








-
Plants frequently encounter various abiotic stresses, such as salt accumulation, drought, and environmental fluctuations such as high and low temperatures. These stresses severely impact crop growth and development, photosynthetic efficiency, and yield; thus, improving crop adaptability under adverse conditions has become a critical goal for sustainable agricultural development[1]. As a widely distributed intracellular second messenger, calcium ions (Ca2+) play a crucial role in plants' responses to these stresses[2]. Transient changes in Ca2+ levels form specific calcium signatures, providing a precise signal encoding basis for downstream sensing systems and triggering a series of complex physiological responses and gene expression adjustments[3]. Plants possess multiple types of Ca2+ sensors, including calmodulin (CaM/CML), calcium-dependent protein kinases (CDPKs), and the CBL-CIPK signaling system. CBL (calcineurin B-like proteins), acting as Ca2+ sensors, interact with CIPKs (serine/threonine protein kinases) to form CBL-CIPK complexes. By regulating various transporters and signaling modules, they participate in responses to abiotic stresses such as salinity, drought, and low temperatures[4]. CIPK, as a downstream target of CBL, undergoes specific interaction with CBL mediated by Ca2+ to form a complex, thereby participating in regulating plant responses to various environmental stimuli. CIPK acts as a key intermediary, bridging Ca2+ sensors and their downstream targets, playing a pivotal role in various protein-protein interactions[5]. Research has revealed that CIPK functions by interacting specifically with one or more CBL proteins, thereby participating in the regulation of plant responses to various abiotic stresses such as drought, salinity, low temperatures, and heavy metals[6].
In plants, CIPK plays a key role in salt stress responses by interacting with CBL (Calcineurin B-like) proteins. CBL proteins act as calcium sensors. When plants undergo salt stress, intracellular calcium concentrations undergo transient changes. Upon detecting these calcium signals, CBL proteins bind to CIPK proteins, thereby activating their kinase activity. Activated CIPK proteins can phosphorylate downstream target proteins, regulating plant physiological and biochemical processes to help plants adapt to salt stress[7]. In Arabidopsis, AtCIPK24 (also known as SOS2) interacts with CBL4 (SOS3) to phosphorylate the SOS1 protein, which is a Na+/H+ counter-transport pump located on the cell membrane[8]. By phosphorylating SOS1, the AtCIPK24/CBL4 complex enhances SOS1 activity, promoting the efflux of excess sodium ions from cells and thereby improving plant salt tolerance[9]. During heterologous expression experiments in yeast, it was discovered that SpCIPK8 from Sesuvium portulacastrum forms a complex with SpCBL10. This complex regulates the activity of the plasma membrane Na+/H+ counter-transporter by recognizing and binding to the C-terminal segment of SpSOS1, targeting specific amino acid residues (Ser1144 and Ser1146)[10]. Overexpression of MdSOS2L1 in apple exhibited significantly increased levels of antioxidants such as anthocyanins and malic acid, while also demonstrating enhanced salt tolerance[11]. In liverwort (Marchantia polymorpha), knockout mutants of MpCIPK-B exhibited leaf yellowing and even wilting under salt stress conditions[12]. CsCBL4 and CsCIPK6 were found to specifically interact in cucumber (Cucumis sativus), jointly responding to salt stress, with their silencing reducing salt tolerance[13]. In Arabidopsis, AtCIPK16 is significantly upregulated under salt stress conditions, suggesting its involvement in salt response pathways[14]. In rice, OsCIPK9 has been demonstrated to be a key regulatory factor for salt-alkali stress tolerance, with inactivating mutant lines exhibiting enhanced stress resistance[15]. Additionally, heterologous expression of the algal-derived CIPK (NtCIPK11) promoted proline accumulation in transgenic tobacco plants, thereby enhancing tolerance under salt stress[16]. Similarly, CIPK proteins play a critical role in drought stress responses through interactions with CBL proteins, which act as calcium sensors, binding to CIPK proteins upon detecting calcium signals during stress. When plants undergo drought stress, intracellular calcium concentrations surge transiently. Upon detecting these calcium signals, CBL proteins bind to CIPK proteins, thereby activating their kinase activity[17]. Activated CIPK proteins can phosphorylate downstream target proteins, regulating plant physiological and biochemical processes to help plants adapt to drought stress[18]. In Arabidopsis, AtCIPK1 negatively regulates ABA signaling in drought responses by phosphorylating the ABA receptor PYL4, thereby achieving negative feedback regulation of the drought response system[19]. In quinoa, most CqCIPK genes showed significant upregulation in roots and leaves following drought stress (6 h), with CqCIPK01 and CqCIPK25 exhibiting expression increases of 77- and 138-fold, respectively[20]. In rice, OsCIPK17 positively regulates drought responses through mechanisms that include promoting citrate accumulation in the TCA cycle, thereby enhancing root development and energy metabolism[21]. In chili peppers, CaCIPK7 participates in drought tolerance responses through ROS scavenging and the ABA signaling pathway, significantly enhancing drought resistance[22]. Analysis of multiple TtCIPK gene families in durum wheat revealed that TtCIPK33 is a candidate gene highly involved in drought response, potentially contributing to enhanced drought tolerance[23].
In Arabidopsis thaliana, a CIPK protein kinase gene, AtCIPK, was identified in higher plants for the first time[24]. With the rapid advancement of high-throughput genome sequencing, CIPK and CBL genes have been systematically analyzed in numerous species. The OsCIPK family was systematically annotated at the whole-genome level in rice (Oryza sativa), and sequence and expression analyses revealed its vital role in stress responses[25]. In Chinese cabbage (Brassica rapa ssp. pekinensis), 18 CBLs and 47 CIPKs were analyzed respectively[26]. Besides, 80 and 78 CIPK genes were identified in upland cotton (Gossypium hirsutum) and sea island cotton (G. barbadense), respectively[27]. Eight CBL and 20 CIPK genes were analyzed in the grapevine (Vitis vinifera) genome and classified into subgroups based on phylogenetic and structural characteristics[28]. In tomato (Solanum lycopersicum), 22 SlCIPK genes were identified and combined with expression analysis, pointed to their functions related to abiotic stress[29]. Twenty-seven StCIPK genes were identified in potato (Solanum tuberosum), categorized into 'few/many introns' based on gene structure, with some genes associated with drought/ABA induction[30]. In pepper (Capsicum annuum), nine CaCBL and 26 CaCIPK genes were identified, most of which showed significant expression changes under salt/osmotic stress and pathogen stress[31]. Nineteen CBLs and 51 CIPKs were identified in Turnip (Brassica rapa var. rapa), with family expansion closely associated with segmental duplication (a type of gene duplication involving large chromosomal segments)[32]. An integrated comparison of hickory (Carya illinoinensis) with other species, highlights the diversity in CBL/CIPK family members and functional variations, serving as one of the representative data sources for woody fruit trees[33]. In bananas (Musa spp.), 34 CIPK members were identified and characterized, filling an information gap regarding this crop at the CIPK family level[34]. These findings collectively highlight the evolutionary diversity and functional specialization of CIPK-CBL signaling across plant species.
Lettuce (Lactuca sativa L.), belonging to the Asteraceae family, is one of the most widely cultivated leafy vegetables globally. It holds significant economic value and serves as an important source of natural plant nutrients in the human diet[35]. The completion of high-quality reference genomes has enabled rigorous gene family analysis. Lettuce is primarily a cold-tolerant annual crop suited for low-temperature growth, yet it is highly sensitive to high temperatures, salt stress, and drought. These abiotic stresses significantly impact its quality and yield[36]. The chromosome-level assembly of cv. Salinas was first reported in 2017, followed by improved gap-free or telomere-to-telomere assemblies and additional morphotype-specific genomes. Recently, the lettuce super-pan-genome, which expanded the catalog of presence-absence and structural variants associated with traits and stress adaptation[37−41]. Despite these genomic resources, the CIPK gene family in lettuce remains unexplored. While extensive research has been conducted on lettuce morphology, cultivation management, stress physiology, and nutritional quality, systematic bioinformatics analyses of lettuce gene families are still limited. With the completion of lettuce whole-genome sequencing, extensive genomic data has enabled comprehensive identification of its gene families. This study aims to systematically identify and analyze the CIPK gene family in lettuce using bioinformatics methods, including phylogenetic relationships, gene structural features, and prediction of promoter cis-acting elements. Simultaneously, transcriptomic and gene expression analyses under salt and drought stress conditions revealed that multiple LsCIPK genes (e.g., LsCIPK7, LsCIPK26, and LsCIPK32) are strongly induced, suggesting their potential roles in abiotic stress responses. This study provides candidate genes and theoretical support for deciphering the regulatory mechanisms of the CBL-CIPK signaling pathways in lettuce and breeding salt-tolerant and drought-tolerant lettuce varieties.
-
Seeds of the uniform and plump lettuce (Lactuca sativa L.) cultivar 'American Fast-Grower' (Institute of Vegetable and Flower Crops, Chinese Academy of Agricultural Sciences, Beijing, China) were selected for subculture propagation. The culture medium was based on MS medium supplemented with 0.5 mg/L 6-BA, and 0.1 mg/L NAA. Meanwhile, 30 g sucrose and 7 g agar were added per L of medium, adjusting the pH to 5.8−6.2. Culture conditions are set at 25 °C with a 16-h light/8-h dark photoperiod at an irradiance of 200 μmol/m2/s. Subculture was performed every 30 d. Subsequently, robust and uniformly developed tissue-cultured seedlings, four weeks old, were selected and treated with 20% PEG6000 and 200 mM NaCl, respectively. After 24 h, apical young leaves from the control (CK) and both stressed groups were collected and frozen immediately in liquid nitrogen, and then stored at −80 °C. Total RNA was extracted using the FastPure Universal Plant Total RNA Isolation Kit (Vazyme, Nanjing, China). cDNA synthesis was performed with the HiScript III 1st Strand cDNA Synthesis Kit (+gDNA wiper) (Vazyme, Nanjing, China).
Identification of the LsCIPK gene in lettuce
-
The lettuce genome and protein sequences were downloaded from the NCBI database (
www.ncbi.nlm.nih.gov ), and the hidden Markov model (HMM, Pfam: PF03822) was obtained from the Pfam database (http://pfam.xfam.org ). Additionally, the protein kinase domain (PF00069) was referenced to assist in the screening process. Arabidopsis and rice CIPK protein sequences were used as query sequences, and BLASTP alignment was performed against the local lettuce protein database (E-value set to 1e-10) to obtain preliminary candidate proteins. Subsequently, the hmmsearch program of HMMER was performed to further screen the candidate sequences based on PF03822[42]. Following domain identification and validation using Pfam and NCBI-CDD databases, the lettuce CIPK gene family members were definitively determined. The gene IDs were then extracted from the lettuce genome annotation files for further analysis.Analysis of phylogenetic, the conserved domains and gene structure of the LsCIPK protein in lettuce
-
To elucidate the evolutionary relationship between the lettuce and Arabidopsis CIPKs, multiple sequence alignment of the LsCIPK and AtCIPK protein sequences was performed using Clustal X 2.0 software. Based on the alignment results, a phylogenetic tree was constructed using the maximum likelihood (ML) method by MEGA 7.0 software. Model selection was based on the minimum Bayesian Information Criterion (BIC) value, and 1,000 bootstrap replicates were performed to assess branch reliability[43].
Conserved motif analysis of LsCIPK protein sequences was performed using the MEME website (
https://meme-suite.org/meme )[44], and their conserved domains were identified via the CDD-search platform. Subsequently, the Gene Structure View feature in TBtools was used, MEME results, genomic annotation information, and domain data were integrated and visualized in a single diagram[45].Characterization of the LsCIPK protein from lettuce
-
The ExPASy ProtParam tool (
https://web.expasy.org/protparam ) was taken to perform a predictive analysis of the fundamental characteristics of the CIPK protein amino acid sequence, including physicochemical properties such as amino acid content, molecular weight, isoelectric point, hydrophobicity, and instability index[46]. Additionally, the subcellular localization was predicted using the online tool Cell-PLoc 2.0 (www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc ).Analysis of the cis-acting elements in the promoter of LsCIPKs
-
The GFF3 file was downloaded from the Phytozome v13 database, and TBtools was used to extract the sequence spanning 2,000 bp upstream of the ATG start codon. Subsequently, the obtained sequences were uploaded to the PlantCare online platform (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) to predict cis-acting elements in the promoter region, and the results would be visualized by TBtools[47].Collinearity and Ka/Ks analysis of CIPKs in lettuce
-
To investigate evolutionary events within gene families, MCScanX software was employed for collinearity analysis to identify homologous blocks within the lettuce genome and between lettuce, Arabidopsis, and rice, thereby detecting tandem and segmental duplication events[48]. Subsequently, the Ka/Ks Calculator module in TBtools was used to compute the non-synonymous substitution rate (Ka), synonymous substitution rate (Ks), and the Ka/Ks ratio for collinear gene pairs. Ka, Ks data, and Ka/Ks visualization plots were generated and displayed by TBtools.
Intersection analysis of differentially expressed genes (DEGs) under salt stress and drought stress
-
To compare the transcriptional responses of lettuce under salt stress vs drought stress conditions, differential expression gene (DEG) analysis was performed on the NaCl-treated group (CK vs NaCl) and the PEG-treated group (CK vs PEG), respectively. Based on the normalized expression matrix, DESeq2 was employed for differential expression analysis. The screening threshold was set to |log2FC| ≥ 1 and FDR < 0.05. The DEGs obtained under the two treatment conditions were imported into TBtools (v1.098) separately. The Venn Diagram module was used to perform intersection analysis, identifying co-differentially expressed genes (co-DEGs) involved in both salt and drought responses, as well as genes specific to each treatment. The final intersection statistics and Venn diagram visualization were automatically generated by TBtools.
GO annotation and KEGG enrichment analysis of specific differentially expressed genes (DEGs)
-
To further elucidate the potential functions of specific differentially expressed genes (specific DEGs) under salt stress and drought stress, as well as their common differentially expressed genes, Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed on the three categories of specific genes. First, the list of differentially expressed genes (DEGs) was imported into TBtools (v1.098). The GO enrichment module was used to perform statistical classification of genes into biological processes (BP), molecular functions (MF), and cellular components (CC). Subsequently, pathway enrichment analysis was conducted using the KEGG Enrichment module based on the official KEGG database, with a significance threshold set at Q < 0.05 (Benjamini–Hochberg correction). Both bar charts and scatter plots of enrichment results were automatically generated by TBtools for subsequent result visualization.
Analysis of LsCIPKs gene expression in lettuce under high salinity and drought stress
-
To analyze the expression patterns of the LsCIPKs under salt stress and drought stress conditions, gene expression levels under various treatment conditions were calculated and normalized based on transcriptome sequencing (RNA-seq) data. The screening criteria for differentially expressed genes were set as: |log2FC| ≥ 1 and FDR < 0.05 (DESeq2, Benjamini–Hochberg correction). The raw sequencing data was first aligned against the lettuce reference genome, using Hisat2 to obtain alignment results. Then, the FPKM expression levels for each gene were calculated using StringTie/FeatureCounts. Subsequently, the expression matrices of all samples were integrated. The expression data of LsCIPK genes were imported using TBtools (v1.098), and the Heatmap Illustrator module was employed to generate an expression heatmap illustrating the expression changes of LsCIPK genes under CK, NaCl, and PEG treatments.
qRT-PCR validation
-
Total RNA was isolated from finely ground lettuce leaf material using the Vanzyme RNA Extraction Kit (Nanjing Vanzyme Biotech Co., Ltd) after thorough grinding in liquid nitrogen. RNA concentration and purity were determined using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA). Samples with an OD260/280 ratio between 1.8 and 2.1 were considered acceptable for quality. Simultaneously, RNA integrity was assessed via 1% agarose gel electrophoresis. Subsequently, 1 μg of total RNA was taken as a template, and first-strand cDNA synthesis was performed in a 20 μL reaction system with the Evo M-MLV reverse transcription kit (Changsha Aikrui Biotechnology Co., Ltd.). The resulting cDNA was stored at −20 °C for subsequent experiments.
Real-time fluorescent quantitative PCR was performed in a 20 μL reaction system containing 10 μL of 2 × SYBR Green qPCR Mix (TSINGKE, Chengdu, China), with cDNA template quantity controlled below 100 ng. Additionally, 0.8 μL forward primer and 0.8 μL reverse primer (10 μmol/L; synthesized by BGI, Shenzhen, China) were added, with the remaining volume supplemented with RNase-free water. PCR amplification was performed on a CFX96 Touch real-time quantitative PCR instrument (Bio-Rad, USA). The reaction program was set as follows: 95 °C pre-denaturation for 5 min, followed by 40 cycles (95 °C for 10 s, 60 °C for 30 s); after amplification completion, a melting curve analysis was performed (95 °C for 15 s, 60 °C for 1 min, 95 °C for 15 s) to confirm the specificity of the amplified products.
Based on drought and salt transcriptome data from lettuce, 12 CIPK genes were randomly selected, and quantitative primers for these genes were designed using Primer 5.0 software (Table 1), and their specificity was verified in the NCBI database. Lettuce Actin was used as a reference gene for expression normalization, with relative expression levels calculated using the 2−ΔΔCt method[49]. Each sample included three biological replicates, with three technical replicates per biological replicate to ensure experimental accuracy and reproducibility.
Table 1. qRT-PCR primers.
Name Forward primers (5'-3') Reverse primers (5'-3') LsActin CTGGTGTGATGGTAGGTATGG CTCGTTGTAGAAAGTGTGATGC LsCIPK1 CGCGAAGGTCAAGTTTGCTC TGGGAGGTATCCAGCCATCA LsCIPK7 GTAGCACACCGTGACCTCAA CCGCCACCGTCTTCTCTATC LsCIPK13 TGAAGGTGACGGATTTCGGG CGACGTCGTCTTCGCAGATA LsCIPK21 CACGAGGTTTTGGCAAGCAA CCTCTGCCCGTTGTGATCTT LsCIPK25 GGCCTAAACCTGTCTGCCTT ACGGAAGCTCCTTCAAGACG LsCIPK26 TTGGACTCAGCGCTTTTTGC CACCAGAAACGACGGAGTGA LsCIPK29 AGGTCTACCATGGCTGGGAT ACGTCCATTTCGGAACACGA LsCIPK31 CCAGCAATTGATCACAGCCG GGTGCCACATAAGCAGGAGT LsCIPK32 CACATGTGGTACTCCGGCTT CACCATGCCATGCACAAACA Experimental results are presented as 'mean ± standard deviation (SD)'. Statistical analysis was performed using SPSS software (IBM SPSS Statistics 26.0). Data was first subjected to one-way analysis of variance (ANOVA), followed by Duncan's multiple comparison test to examine differences between treatments. A p-value < 0.05 was considered statistically significant.
Determining the subcellular localization of LsCIPK32
-
The complete coding sequence (CDS) of the LsCIPK32 lacking a stop codon was cloned into the 35S::GFP vector to generate the recombinant construct 35S::LsCIPK32-GFP. 35S::GFP and 35S::LsCIPK32-GFP plasmids were then transformed into Agrobacterium GV3101, suspended in 100 mL of infiltration buffer (0.042 g MES; 0.048 g MgCl2·6H2O; 100 mM acetophenone; pH 5.5), adjusted to an OD600 of 0.4–0.6, and injected into four-week-old Nicotiana benthamiana leaves. After 24 h of dark incubation and 48 h post-infiltration, imaging was performed using a Nikon C2-ER confocal microscope (Nikon, Tokyo, Japan) equipped with a 488 nm argon ion laser (15% intensity), a 515/30 nm emission filter, and a CFI Plan Apo λ 40 × oil immersion objective (NA = 1.3) to scan the Z-axis at 1 μm intervals.
-
We identified 32 CIPK genes in lettuce (designated LsCIPK1–LsCIPK32) through combined BLASTP and HMMER screening using Arabidopsis and rice CIPK protein sequences. Chromosomal localization analysis revealed that these genes were distributed across nine chromosomes, and exhibited a markedly unbalanced distribution (Fig. 1). Among them, LsChr1, LsChr5, and LsChr7 had relatively high numbers of genes (five, eight, and five members, respectively), while other chromosomes contained only one to four genes. Most genes exhibited scattered distribution, though localized gene clusters were also observed. For instance, LsCIPK14–LsCIPK17 formed a tightly clustered arrangement on LsChr5, potentially associated with gene duplication events.
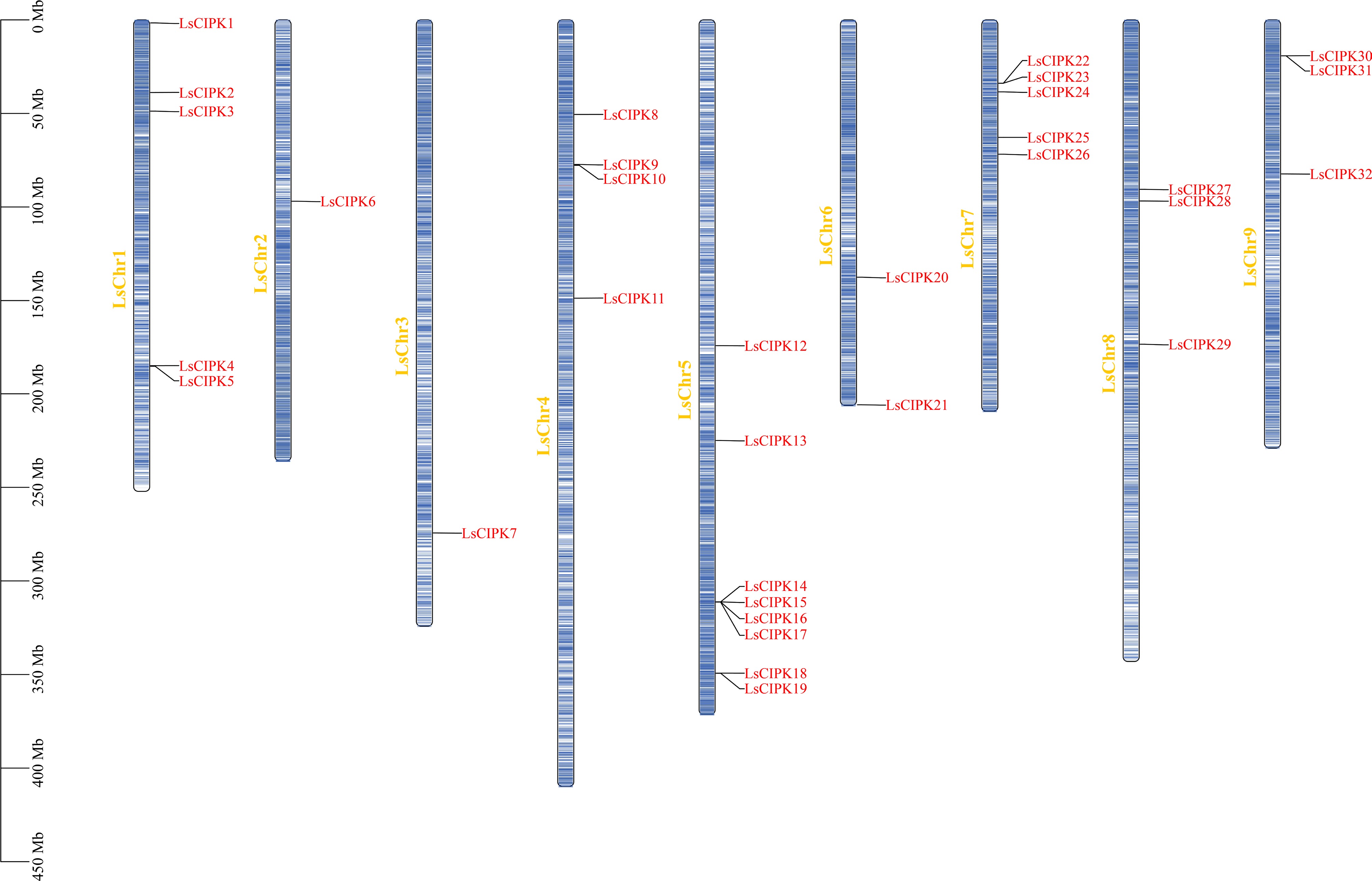
Figure 1.
Chromosomal distribution of the lettuce CIPK gene family. Blue bars represent lettuce chromosomes (LsChr1–LsChr9), with physical distances (Mb) shown on the left. Red text indicates CIPK gene names.
Physicochemical characterization of the lettuce CIPK gene family
-
To understand the fundamental characteristics of the LsCIPK protein, its amino acid length, molecular weight, isoelectric point, instability index, aliphatic index, hydrophilicity, and subcellular localization were analyzed (Table 2). The results indicated that LsCIPK proteins exhibited considerable variation, and the molecular weight of most members was concentrated within the 50−55 kDa. The theoretical isoelectric points predominantly fell within the alkaline range (pI > 8), consistent with the typical physicochemical characteristics of CIPK. The instability index results indicated that most members exhibited a tendency toward instability; however, their generally high aliphatic indices suggested overall good thermal stability. All LsCIPK GRAVY values were negative, indicating pronounced hydrophilic characteristics. Subcellular localization predictions suggested that LsCIPKs were primarily distributed in the cytoplasm and plasma membrane, while some members localize to chloroplasts, nuclei, or the endoplasmic reticulum, reflecting their potential involvement in signal transduction processes across multiple organelles.
Table 2. Physicochemical properties of the lettuce CIPK gene family.
Gene name The number
of amino acidMolecular
weight (Da)Theoretical pI Instability index Aliphatic
indexGrand average of hydropathicity Subcellular localization prediction LsCIPK1 904 102,285.14 6.54 40.05 88.36 −0.306 Plasma membrane; Cytoplasm; Endoplasmic Reticulum LsCIPK2 458 52,131.89 8.82 41.22 78.08 −0.398 Cytoplasm; Chloroplast; Nucleus LsCIPK3 446 5,0691.35 8 31.65 90.45 −0.4 Cytoplasm; Peroxisome; Nucleus LsCIPK4 483 54,689.08 8.88 42.81 88.36 −0.295 Chloroplast; Mitochondrion; Extracellular space LsCIPK5 482 54,396.49 8.96 29.92 77.86 −0.461 Chloroplast; Endoplasmic Reticulum; Plasma membrane LsCIPK6 2,890 328,456.35 7.36 38.07 83.37 −0.412 Plasma membrane; Nucleus; Endoplasmic Reticulum LsCIPK7 437 48,880.63 9.07 35.68 91.24 −0.308 Chloroplast; Golgi apparatus; Plasma membrane LsCIPK8 440 50,107.46 8.92 42.65 79.07 −0.526 Mitochondrion; Plasma membrane; Nucleus LsCIPK9 499 56,623.4 8.93 43.68 88.28 −0.323 Chloroplast; Vacuole; Cytoplasm LsCIPK10 462 52,465.55 9.03 28.57 81.43 −0.443 Cytoplasm; Endoplasmic Reticulum; Chloroplast LsCIPK11 441 50,187.94 9.02 35.63 84.22 −0.323 Cytoplasm; Nucleus; Endoplasmic Reticulum LsCIPK12 460 51,846.01 9.2 44.36 91.78 −0.272 Chloroplast; Plasma membrane; Cytoplasm LsCIPK13 427 47,976.37 8.86 36.01 82.41 −0.371 Plasma membrane; Endoplasmic Reticulum; Cytoplasm LsCIPK14 806 91,426.57 7.9 22.16 99.85 −0.109 Plasma membrane; Cytoplasm; Endoplasmic Reticulum LsCIPK15 878 99,016.35 8.26 32.55 89.25 −0.324 Plasma membrane; Endoplasmic Reticulum; Cytoplasm LsCIPK16 439 49,560.31 7.24 34.87 91.46 −0.294 Cytoplasm; Chloroplast; Endoplasmic Reticulum LsCIPK17 437 49,440.97 8.66 34.67 97.21 −0.25 Cytoplasm; Cytoskeleton; Chloroplast LsCIPK18 871 99,290.81 9.24 43.07 92.55 −0.367 Cytoplasm; Nucleus; Chloroplast LsCIPK19 432 48,527.79 8.85 40.83 90.67 −0.278 Chloroplast; Nucleus; Cytoplasm LsCIPK20 445 49,952.97 7.07 31.66 78.63 −0.356 Chloroplast; Plasma membrane; Nucleus LsCIPK21 451 50,541.88 6.3 35.49 93.08 −0.345 Cytoplasm; Nucleus; Chloroplast LsCIPK22 455 51,728.97 6.55 26.87 92.13 −0.283 Cytoplasm; Chloroplast; Plasma membrane LsCIPK23 459 52,112.4 9.07 48.33 85.14 −0.283 Chloroplast; Cytoplasm; Mitochondrion LsCIPK24 454 51,137.91 9.02 36.47 85.46 −0.369 Chloroplast; Mitochondrion; Golgi apparatus LsCIPK25 1,335 151,026.24 8.61 29.65 91.98 −0.245 Plasma membrane; Endoplasmic Reticulum; Vacuole LsCIPK26 428 48,055.44 9.06 32.74 82.69 −0.34 Plasma membrane; Cytoplasm; Endoplasmic Reticulum LsCIPK27 829 94,260.65 8.64 34.86 91.71 −0.243 Plasma membrane; Endoplasmic Reticulum LsCIPK28 428 47,731.15 9.23 32.27 84.28 −0.31 Plasma membrane; Endoplasmic Reticulum; Cytoplasm LsCIPK29 466 52,966.58 6.48 42.05 87.88 −0.283 Chloroplast; Extracellular space; Cytoplasm LsCIPK30 429 49,029.58 9.03 43.56 92.45 −0.316 Cytoskeleton; Cytoplasm; Cytoplasm & Nucleus LsCIPK31 458 52,243.51 8.99 42.3 88.54 −0.441 Cytoplasm; Cytoskeleton; Chloroplast LsCIPK32 456 5,1765.2 8.63 35.59 86.1 −0.269 Cytoplasm; Cytoskeleton; Golgi apparatus Analysis of the conservative domains and gene structure of the lettuce CIPK gene family
-
To reveal the structural characteristics of the LsCIPKs, its protein conserved domains and gene structures were analyzed. The results showed that all LsCIPK proteins contained the characteristic PKc-like kinase domain and the NAF (PF03822) regulatory domain, indicating they are typical members of the CIPK family (Fig. 2a). Most of proteins also harbored conserved segments such as CIPK_C or STKc_SnRK3, with highly consistent overall domain compositions, suggesting strong structural conservation within this family during evolution. In terms of gene structure, the number of exons varied considerably among different LsCIPK genes, ranging from 1 to 16, but nearly half of the members exhibit a single-exon structure (Fig. 2b). Some genes possessed a higher number of exons (12–16), indicating structural diversity within the family.
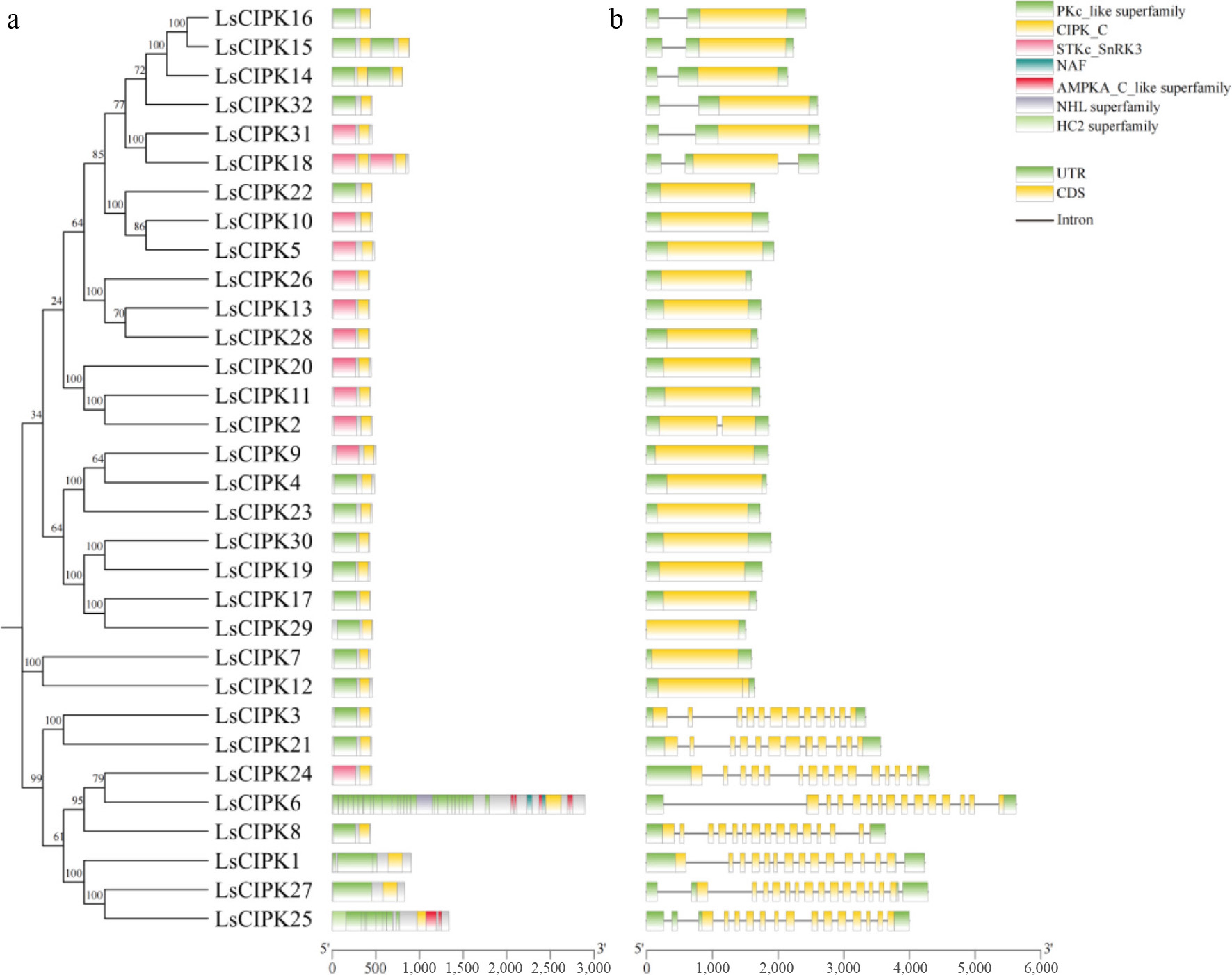
Figure 2.
Conserved domains and gene structure analysis of the CIPK gene family in lettuce. (a) Conserved domain distribution of the LsCIPK protein. Different colors represent different domain types; the corresponding domain names are labeled on the right. (b) Exon-intron structure of the LsCIPK gene. Yellow boxes represent coding regions (CDS), green boxes represent untranslated regions (UTR), and black lines represent introns.
Phylogenetic analysis of the lettuce CIPK gene family
-
To analyze the phylogenetic relationships of the lettuce CIPKs, multiple sequence alignments were performed using CIPK protein sequences from lettuce and Arabidopsis, and the phylogenetic tree was constructed based on the Neighbor-Joining (NJ) method (Fig. 3). The results indicated that the CIPK family is divided into five major branches (Group I–V), with a structural classification pattern highly consistent with that of Arabidopsis. Among the five branches, Group III, Group IV, and Group V contain a higher number of lettuce genes. Most of the lettuce members cluster closely with Arabidopsis homologs, indicating high evolutionary conservation within this family. Additionally, some genes cluster in pairs (e.g., LsCIPK15/16, LsCIPK3/21), which might result from gene duplication events.
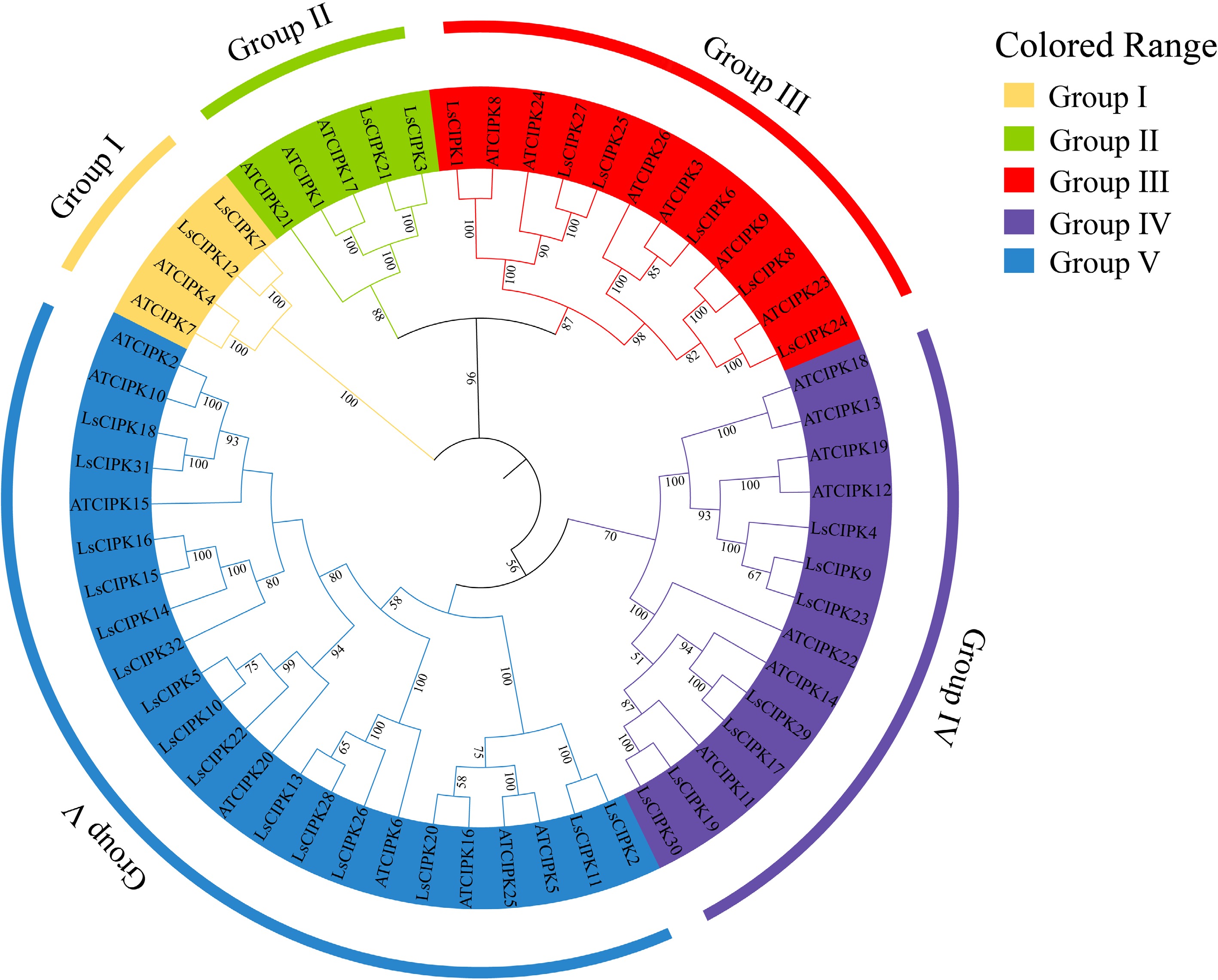
Figure 3.
Phylogenetic tree of the CIPK gene family in lettuce and Arabidopsis. Different colors represent five evolutionary branches (Group I–V). CIPK proteins in lettuce and Arabidopsis are distinguished as 'LsCIPK' and 'AtCIPK'.
Analysis of cis-acting elements in the lettuce CIPK gene family
-
To elucidate the potential regulatory mechanisms of the LsCIPK gene, cis-acting elements within the 2,000 bp region upstream of its promoter were analyzed. A total of 49 classes and 813 cis-elements were identified, which can be categorized into four major functional groups: light response, hormone response, stress response, and growth-and-development-related responses (Fig. 4). The light-responsive elements were the most abundant, with Box4 and G-Box appearing 90 and 76 times, respectively. Among hormone response elements, the ABA response element ABRE is the most abundant (67 occurrences). Most genes simultaneously contained both MeJA-related CGTCA-motifs and TGACG-motifs, suggesting that LsCIPKs might be broadly involved in hormone-mediated signal regulation. Among stress response elements, ARE (67), MBS (24), and LTR (19) showed widespread distribution, indicating that the LsCIPK gene might play a role in responses to abiotic stresses such as anaerobiosis, drought, and low temperature. The marked variation in the number of stress elements among different members suggested functional differentiation in stress regulation. Additionally, multiple cis-elements associated with growth and development were detected, such as MSA-like, CAT-box, and circadian elements, indicating that the expression of LsCIPKs might also be regulated by developmental stages and circadian rhythms. Overall, the LsCIPK promoter region was rich in light-, hormone-, and stress-response elements, suggesting that this gene family might play a crucial role in environmental signal perception and response.
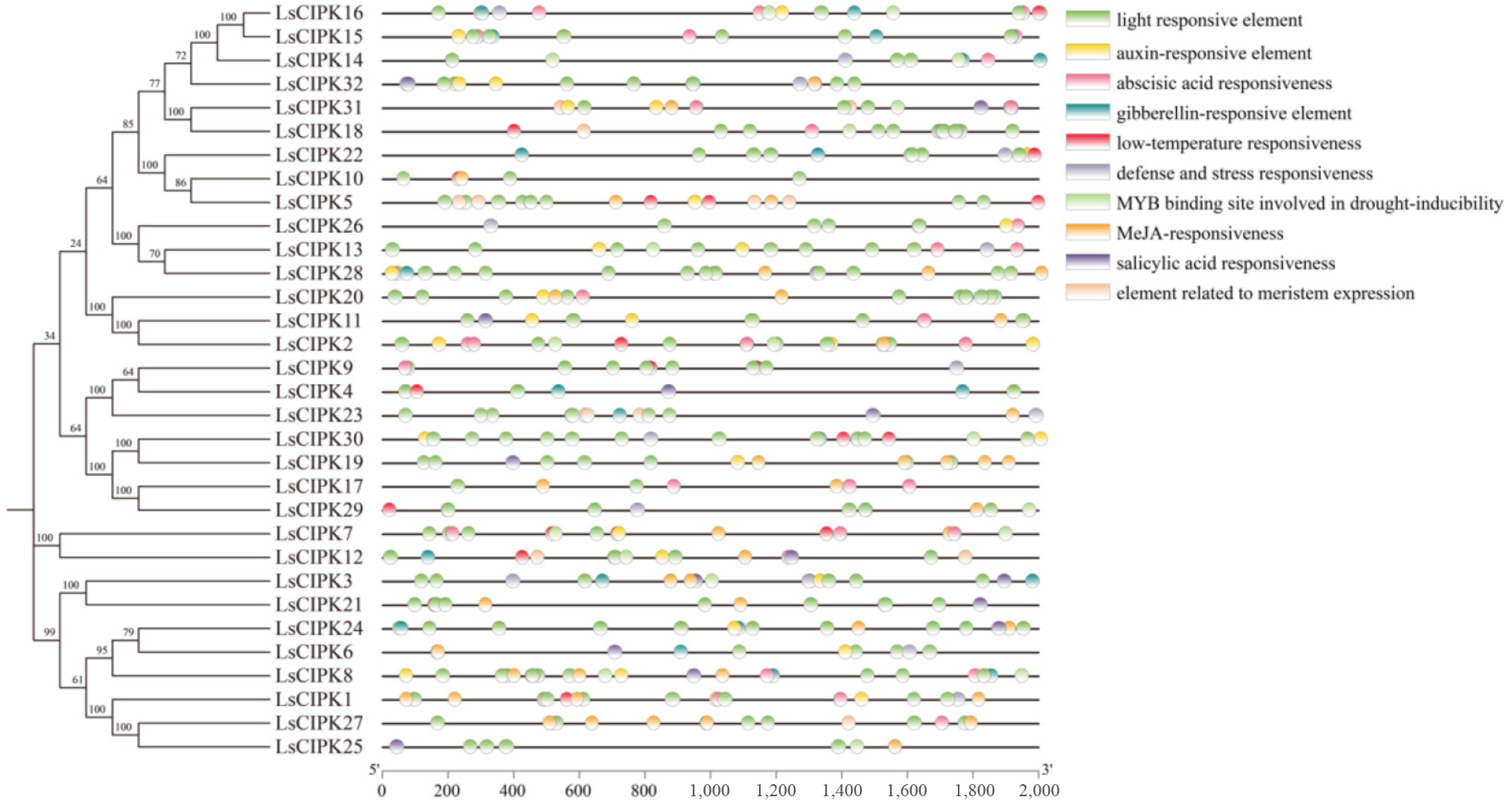
Figure 4.
Analysis of cis-acting elements associated with specific plant hormones and abiotic stresses within 2,000 bp upstream of the LsCIPKs sequence.
Intraspecific, cross-species collinearity analysis and Ka/Ks analysis of the CIPK gene family in lettuce
-
To analyze the expansion pattern and evolutionary characteristics of the CIPK gene family in lettuce, MCScanX was employed to examine their collinearity relationships within the genome (Fig. 5). The results revealed that 32 LsCIPK genes are distributed across nine chromosomes, forming eight pairs of collinear duplicate genes, including LsCIPK2/LsCIPK11, LsCIPK9/LsCIPK22, LsCIPK13/LsCIPK26, LsCIPK13/LsCIPK28, LsCIPK25/LsCIPK27, LsCIPK26/LsCIPK28, LsCIPK31/LsCIPK32, and LsCIPK30/XM_023875268.3. Although the LsCIPK genes exhibit a relatively dispersed distribution across chromosomes, they maintain clear collinearity, indicating that this gene family underwent expansion driven by segmental duplication in lettuce, and possesses high structural conservation.
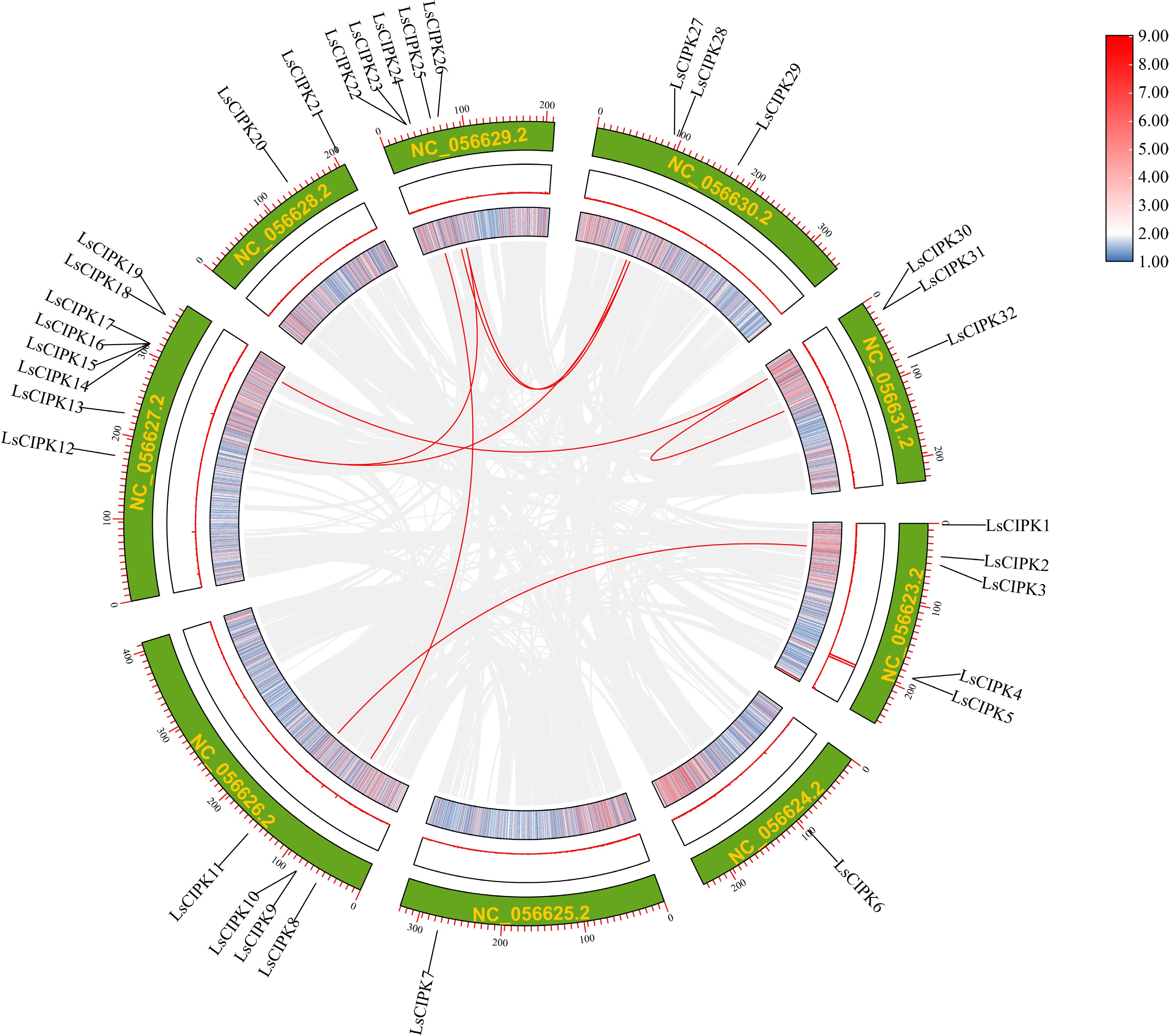
Figure 5.
Genome-wide colinearity analysis of the lettuce CIPK gene family. Arcs of different colors represent homologous gene pairs between chromosomes; the green outer ring depicts the nine chromosomes of lettuce (LsChr1–LsChr9). The heatmap colors (blue, red) indicate a gradient of sequence similarity (from low to high).
To further evaluate the selective pressure following gene duplication, Ka/Ks analysis was performed on the duplicated genes mentioned above (Fig. 6). The Ka/Ks ratios for all gene pairs were less than 1 (0.0588–0.1278), indicating that the LsCIPK gene family as a whole underwent strong purifying selection, with its function remaining stable throughout evolution. Among these, LsCIPK2/LsCIPK11 and LsCIPK25/LsCIPK27 exhibit slightly higher Ka/Ks ratios, suggesting a potential trend toward weak functional differentiation. Conversely, LsCIPK13/LsCIPK26 and LsCIPK26/LsCIPK28 display the lowest Ka/Ks ratios, indicating they were subject to more stringent conservation selection. Combining the results of collinearity analysis and Ka/Ks ratios, we revealed that the expansion of the lettuce CIPK gene family was primarily driven by segmental duplication events. This family has maintained highly conserved molecular characteristics throughout long-term evolution, consistent with the evolutionary patterns observed in CIPK families of plants such as Arabidopsis and rice.
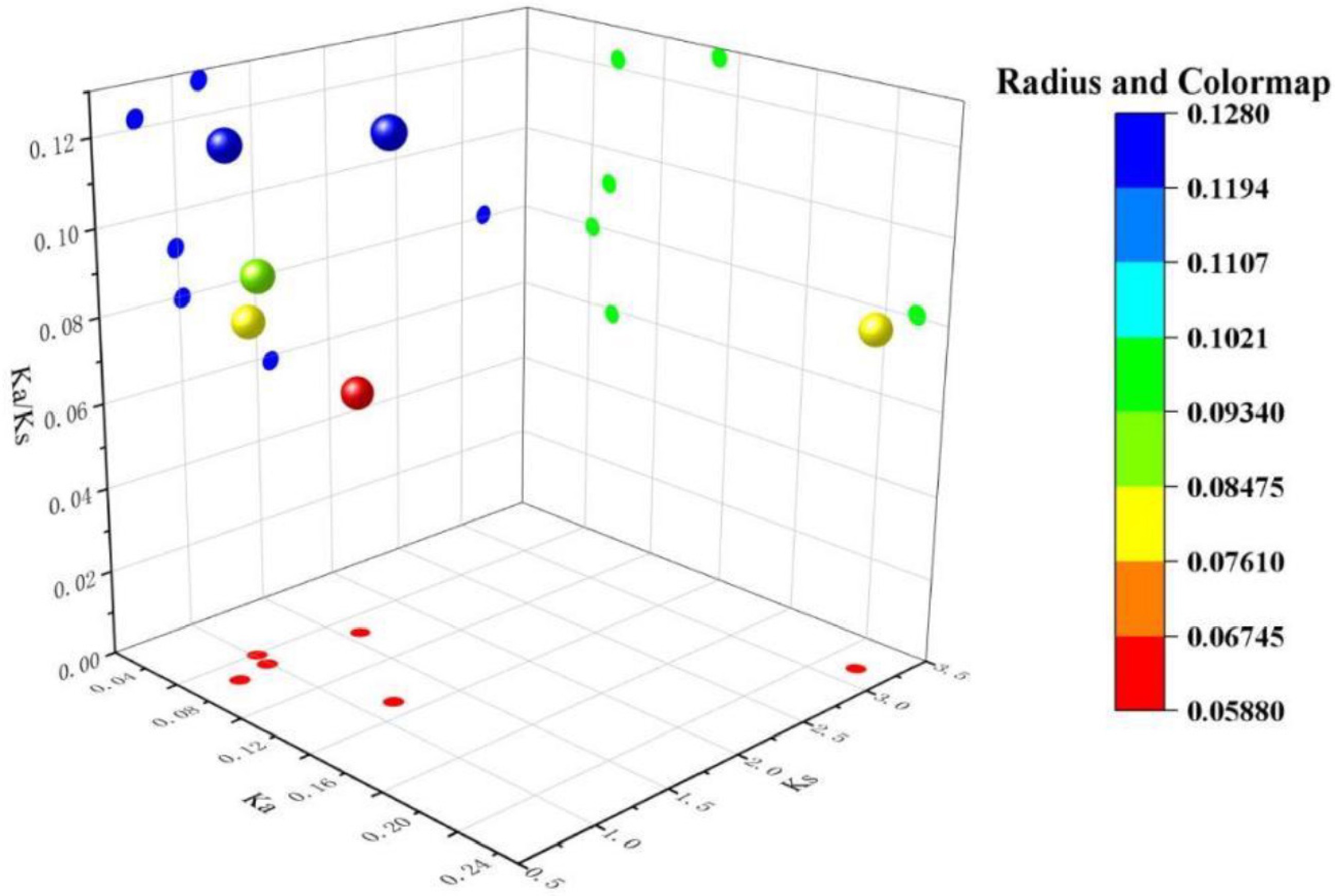
Figure 6.
Analysis of Ka/Ks ratios for collinear genes in the lettuce CIPK gene family. The bubble chart displays the distribution of Ka, Ks, and Ka/Ks values for each gene pair, with color and bubble size indicating the range of Ka/Ks ratios.
To assess the evolutionary conservation of the lettuce CIPK gene family across different plant species, a collinearity relationship map was constructed using MCScanX, between lettuce, Arabidopsis, and rice (Fig. 7). The results revealed abundant collinearity connections between lettuce and Arabidopsis thaliana, primarily involving regions such as AtChr1, AtChr2, AtChr3, AtChr4, AtChr5, and lettuce LsChr1, LsChr4, LsChr5, LsChr6, LsChr7, LsChr8, and LsChr9. This indicated high homology and conservation in the CIPK gene family between the two species. Some LsCIPK genes also form one-to-many or many-to-one correspondences with multiple Arabidopsis CIPK members, reflecting the duplication and retention phenomena within the CIPK family in dicotyledonous plants. In contrast, the number of colinear connections between lettuce and rice was significantly lower, with limited homologous regions detected only on OsChr1, OsChr5, OsChr6, OsChr7, OsChr9, OsChr11, and OsChr12, indicating that following the divergence of monocots and dicots, the CIPK family underwent substantial structural and sequential diversification, resulting in reduced conservation. In summary, the lettuce CIPK gene family exhibits significantly higher conservation within dicotyledons than in homologous relationships with monocotyledons. This suggested that the family had maintained a more consistent evolutionary pattern within the dicotyledonous lineage, whereas monocotyledons had undergone more pronounced rearrangement and diversification.
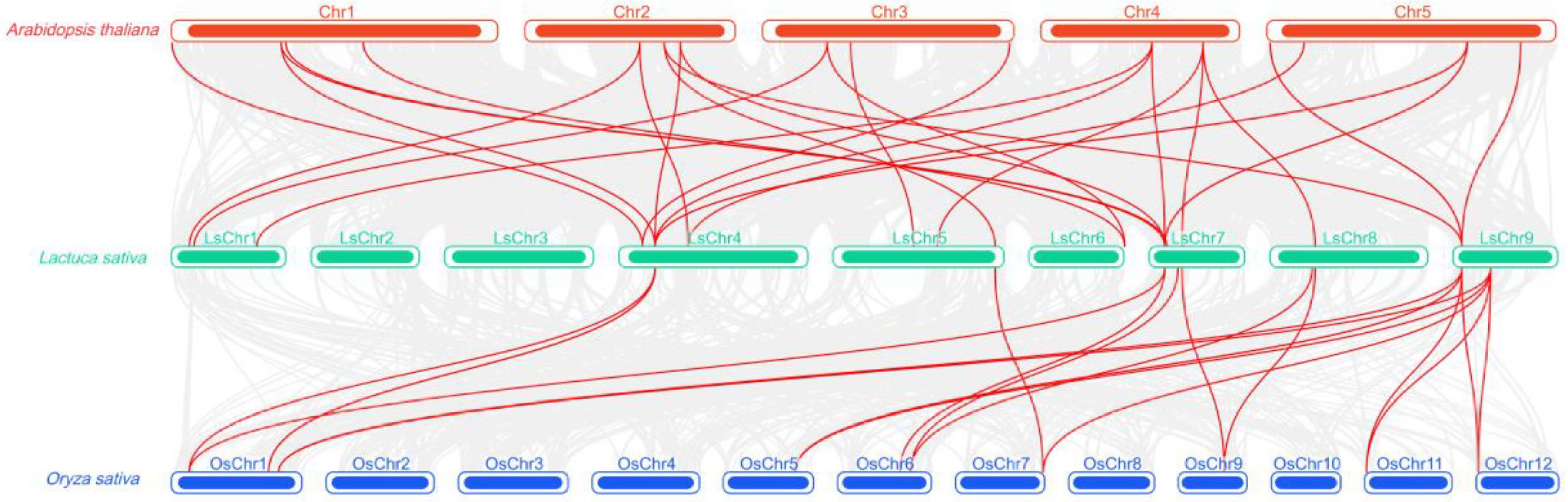
Figure 7.
Cross-species orthology analysis of the CIPK gene families in lettuce, Arabidopsis, and rice. Red lines indicate homologous gene pairs between different species; the top panel shows Arabidopsis chromosomes (red), the middle panel shows lettuce chromosomes (green), and the bottom panel shows rice chromosomes (blue); the density of lines reflects the degree of colinearity among CIPK genes across different species.
Intersection analysis of differentially expressed genes in lettuce under salt and drought stress
-
To compare the transcriptional response patterns of lettuce to different abiotic stresses, an intersection analysis was performed on differentially expressed genes (DEGs) from salt stress (CK vs NaCl) and drought stress (CK vs PEG) treatments (Fig. 8). Results revealed that 4,806 differentially expressed genes (DEGs) were identified under salt stress, including 1,619 salt-specific genes; 4,438 DEGs were identified under drought stress, including 1,251 drought-specific genes, and 3,187 genes exhibited consistent expression changes across both stresses.
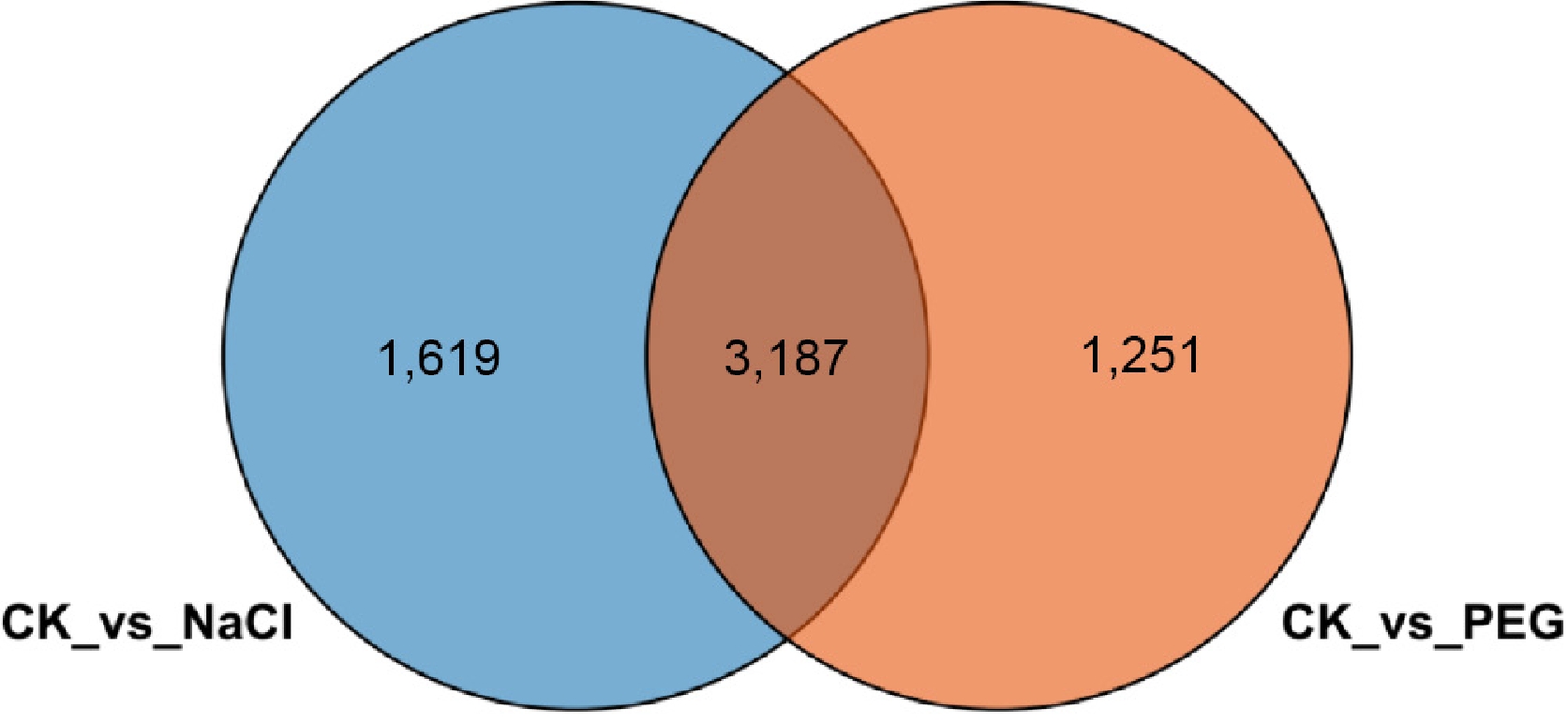
Figure 8.
Venn diagram of differentially expressed genes in lettuce under salt stress and drought stress. Blue and orange circles represent DEGs under salt stress and drought stress, respectively. The overlapping region indicates differentially expressed genes common to both treatments.
GO and KEGG functional enrichment analysis of differentially expressed genes
-
To analyze the functional characteristics of genes specifically responding to salt stress, GO and KEGG enrichment analyses were performed on 1,619 genes differentially expressed in response to NaCl treatment (Fig. 9). GO analysis revealed that these genes were primarily enriched in entries related to transferase activity, kinase activity, transcriptional regulation activity, protein modification processes, plasma membrane, and chloroplast-associated functions (Fig. 9a). This indicated that salt stress significantly impacted signal transduction, membrane structural homeostasis, and photosynthetic organ function. KEGG analysis further revealed that the specific DEGs were significantly enriched in multiple metabolism-related pathways, including metabolic pathways, mitochondrial biogenesis, transport proteins, DNA replication proteins, and amino acid metabolism (Fig. 9b). Additionally, the enrichment of pathways such as ubiquitin-mediated protein degradation and P450 redox reactions indicated that salt stress might also participate in the response by regulating protein turnover and redox balance. In summary, genes specifically responsive to salt stress primarily participate in processes such as signal perception, transmembrane transport, energy and nitrogen-carbon metabolism, protein processing, and redox regulation.
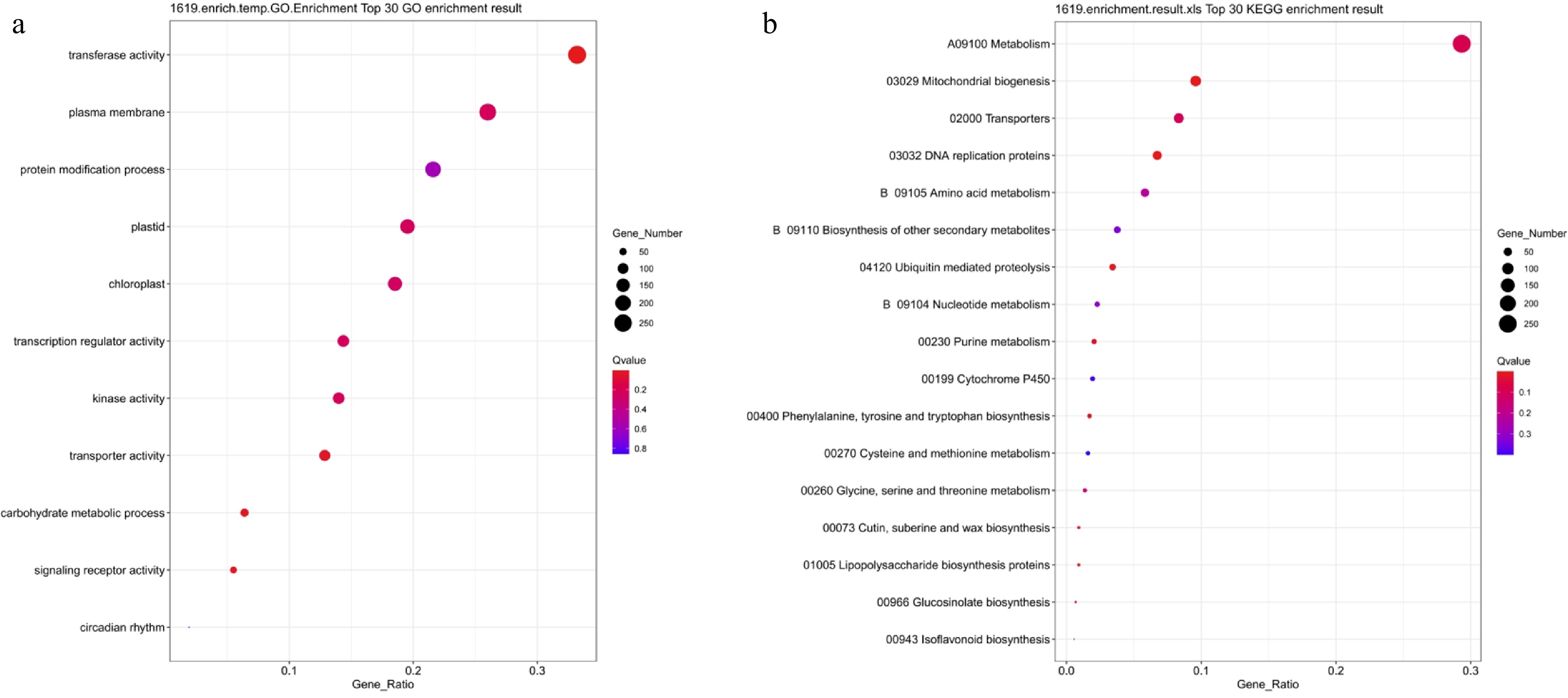
Figure 9.
GO and KEGG functional enrichment analysis of NaCl-treated differentially expressed genes. (a) GO enrichment analysis; (b) KEGG enrichment analysis. Bubble size indicates the number of enriched genes, color represents significance (Q-value), and the horizontal axis shows enrichment factor. A smaller Q-value indicates more significant enrichment.
To elucidate the specific response mechanisms to drought stress, GO and KEGG enrichment analyses were performed on 1,251 PEG-treated differentially expressed genes (Fig. 10). GO analysis revealed that these genes were primarily enriched in entries such as cellular processes, intracellular structures, nucleotide metabolism, hydrolase activity, RNA binding, and mitochondrial-related functions (Fig. 10a). These enriched items indicated that drought stress significantly impacted fundamental cellular metabolic activities and enhanced processes related to energy production, RNA processing, and transcriptional regulation. KEGG analysis revealed that drought-specific DEGs were primarily enriched in pathways, including genetic information processing, chromosome-associated proteins, ribosome biosynthesis, mRNA and tRNA synthesis, and plant–pathogen interactions (Fig. 10b). The activation of these pathways indicated that lettuce might maintain physiological function by regulating transcriptional and translational processes, cellular defense responses, and nucleic acid metabolism under drought stress. In summary, PEG-induced stress-responsive genes are primarily involved in genetic information processing, cellular metabolic regulation, and stress signal response.
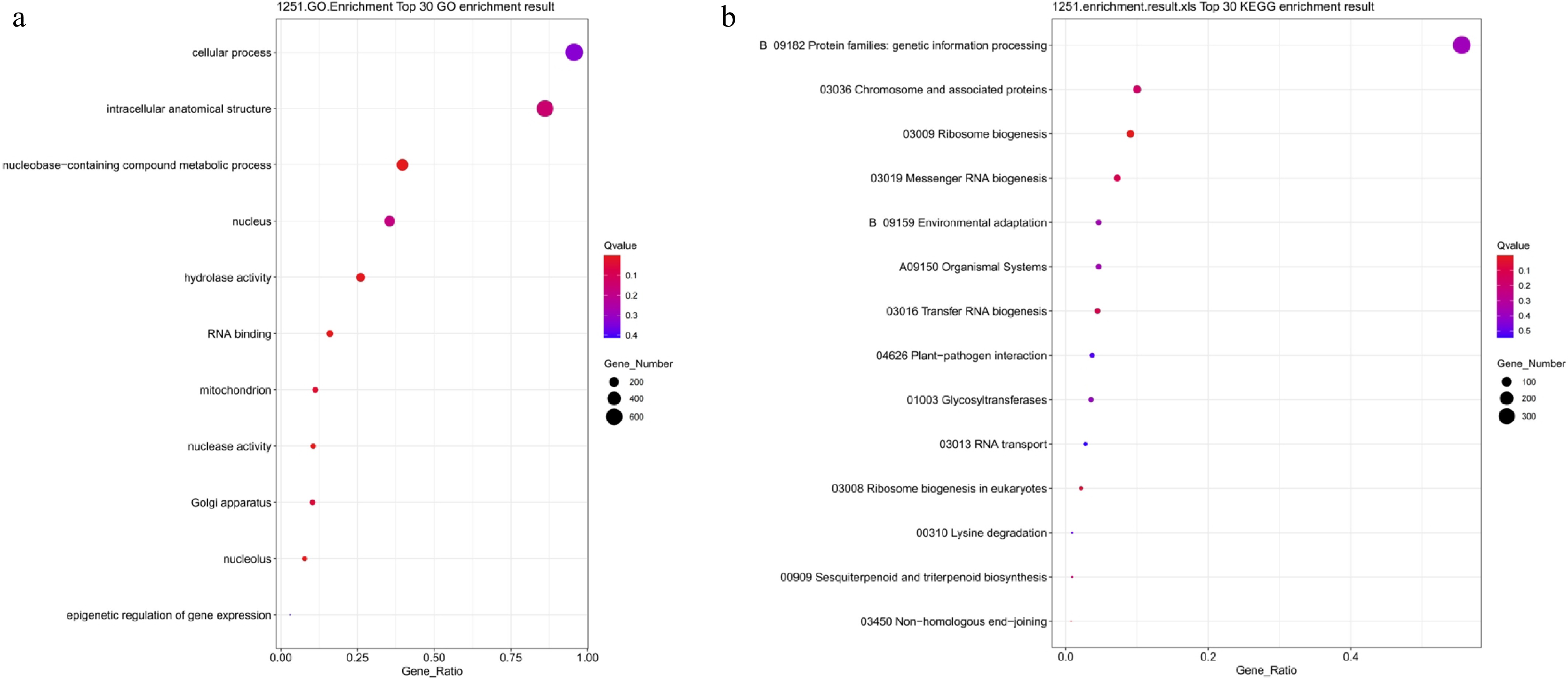
Figure 10.
GO and KEGG functional enrichment analysis of PEG-treated differentially expressed genes. (a) GO enrichment analysis; (b) KEGG enrichment analysis. Bubble size indicates the number of enriched genes, color represents significance (Q-value), and the horizontal axis shows enrichment factor. A smaller Q-value indicates more significant enrichment.
To investigate the common response mechanisms to salt and drought stress, GO and KEGG enrichment analyses were performed on 3,187 co-differentially expressed genes (Fig. 11). GO enrichment analysis revealed that these common DEGs were primarily distributed across entries such as intracellular structures, cytoplasm, nucleotide metabolism, chemical stimulus response, stress response, and abiotic stress response (Fig. 11a). Additionally, it was significantly enriched in functional categories such as chloroplasts, mitochondria, DNA-binding transcription factor activity, and nuclease activity, indicating that both salt and drought stress affected core cellular processes including basal metabolism, energy conversion, and transcriptional regulation. KEGG analysis revealed that these genes were primarily enriched in pathways, including metabolism, signaling and cellular process-related protein families, substance transport, mitochondrial biogenesis, chaperone protein folding, amino acid metabolism, and ribosome biogenesis (Fig. 11b). Concurrently, significant enrichment was observed in pathways related to protein degradation, glutathione metabolism, and photosynthesis, suggesting that these shared DEGs were broadly involved in stress signal regulation, antioxidant defense, and energy balance maintenance. Overall, differentially expressed genes induced by combined salt and drought stress are primarily involved in fundamental processes such as cellular metabolic regulation, redox balance, and energy conversion, reflecting their shared molecular response patterns in lettuce's adaptation to various abiotic stresses.
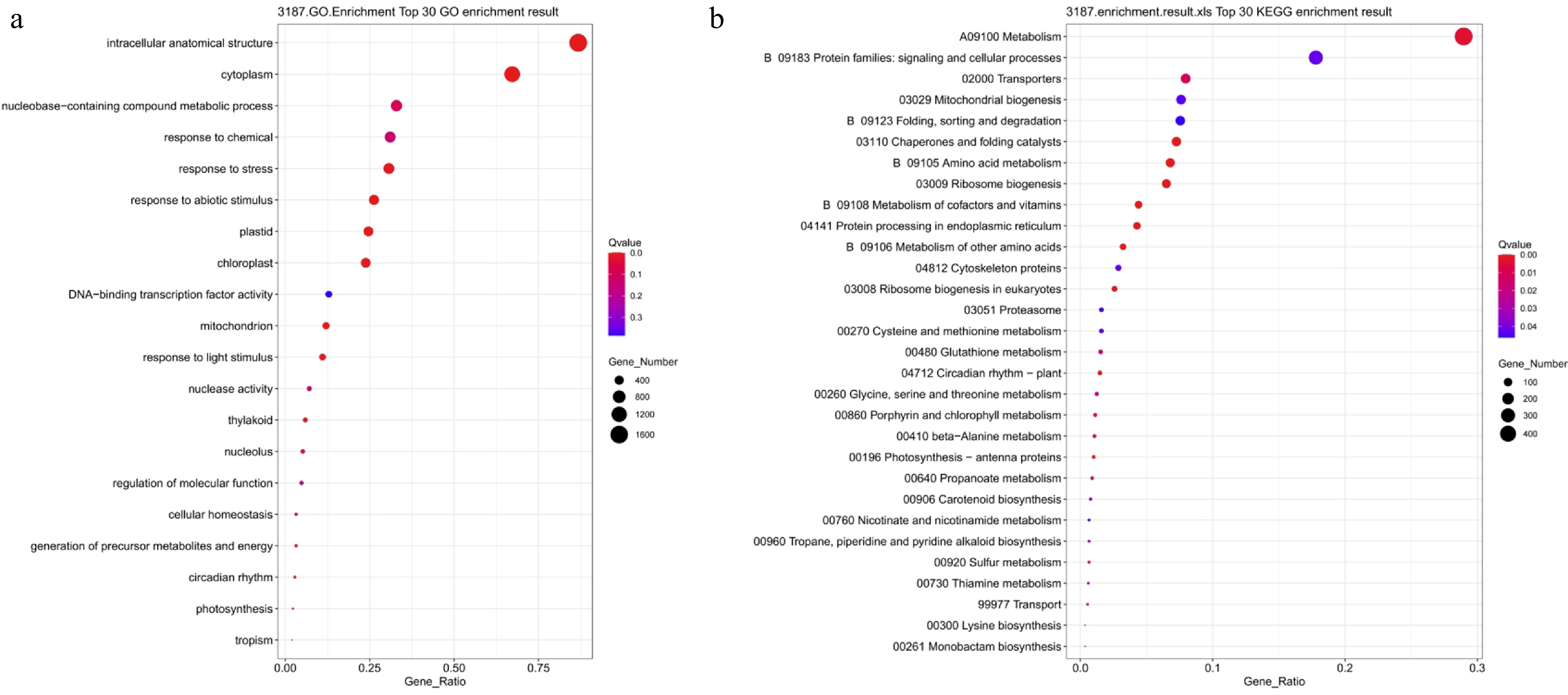
Figure 11.
GO and KEGG functional enrichment analysis of genes differentially expressed in both salt stress and drought stress. (a) GO enrichment analysis; (b) KEGG enrichment analysis. Bubble size indicates the number of enriched genes, color represents significance (Q-value), and the horizontal axis shows enrichment factor. A smaller Q-value indicates more significant enrichment.
Expression analysis of the lettuce CIPKs under salt and drought stress
-
To evaluate the expression patterns of the lettuce CIPK genes under abiotic stress, transcriptomic data was analyzed to examine the expression levels of 32 LsCIPKs under salt stress (NaCl) and drought stress (PEG) treatments (Fig. 12). The results indicated that the LsCIPK gene family exhibited extensive expression changes under both stress conditions. The expression level of multiple members (such as LsCIPK2, LsCIPK7, LsCIPK8, LsCIPK26, LsCIPK27, and LsCIPK32) were upregulated under both NaCl and PEG treatments. Among them, the expression of LsCIPK26 and LsCIPK28 was upregulated significantly. Another group of members (such as LsCIPK5, LsCIPK6, LsCIPK13, LsCIPK23, LsCIPK25, and LsCIPK29) showed consistent downregulation under both treatment conditions. Additionally, certain genes exhibited differential responses under different stress conditions. For instance, the expression patterns of LsCIPK5, LsCIPK9, and LsCIPK16 were inconsistent under salt and drought stresses.
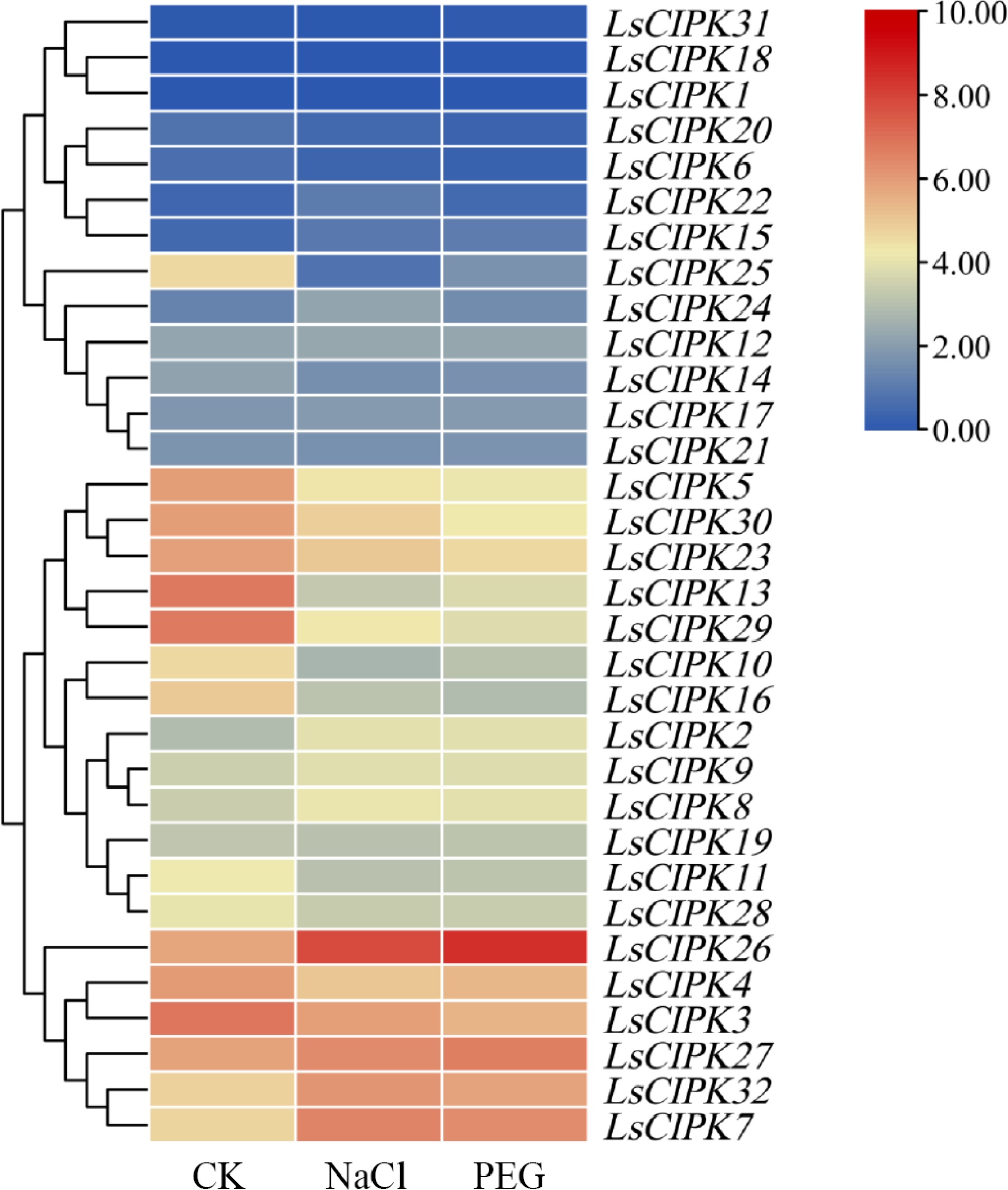
Figure 12.
Heatmap of expression patterns of the LsCIPK genes under salt stress (NaCl) and drought stress (PEG) treatments. The color bars in the heatmap represent expression levels converted to log2 (log2[TPM + 1]), with values ranging from 0 to 10: dark blue indicates lower expression, while dark red indicates higher expression. CK denotes the control group, NaCl represents the salt stress treatment group, and PEG indicates the drought stress treatment group.
qRT-PCR validation of the LsCIPKs under salt stress and drought stress
-
To validate the expression patterns detected by transcriptomics, nine LsCIPK genes (LsCIPK7, LsCIPK12, LsCIPK13, LsCIPK19, LsCIPK21, LsCIPK25, LsCIPK26, LsCIPK29, and LsCIPK32) were selected for qRT-PCR analysis to examine their expression under salt stress (NaCl) and drought stress (PEG) treatments (Fig. 13). The results indicated that gene expression responses to the two types of abiotic stress varied significantly. Among them, LsCIPK7, LsCIPK26, and LsCIPK32 showed significant upregulation under both NaCl and PEG treatments, consistent with the transcriptome results. The expression levels of LsCIPK12, LsCIPK19, and LsCIPK21 showed minimal variation under both stress conditions, remaining relatively stable overall. LsCIPK13, LsCIPK25, and LsCIPK29 were all downregulated under both NaCl and PEG treatment conditions. qRT-PCR results showed high consistency with transcriptomic data.
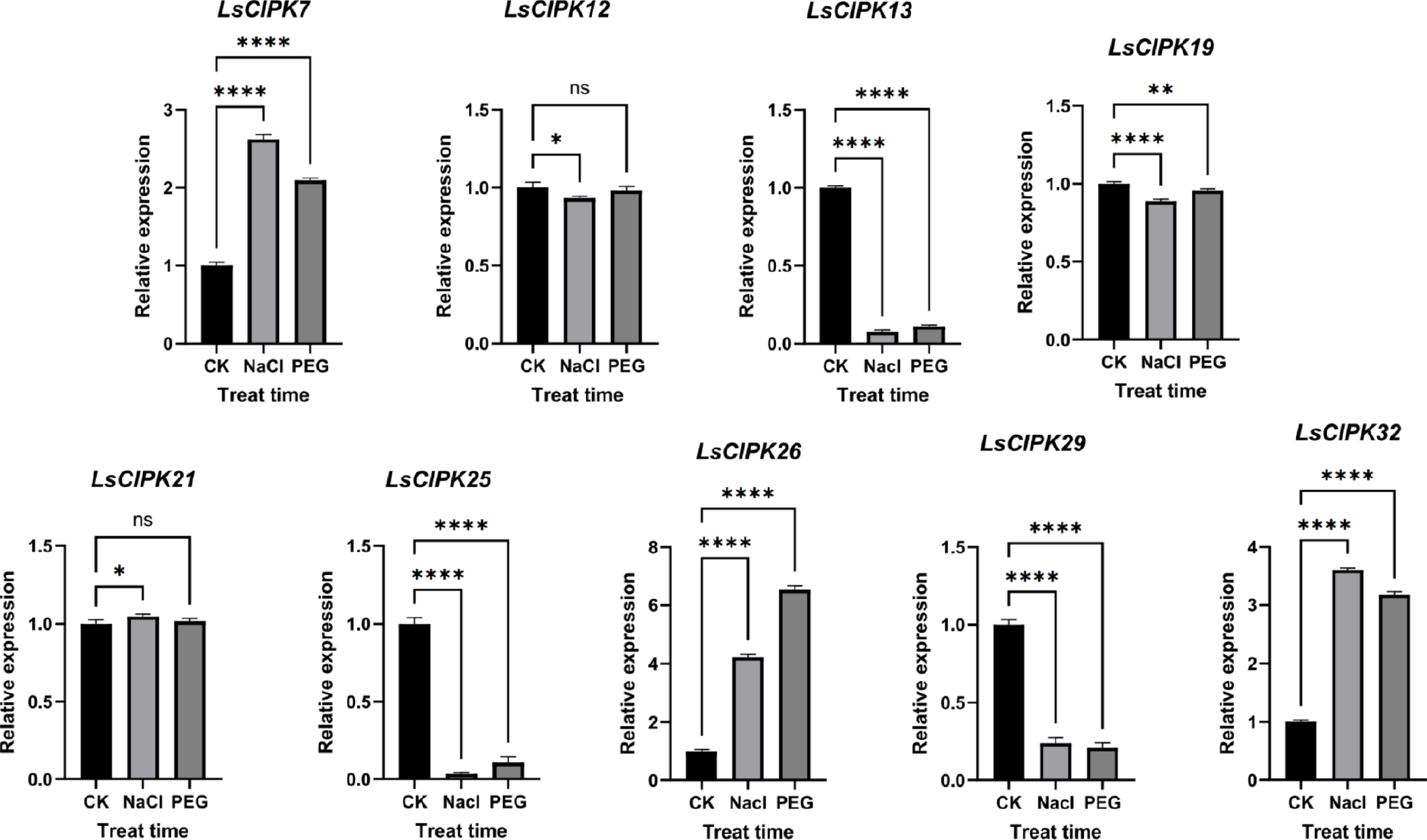
Figure 13.
qRT-PCR expression analysis of the lettuce CIPK genes under salt (NaCl), and drought (PEG) stress. CK represents the control group, NaCl denotes the salt stress treatment group, and PEG indicates the drought stress treatment group; error bars represent standard errors (n = 5); ns indicates p > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
Subcellular localization of LsCIPK32 in lettuce
-
To determine the intracellular localization of LsCIPK proteins, a 35S:LsCIPK32-GFP fusion expression vector was constructed using LsCIPK32 as a representative protein. This vector was transiently transformed into tobacco epidermal cells for fluorescence observation, with 35S:GFP serving as the empty vector control (Fig. 14). Confocal microscopy observations revealed that the empty vector GFP exhibited a freely diffusing distribution within cells, with fluorescent signals detectable in both the cytoplasm and the nucleus. The fluorescence signal of LsCIPK32-GFP was predominantly localized to the nucleus, with distinct membrane-bound distribution observed in the plasma membrane region. No widespread cytoplasmic diffusion was detected. These results indicated that the LsCIPK32 protein exhibited dual localization to both the nucleus and plasma membrane, consistent with its potential signaling function.
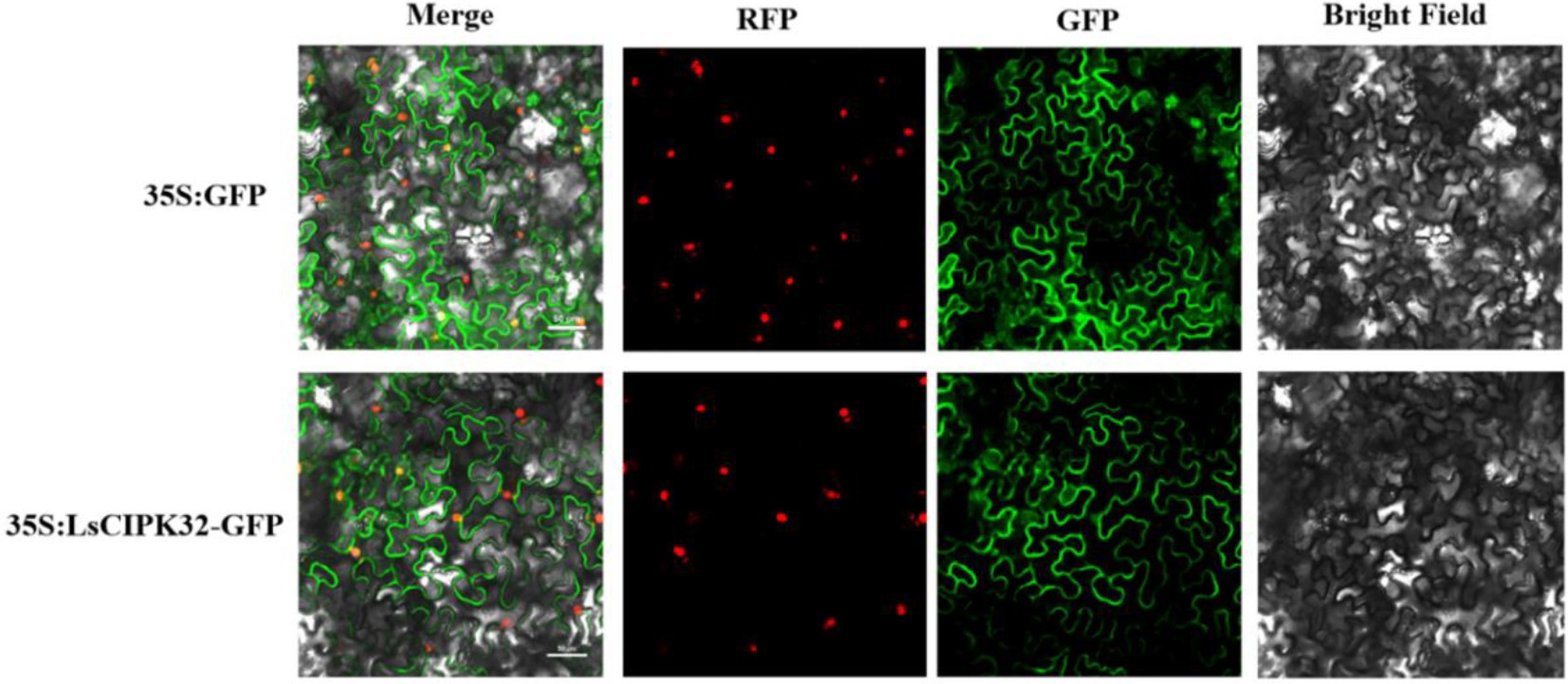
Figure 14.
Subcellular localization analysis of the LsCIPK32 protein. Merge, RFP, GFP, and Bright Field represent signal merging, red fluorescence channel, green fluorescence channel, and bright field image, respectively. Scale bar = 50 μm.
-
The CBL–CIPK signaling system, as a core regulatory module mediating Ca2+-dependent stress responses in plants, plays a pivotal role under salt stress, drought stress, and various environmental stimuli[50]. Research indicated that CIPK proteins regulated various adaptive mechanisms, including ion homeostasis, osmotic regulation, and redox balance, by forming complexes with CBL proteins, thereby enhancing plant tolerance to adverse environmental conditions[2]. In this study, 32 CIPK genes were systematically identified in the lettuce genome, a number comparable to those in Arabidopsis[24], rice[25], tomato[29], potato[30], and pepper[31], indicating high conservation of the CIPK family across dicotyledonous plants. Chromosomal mapping results indicated that LsCIPK genes exhibited an uneven distribution across nine chromosomes, with some members clustered together, suggesting potential tandem duplication or segmental duplication events. This expansion pattern aligns with the formation mechanism of CIPK gene families in crops such as Chinese cabbage[26], cotton[27], and grape[28], suggesting that segmental duplication is a common evolutionary mechanism in dicotyledonous plants.
Phylogenetic analysis revealed that the lettuce CIPK family can be divided into five major branches, with clear correspondence between its members and the CIPKs members of Arabidopsis, indicating that the gene family had maintained high sequence conservation during evolution, consistent with previous findings in grape[28], tomato[29], and pecan[33]. Further conservative domain analysis indicated that all LsCIPK proteins contained the characteristic PKc-like and NAF domains, which constituted the structural basis for CIPK protein interaction with CBL[4,5,24]. Additionally, gene structures exhibited distinct 'high/low intron' patterns, a phenomenon also observed in species such as rice[25], potato[30], and quinoa[20], which suggested that CIPK genes with different structural categories might perform distinct regulatory functions in lettuce as well.
Analysis of cis-acting elements revealed that most of the LsCIPK gene promoter regions contained abundant light-responsive, hormone-responsive, and stress-responsive elements, particularly those associated with ABA, MeJA, and drought/cold stress responses. This feature was highly consistent with the regulatory functions of the CIPK family in the ABA signaling pathway, osmotic stress, and light signaling[17−22]. In Arabidopsis, CIPK1 participates in regulating drought responses by phosphorylating the ABA receptor PYL4[19]; in chili peppers, CIPK genes have also been demonstrated to participate in ABA-dependent stress signaling pathways[22]. Therefore, it is hypothesized that the lettuce CIPK gene might participate in stress regulation within a multi-hormone network, but its specific role required further functional validation.
Genome-wide synteny analysis revealed that the expansion of the lettuce CIPK gene family was primarily driven by segmental duplication. Furthermore, the Ka/Ks ratios for all eight duplicated gene pairs was less than 1, indicating that this family was under intense purifying selection, which was highly consistent with research on some crops, such as Chinese cabbage[26], cotton[27], and turnips[32], indicating that CIPK genes, as core members of signaling pathways, have remained highly conserved throughout plant evolution.
Based on transcriptome expression analysis, most LsCIPK genes showed significant upregulation or downregulation under salt and drought stress, indicating that these genes might be extensively involved in the regulation of stress signaling. Among these, LsCIPK7, LsCIPK26, and LsCIPK32 were significantly upregulated under both types of stress, consistent with previously reported positively regulated CIPK members in species, such as quinoa[20], potato[30], and cassava[17]. However, LsCIPK13, LsCIPK25, and LsCIPK29 exhibited sustained downregulation under both stress conditions, suggesting these genes might function as negative regulators in lettuce stress response pathways, which was analogous to the negative regulatory function of AtCIPK1 in Arabidopsis under drought stress[19]. qRT-PCR results showed high consistency with transcriptomic data, further validating the reliability of the expression patterns.
Additionally, the expression of LsCIPK32 was upregulated under both NaCl and PEG treatments based on qRT-PCR and transcriptomic data. To further clarify its function, subcellular localization of LsCIPK32 was detected, and the results indicated that LsCIPK32 is primarily distributed in the nucleus and cytoplasm, consistent with the functional mode of most CIPKs that regulate SOS1 or other ion transporters through membrane localization[8−11]. In various plant species, CIPK membrane localization typically indicate involvement in ion efflux, osmotic regulation, or membrane signaling[7−11,31]. Therefore, LsCIPK32 might play a crucial role in salt stress and drought responses in lettuce.
In summary, the composition, structural characteristics, evolutionary relationships, and expression patterns of lettuce CIPKs under salt and drought stress was systematically analyzed. Subsequently, CRISPR/Cas9 and overexpression materials would be taken to validate the functions of LsCIPK genes, such as LsCIPK7, LsCIPK26, and LsCIPK32, which would further elucidate the regulatory network of the CBL-CIPK complex in lettuce stress responses, providing theoretical foundations and candidate gene resources for molecular breeding of stress-tolerant lettuce.
-
In this study, 32 LsCIPK genes were identified in the lettuce genome, and their chromosomal distribution, structural characteristics, phylogenetic relationships, and duplication patterns were systematically analyzed. Collinearity and Ka/Ks analysis indicated that the expansion of this family was primarily driven by segmental duplication, and had maintained high conservation over long term evolution. Cis-element analysis revealed that LsCIPKs were broadly involved in light signaling, hormone regulation, and stress responses. Transcriptome and qRT-PCR results indicated that multiple genes (such as LsCIPK7, LsCIPK26, and LsCIPK32) were significantly induced under salt and drought stress, suggesting their potential key roles in stress responses. Subcellular localization revealed that LsCIPK32s were distributed in both the nucleus and cytoplasm. This study lays the foundation for functional research on the CIPK genes in lettuce and provides important genetic resources for subsequent analysis of stress signaling networks, and the development of stress-tolerant lettuce breeding programs.
-
The authors confirm contribution to the paper as follows: study conception and design, analysis and interpretation of results: Li J, Li K, Li Z; data collection: Li J, Chu W, Zhao Q; draft manuscript preparation: Wu R, Yin L. All authors reviewed the results and approved the final version of the manuscript.
-
The sequence data for the expression pattern analysis of LsCIPK genes in response to salt or drought conditions is available at NCBI: PRJNA1367680.
-
This study was supported by the Handan Science and Technology Research and Development Plan Project (21422012321).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jiawei Li, Zixuan Li, Ke Li
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li J, Li Z, Li K, Chu W, Zhao Q, et al. 2026. Genome-wide identification of the CIPK gene family in lettuce and expression analysis under salt and drought stress. Vegetable Research 6: e012 doi: 10.48130/vegres-0026-0005 

Genome-wide identification of the CIPK gene family in lettuce and expression analysis under salt and drought stress
- Received: 13 December 2025
- Revised: 22 January 2026
- Accepted: 04 February 2026
- Published online: 31 March 2026
Abstract: CBL-interacting protein kinases (CIPKs) serve as key regulators in plant Ca2+ signaling pathways, playing a vital role in responding to abiotic stresses such as salinity and drought. However, systematic studies on the CIPK gene family in lettuce (Lactuca sativa L.) remain scarce. In this study, comprehensive identification, structural analysis, evolutionary analysis, and stress response analysis of the CIPK gene family were conducted based on the lettuce reference genome. A total of 32 LsCIPK genes were identified, which were distributed on nine chromosomes. Phylogenetic analysis demonstrated that 32 LsCIPKs could be divided into five evolutionary branches. All LsCIPK proteins contained a typical protein kinase domain and an NAF domain (CBL-interacting domain). Promoter cis-element analysis indicated that most LsCIPK genes are rich in cis-elements associated with light response, hormone response, and abiotic stress. Collinearity analysis indicates that the expansion of this gene family was primarily driven by segmental duplication, and the Ka/Ks ratios for all eight duplicate gene pairs were less than 1, suggesting that the family as a whole is under purifying selection. Expression analysis revealed significant expression changes in multiple LsCIPK genes. Among them, the expression of LsCIPK7, LsCIPK26, and LsCIPK32 was strongly induced. Further qRT-PCR results confirmed the transcriptome trends. Subcellular localization analysis of LsCIPK32 revealed that this protein is primarily distributed in the nucleus and cell membrane. This study provides candidate genes and theoretical support for deciphering the regulatory mechanisms of the lettuce CBL-CIPK signaling pathway and for breeding salt-tolerant and drought-resistant lettuce varieties.
-
Key words:
- Lettuce /
- CIPK gene family /
- Abiotic stress /
- Genome-wide identification /
- Transcriptome analysis