









-
As sessile organisms, plants are constantly exposed to fluctuating environmental conditions that can pose significant challenges to their growth, development, and productivity. Environmental stresses are a major cause of global agricultural yield loss[1−3]. Through evolution, plants have developed intricate signaling networks capable of sensing stressors, processing endogenous and environmental information, and dynamically redirecting growth and defense outputs[4].
The phytohormone signaling network is central to plant adaptation to the environment, among which auxin is one of the most extensively studied multifunctional hormones. Auxin primarily forms concentration gradients within tissues through polar auxin transport, thereby regulating processes such as organogenesis, vascular differentiation, and environmental responses[5]. At the molecular level, the Aux/IAA gene family occupies a pivotal position in the auxin signaling pathway. The short-lived transcriptional repressor proteins encoded by this family bind to Auxin Response Factors (ARFs), repressing the expression of downstream genes under low auxin concentrations[6,7]. When auxin levels rise, the SCFTIR1/AFB receptor complex mediates the ubiquitination and degradation of Aux/IAA proteins, releasing ARFs to activate or inhibit target gene expression, thereby achieving precise regulation of plant growth and development[8].
Accumulating evidence has demonstrated that the Aux/IAA gene family members play crucial regulatory roles in shaping plant traits[9]. In tomato, the SlERF.J2–IAA23 module integrates hormonal signals to regulate hypocotyl elongation and plant height[10]. Beyond development, Aux/IAA genes are widely involved in abiotic stress responses. Under low-light treatment, auxin-related genes in watermelon exhibit significant genotype-specific expression[11]. In rice, OsIAA29 interacts with OsIAA21 and binds to OsARF17, regulating the auxin signaling cascade and altering seed development under high-temperature conditions[12]. In wheat, TaIAA15-1A enhances drought tolerance in Brachypodium by regulating ABA-related genes and strengthening antioxidant capacity[13]. In Arabidopsis, the loss of AtIAA5, AtIAA6, and AtIAA19 leads to reduced glucosinolate levels, rendering single and triple mutants more susceptible to drought stress compared to the wild type[14].
In terms of pathogen response, Shi et al. revealed that GhTIR1 specifically interacts with GhIAA14 in cotton, modulating plant defense against Verticillium dahliae through auxin perception and signaling[15]. A dominant mutation in OsIAA13 was shown to enhance resistance against Xanthomonas oryzae pv. oryzae (Xoo), by fine-tuning jasmonic acid and auxin-mediated defense mechanisms in rice[16]. Furthermore, Wang et al. demonstrated that the Puccinia striiformis effector PNPi suppresses host cell death by specifically targeting TaIAA14 in Nicotiana benthamiana, thereby promoting pathogen infection[17]. These findings collectively provide mechanistic insights into how pathogens manipulate host auxin signaling to subvert immunity, and offer a conceptual foundation for developing innovative and sustainable disease management strategies.
The advancement of genomic sequencing technologies and bioinformatics has facilitated systematic exploration of the Aux/IAA gene family across diverse plant species. Studies in model plants such as Arabidopsis thaliana (L.) with 29 members[18], Oryza sativa L. with 31[19], and Solanum lycopersicum L. with 25[20], along with research in non-model species, including Asparagus officinalis with 17[21], Hordeum vulgare L. with 36[22], and Solanum tuberosum L. with 26[23], have enabled comprehensive identification and annotation of Aux/IAA genes in these organisms.
Melon (Cucumis melo L.) is a major horticultural and economic crop of significant commercial value, cultivated worldwide. In 2023, global melon production reached 29.54 million tons, reflecting a substantial increase of over 8% compared to the past decade (Food and Agriculture Organization of the United Nations, 2025,
www.fao.org/faostat/en/#data/QCL ). The sequencing of the melon DHL92 genome by the Spanish Agricultural Genomics Research Center in 2012 laid the foundation for bioinformatics research in melon[24]. With continuous advances in bioinformatics, the DHL92 genome has been further refined to version V4.0[25]. Leveraging melon genomic data, researchers have conducted extensive transcriptome sequencing and analyses[26]. These rich datasets not only demonstrate the rapid progress in melon research, but also facilitate the work of a growing number of researchers. By deeply mining available transcriptome data, gene expression profiles can be efficiently obtained.Although the biological functions of Aux/IAA genes have been widely reported, studies on this gene family in melon remain limited. To address this gap, this study employed bioinformatic approaches to analyze the chromosomal localization, phylogenetic evolution, gene structure, and promoter sequences of Aux/IAA genes in melon. To investigate the expression patterns of Aux/IAA genes across different melon tissues and under various stresses, we systematically applied multiple biotic and abiotic stress treatments and performed transcriptome sequencing on the collected samples. In addition, we integrated publicly available transcriptome data to comprehensively analyze the expression profiles of the Aux/IAA gene family. The findings of this study establish a foundation for future research on the Aux/IAA gene family in melon, and provide potential candidate targets for resistance breeding in this crop.
-
All melon materials used in this study belong to Cucumis melo L. subsp. melo, with the exception of cultivar 'HMC1016' which is classified as Cucumis melo L. subsp. agrestis. These materials were provided by the Biological Breeding Laboratory of Xinjiang Academy of Agricultural Sciences (XAAS). Publicly available transcriptomic data was obtained from open databases.
Drought stress
-
Six melon cultivars ('HMC1013', 'HMC1014', 'HMC1015', 'HMC1016', 'HMC1017', and 'HMC1019') were subjected to drought stress under field conditions at the Turpan Experimental Base of the Xinjiang Academy of Agricultural Sciences (XAAS), China. At the early fruit-setting stage, plants in the drought treatment group (D–) were deprived of irrigation for 10 d, while the control group (W–) received normal watering. The 10th true leaf from both groups was collected, immediately flash-frozen in liquid nitrogen, and stored at –80 °C for subsequent RNA sequencing and expression profiling. Unless stated otherwise, all subsequent tissue samples in this section were processed identically.
High-temperature
-
High-temperature stress was applied to cultivar 'HMC-247' under controlled growth-chamber conditions. Seeds were surface-sterilized, germinated, and seedlings were transplanted into a soil mixture (nutrient soil : vermiculite : perlite = 3:1:1). Plants were grown at 28/18 °C (day/night) under a 16 h photoperiod (180 μmol·m−2·s−1). At 15 d after germination, seedlings were exposed to 42/30 °C (day/night) for 6 d. The first true leaves from stressed and control plants (five biological replicates per group) were collected.
Low-light stress
-
Using the same cultivar and growth conditions as in Section 2.1.2, seedlings at 15 d after germination were transferred to a light intensity of 40 μmol·m−2·s−1 for 56 h. Hypocotyls from stressed and control plants (three biological replicates per group) were harvested.
Downy mildew inoculation
-
Two melon cultivars with contrasting resistance, 'R' (resistant) and 'S' (susceptible), were inoculated with Pseudoperonospora cubensis (downy mildew). Plants were grown in a seedling greenhouse at the Hainan Sanya Breeding Station of XAAS. Seeds were disinfected, germinated, and sown in trays. At 15 d after germination, seedlings were spray-inoculated following Ling et al[27]. Leaf samples were collected at 0, 12, 24, 48, and 72 h post-inoculation (hpi).
Bacterial fruit blotch inoculation
-
The same cultivars ('R' and 'S') were inoculated with Acidovorax citrulli (bacterial fruit blotch) under identical growth conditions as those used for the downy mildew inoculation. The bacterial strain Pslb 38, provided by Prof. Tingchang Zhao (Institute of Plant Protection, Chinese Academy of Agricultural Sciences), was cultured as described by Wei et al[28]. Inoculation was performed according to Hopkins & Thompson[29], with three independent biological replicates. Leaf samples were collected at 0, 3, 6, 12, and 24 hpi.
Identification of the putative Aux/IAA gene family in C. melo and analysis of its physicochemical properties
-
The melon DHL92 v4.0 genome, protein sequences, and annotations were downloaded from the CuGenDBv2[30]. The Hidden Markov Model (HMM) for the Aux/IAA [AUX_IAA (PF02309)] gene family was downloaded from Pfam[31]. Putative Aux/IAA protein sequences were obtained using TBtools-II v2.336[32], to search the HMM against the proteome. Predicted amino acid sequences were examined using the NCBI CDD for the presence of characteristic domains. Theoretical isoelectric points (pIs), molecular weights, and grand average of hydropathicity (GRAVY) of the Aux/IAA proteins were predicted via the online software tools on the ExPASy website[33]. WoLF PSORT (
http://wolfpsort.hgc.jp ) was used to predict the subcellular location of the predicted Aux/IAA proteins.Chromosome locations, duplication events, and synthetic analysis of CmAux/IAAs
-
The chromosome location information of the CmAux/IAA's was obtained from the GFF annotation file of the genome, and diagrammed using the software TBtools[32]. CmAux/IAA duplication events were examined using MCScanX via TBtools[32]. Advanced Circos was employed to visualize segmental duplications. One Step MCScanX was employed to predict synteny between Aux/IAA genes in C. melo and those in C. sativus, S. lycopersicum, and A. thaliana, utilizing genome annotation and sequence files.
Aux/IAAs phylogenetic analysis and classification
-
A phylogenetic tree was constructed to elucidate the relationships and classification of Aux/IAA proteins among C. melo, A. thaliana, and C. sativus. Aux/IAA proteins were categorized based on their phylogenetic relationships, aligning them with corresponding members. The Arabidopsis thaliana and Chinese Long v3.0 (Cucumis sativus L.) Aux/IAA protein sequences were downloaded from the Arabidopsis Information Resource (TAIR,
www.arabidopsis.org ), and the CuGenDBv2[30], respectively. The 87 Aux/IAA protein sequences (29 Arabidopsis, 29 cucumber, and 29 melon) were aligned using the ClustalX2 software[34]. The aligned sequences were used to construct a maximum likelihood phylogenetic tree in MEGA 11[35], with 1,000 bootstrap replicates and default parameters.Analysis of conserved motifs, gene structure, and cis-element in CmAux/IAAs
-
The conserved motifs within the CmAux/IAA gene family were identified using the online tool MEME suite[36]. The intron-exon structures of these genes were visualized with the Gene Structure View (Advanced) module of TBtools[32]. For cis-acting regulatory element analysis, the 2.0 kb promoter sequences upstream of the CmAux/IAA genes were extracted and examined using the PlantCARE database[37]. The identified cis-elements were subsequently visualized using TBtools[32], statistically analyzed, and presented as a stacked bar chart using Microsoft Excel, with final composition and refinement performed in Adobe Illustrator.
Tissue-specific expression of CmAux/IAA family genes in C. melo
-
Tissue-specific expression patterns of the CmAux/IAA gene family were analyzed using publicly available RNA-seq datasets from NCBI BioProjects PRJDB6414 and PRJNA603204, obtained through the Cucurbit Genomics Database[30]. These datasets were generated from 42 distinct tissues and developmental stages of the Japanese muskmelon cultivar 'Earl's Favorite Harukei-3', covering vegetative, reproductive, and fruit development processes.
Additionally, this study utilized the melon cultivar 'Vedrantais', sampling the second true leaf as senescent tissue, and the 22nd true leaf as young tissue for transcriptome sequencing. FASTQ files that were too short, contained too many N bases, had a quality score lower than 20, or contained adapter sequences were considered to be of insufficient quality and were removed. The data was preprocessed and normalized to obtain TPM values according to standard transcriptome analysis protocols[38].
A heatmap was constructed using TBtools to visualize the expression patterns of CmAux/IAA genes across the different developmental tissues.
Analysis of CmAux/IAA genes expression profiles under abiotic and biotic stresses
-
Expression profiling of CmAux/IAA genes was conducted under multiple stress conditions. Under abiotic stress (salt, drought, high-temperature, and low light), the transcriptome data for salt and chilling stresses were obtained from NCBI BioProjects PRJNA987131[39] and PRJNA553119[40], respectively; the data for the remaining stresses were experimentally generated in this study. Under biotic stress, melon leaves were challenged with Pseudoperonospora cubensis (downy mildew), Podosphaera xanthii (powdery mildew), and Acidovorax citrulli (bacterial fruit blotch). Among these, the transcriptome data for powdery mildew infection was acquired from NCBI BioProjects PRJNA358655 and PRJNA358674[30], while the data for downy mildew and bacterial fruit blotch was produced in this study.
Differential expression analysis between the two groups was performed using DESeq2[41]. The p-value was adjusted using the Benjamini-Hochberg method. Differentially expressed genes (DEGs) were identified with the following thresholds: an adjusted p-value (padj) < 0.05, and an absolute log2 fold change ≥ 1.
-
After performing HMMER searches, 43 candidate genes were initially identified in the melon genome. Further validation through NCBI CDD confirmed the presence of the conserved Aux/IAA domain (PF02309) in polypeptides, resulting in the identification of 29 CmAux/IAA genes after excluding auxin response factors (ARFs) containing the Aux/IAA domain. All verified genes were systematically designated as CmAux/IAA1 to CmAux/IAA29 in ascending order, according to their chromosomal positions (Supplementary Fig. S1 and Supplementary Table S1). The peptide residue length, isoelectric point, and relative molecular weights (in kilodalton) varied greatly within the family, ranging from 127 aa/14.2 kDa (CmAux/IAA21), to 413 aa/45.4 kDa (CmAux/IAA11), and from 4.69 pI (CmAux/IAA21) and CmAux/IAA26), to 9.49 pI (CmAux/IAA18). Of the protein sequences analyzed, 16 were acidic (pI < 6.5), 11 were alkaline (pI > 7.5), and two were neutral (pI between 6.5 and 7.5). Proteins with instability index values exceeding 40 are considered unstable. CmAux/IAA2, CmAux/IAA4, CmAux/IAA18, CmAux/IAA24, and CmAux/IAA29 are classified as stable, whereas the remaining are categorized as unstable. The aliphatic index of 29 CmAux/IAA's ranged from 56.61 (CmAux/IAA1), to 89.21 (CmAux/IAA21), all less than 100, indicating that they were hydrophilic proteins. The grand average of hydropathicity (GRAVY) of CmAux/IAA proteins ranked from −0.898 (CmAux/IAA1), to −0.263 (CmAux/IAA21), indicating that CmAux/IAA proteins were hydrophilic. The majority of putative CmAux/IAA's were localized in the nucleus, while a few were found in the cytoplasm and extracellular.
Chromosomal distribution and synteny analysis of Aux/IAA genes in melon
-
To elucidate the chromosomal distribution of CmAux/IAA genes, we generated a chromosome map. The analysis revealed that CmAux/IAA genes were present on all chromosomes except chromosomes 7, 11, and 12 (Fig. 1). Chromosomes 5 and 4 contained the highest numbers of CmAux/IAA genes, with seven and six genes, respectively, followed by chromosomes 1 and 2 (four genes each). Chromosomes 3 and 6 harbored three and two genes, respectively, while chromosomes 8, 9, and 10 each contained only one CmAux/IAA gene. Collinearity analysis of the CmAux/IAA genes revealed 12 segmentally duplicated (SD) pairs (Fig. 1 and Supplementary Table S2), suggesting that segmental duplication may represent a primary mechanism for the expansion of this gene family[42]. We further identified orthologous Aux/IAA gene pairs between melon and three representative species: cucumber, tomato (Solanum lycopersicum), and Arabidopsis (Supplementary Fig. S2). A total of 26, 36, and 40 orthologous pairs were identified for cucumber, tomato, and Arabidopsis, respectively (Supplementary Table S3). The highest number of orthologs with Arabidopsis suggests that the Aux/IAA protein sequence and function are highly conserved, and that melon may share a closer evolutionary relationship with Arabidopsis.
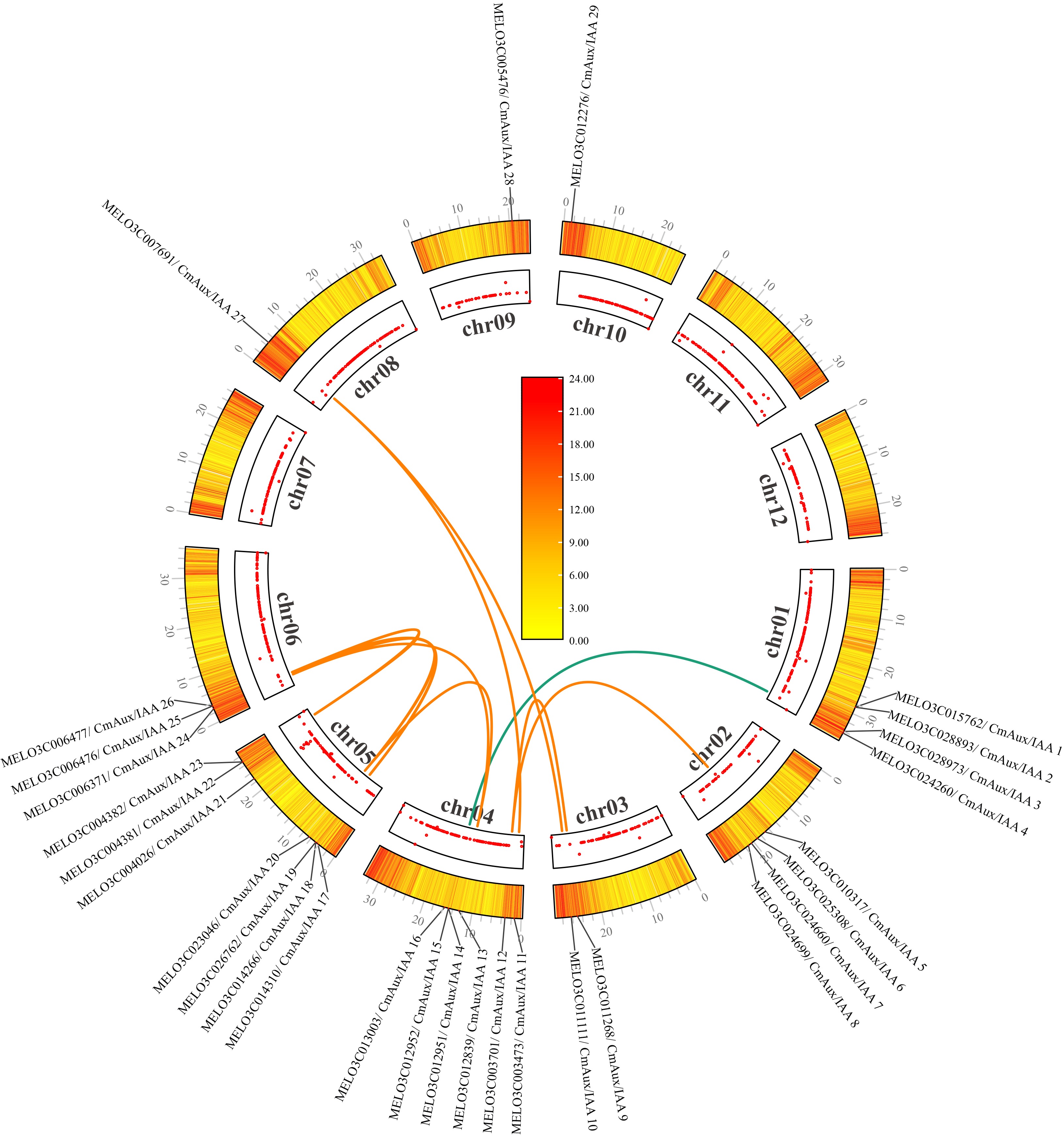
Figure 1.
Genomic distribution and collinearity analysis of AuxIAA genes in Cucumis melo.
Phylogenetic analysis of Aux/IAAs in A. thaliana, C. sativus and C. melo
-
To investigate the cross-species relationship of Aux/IAA family proteins, phylogenetic evolutionary trees of 87 Aux/IAA protein sequences (29 melon, 29 cucumber, and 29 Arabidopsis), are constructed using MEGA 11, and the phylogenetic trees was generated by the maximum likelihood method. According to the clustering with known Arabidopsis and cucumber orthologs, melon Aux/IAA proteins were divided into eight clades (Fig. 2). Clade I contains the fewest members, and sequences from cucumber and melon are absent from this clade. Clade II and Clade III contain the same total number of IAA proteins, with three and two CmAux/IAA proteins respectively. In Clade III, CmAux/IAA29 is the orthologous gene of IAA32/34. Therefore, CmAux/IAA29 may similarly function in differential cell elongation in the apical hook, a process governed in Arabidopsis by the antagonism between TMK1c and the CKRW1/WAV3 E3 ubiquitin ligase subfamily that controls IAA32/34 levels[43]. Clade IV consists of two subclades: Clade IVa (2 CmAux/IAAs), and Clade IVb (3 CmAux/IAAs). Within subclade IVb, CmAux/IAA20 and CmAux/IAA24 are identified as the likely melon orthologs of Arabidopsis IAA12/13. Notably, IAA12 plays a critical role in auxin-mediated apical-basal patterning during embryogenesis, and mutants lacking IAA12 function fail to develop a primary root meristem[44]. Clade V comprises five CmAux/IAA proteins, and Clade VI is partitioned into two subclades: VIa (2 CmAux/IAA's), and VIb (3 CmAux/IAA's). The Clade VIb proteins include the four highly sequence-similar Arabidopsis IAA1/2/3/4 proteins, where CmAux/IAA18 is grouped as their homolog. Clade VII consists of two CmAux/IAA proteins. Clade VIII incorporates the largest number of CmAux/IAAs, and is subdivided into VIIIa (four CmAux/IAA's), and VIIIb (five CmAux/IAA's). Phylogenetic analysis positions CmAux/IAA2 within VIIIb as the likely ortholog of Arabidopsis IAA16. Based on this homology and the critical role of the IAA1/2/3/4/7/16 gene set[45], it is plausible that CmAux/IAA2 and CmAux/IAA18 (homologs of Arabidopsis IAA1/2/3/4) are functionally analogous, potentially mediating low light sensitivity, and hypocotyl cell elongation.
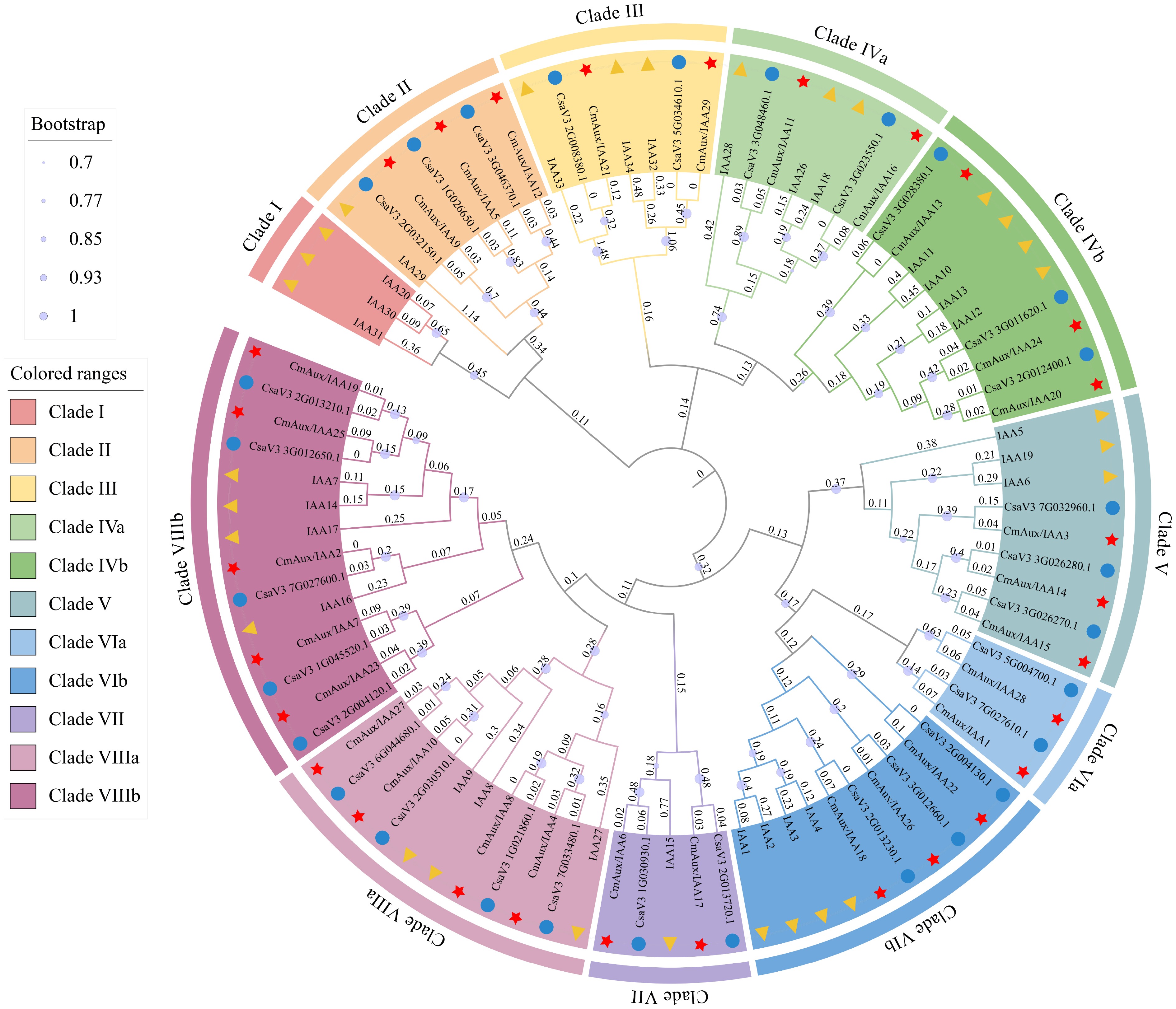
Figure 2.
Phylogenetic analysis of Aux/IAA proteins. The Aux/IAA amino acid sequences of A. thaliana, C. sativus, and C. melo were aligned via ClustalX 2.1, and the phylogenetic tree was constructed via the maximum likelihood method with 1,000 bootstrap replicates. The sequences were divided into eight clades. Different clades in the outer ring are distinguished by different colors, and different colors/shapes in the inner ring represent different species (yellow triangle is Arabidopsis, blue circle is cucumber, and red star is melon). This phylogenetic tree was generated by MEGA 11 (
www.megasoftware.net ), and iTOL online site (https://itol.embl.de ).Gene structure and conserved motif analysis of Aux/IAA genes in melon
-
A phylogenetic tree reconstructed with MEGA 11.0, comprising 29 melon Aux/IAA proteins, classified the genes into three distinct clades: Group I (21 members), Group II (4 members), and Group III (4 members) (Fig. 3a). Analysis of conserved motifs identified four characteristic motifs across the family (Fig. 3b, d). Notably, Motif 3 was universally conserved in all CmAux/IAA proteins, and Motif 2 was present in 93.10% of members. In contrast, Motif 1 was absent in nine members, and Motif 4 was missing in 12. Gene structure analysis revealed substantial variation in exon-intron architecture among CmAux/IAA genes, which ranged from a 7-exon/6-intron structure (CmAux/IAA10), to single-exon, intronless genes (CmAux/IAA15/21) (Fig. 3c). In terms of non-coding regions, 72.41% of members contained complete UTRs, though the number of UTRs varied significantly—from a maximum of six in CmAux/IAA10, to a complete absence in genes such as CmAux/IAA21 and 23, suggesting distinctive structural configurations. The number of CDS regions also differed considerably, with CmAux/IAA11 possessing the highest count (6), whereas CmAux/IAA15 and 21 contained only one. Collectively, these results demonstrate a remarkable level of structural diversification within the CmAux/IAA gene family.
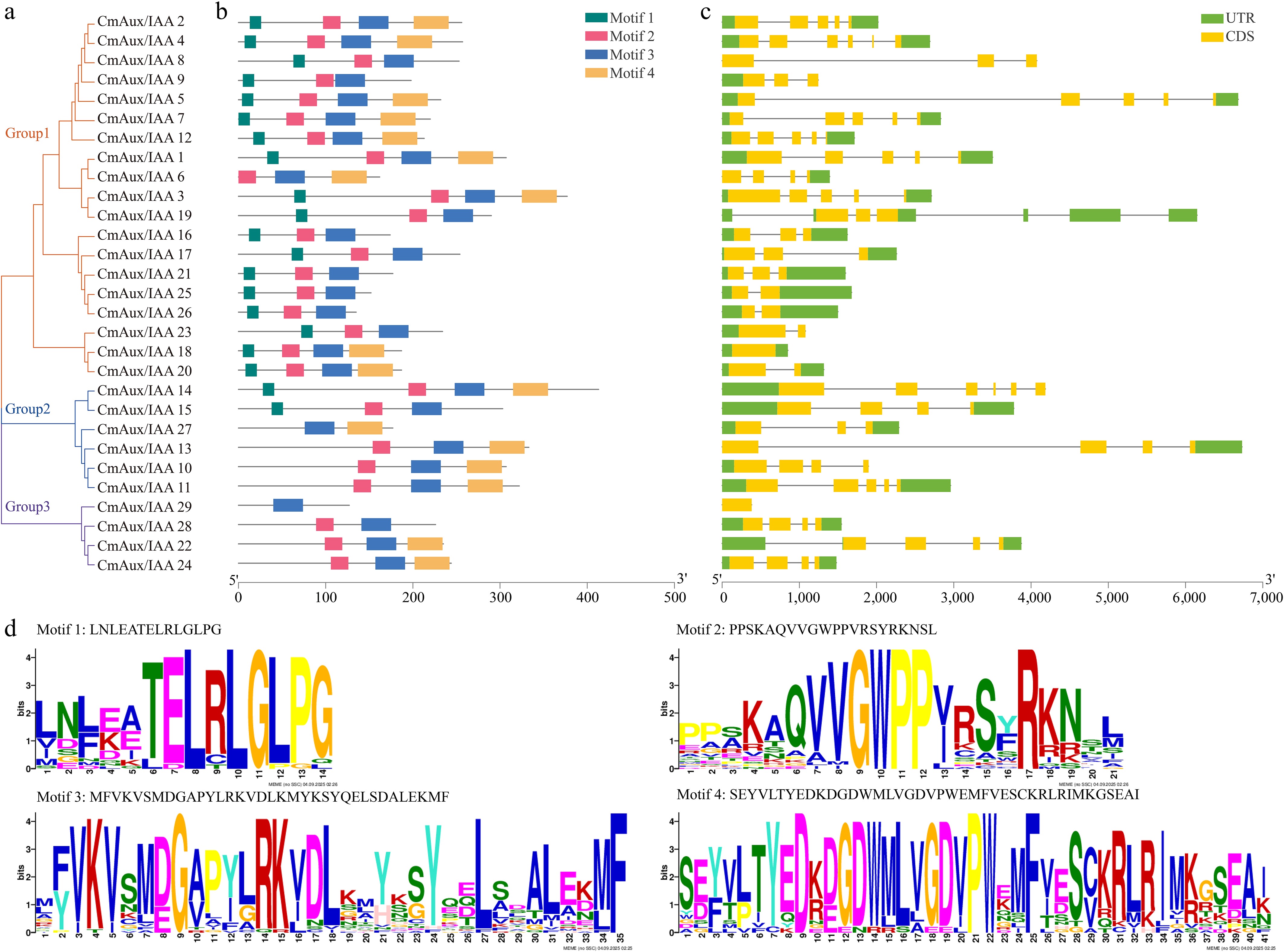
Figure 3.
Phylogenetic relationships, gene structure, and motif distribution of CmAux/IAA > gene family. (a) Phylogenetic tree of CmAux/IAA protein with 1,000 replicates per node. (b) The distribution of conserved motifs of CmAux/IAA proteins by the MEME. Different color boxes represent four conserved motifs. (c) Exon-intron structures of CmAux/IAA genes. Green rectangles, yellow rectangles, and black lines indicate UTR (non-coding region), CDS (coding sequence or exons), introns, respectively. (d) Amino acids' sequence logos for the four motifs.
Cis-acting elements in the promoters of CmAux/IAA genes
-
To elucidate the potential regulatory mechanisms of CmAux/IAA gene expression, a 2000-bp promoter region of each CmAux/IAA gene was analyzed using PlantCARE. Comprehensive analysis revealed a diverse array of cis-elements implicated in various biological processes (Fig. 4, Supplementary Fig. S3, and Supplementary Table S4). Notably, the promoter sequences contained an abundance of light-responsive elements, including G-box, Box 4, TCT-motif, and GT1-motif. We also detected 336 short function elements, a finding consistent with the canonical characterization of Aux/IAA proteins as 'short-lived nuclear proteins'[46]. The search further revealed hormone-responsive elements, such as ABRE, CGTCA-motif, TCA-element, and TGACG-motifs, which are known to mediate responses to abscisic acid, methyl jasmonate, and salicylic acid. Among the elements associated with environmental stress were ARE, TC-rich repeats, MBS and LTR, suggesting putative roles in anaerobic stress response, defense mechanisms, low-temperature adaptation, and drought inducibility. The promoters also harbored seed- and endosperm-specific regulatory elements, along with elements related to meristem expression, pointing to their involvement in developmental processes. Finally, the presence of cis-elements involved in circadian control (e.g., circadian), and cell cycle regulation (e.g., MSA-like), indicates a potential role for Aux/IAA genes in modulating the plant's rhythmic physiological processes.
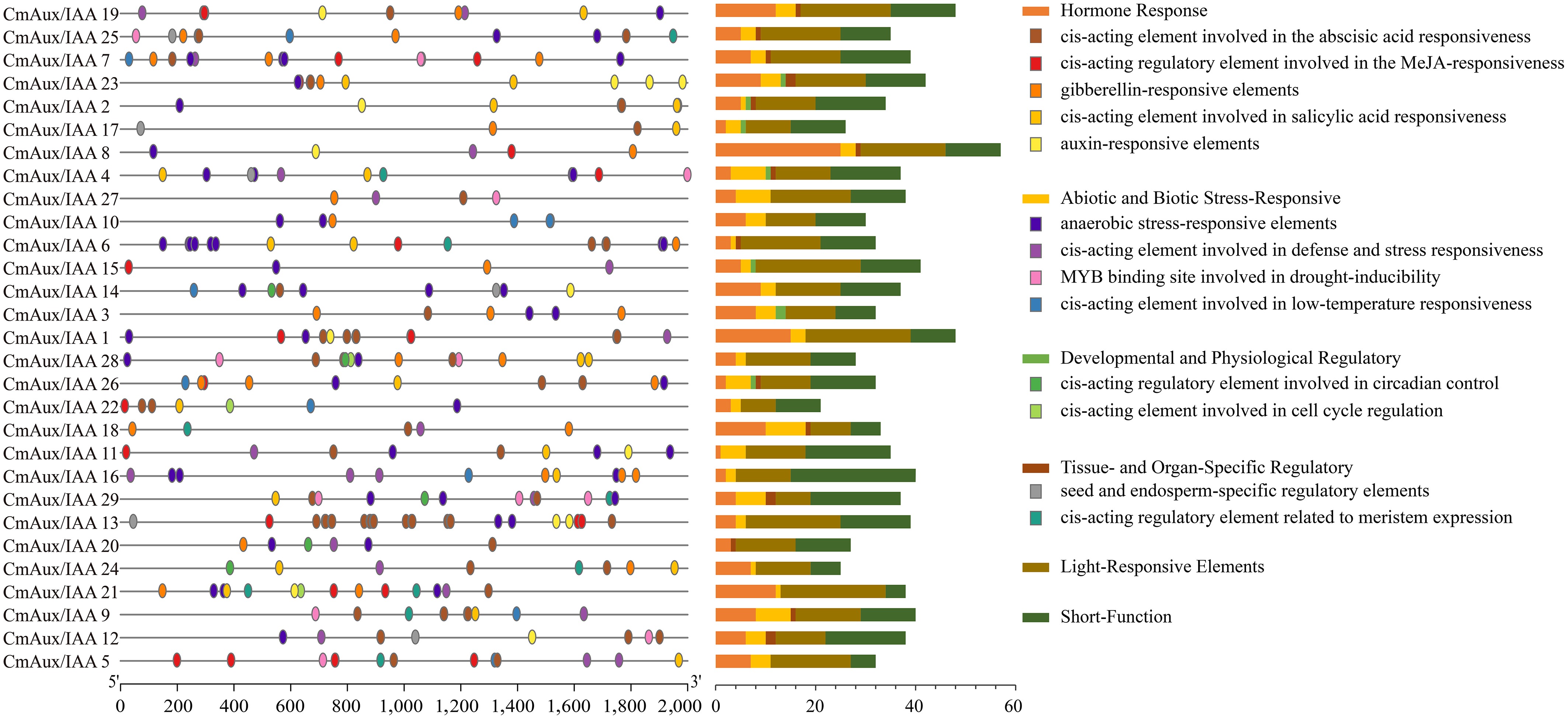
Figure 4.
Cis-acting element of the Aux/IAA gene family in melon. Elements are represented by differently colored ovals on the left, with each type possessing a distinct biological role. The number of elements within each major category is presented in the stacked bar chart on the right.
CmAux/IAA gene expression patterns in melon growth and development
-
We characterized the expression patterns of the CmAux/IAA gene family in the Japanese muskmelon 'Earl's Favorite Harukei-3' across a set of 42 distinct tissues and developmental stages, using publicly available RNA-seq datasets (Fig. 5 and Supplementary Table S5). Our analysis revealed pronounced tissue-specific expression profiles. Specifically, CmAux/IAA6, 7, 8, 10, 18, and 20 were predominantly expressed in the epicarp, while a distinct set of genes—CmAux/IAA1, 9, 12, 16, 19, 26, 28, and 29—showed higher expression in flowers than in the epicarp. Another gene subset, including CmAux/IAA3, 5, 15, and 22, exhibited peak expression in flesh rather than leaves, whereas CmAux/IAA17 was most highly expressed in leaves. Notably, CmAux/IAA17 displayed strict leaf-specific expression, with transcripts detected exclusively in cotyledons and leaves, and absent from stems, roots, flowers, and fruit tissues. In contrast, CmAux/IAA14 and 15 were not expressed in leaves (Fig. 5, Supplementary Fig. S4, Supplementary Tables S5 and S6). Meanwhile, CmAux/IAA21 showed high expression restricted to tendrils, with no expression detected in the epicarp or stem. Notably, CmAux/IAA2, the homolog of Arabidopsis IAA16, exhibited the highest expression level in the hypocotyl, suggesting its potential role in mediating hypocotyl cell elongation, and low light sensitivity[45].
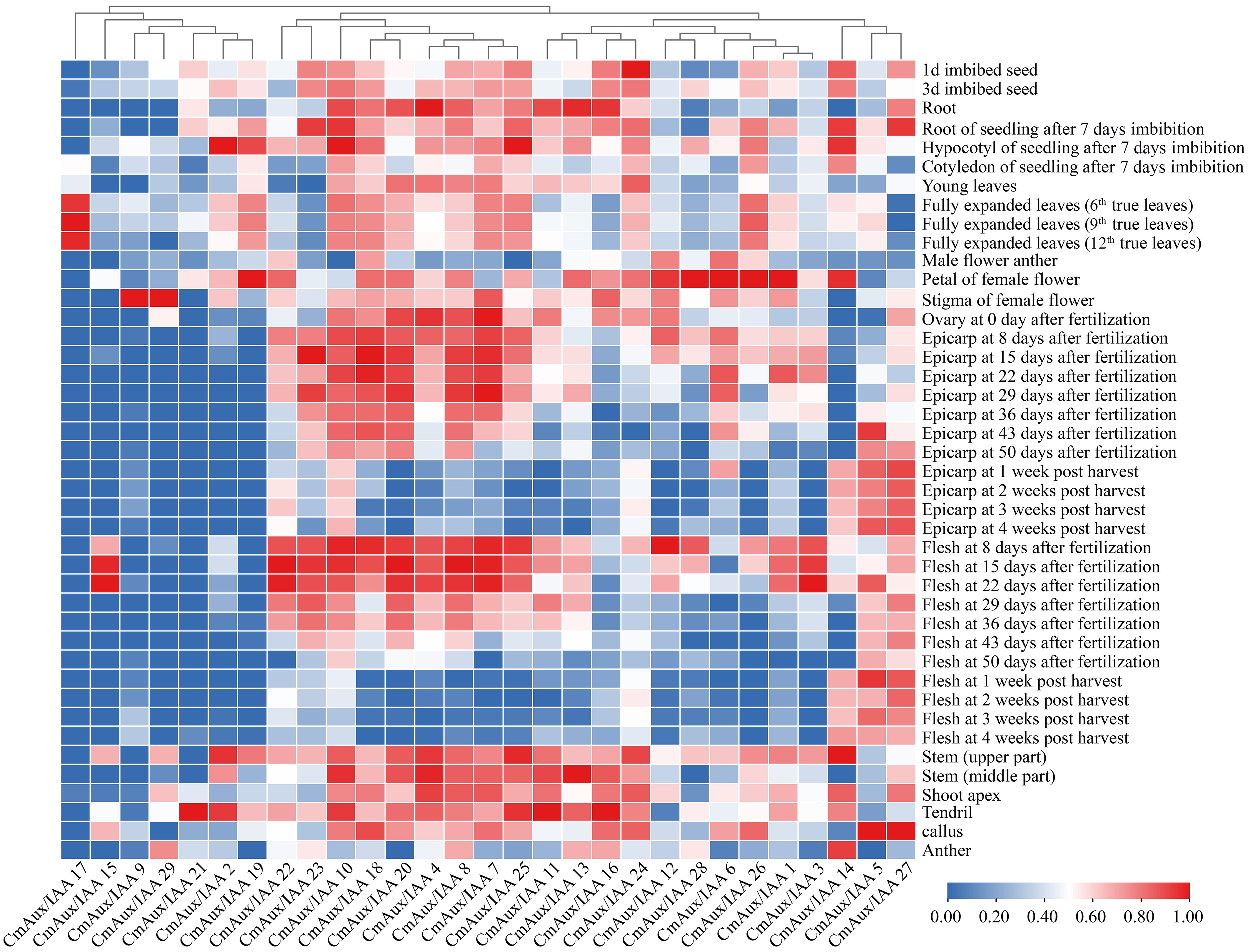
Figure 5.
Tissue-specific expression patterns of the Aux/IAA gene family in melon. Integration of RNA-seq data from 27 and 15 developmental tissues (NCBI BioProjects PRJDB6414 and PRJNA603204). The heatmap was generated using TBtools based on log2-transformed fluorescence intensity values. Expression values for each gene (columns) across different samples (rows) were standardized by Z-score normalization. Genes were clustered using Pearson correlation coefficients. The color gradient from blue to red represents relative expression levels from low (0.00) to high (1.00).
From a developmental timing perspective, most genes showed higher expression levels in the epicarp and flesh at post-pollination stages, compared to post-harvest stages (Fig. 5 and Supplementary Table S5). In contrast, CmAux/IAA14 exhibited an opposite expression pattern. CmAux/IAA27 maintained consistently high expression in both post-pollination and post-harvest fruit tissues, with no significant changes observed. A similar trend was observed in leaves: while most genes were more highly expressed in young leaves than in senescent leaves, CmAux/IAA16 and 27 showed higher expression in senescent leaves (Supplementary Fig. S4, Supplementary Table S6). Notably, CmAux/IAA14 and 15 were nearly undetectable in leaf tissues across all developmental stages. Collectively, these findings underscore the diverse and tissue-specific regulatory roles of CmAux/IAA genes during melon development.
Expression patterns of CmAux/IAA gene in melon under abiotic stresses
-
Promoter analysis of the CmAux/IAA gene family uncovered multiple cis-acting elements associated with hormones and stress, suggesting their potential roles in development and stress responses. To further investigate their stress responsiveness, we examined the expression profiles of CmAux/IAA genes in melon under salt, drought, chilling, high-temperature, and low light stress. The transcriptome data for salt and chilling stress was obtained from public databases (NCBI BioProjects: PRJNA987131 and PRJNA553119), while the data for the other abiotic stress conditions was generated in this study.
Utilizing RNA-Seq data from the melon cultivar 'Longqing' under salt stress[39], expression dynamics of CmAux/IAA genes at 0, 6, 12, 24, and 48 h post-treatment was analyzed (Fig. 6a, Supplementary Table S7). The results showed that CmAux/IAA29 was virtually undetectable in roots, corroborating previous findings in the 'Harukei-3' cultivar (Fig. 5, Supplementary Table S5). Under salt stress, expression of CmAux/IAA 1, 2, 6, and 11 was up-regulated at 6 h post-treatment. By 12 h, 12 genes (CmAux/IAA 1, 2, 5, 6, 9, 10, 11, 18, 19, 22, 25, and 26) showed increased expression. At the 24-h time point, up-regulation was observed only for CmAux/IAA 9 and 18. After 48 h of treatment, most CmAux/IAA genes were down-regulated, with CmAux/IAA 9 being the sole gene that remained up-regulated. Based on the above expression profiling data, CmAux/IAA1, 2, 6, 9, 11, and 18 were up-regulated at least two time points and were therefore selected as candidate genes for subsequent studies on salt stress response.
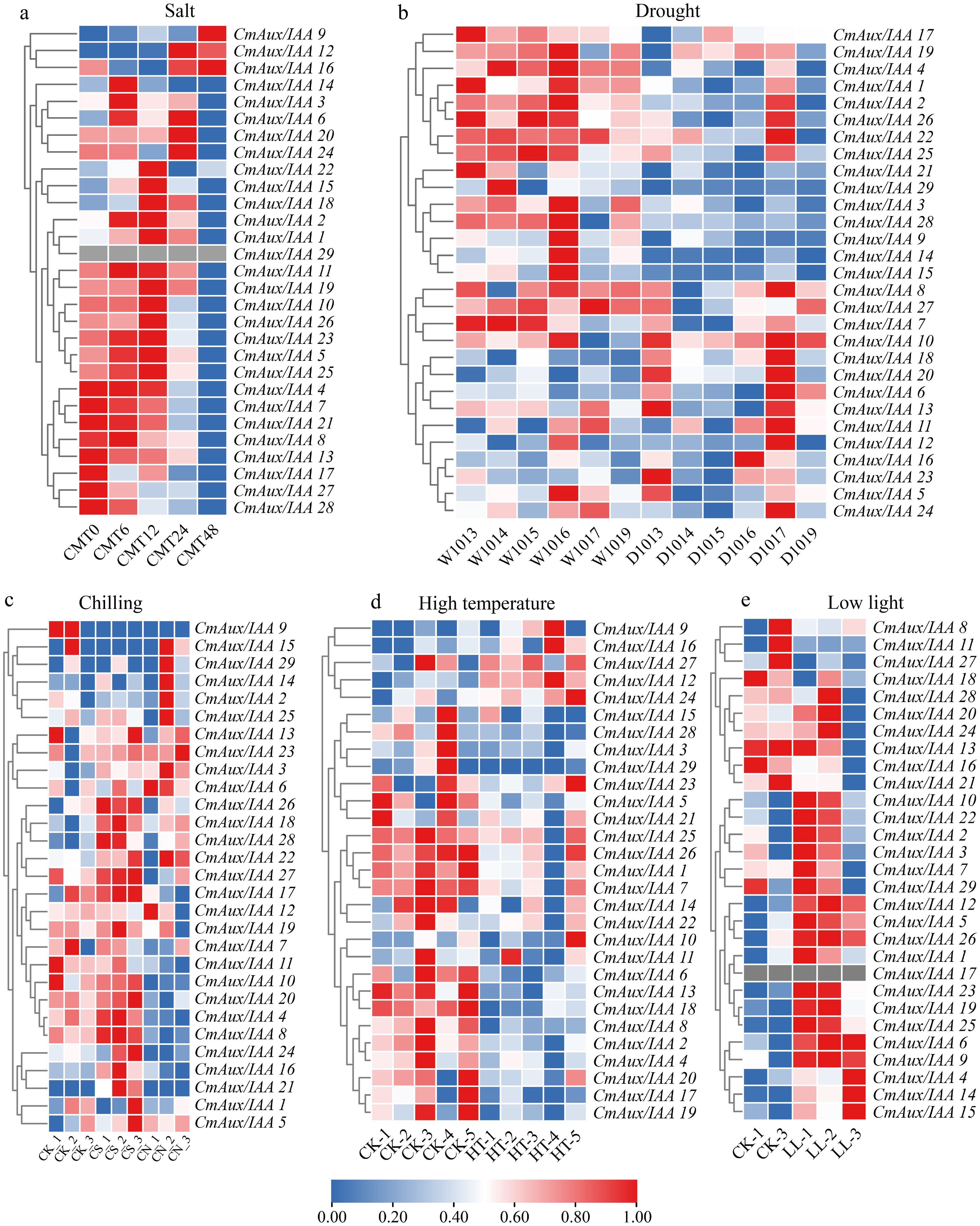
Figure 6.
Expression profiles of CmAux/IAA genes under abiotic stresses. (a) Root expression in cultivar 'Longqing' under salt stress at indicated time points (0, 6, 12, 24, 48 h). (b) Response to drought stress across six distinct melon varieties. (c) Response to chilling stress (CS) and the combined treatment of chilling with the nitric oxide donor sodium nitroprusside (CN) in seedlings. (d) Response to high-temperature stress. (e) Response to low-light stress. Data for (a) and (c) were obtained from public databases (NCBI BioProject: PRJNA987131 and PRJNA553119, respectively); data for (b), (d), and (e) were generated in this study. For the salt stress condition, expression values are reported as the median of three biological replicates. The heatmap was generated in TBtools based on log2-transformed fluorescence intensity values, with Z-score normalization applied to each gene (rows) across all samples (columns). Hierarchical clustering was performed using Pearson correlation coefficients. A blue-to-red color gradient indicates relative expression levels from low (0.00) to high (1.00).
We conducted natural drought treatment on six melon varieties and obtained RNA-Seq data to investigate the expression patterns of CmAux/IAA gene family members in response to drought stress (Fig. 6b, Supplementary Table S8). Drought stress analysis revealed that CmAux/IAA11 was the core responsive gene, significantly up-regulated in all six melon varieties. Additionally, CmAux/IAA18 and CmAux/IAA20 were broadly up-regulated in five varieties ('HMC1013', 'HMC1014', 'HMC1016', 'HMC1017', and 'HMC1019'), and CmAux/IAA10, CmAux/IAA16, and CmAux/IAA23 showed increased expression in four varieties.
We analyzed the expression patterns of CmAux/IAA genes in melon seedlings under chilling stress, and under combined treatment with the nitric oxide (NO) donor sodium nitroprusside (chilling + SNP, CN), using RNA-seq data from the NCBI public database (Fig. 6c, Supplementary Table S9)[40]. The results showed that chilling stress induced the up-regulation of CmAux/IAA26 and 28. Previous studies have reported that exogenous NO application enhances plant tolerance to chilling stress. At the gene expression level, under the combined CN treatment, CmAux/IAA3 was expressed at higher levels compared to the control (CK), whereas CmAux/IAA26 and 28 showed no significant difference in expression. Moreover, CmAux/IAA4 and 11, which were down-regulated in the CN group, displayed no significant expression changes under chilling stress alone. Based on these results, we speculate that CmAux/IAA26 and 28 may play a key role in the response to chilling injury.
The expression of CmAux/IAA genes under high-temperature and low light conditions was further analyzed (Fig. 6d, e, and Supplementary Tables S10, S11). Expression analysis revealed that only CmAux/IAA12 was up-regulated in response to high-temperature stress, while a set of members—including CmAux/IAA1, 2, 4, 6, 7, 8, 17, 18, 25, 26, and 28 were consistently down-regulated. Under low light conditions, CmAux/IAA17 expression was undetectable in hypocotyl tissues, consistent with the aforementioned findings in the 'Harukei-3' cultivar (Fig. 5, Supplementary Table S5). In addition, CmAux/IAA2 and 25 exhibited the highest expression level among all CmAux/IAA genes in the control group, supporting their potential key role in hypocotyl development. However, under low light treatment, CmAux/IAA2 and 25 expression remained largely unchanged. Notably, the expression of CmAux/IAA14 increased significantly, suggesting it plays an important role in adaptation to low light.
Expression patterns of CmAux/IAA gene in melon under biotic stresses
-
In addition to abiotic stresses, plants in natural environments face threats from pathogenic infections (including bacteria, fungi, and viruses). As a key phytohormone, auxin not only interacts with other endogenous hormones but also responds to various biotic stresses. Previous studies have reported that multiple pathogens have evolved strategies to disrupt Aux/IAA-mediated signaling. To investigate whether CmAux/IAA genes in melon are targeted by such strategies and how they respond to pathogen infection, we examined the expression profiles of CmAux/IAA genes in melon leaves exposed to two fungal and one bacterial pathogen, including Pseudoperonospora cubensis (downy mildew), Podosphaera xanthii (powdery mildew), and Acidovorax citrulli (bacterial fruit blotch). The transcriptome data for powdery mildew infection was obtained from public databases (NCBI BioProjects: PRJNA358655 and PRJNA358674), while the data for the other pathogens was generated in this study.
We inoculated both the resistant cultivar 'R' and the susceptible cultivar 'S' with downy mildew, and collected samples at 0, 12, 24, 48, and 72 hpi for transcriptome sequencing (Fig. 7, Supplementary Table S12). The results revealed that several genes exhibited consistent expression patterns in both 'R' and 'S' genotypes. For example, CmAux/IAA16 was consistently up-regulated from 12 to 72 hpi, while CmAux/IAA28 showed up-regulated expression at 12 hpi, and both CmAux/IAA11 and CmAux/IAA22 were up-regulated at 72 hpi. In contrast, some genes displayed genotype-specific expression patterns. At 12 hpi, CmAux/IAA9 was up-regulated exclusively in the 'R' genotype, whereas CmAux/IAA20 was specifically induced only in 'S'. Similarly, at 72 hpi, CmAux/IAA1 and CmAux/IAA2 were up-regulated specifically in 'R', but showed no significant expression changes in 'S'.
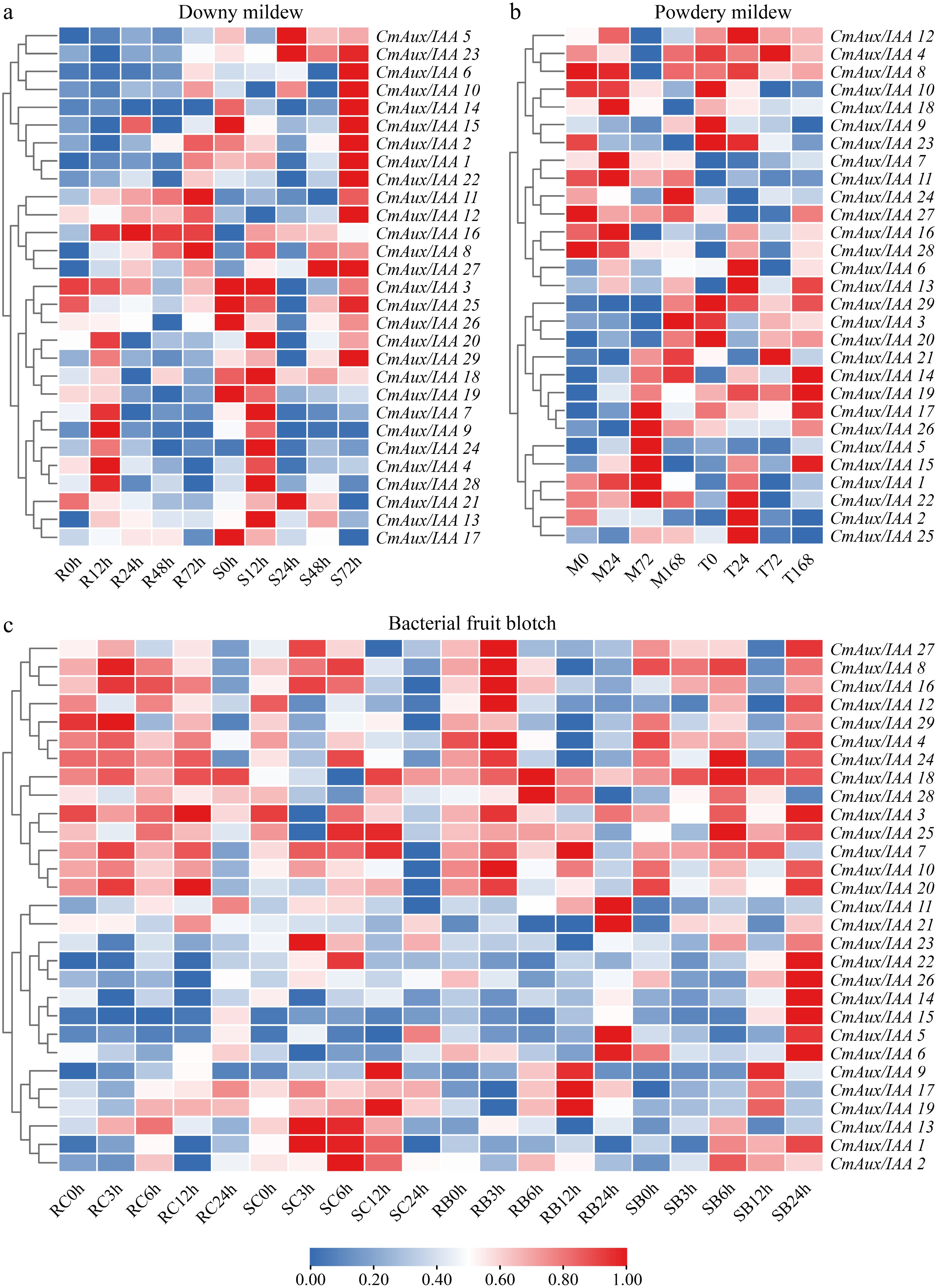
Figure 7.
Disease-responsive expression patterns of Aux/IAA genes in melon. (a) Expression profiles in downy mildew-resistant 'R' and -susceptible 'S' at 0, 12, 24, 48, and 72 h post-inoculation (hpi). (b) Expression dynamics in cultivars 'MR-1' (M) and 'Top Mark' (T) at 0, 24, 72, and 168 hpi with powdery mildew. (c) Expression profiles in bacterial fruit blotch-resistant 'R' and -susceptible 'S' at 0, 3, 6, 12, and 24 hpi. Data for (b) were obtained from public databases (BioProject: PRJNA358655 and PRJNA358674); data for (a) and (c) were generated in this study. Expression values for downy mildew and bacterial fruit blotch infections represent median values of three biological replicates. The heatmap was generated using TBtools based on log2-transformed fluorescence intensity values. The heatmap illustrates Z-score normalized expression values for each gene (rows) across different samples (columns). Hierarchical clustering was performed using Pearson correlation coefficients. A blue-to-red color gradient indicates expression levels from low (0.00) to high (1.00).
Utilizing RNA-Seq data obtained from the NCBI, we conducted expression profiling of CmAux/IAA genes in two melon cultivars, 'MR-1' (resistant cultivar), and 'Top Mark' (susceptible cultivar), at 0, 24, 72, and 168 h post-inoculation with powdery mildew. Our results (Fig. 7, Supplementary Table S13) revealed substantial differences in CmAux/IAA expression patterns between the two cultivars under powdery mildew stress. In 'MR-1', CmAux/IAA genes exhibited a phased induction pattern: CmAux/IAA5, 14, 17, 19, 21, and 26 were up-regulated at 72 hpi, while CmAux/IAA14, 17, 19, 20, 21, and 26 were induced at 168 hpi; no significant changes were observed at 24 hpi. In contrast, 'Top Mark' displayed a distinct temporal induction profile, with up-regulation of CmAux/IAA1, 11, 14, 16, and 28 specifically at 24 hpi, and no significant changes were detected at any other time point. It is noteworthy that in 'MR-1', while the expression level of CmAux/IAA11 did not change significantly; it remained consistently higher than that in 'Top Mark', suggesting that it may play a constitutive resistance role. Additionally, CmAux/IAA5, 17, 19, 20, 21, and 26, which were specifically induced and up-regulated by powdery mildew infection only in 'MR-1', exhibited lower basal expression levels than in 'Top Mark' under uninfected conditions. However, among these, the expression levels of CmAux/IAA5 and 26 following pathogen infection became significantly higher in 'MR-1' compared to 'Top Mark', suggesting they may serve as key responsive genes in defense against powdery mildew infection.
Transcriptional analysis following bacterial fruit blotch inoculation revealed distinct expression dynamics of CmAux/IAA genes between the resistant 'R', and susceptible 'S' cultivars (Fig. 7, Supplementary Table S14). In the 'R', the CmAux/IAA genes exhibited a phased induction pattern: CmAux/IAA6 was induced at 3 hpi; CmAux/IAA9 and 28 were induced at 6 hpi; CmAux/IAA1, 2, 5, 9, 11, 14, 17, 19, 22, 26, and 28 were induced at 12 hpi; and CmAux/IAA5, 6, and 21 were induced at 24 hpi. In contrast, the 'S' displayed a distinct temporal induction profile: CmAux/IAA3, 4, and 6 were up-regulated as early as 3 hpi; CmAux/IAA18 and 28 were up-regulated at 6 hpi; CmAux/IAA5, 6, and 26 were up-regulated at 12 hpi; and by 24 hpi, CmAux/IAA1, 3, 4, 6, 8, 12, 14, 15, 16, 20, 22, 24, 26, and 27 all showed up-regulated expression. Furthermore, CmAux/IAA11 and 19, which was up-regulated in response to pathogen infection in the 'R', was conversely down-regulated in the 'S'. Notably, several genes exhibited consistent expression patterns in both the 'R' and 'S'. For instance, CmAux/IAA6 was induced at both 3 and 24 hpi, while CmAux/IAA28 was up-regulated at 6 hpi, and both CmAux/IAA5 and 26 were up-regulated at 12 hpi in both genotypes. Based on the above analysis, although the expression levels of multiple CmAux/IAA genes changes upon bacterial fruit blotch infection; no gene in the 'R' exhibited sustained up-regulation across multiple time points during the inoculation period, nor did its expression level significantly exceed that in the 'S'. Therefore, it remains difficult to identify key CmAux/IAA genes responsible for defense against bacterial fruit blotch. However, in the 'S', we observed that CmAux/IAA14, 15, and 26 were significantly up-regulated at 24 hpi, and their expression levels were higher than those in the 'R'. This suggests that these genes may play a negative regulatory role in the disease resistance process.
-
Recent advances in melon whole-genome sequencing have enabled genome-wide identification and analysis of functional gene families. The auxin signaling pathway, in which Aux/IAA proteins play a key role, regulates diverse processes including development, organogenesis, and stress responses. This study systematically identifies the Aux/IAA gene family in melon, providing new insights into its roles in growth, development, and adaptation to biotic and abiotic stresses.
Characteristics and molecular evolution of the CmAux/IAA gene family
-
Aux/IAA genes have been identified in multiple plant species. In this study, 29 Aux/IAA genes were identified in the melon genome, a number comparable to those found in rice (31)[19], Arabidopsis (29)[18], and tomato (25)[20]. Research indicates functional divergence among Aux/IAA genes. In Arabidopsis, this divergence may depend on protein molecular structure and expression patterns. Previous studies have reported that the Aux/IAA gene family typically contains four conserved domains: I, II, III, and IV. In this study, four conserved motifs were identified in the CmAux/IAA gene family. Among them, 10 members (CmAux/IAA6, 7, 8, 11, 14, 15, 17, 19, 25, and 27) possess all four typical domains, while the remaining 19 non-classical Aux/IAA proteins lack one or more domains. Specifically, Domain I (Motif 1) contains conserved leucine residues and an LxLxL motif, which is crucial for the repressive function of IAA proteins[47]. Nine CmAux/IAA proteins (CmAux/IAA4, 5, 9, 12, 13, 20, 21, 24, and 29) lack this domain and may have lost their auxin signaling repression function. Two CmAux/IAA proteins (CmAux/IAA21 and 29) lack Domain II (Motif 2). Similar to Arabidopsis AtIAA20, these proteins may be unable to degrade rapidly under basal or high auxin conditions[48], thereby interfering with the auxin signal transduction pathway. Domains III and IV mediate the interaction between Aux/IAA and ARF proteins[49]. All CmAux/IAA proteins retain Domain III (Motif 3), which is essential for regulating the expression of downstream auxin-responsive genes[50]. Twelve CmAux/IAA proteins (CmAux/IAA1, 2, 3, 5, 10, 16, 18, 21, 22, 23, 26, and 28) lack Domain IV (Motif 4), and may have lost their ARF-binding capacity.
The Aux/IAA gene family encodes a class of rapidly auxin-induced, short-lived nuclear proteins[46]. Functioning as key repressors in the nuclear auxin signaling pathway, Aux/IAA proteins inhibit transcription when stable and promote it upon degradation. This regulation is achieved through their ability to form various dimers with ARFs, thereby modulating gene expression through multiple mechanisms. This study revealed that the promoter regions of CmAux/IAA family members are enriched with short-lived functional elements, which further corroborates the classical theory of Aux/IAA proteins as 'short-lived nuclear proteins'[46].
Auxin, a pivotal phytohormone regulating plant development, interacts with other hormones—including abscisic acid, brassinosteroids, cytokinins, ethylene, gibberellins, jasmonic acid, and salicylic acid[51]. As a crucial gene family in auxin signaling, Aux/IAA genes also mediate responses to other hormonal signals, such as abscisic acid, cytokinin, and salicylic acid. For instance, the Arabidopsis mutant iaa16-1 exhibits reduced sensitivity to both auxin and abscisic acid, severe growth impairment at maturity, and complete sterility in the homozygous state[52]. Our promoter analysis identified a notable enrichment of abscisic acid-responsive elements in CmAux/IAA genes, in line with the previously suggested involvement of these proteins in mediating auxin and ABA signal integration.
Involvement of CmAux/IAA genes in abiotic stress responses
-
Auxin and abscisic acid (ABA) are key regulators of plant abiotic stress tolerance. Previous work has shown that Aux/IAA genes, integral to auxin signaling, can be induced by stresses like drought. For example, silencing OsIAA20 in rice compromised drought and salt tolerance, in part through its regulation of the ABA-responsive gene OsRab21[53]. In our study, several CmAux/IAA genes were identified as salt-responsive (CmAux/IAA1, 2, 6, 9, 11, and 18), while CmAux/IAA11 emerged as a core, consistently up-regulated gene under drought stress across all six melon varieties.
Furthermore, previous studies have demonstrated that heterologous overexpression of the grape VaIAA3 gene significantly enhances Arabidopsis thaliana low-temperature stress tolerance. Through RNA-seq analysis, it was revealed that this gene regulates cold resistance in Arabidopsis via the auxin, ABA, jasmonic acid (JA), and ethylene signaling pathways[54]. Public RNA-Seq data implicated CmAux/IAA genes in melon chilling response. Among them, CmAux/IAA26 and 28 were up-regulated, suggesting key roles in chilling injury response. Beyond low-temperature stress, elevated temperatures also trigger the up-regulation of CmAux/IAA12. In rice, thermal stress induces expression of the grain-specific OsIAA29, which competitively displaces OsIAA21 from OsARF17 binding, thereby enhancing OsARF17-mediated transcriptional activation of starch and protein synthesis-related genes, and ultimately modulating grain development through the auxin signaling pathway[12].
As key regulators of phototransduction, PHYTOCHROME INTERACTING FACTOR 4 (PIF4) directly binds to the promoter regions of IAA19 and IAA29 to activate their expression, thereby negatively regulating phototropism and auxin signaling pathways through repression of ARF7 activity[55]. Recent studies demonstrate that among 29 Aux/IAA family genes, 21 are directly regulated by PIF3/4/5, with 12 showing significant up-regulation under natural shade conditions[56], highlighting the central regulatory role of Aux/IAA genes in plant light responses. Notably, our study reveals that all CmAux/IAA members contain light-responsive elements, with each CmAux/IAA gene promoter harboring at least seven such elements, further suggesting the potential importance of this gene family in light signaling networks. Under low-light stress, CmAux/IAA14, a homolog of Arabidopsis IAA19, was significantly up-regulated, suggesting it may be a key gene involved in the response to low light (Fig. 8). Furthermore, in the control group, CmAux/IAA2 and CmAux/IAA25, which belong to the same evolutionary clade VIIIb as Arabidopsis IAA16, exhibited the highest expression levels among all CmAux/IAA genes, supporting their potential key role in hypocotyl development.
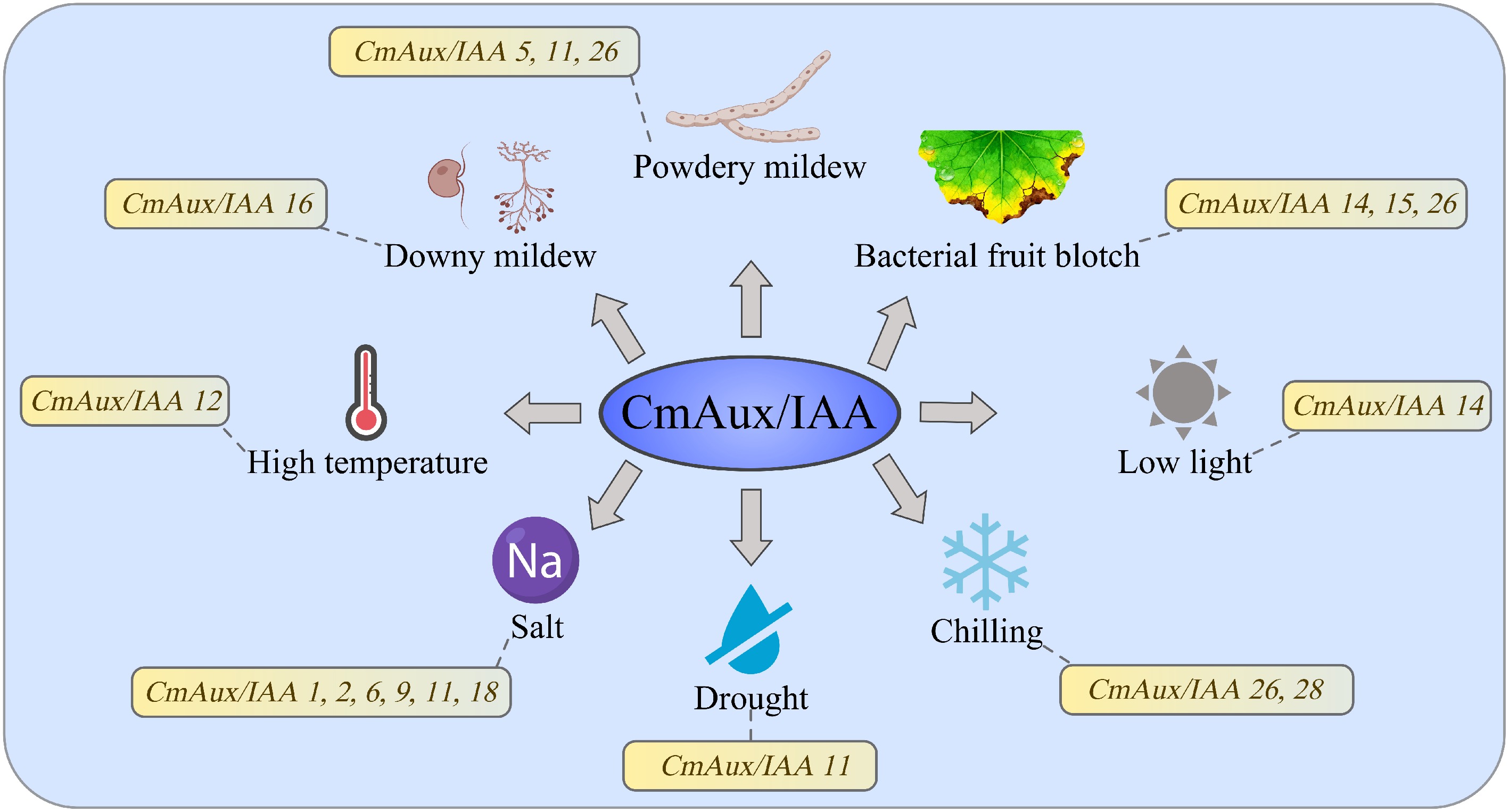
Figure 8.
Prediction and screening of key CmAux/IAA candidate genes responsive to biotic and abiotic stresses based on transcriptome data.
Roles of CmAux/IAA genes in biotic stress responses
-
Auxin signaling plays a central role in plant-pathogen interactions. The core regulatory module involves Aux/IAA proteins, which are targeted for ubiquitination and degradation by the SCFTIR1 complex to activate auxin responses. Notably, pathogens can subvert this pathway; for instance, the Tomato chlorosis virus p22 protein disrupts SCFTIR1 assembly to inhibit Aux/IAA turnover and promote infection[57]. Conversely, in cassava, enhanced expression of MeAux/IAA genes boosts resistance to Xanthomonas axonopodis pv. manihotis[58], illustrating the dual role of this family in immunity.
In this study, transcriptomic profiling revealed dynamic, genotype-specific expression of CmAux/IAA genes under pathogen challenge. CmAux/IAA16 was steadily induced by downy mildew in both resistant and susceptible lines, indicating a conserved response role. For powdery mildew, CmAux/IAA11 showed constitutively higher expression in the resistant genotype 'MR-1', whereas CmAux/IAA5 and 26 were specifically induced only in 'MR-1', suggesting distinct constitutive and inducible resistance mechanisms. Following bacterial fruit blotch infection, CmAux/IAA14, 15, and 26 were markedly upregulated at 24 hpi specifically in the susceptible cultivar. Based on established mechanisms where pathogen effectors directly target Aux/IAA proteins[15−17], we propose that these three genes may represent susceptibility-associated factors potentially exploited by the pathogen to dampen host defense. An alternative hypothesis is that their induction reflects a disrupted auxin signaling feedback loop during infection, tilting the balance from defense toward growth. Direct protein-interaction studies and functional validation in melon are needed to distinguish whether these genes are effector targets, or components of a dysregulated signaling circuit that promotes susceptibility.
Collectively, CmAux/IAA genes exhibit tissue-specific and temporal-specific expression patterns, and are differentially expressed in response to abiotic stresses such as salinity, drought, chilling, high temperature, and low light, as well as to infections by fungal and bacterial pathogens including downy mildew, powdery mildew, and bacterial fruit blotch (Fig. 8). Future research may employ transgenic and gene-editing technologies to further elucidate the functional mechanisms of CmAux/IAA genes, particularly in plant development, responses to biotic and abiotic stresses, and hormonal signaling pathways. It should be noted that the sampling sites in this study varied across different stress treatments. This design was based on classical physiological studies identifying the most sensitive responding tissues for each specific stress type, which facilitates the capture of early-stage, stress-specific signals. However, differences in baseline gene expression across tissues may introduce some influence on the direct comparability of expression patterns between different stress types. Therefore, this study primarily focuses on comparisons across genotypes or time points within the same stress type, while cross-stress conclusions are intended to indicate the potential of genes to respond to multiple stresses. The precise mechanisms underlying such responses will require further validation through experiments with uniform tissue sampling in future studies.
-
In summary, this study identified 29 Aux/IAA genes in the melon genome, and classified them into eight clades based on phylogenetic analysis. Gene structure and motif composition were largely conserved within subfamilies. Promoter analysis revealed an enrichment of cis-acting elements related to development, hormone response, and stress, consistent with the multi-faceted regulatory roles of Aux/IAA proteins. Transcriptomic profiling further showed that CmAux/IAA expression is both tissue-specific and temporally regulated, and is differentially modulated under abiotic stresses. Specifically, under abiotic stress, CmAux/IAA11 emerged as a core drought-responsive gene, while CmAux/IAA26 and 28 were markedly up-regulated under chilling stress. CmAux/IAA12 and CmAux/IAA14 appear to be involved in adaptation to high temperature and low light, respectively. In response to biotic stress, CmAux/IAA16 was identified as a key gene for downy mildew response. During powdery mildew infection, CmAux/IAA11 may play a constitutive resistance role, whereas CmAux/IAA5 and 26 likely function as inducible resistance genes. In bacterial fruit blotch-susceptible cultivars, the elevated expression of CmAux/IAA14, 15, and 26 implies their potential negative regulatory role in disease response. Collectively, these findings provide a valuable resource for future functional characterization of Aux/IAA genes, and offer new insights for research into plant growth, development, and adaptation to environmental stresses.
This research was supported by Xinjiang Academy of Agricultural Sciences Young Science and Technology Core Talents Innovation Capacity Building Project (xjnkq-2023032), Xinjiang Academy of Agricultural Sciences Self-Supported Cultivation Special Project (xjnkycxzx-2022002), Special Fund for Basic Scientific Research Operating Funds of Public Welfare Research Institutes in Xinjiang Uygur Autonomous Region (KY2023027) and National Natural Science Foundation of China (32460758). We are grateful to Professor Tingchang Zhao from the Institute of Plant Protection, Chinese Academy of Agricultural Sciences for kindly providing the Acidovorax citrulli strain Pslb 38 used for bacterial fruit blotch inoculation in this study.
-
The authors confirm their contributions to the paper as follows: study conception and design: Liu B; data collection: Ling Y, Wei Y, Muhamatihan M; analysis and interpretation of results: Ling Y, Chen X, Zhang X; draft manuscript preparation: Ling Y, Chen X, Yang S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yueming Ling, Xuanye Chen
- Supplementary Table S1 Physicochemical properties of Aux/IAA gene family members in melon.
- Supplementary Table S2 Segmentally duplicated Aux/IAA gene pairs identified in the melon genome.
- Supplementary Table S3 Orthologous relationships of Aux/IAA genes between melon and three representative plants.
- Supplementary Table S4 Prediction of cis-regulatory elements in promoter regions of Aux/IAA gene family in melon.
- Supplementary Table S5 Global expression profiling of CmAux/IAA genes in Japanese muskmelon 'Harukei-3' utilized FPKM values from a combined dataset of 27 and 15 developmental tissues, sourced from NCBI BioProjects PRJDB6414 and PRJNA603204.
- Supplementary Table S6 Expression profiling of the CmAux/IAA gene family in melon 'Vedrantais', contrasting young and senescent leaves. Data are presented as TPM values.
- Supplementary Table S7 Expression profiling of the CmAux/IAA gene family in melon under salt stress. Gene expression values are derived from a RNA-seq dataset of the muskmelon cultivar 'Longqing' (NCBI BioProject: PRJNA987131). Data are presented as TPM values.
- Supplementary Table S8 Expression profiles of CmAux/IAA gene family in melon under drought stress. Gene expression levels are represented by TPM values.
- Supplementary Table S9 Expression profiles of CmAux/IAA gene family in melon under chilling stress. Gene expression levels (FPKM) were derived from RNA-seq dataset of NCBI BioProject PRJNA553119.
- Supplementary Tables S10 Expression profiles of CmAux/IAA gene family in melon under high tempreture stress. Gene expression levels are represented by TPM values according to standard transcriptome analysis protocols.
- Supplementary Table S11 Expression profiles of CmAux/IAA gene family in melon under low light stress. Gene expression levels are represented by TPM values.
- Supplementary Table S12 Expression profiles of CmAux/IAA gene family in melon following downy mildew (DM) inoculation. Gene expression levels are represented by TPM values.
- Supplementary Table S13 Expression profiling of the CmAux/IAA gene family in melon cultivars 'MR-1' and 'Top Mark' following inoculation with powdery mildew (PM). Gene expression levels (FPKM) were derived from RNA-seq dataset of NCBI BioProjects PRJNA358655 and PRJNA358674.
- Supplementary Table S14 Expression profiles of CmAux/IAA gene family in melon under bacterial fruit blotch (BFB) infection. Expression levels are presented as FPKM values, normalized through standard transcriptomic analysis workflows.
- Supplementary Fig. S1 Chromosomal mapping of the CmAux/IAA gene family. The number in the left represent the length of chromosome.
- Supplementary Fig. S2 Pairwise collinearity analysis of Aux/IAA genes between C. melo and each of the three species (C. sativus, S. lycopersicum, A. thaliana).
- Supplementary Fig. S3 Distribution of light-responsive elements and short-functional motifs in the predicted cis-regulatory elements of the melon Aux/IAA gene family.
- Supplementary Fig. S4 Differential expression of CmAux/IAA genes between young and senescent leaves in 'Vedrantais'.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ling Y, Chen X, Muhamatihan M, Wei Y, Yang S, et al. 2026. Genome-wide identification and expression analysis of the melon (Cucumis melo L.) Aux/IAA gene family in response to abiotic and biotic stresses. Vegetable Research 6: e005 doi: 10.48130/vegres-0026-0002 

Genome-wide identification and expression analysis of the melon (Cucumis melo L.) Aux/IAA gene family in response to abiotic and biotic stresses
- Received: 13 December 2025
- Revised: 04 January 2026
- Accepted: 13 January 2026
- Published online: 31 January 2026
Abstract: Cucumis melo L. is an important horticultural crop whose cultivation is limited by environmental stresses. The Auxin/Indole-3-Acetic Acid (Aux/IAA) gene family is central to auxin signaling and plant stress adaptation, yet its functions in melon are unclear. This study identified and characterized the Aux/IAA family in melon to elucidate its roles under stress. Twenty nine CmAux/IAA genes from the melon genome were identified, and phylogenetically classified into eight clades. Collinearity analysis indicated that segmental duplication primarily drove the family's expansion. Promoter analysis revealed cis-regulatory elements linked to development, hormone response, and stress. Transcriptome analysis showed that these genes display characteristic spatiotemporal expression patterns across tissues, and respond differentially to abiotic (salt, drought, chilling, heat, low light), and biotic (downy mildew, powdery mildew, bacterial fruit blotch) stresses. Specifically, CmAux/IAA11 was a core drought-responsive gene; CmAux/IAA26 and 28 were up-regulated under chilling; CmAux/IAA12 and 14 likely function in heat and low light adaptation, respectively. Regarding biotic stress, CmAux/IAA16 responded to downy mildew, CmAux/IAA11 may confer constitutive resistance to powdery mildew, while CmAux/IAA5 and 26 are inducible resistance genes, and elevated expression of CmAux/IAA14, 15, and 26 in susceptible cultivars suggested a negative regulatory role during bacterial fruit blotch. Our findings provide a foundation for functional studies of CmAux/IAA genes, and new insights into melon stress responses.