MAFFT plugin implemented in PhyloSuite v1.2.3[31]. Maximum likelihood phylogenies were inferred using IQ-TREE[32], also within the PhyloSuite environment, with optimal substitution models selected automatically. In parallel, to reconstruct species relationships using nuclear markers, we analyzed the ITS and ETS regions by combining 31 newly generated sequences. Alignment and phylogenetic inference followed the same pipeline as described for the chloroplast data, ensuring methodological comparability between the plastomic and nuclear datasets.
Evolutionary divergence time inference
-
Divergence times were estimated using BEAST v2.6.6[33] under a GTR + G substitution model and a birth-death tree prior. Two independent Markov chain Monte Carlo (MCMC) runs were conducted, each with 500 million generations and sampling every 1,000 steps. Fossil-based calibration points were applied using lognormal priors: the node encompassing Diabelia serrata and D. sanguinea as well as Dipelta floribunda was constrained to 35 Ma (95% CI: 36.0–40.18 Ma[34]), and the root age was set to 115 Ma (95% CI: 115–123 Ma[35]). Convergence and sufficient sampling were assessed using Tracer v1.7[36], confirming that all effective sample size (ESS) values exceeded 200. The first 25% of trees from each run were discarded as burn-in, and the remaining trees were combined using LogCombiner. A maximum clade credibility (MCC) tree with mean node heights was generated in TreeAnnotator, and the resulting chronogram with 95% highest posterior density (HPD) intervals was visualized in FigTree v1.4.2[36].
Species diversity estimates
-
To investigate diversification dynamics within Viburnum, we applied Bayesian Analysis of Macroevolutionary Mixtures (BAMM v2.5.0[37]) to the time-calibrated chloroplast genome phylogeny. Prior configurations were optimized using the setBAMMpriors function in the R package BAMMtools v2.1.10[37]. Four independent Markov chain Monte Carlo (MCMC) runs were conducted, each with 10 million generations and sampling every 1,000 steps. The first 25% of samples were discarded as burn-in after verifying stationarity and adequate mixing of likelihood traces. Convergence was assessed using the coda v0.19-4.1 package[38], ensuring all parameters achieved effective sample sizes (ESS) greater than 200.
To evaluate whether climatic fluctuations influenced macroevolutionary rates in Viburnum, we compared inferred diversification rate shifts through time with a continuous paleotemperature curve derived from global benthic foraminiferal records[39]. Temporal correlation between these rate-through-time and temperature-through-time profiles was assessed to identify potential climate-driven evolutionary dynamics.
Biogeographic analysis
-
To reconstruct the historical biogeography of Viburnum, we employed BioGeoBEARS v1.1.2[40] in R to compare six biogeographic models: DEC, DEC + J, DIVALIKE, DIVALIKE + J, BAYAREALIKE, and BAYAREALIKE + J. Species distribution data were compiled from both Plants of the World Online (
https://powo.science.kew.org ) and original field observations. We defined six operational biogeographic units based on major floristic regions: (A) East Asia (China, Japan, South Korea, North Korea, and Mongolia); (B) Southeast Asia (south of the Lancang-Mekong River); (C) South Asia (Himalayas and regions to the south); (D) Europe; (E) North America; and (F) South America. These units reflect major geological and biogeographic barriers, such as the Lancang-Mekong River and the Himalayas, which have likely shaped species distributions. The maximum number of ancestral areas per node was set to six.Impacts of environmental variables on global patterns of Viburnum diversity
-
Global occurrence records of Viburnum were obtained from the Global Biodiversity Information Facility (GBIF, 2024;
https://doi.org/10.15468/dl.vacj4p ). To assess the influence of environmental factors on species richness, we constructed regression models with species richness per 100 km × 100 km grid cell as the response variable (Fig. 2), and all 19 bioclimatic variables from WorldClim v2[41] along with elevation as predictors. Predictor values were averaged within each grid cell, and species richness was standardized using Z-score transformation ('stats' v4.6.0 in R). We first fitted ordinary least squares (OLS) multiple regression models and assessed multicollinearity using variance inflation factors (VIF) computed with the 'car' package v3.1-3[42]. All retained variables had VIF values below 6.005, indicating acceptable levels of collinearity (Supplementary Table S2). Following variable screening based on statistical and ecological relevance, six predictors were retained: BIO1 (annual mean temperature), BIO2 (mean diurnal range), BIO7 (temperature annual range), BIO12 (annual precipitation), BIO17 (precipitation of driest quarter), and elevation. To account for spatial autocorrelation, we implemented a spatial error autoregression model (SAR) using the "spdep" package v0.7-7[43]. Model comparison based on Akaike's Information Criterion (AIC), coefficient of determination (R2), and Moran's I of residuals indicated that the SAR model (AIC = 6,410; R2 = 0.477; Moran's I ≈ −0.038, p = 0.999) substantially outperformed the OLS model (AIC = 7,395; R2 = 0.119). The SAR model effectively eliminated spatial autocorrelation and provided a significantly better fit. Therefore, we based our final inference—including residual diagnostics and visualization of standardized regression coefficients—on the SAR model to ensure robust interpretation of environmental effects on species richness.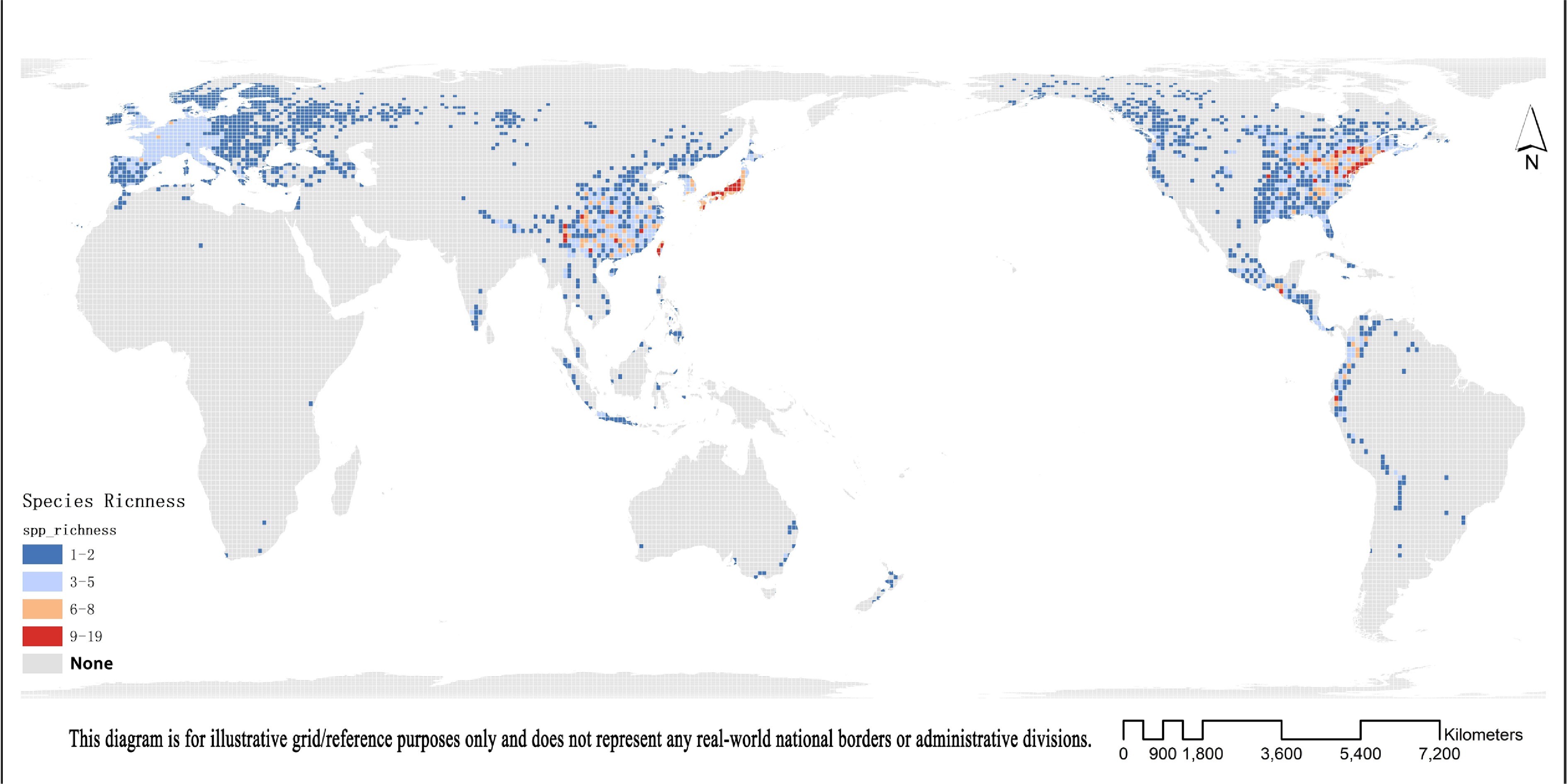
Figure 2.
Global patterns of Viburnum species diversity mapped in 100 km × 100 km grid cells.
-
The chloroplast genomes of Viburnum exhibited limited structural variation across the sampled species (Supplementary Table S3). Total genome length ranged from 158,031 bp to 158,726 bp, representing a maximum divergence of only 695 bp. The large single-copy (LSC) region varied from 86,838 bp to 88,546 bp (difference = 523 bp), while the small single-copy (SSC) region showed greater length heterogeneity, ranging from 18,231 bp to 18,754 bp (difference = 1,703 bp). Each chloroplast genome displayed the typical quadripartite structure, comprising one LSC, one SSC, and two inverted repeat (IR) regions, the latter ranging between 26,153 bp and 26,544 bp. Overall GC content was highly conserved, varying narrowly from 38.0% to 38.3%, with a mean value of 38.1% (Supplementary Table S4).
Phylogenetic results
-
The chloroplast genome phylogeny revealed strongly supported relationships among major clades of Viburnum (Fig. 3). Sect. Odontotinus and Sect. Megalotinus were recovered as a sister lineage with maximum support (BS = 100). This clade (Sect. Odontotinus + Sect. Megalotinus) subsequently formed a well-supported sister group (BS = 100) with Sect. Opulus. The combined clade ([Sect. Odontotinus + Sect. Megalotinus] + Sect. Opulus) was sister to Sect. Tinus, again, with full support (BS = 100). In a distinct lineage, Sect. Pseudopulus and Sect. Platyphylla were resolved as sister taxa (BS = 100), and this pair formed a highly supported clade (BS = 100) with Sect. Thyrsosma. Additionally, Sect. Pseudotinus and Sect. Viburnum was strongly supported as a sister group (BS = 100). Notably, Viburnum plicatum (traditionally placed in Sect. Pseudopulus) was nested within Sect. Pseudotinus, indicating a possible misalignment between current taxonomic boundaries and phylogenetic history.
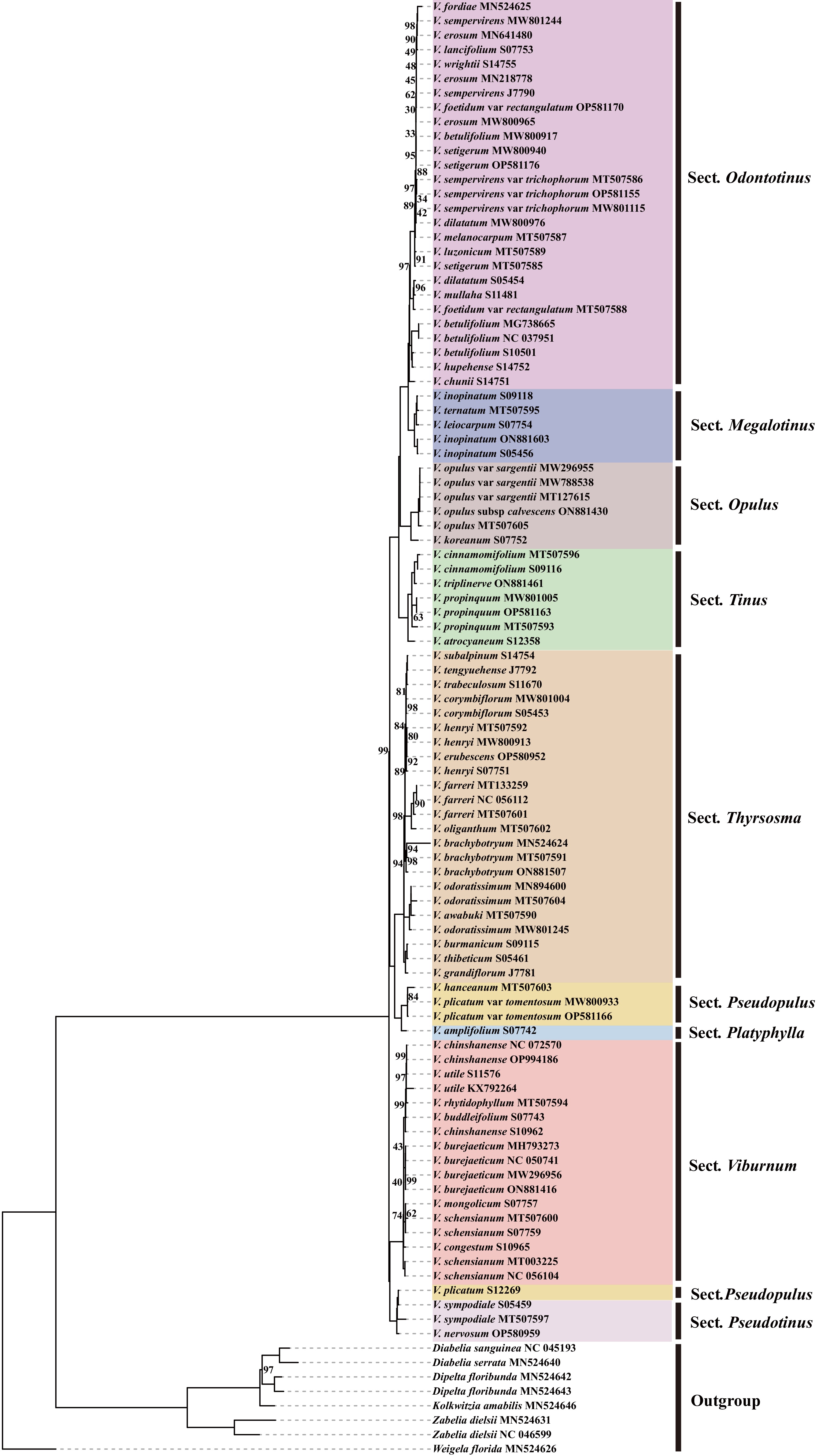
Figure 3.
IQtree constructed based on chloroplast whole genome data.
Simultaneously, we constructed a phylogenetic tree based on Internal Transcribed Spacer (ITS) and extra-transcriptional spacer (ETS) sequences and compared it with the chloroplast genome phylogenetic tree (Fig. 4). In contrast, the phylogeny reconstructed from nuclear ITS and ETS sequences presented both congruent and conflicting topologies (Fig. 5). Section Thyrsosma, Platyphylla, Viburnum, Pseudotinus, Pseudopulus, Megalotinus, and Tinus were each recovered as monophyletic. However, Sect. Megalotinus was embedded within Sect. Odontotinus, suggesting incomplete lineage sorting or hybridization. Sect. Thyrsosma and Sect. Platyphylla formed a highly supported sister clade (BS = 100), which was sister to Sect. Viburnum with moderate support (BS = 89). Additionally, Sect. Pseudotinus and Sect. Pseudopulus was strongly supported as a sister lineage (BS = 100).
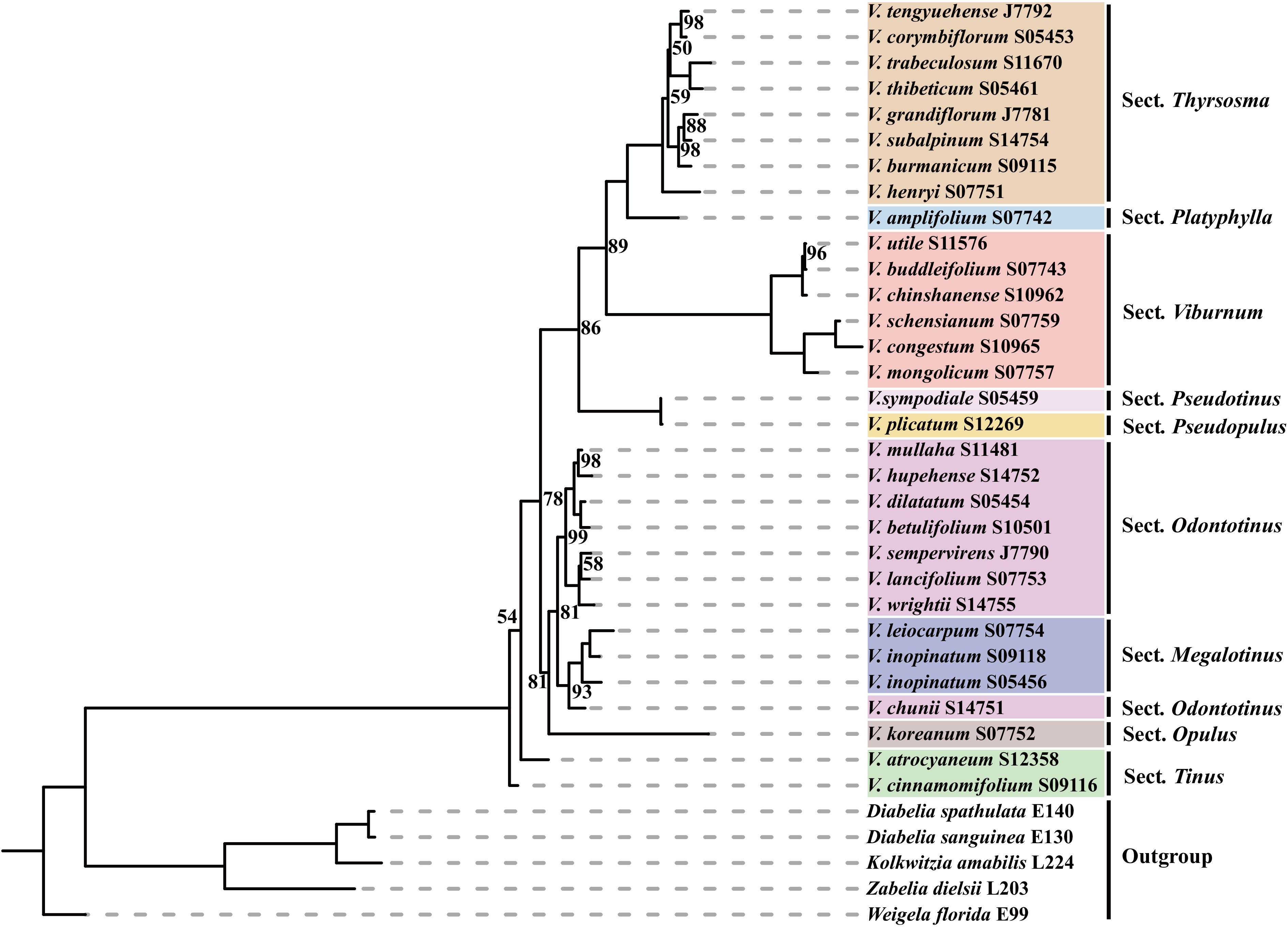
Figure 4.
IQtree constructed based on internal and external transcribed spacers (ITS and ETS) of nuclear genes.
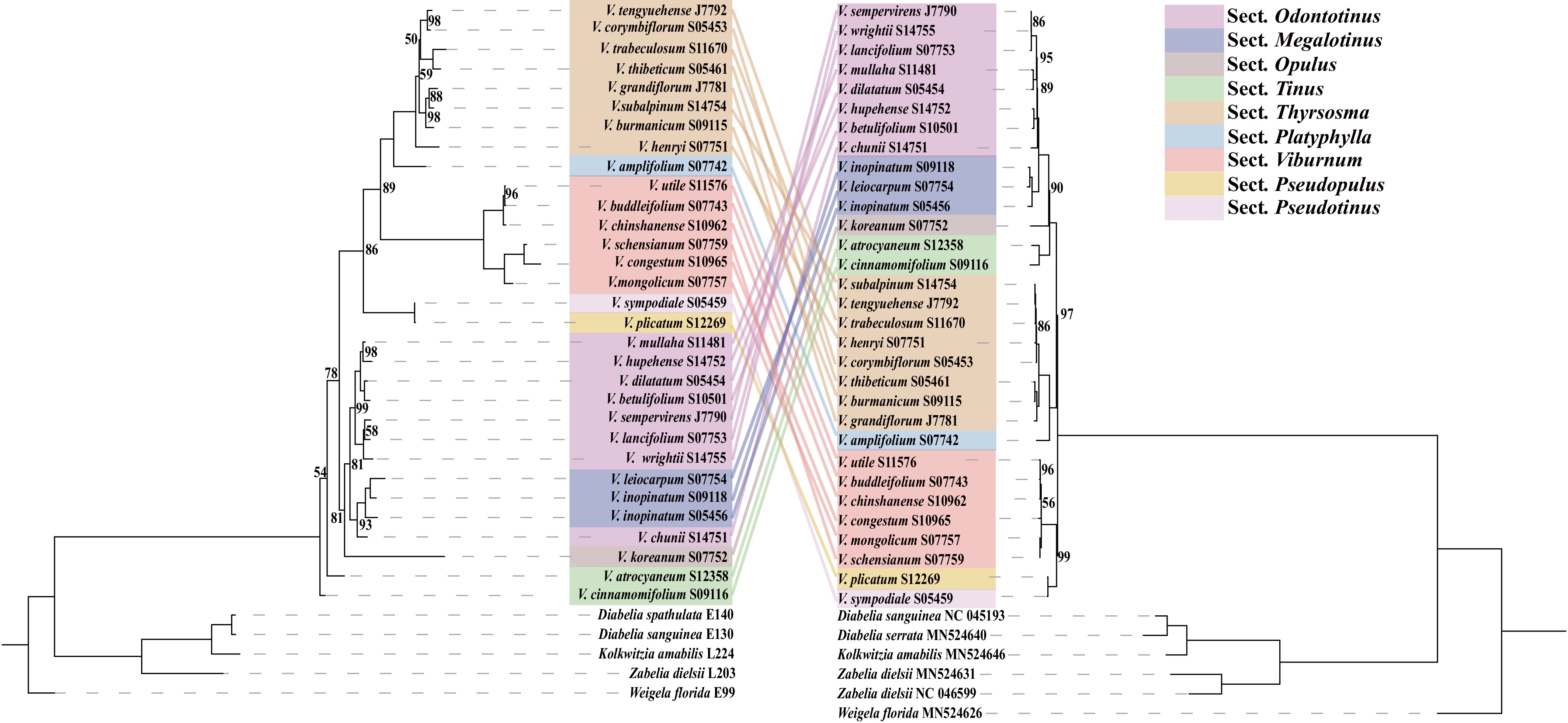
Figure 5.
Comparative phylogenetic trees of nuclear genes (left) and chloroplasts (right) are shown. The dotted lines connect homologous taxa on both phylogenetic trees.
Divergence time estimation
-
Divergence time estimation based on complete chloroplast genome data indicates that the crown group of Viburnum originated during the Eocene, with an initial divergence at 54.36 Ma (95% HPD: 31.80–76.09 Ma). The common ancestor of sections Opulus, Odontotinus, Megalotinus, and Tinus began to diversify around 40.03 Ma (95% HPD: 23.99–56.74 Ma) during the late Oligocene, followed by a rapid radiation that led to the establishment of the crown groups of these four sections. Subsequent divergence events occurred as follows: the crown age of Sect. Odontotinus was estimated at 23.62 Ma (95% HPD: 11.69–37.76 Ma); Sect. Megalotinus at 16.24 Ma (95% HPD: 5.10–31.17 Ma); Sect. Opulus at 17.07 Ma (95% HPD: 5.29–32.57 Ma); and Sect. Tinus at 24.25 Ma (95% HPD: 10.59–41.26 Ma). Sections Thyrsosma, Pseudopulus, and Platyphylla form a monophyletic group with Thyrsosma and Pseudopulus resolved as sister lineages. The crown age of sect. Thyrsosma was estimated at 27.55 Ma (95% HPD: 14.84–41.91 Ma), while sect. Pseudopulus exhibited a much more recent origin at 6.51 Ma (95% HPD: 1.13–16.67 Ma). In contrast, sect. Platyphylla diverged earlier, at 19.02 Ma (95% HPD: 6.02–35.57 Ma). The split between Sect. Viburnum and Sect. Pseudotinus occurred during the Oligocene, at 38.39 Ma (95% HPD: 20.28–58.86 Ma). The crown group of Sect. Viburnum was dated to 20.19 Ma (95% HPD: 7.39–35.62 Ma), while that of Sect. Pseudotinus was considerably younger, at 15.57 Ma (95% HPD: 3.26–34.54 Ma) (Fig. 6).
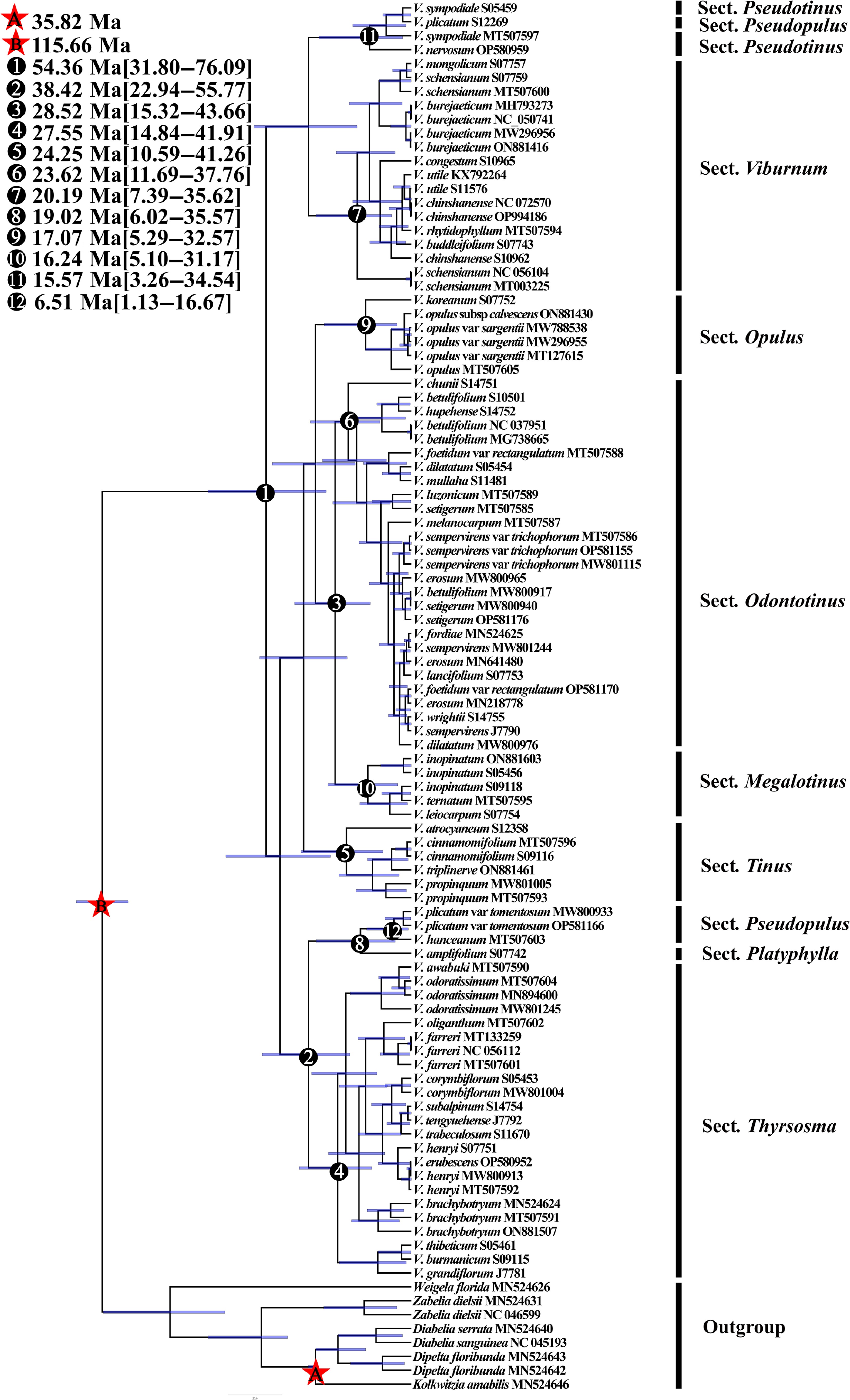
Figure 6.
BEAST divergence time analysis based on whole chloroplast genome construction. Calibration points are indicated by asterisks. Numbers 1–12 represent major divergence events in Viburnum.
Biogeography analysis of Viburnum
-
In our biogeographic analysis, we defined six major geographic regions (Supplementary Fig. S2) and identified the DEC+J model as the best-supported scenario based on comparative model fitting (Supplementary Table S2). East Asia represents the center of species richness with 35 recorded species, followed by South Asia (16 species), Southeast Asia and Europe (each with eight species), North America (six species), and South America (four species). East Asia also exhibits the highest phylogenetic diversity, with species distributed across all nine major clades of Viburnum, whereas South America shows the lowest, with only four species each representing distinct and phylogenetically divergent lineages. Notably, European species such as V. opulus and V. farreri belong to evolutionarily distant clades, a pattern also observed in South America, where V. nervosum and V. lancifolium are not closely related to each other, but instead show affinities to lineages outside the continent. This suggests that the South American species assemblage is largely the result of multiple independent colonization events rather than in situ diversification. Ancestral range reconstruction supports an East Asian origin for Viburnum, with subsequent dispersals into North America via the Bering land bridge during periods of climatic and geological connectivity. From North America, lineages further extended into South America, likely facilitated by late Cenozoic land connections and environmental opportunities.
Changes in species diversification rates
-
Our analyses indicate that the diversification rate of Chinese Viburnum has experienced a sustained increase over the past 54.36 (95% HPD: 31.80−76.09 Ma) million years, marking the onset of a major evolutionary radiation. Cross-referencing with paleotemperature reconstructions reveals that this initial divergence phase (54.36 Ma) coincided with a period of pronounced global climatic instability, characterized by substantial temperature fluctuations (Fig. 7). During this interval, the evolutionary rate of Viburnum initially decelerated before entering a phase of acceleration. In the subsequent Oligocene (33–25 Ma), an interval of relative climatic stability, the diversification rate continued to rise steadily. Notably, even as global temperatures declined during the Neogene and Quaternary, the diversification rate of Chinese Viburnum maintained an upward trajectory, suggesting that other factors—such as key morphological innovations, geographic expansion, or ecological opportunity—may have played a critical role in sustaining lineage accumulation despite cooling conditions.
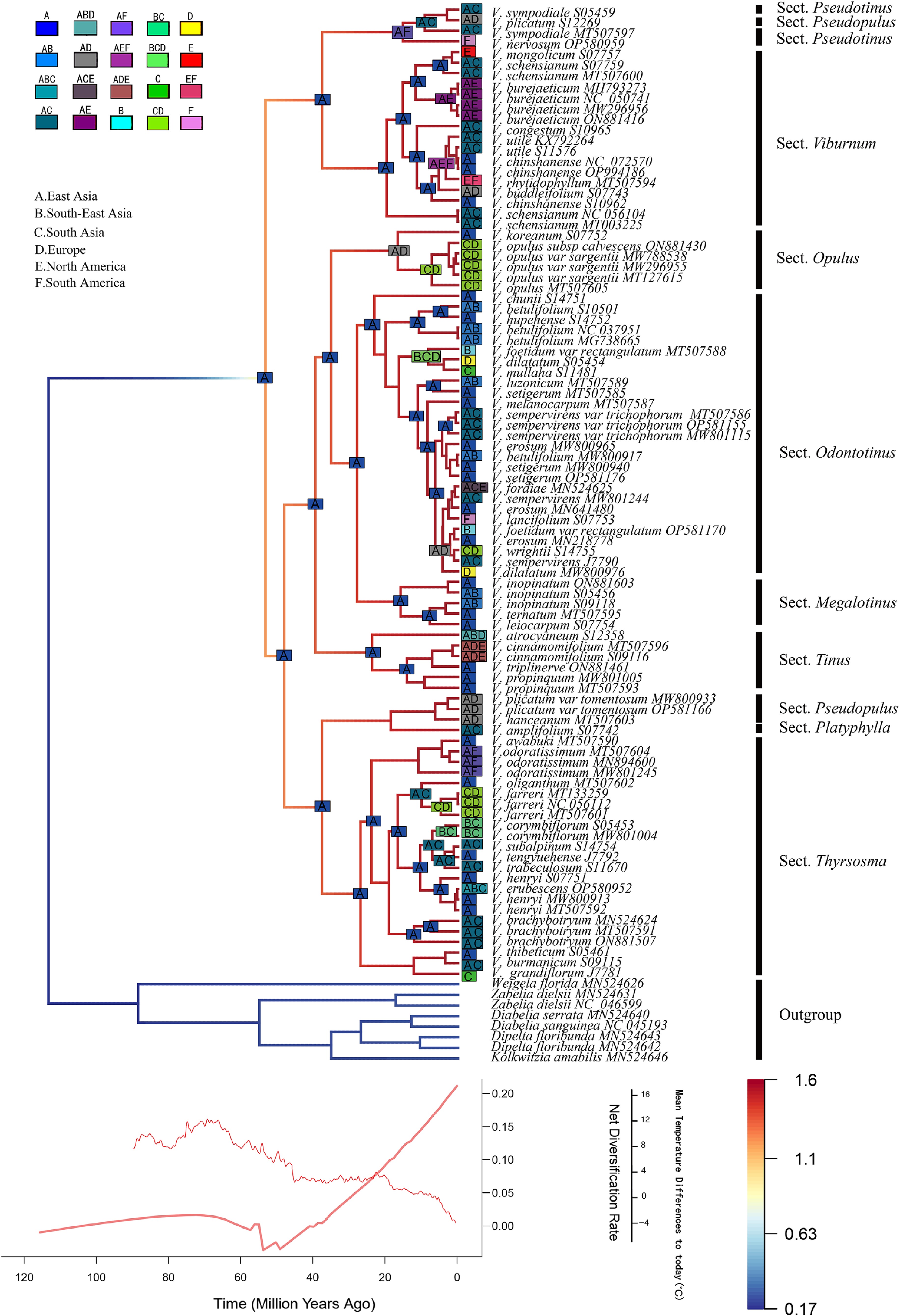
Figure 7.
DEC + J model used in BioGeoBEARS to infer ancestral distribution areas of Viburnum, (a) East Asia, (b) Southeast Asia, (c) South Asia, (d) Europe, (e) North America, and (f) South America. BAMM plot depicting trajectory of net diversification rate over time vs. global temperature change[39].
Ecological factor modeling results
-
Our analyses identified mean annual temperature (BIO1), temperature annual range (BIO7), and elevation as consistently significant environmental predictors of Viburnum species richness in both ordinary least squares (OLS) and spatial autoregressive (SAR) models (p < 0.01). Standardized regression coefficients for all variables were generally higher in the OLS model than in the SAR model, likely reflecting overestimated effect sizes due to unaccounted spatial autocorrelation.
Notably, diurnal temperature range (BIO2) showed a significant negative relationship with species richness in the OLS model (estimate = –0.097, p < 0.05) but was not significant in the SAR model (p = 0.135), indicating that its apparent effect may be confounded by spatial structure. Similarly, precipitation of the driest quarter (BIO17) was significant under OLS but non-significant under SAR, suggesting that its influence may also be attributable to spatial noise rather than direct ecological causation. In contrast, BIO1 and BIO7 retained stable, positive, and statistically significant effects in both models, supporting their roles as robust environmental drivers of species richness in Viburnum independent of spatial autocorrelation (Fig. 8).
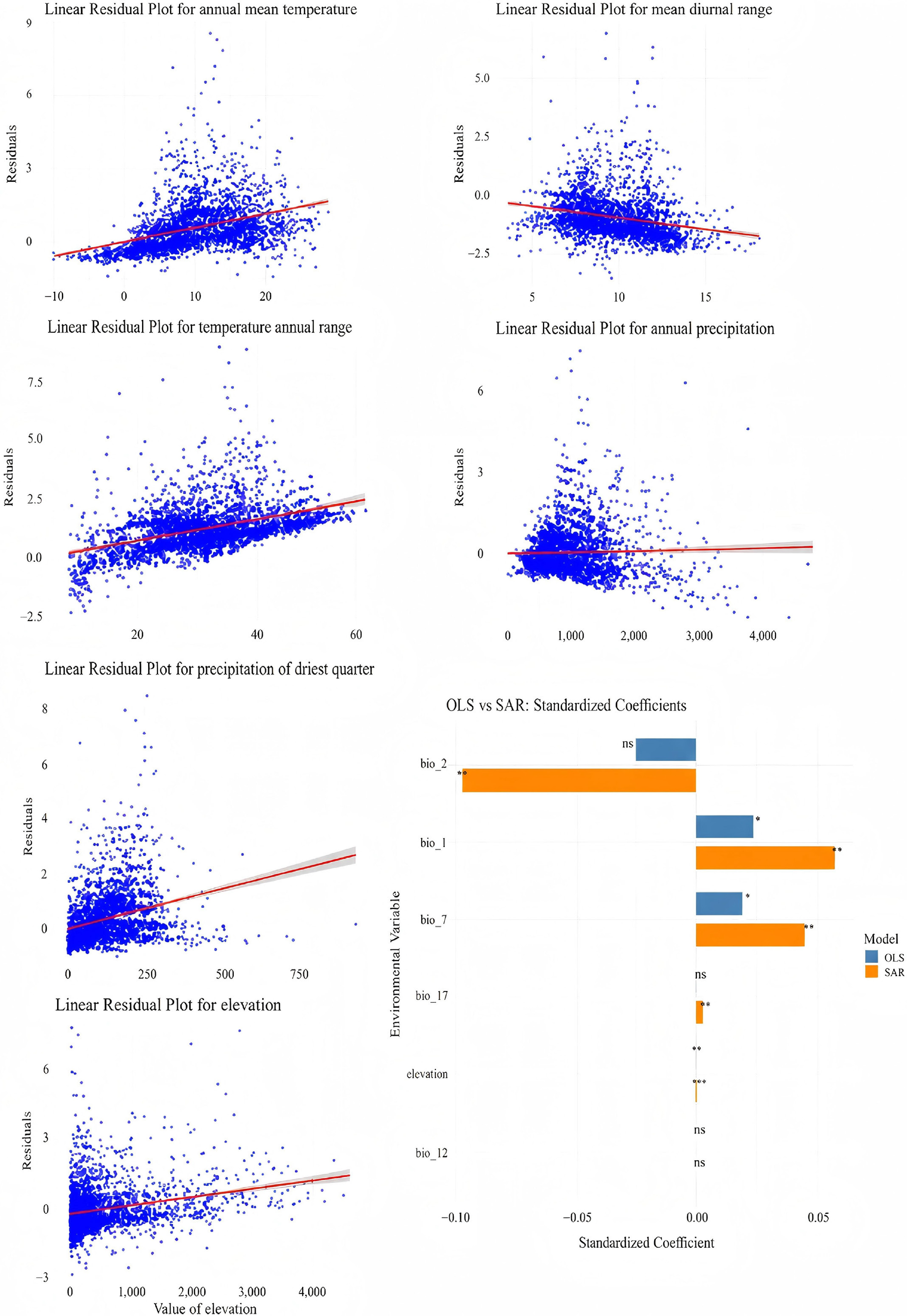
Figure 8.
Effects of different environmental factors on Viburnum species diversity. Bar graphs show the correlation between six variables; BIO1: mean annual air temperature; BIO2: mean diurnal temperature difference; BIO7: annual temperature difference; BIO12: annual precipitation; BIO17: precipitation in the driest season; and a topographic factor (altitude).
-
Phylogenetic trees reconstructed from the Viburnum chloroplast genome and the nuclear loci ITS and ETS exhibited notable topological incongruences, particularly in intergroup relationships (Fig. 5). In the chloroplast phylogeny, the toothed-leaf group (sect. Odontotinus) and the large-leaf group (sect. Megalotinus) formed a highly supported sister clade (BS = 100). This clade was sister to the cleft-leaf group (sect. Opulus; BS = 100), and the bulbous-kernel group (sect. Tinus) was sister to the combined clade ([Odontotinus + Megalotinus] + Opulus) with full support (BS = 100). In contrast, the nuclear phylogeny placed the large-leaf group (Megalotinus) within the toothed-leaf group (Odontotinus), indicating non-monophyly and conflicting with the chloroplast topology.
Such discordance between organellar and nuclear genealogies is commonly attributed to processes including incomplete lineage sorting (ILS), hybridization/introgression, horizontal gene transfer, or systematic error in tree estimation[44,45]. These factors may operate in isolation or concertedly[46], and both ILS and gene tree estimation inaccuracies can substantially affect phylogenetic reconstruction, particularly in rapidly radiating clades[47].
Statistically distinguishing between introgression and ILS—using model-based approaches such as PhyloNet or HyDe—requires multi-locus nuclear genomic data (e.g., from target enrichment or genome-wide SNPs). Our current dataset, comprising only the multi-copy nrDNA regions (ITS + ETS) and plastome sequences, falls short of this requirement. The phylogenetic signal from a single nrDNA locus can be confounded by concerted evolution and homogenization among paralogous copies, making it unsuitable for reliable inference under these sophisticated models. Therefore, we have refrained from applying such analyses here and explicitly acknowledge this limitation.
Although most sections of Viburnum were recovered as monophyletic in at least one genome, the widespread topological conflicts—coupled with short branch lengths across the species tree—suggest that Viburnum represents a complex evolutionary assemblage characterized by rapid diversification. Incomplete lineage sorting is likely a major contributor to the observed phylogenetic discordance, although the potential role of hybridization cannot be ruled out without further genomic evidence. A systematic evaluation of the relative contributions of ILS vs. introgression thus represents a critical goal for future research with more comprehensive nuclear genomic datasets.
Viburnum divergence time and biogeography
-
In this study, the stem group age of Viburnum was estimated using chloroplast genomic data and calibrated under a Bayesian framework, yielding an origin during the Eocene at 54.36 Ma (95% HPD: 31.80−76.09 Ma). This estimate aligns broadly with the Late Cretaceous stem age proposed by Spriggs et al.[16] (~46 Ma), which was inferred using contested Paleocene leaf fossils. Earlier work by Moore & Donoghue[48], employing fossil pollen assigned to Valvatotinus, suggested a younger crown age of 28 Ma (95% HPD: 17–40 Ma). In contrast, Spriggs et al.[16], using the same pollen fossil but with an expanded molecular dataset, inferred an older Eocene crown age of 55 Ma (95% HPD: 46–73 Ma). A consistent trend across studies is that increased taxonomic sampling and the incorporation of additional fossils have progressively pushed back estimated ages for major nodes within Viburnum, irrespective of calibration strategies. Importantly, regardless of the exact crown age, our results indicate that the majority of extant Viburnum species diverged after the Eocene, during more recent geological periods.
Biogeographic reconstructions indicate that Viburnum originated in Asia and subsequently dispersed to North America on multiple occasions, though back-dispersal from North America to Asia was not supported. These findings suggest a pattern of asymmetric colonization consistent with the Boreotropical hypothesis and the existence of Eocene–Oligocene land connections. The inferred dispersals are associated with several well-established intercontinental disjunctions—such as those between East Asia and Eastern North America, and between East Asia and Europe—mirroring biogeographic patterns observed in numerous other Northern Hemisphere plant clades[15]. Regions of high species richness and endemism, particularly in East and Southeast Asia, coincide with topographically complex mountain systems exhibiting high habitat heterogeneity. This heterogeneity, driven by pronounced altitudinal gradients and variability in hydrothermal conditions[49,50], has likely facilitated species coexistence, population isolation, and ecological diversification[16,51]. Moreover, these montane regions served as critical refugia during Pleistocene glaciations, mitigating extinction risks through microclimatic buffering and promoting allopatric speciation via habitat fragmentation and limited migration[52] In particular, the mountainous regions of Southern and Southwestern China, characterized by highly heterogeneous tropical and subtropical environments, represent not only the center of diversity for Viburnum but also a macroevolutionary cradle for the flora of Eastern Asia[50,52,53].
Our biogeographic analysis results, particularly the inferred timing of dispersal events across the Bering Strait, align with predictions from the land bridge hypothesis. Analysis indicates that Viburnum lineages dispersed multiple times from East Asia to North America during the Early to Middle Miocene. This period highly overlaps with the geologically confirmed existence window of the Bering Land Bridge, which emerged repeatedly throughout the Cenozoic and provided a viable corridor for temperate flora migration, especially during the Miocene[5,54]. Although our analysis cannot rule out alternative possibilities such as long-distance dispersal, multiple, temporally concentrated dispersal events are more consistent with migration via a persistently available land bridge corridor than with stochastic transoceanic dispersal[6]. We therefore propose that the land bridge hypothesis provides the most parsimonious and geologically coherent mechanistic explanation for the observed intercontinental disjunctions in Viburnum.
Factors influencing modern global patterns of Viburnum species diversity
-
The distributional diversity of Viburnum is shaped by a complex interplay of evolutionary history, ecological adaptation, and geographical processes[55]. Among these, rapid evolutionary radiation stands out as a predominant diversification mode, typically initiated by the emergence of new ecological opportunities or adaptive evolution in response to environmental shifts[56]. Our analyses further identify mean annual temperature, precipitation of the driest season, and elevation as the most significant environmental predictors underlying the spatial diversity patterns of the genus. These factors collectively delineate the ecological niche space of Viburnum, with most species exhibiting low water requirements and considerable drought tolerance—traits that may represent key adaptations to seasonal climates and may have been shaped by niche conservatism over deep evolutionary timescales[57]. Notably, lineage differentiation appears to be initiated at a mean annual temperature threshold of approximately 10 °C and accelerates under cooler conditions, pointing to a marked thermal sensitivity within the genus.
This pronounced temperature dependence suggests that global cooling during the Oligocene[39] served as a pivotal external driver (Fig. 8), likely triggering the rapid radiation event observed in Viburnum by creating new temperate zones and selective pressures[9,58]. As global climates deteriorated, cooling temperatures may have created widespread opportunities for adaptation and niche differentiation across the Northern Hemisphere, thereby promoting lineage splitting and ecological diversification.
Geographically, this evolutionary dynamism is reflected in the concentration of species within the 0–1,000 m elevation belt, which encompasses the three major centers of diversity: East Asia, Europe, and the Americas. These regions are characterized by topographically complex, low-elevation mountain systems that generate high habitat heterogeneity through varied slopes, aspects, and microclimates[59,60]. Such environmental mosaics are known to facilitate species coexistence by reducing competitive exclusion, promote ecological divergence through local adaptation, and ultimately drive allopatric speciation by subdividing populations across physical barriers[61,62]. The coincidence of high diversity with these geographically heterogeneous regions underscores the importance of landscape complexity in shaping the evolutionary trajectory of Viburnum, providing the stage upon which climatic drivers acted to generate its remarkable species richness.
-
Our phylogenomic analyses provide a robust framework for understanding the evolutionary history of Viburnum. We resolve most sections as monophyletic, with observed cytonuclear discordance largely attributable to incomplete lineage sorting rather than hybridization. Divergence time estimation indicates an Eocene origin (c. 54 Ma) and a sustained increase in diversification rates, coinciding with global cooling and Qinghai-Xizang Plateau uplift. Biogeographic reconstructions support an East Asian origin, with subsequent migrations to Europe and the Americas via the Bering and North Atlantic land bridges, consistent with the land-bridge hypothesis. The 'Out-of-Xizang' hypothesis further explains the colonization of high-elevation and cooler temperate regions. Centers of diversity in East Asia, Europe, and Eastern North America reflect both historical refugia and ongoing diversification in topographically complex regions. Importantly, we show that global cooling since the Oligocene and regional tectonic dynamics were critical drivers of Viburnum radiation. This study highlights the value of combining plastid-based phylogenetic, biogeographic, and ecological data to decipher the evolutionary mechanisms shaping plant diversity in response to environmental change.
-
The authors confirm their contributions to the paper as follows: conceptualization, project administration, funding acquisition: Wang HF; methodology: Liu PF, Yang WL; validation, supervision, writing − review and editing: Wang HF, Landis JB; resources, data curation: He YX; writing − original draft preparation, visualization: Di YX, Chen B. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the currentstudy are available from the corresponding author upon reasonable request.
-
This study was supported by grants from the National Natural Science Foundation of China (32270221), and the Natural Science Foundation of Hainan Province (421RC486 and 822QN314). We thank the staff of the Laboratory of Analytical Biology of the National Museum of Natural History for technical support and assistance. We thank the High Performance Computing Platform of the Department of Science and Technology of Hainan Province for supporting the data analysis process.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/tp-0026-0025.
-
Received 28 November 2025; Accepted 6 May 2026; Published online 15 June 2026
-
# Authors contributed equally: Yu-Xin Di, Bing Chen
- Supplementary Table S1 Information on plant species and source locations used in this study.
- Supplementary Table S2 Statistics for the six models considered in the nuclear‐based analysis in BioGeoBEARS.
- Supplementary Table S3 Overview of the main features of the chloroplast genome of Viburnum and its related outgroups.
- Supplementary Table S4 Overview of the main features of the Viburnum chloroplast genome.
- Supplementary Fig. S1 Previous studies on the phylogenetic relationships of Viburnum.
- Supplementary Fig. S2 Global distribution maps of Viburnum species constructed based on chloroplast divergence times for East Asia (A), Southeast Asia (B), South Asia (C), Europe (D), North America (E), and South America (F).
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Di YX, Chen B, Landis JB, Liu PF, Yang WL, et al. 2026. Phylogenomics and biogeography of Viburnum (Adoxaceae) based on plastid genomes reveal the roles of Qinghai-Xizang Plateau uplift and global cooling in its rapid radiation. Tropical Plants 5: e020 doi: 10.48130/tp-0026-0025 

Phylogenomics and biogeography of Viburnum (Adoxaceae) based on plastid genomes reveal the roles of Qinghai-Xizang Plateau uplift and global cooling in its rapid radiation
- Received: 28 November 2025
- Revised: 25 April 2026
- Accepted: 06 May 2026
- Published online: 15 June 2026
Abstract: Viburnum is a genus of over 160 species with a disjunct distribution across the Northern Hemisphere, making it an ideal system for studying biogeographic patterns and diversification processes. Despite previous phylogenetic work, the drivers underlying its rapid radiation and biogeographic history remain poorly understood. Here, we present a phylogenomic analysis based on complete chloroplast genomes supplemented by nuclear ribosomal ITS/ETS (Internal and External Transcribed Spacers of Nuclear Genes) sequences from 31 Chinese endemic species and 61 publicly available genomes, covering all major clades. We reconstructed phylogenetic relationships, estimated divergence times, inferred biogeographic history, and assessed diversification rates in the context of paleoclimate and tectonics. Our results strongly support the monophyly of most recognized sections, though with notable cytonuclear discordance likely due to incomplete lineage sorting. The crown age of Viburnum was estimated at 54.36 Ma (95% HPD: 31.80–76.09 Ma), with subsequent diversification linked to global cooling and the uplift of the Qinghai-Xizang Plateau. Biogeographic reconstructions support an East Asian origin, with multiple dispersals to North America via the Bering land bridge and later to South America. Niche modeling identified mean annual temperature and elevation as key factors shaping species richness. We conclude that the interplay of tectonic uplift and climate cooling during the Oligocene–Miocene transition promoted rapid radiation in Viburnum, supporting both the 'land-bridge' and 'Out-of-Xizang' hypotheses. This study demonstrates the utility of integrating plastid-based phylogenetic and ecological data to unravel the evolutionary history of temperate plant groups.
-
Key words:
- Phylogenomics /
- Biogeography /
- Divergence time /
- Ecological analysis /
- Environmental factors /
- Viburnum /
- Adoxaceae