










-
Graves' ophthalmopathy (GO) is an autoimmune inflammatory disorder characterized by orbital tissue remodeling, including extraocular muscle swelling and orbital fat expansion[1]. As the most prevalent extrathyroidal manifestation of Graves' disease (GD), GO affects 25%–50% of GD patients and significantly complicates the clinical management of autoimmune thyroid dysfunction[2]. Clinically, GO presents with debilitating symptoms such as lid retraction, proptosis, and diplopia, which frequently lead to marked facial disfigurement and subsequent social stigmatization or withdrawal[2,3]. In severe cases, the condition can escalate to sight-threatening dysthyroid optic neuropathy (DON), resulting in irreversible vision loss and imposing an immense psychological and socioeconomic burden on both individuals and society[4].
Currently, the management of moderate to severe GO relies heavily on glucocorticoids, yet this approach is often hampered by therapeutic resistance and systemic toxicity[5]. Although the advent of novel biological agents, such as the insulin growth factor 1 receptor (IGF-1R) inhibitor teprotumumab, has provided a potent alternative for reducing proptosis, their clinical application is constrained by potential adverse events, including hearing impairment, muscle spasms, and hyperglycemia[6]. Emerging evidence suggests that the pathogenesis of GO is driven by intricate immune-inflammatory cascades, where various inflammatory proteins serve as critical mediators in the transition from early active inflammation to chronic fibrosis[7]. Consequently, elucidating the specific roles of these inflammatory mediators is crucial for clarifying the molecular mechanisms underlying GO's pathogenesis and may further inform the development of targeted therapeutic strategies and improved clinical management[7].
Thyrotropin receptor autoantibody (TRAb) is an autoantibody targeting the thyrotropin receptor (TSHR)[8]. Mechanistically, the binding of TSAb to the TSHR, which frequently cross-talks with the IGF-1R on orbital fibroblasts, serves as the primary trigger for orbital pathology[9]. Crucially, rather than causing isolated structural changes directly, this receptor's activation first orchestrates a profound local immune response, characterized by the recruitment of immune cells and the robust secretion of various inflammatory cytokines and chemokines[10]. It is this intricate inflammatory cascade that subsequently acts as the core mediator, driving the proliferation of orbital fibroblasts, adipogenesis, and the excessive accumulation of hyaluronic acid, ultimately leading to extraocular muscle enlargement, orbital fat expansion, and chronic fibrosis[11,12]. Given its role in initiating this process, TRAb has become an indispensable biomarker for diagnosing GO[8]. Beyond diagnosis, serum TRAb titers strongly correlate with the clinical activity score (CAS), serving as a valuable predictor for disease progression and therapeutic outcomes[13]. Identifying the key downstream inflammatory mediators that translate the causal pathway from TRAb to GO is essential for unraveling this complex pathogenic cascade, clarifying disease mechanisms, and discovering novel, precise therapeutic targets.
Inflammatory proteins, encompassing a diverse array of cytokines and chemokines, serve as essential effectors in the immunopathogenesis of GO by driving orbital fibroblast activation, adipogenesis, and tissue remodeling[10,14]. The aberrant production of these inflammatory mediators is intricately driven by upstream autoimmune triggers, most notably TRAb. Indeed, serum TRAb titers exhibit a strong positive correlation with both local and systemic proinflammatory cytokine profiles, acting as a quantitative indicator of pathological inflammation in GO[14]. For example, interleukins (ILs) such as IL-17, alongside Th1-chemokines like CXCL10, are significantly upregulated in the serum and orbital tissues of GO patients, orchestrating immune cell infiltration and extracellular matrix expansion[15−17]. These observations confirm that a multitude of specific inflammatory factors actively participate in and propel the disease cascade. Furthermore, the therapeutic success of the IL-6 receptor antagonist tocilizumab has demonstrated immense potential in managing active GO[5,18,19]. This targeted therapeutic efficacy robustly corroborates the pathogenic centrality of the inflammatory network in GO development. Despite these advances, the inflammatory cascade in GO is highly complex and dynamic. Traditional observational studies are inherently limited by potential confounders and the high risk of reverse causation. Consequently, a critical knowledge gap persists regarding which of the multitude of dysregulated inflammatory markers are genuine causal mediators lying directly on the pathway from TRAb elevation to the development of GO, rather than merely downstream byproducts of the disease state. Resolving this causal dilemma is imperative for unraveling the precise pathogenic mechanisms of GO and identifying novel, highly specific therapeutic targets.
To overcome the inherent limitations of traditional observational designs, Mendelian randomization (MR) is widely utilized as a robust epidemiological approach for causal inference[20,21]. By using randomly allocated genetic variants as instrumental variables, MR effectively minimizes environmental confounding and eliminates reverse causation, serving as a natural randomized controlled trial to establish reliable causal relationships. This study aimed to systematically investigate the mediating roles of specific circulating inflammatory proteins in the causal pathway between TRAb and GO using a comprehensive mediation MR framework. Elucidating these critical mediators will contribute to a better understanding of the complex immunopathogenesis of GO.
-
In MR analyses, single-nucleotide polymorphisms (SNPs) are commonly used as instrumental variables (IVs) to explore potential causal relationships between exposures, such as TRAb, and clinical outcomes (e.g., GO). The validity of this approach relies on three core assumptions: (1) The selected IVs must show a robust association with the exposure of interest, (2) the IVs should be independent of any confounders that may bias the exposure–outcome relationship, and (3) the IVs must affect the outcome exclusively through their influence on the exposure, with no alternative biological pathways involved[22,23].
From a causal inference perspective, the impact of an exposure on an outcome can be decomposed into direct and mediated components. The total effect represents the overall influence of the exposure on the outcome, whereas the indirect effect captures the portion of this influence that operates through the intermediary variable[24]. In the present study, TRAb was treated as the primary exposure and GO as the outcome of interest, whereas circulating inflammatory proteins were considered as potential mediators within this causal framework.
To investigate the roles of inflammatory proteins in the effect of TRAb on GO, we applied a two-step MR analysis, which involved sequentially estimating: (1) The causal effect of TRAb on GO, (2) the causal associations between circulating inflammatory proteins and GO, and (3) the causal influence of TRAb on circulating inflammatory protein levels. The indirect (mediated) effect was then quantified using the product of coefficients method, allowing us to delineate the specific contribution of inflammatory proteins within the overall causal pathway linking TRAb to GO[25], thereby providing further insight into the pathogenic processes involved.
All data utilized in this analysis were derived from previously published studies in which informed consent had been obtained from all participants. As a result, no additional ethical approval was required for the present study. All procedures were conducted in accordance with the principles outlined in the Declaration of Helsinki.
Data sources
-
The genome-wide association study (GWAS) summary statistics for TRAb were derived from the most recent release of the FinnGen project (
https://labvalues.finngen.fi/ ) and comprised a total of 3,690 participants. These data were generated from routine clinical laboratory measurements collected as part of standard healthcare practice across Finland, encompassing both public and private medical services. All laboratory records were systematically aggregated within the national KANTA health information system. Prior to incorporation into the FinnGen resource, all laboratory data underwent rigorous quality control procedures conducted by the FinnGen research team to ensure accuracy and reliability. Furthermore, the laboratory measurements were standardized and harmonized according to international standards ((e.g., observational medical outcomes partnership), thereby enabling consistent integration and efficient use of the data for downstream research[26].GWAS summary-level data for circulating inflammatory proteins were obtained from a large meta-analysis that integrated 11 independent cohorts, comprising 14,824 individuals of European descent (
www.ebi.ac.uk/gwas/publications/37563310 )[27]. Using paired genotype data and plasma proteomic measurements generated with the Olink Target platform, the study ultimately provided genetic association results for 91 circulating inflammatory proteins[27]. For GO, the summary statistics were collected from two major GWAS resources to enable cross-dataset discovery and validation. Specifically, data were drawn from the VA Million Veteran Program (710 cases and 449,706 controls) (www.ebi.ac.uk/gwas/studies/GCST90477300 ) and UK Biobank (138 cases and 391,429 controls) (www.ebi.ac.uk/gwas/studies/GCST90435691 )[28,29]. All GWAS datasets were restricted to participants of European ancestry, and no sample overlap was known between the sources.Statistical analysis
-
To infer causality across the TRAb–inflammatory protein–GO axis, we first implemented two-sample MR analyses. The detailed criteria and procedures used for selecting instrumental variables are described in the Supplementary File 1[20,23,30−35]. When an exposure was instrumented by a single genetic variant, causal inference was performed using the Wald ratio approach. This method is specifically suited to single-instrument scenarios, as it estimates the causal effect by taking the ratio of the genetic association with the outcome to that with the exposure, yielding a direct and interpretable effect estimate[24]. In cases where two or more independent SNPs were available as instruments, causal effects were estimated using the inverse variance weighted (IVW) method. This approach integrates effect estimates across multiple genetic variants within a meta-analysis framework. By leveraging information from multiple instruments, the IVW method increases statistical efficiency and improves the robustness of causal effect estimation[36]. Cochran's Q statistic was derived under a fixed-effects IVW framework to assess heterogeneity among the causal estimates. A p-value below 0.05 was taken as evidence of significant heterogeneity[37]. If such heterogeneity was observed, the analysis was repeated using a multiplicative random effects IVW approach to appropriately account for this variability[20].
To evaluate whether circulating inflammatory proteins mediate the association between TRAb and GO, we first estimated the overall causal effect of TRAb on GO (β overall). We then separately derived the causal effect of TRAb on circulating inflammatory protein levels (β1), as well as the causal effect of circulating inflammatory proteins on GO (β2). The indirect (mediated) effect was calculated as the product of these two coefficients (β1 × β2), representing the component of the association transmitted through inflammatory proteins[25]. To further characterize the extent of mediation, we calculated the proportion of mediation by dividing the indirect effect by the total effect (β1 × β2 / β overall). This metric quantifies the fraction of the overall TRAb–GO relationship that can be attributed to the inflammatory protein-mediated pathway[25]. Statistical testing of the mediation effect was performed using a product of coefficients approach implemented in the Interactive Mediation Tests platform (
https://quantpsy.org/sobel/sobel.htm ), which is based on the Sobel test's methodology. The MR-Egger method evaluates whether the regression intercept deviates from zero, which helps identify and measure directional pleiotropy[20]. When evidence of pleiotropy is present, radial plots and radial regression are subsequently used to detect and exclude variants that behave as pleiotropic outliers[38]. After removing these outliers, the mediation analysis is recalculated; if pleiotropy persists, the results are considered to be unreliable and are excluded. To further explore the biological context of the identified mediating inflammatory protein, we constructed an interaction network using GeneMANIA. GeneMANIA integrates multiple evidence sources to infer functionally related genes/proteins, supporting functional interpretation and hypothesis generation regarding potential disease-related mechanisms[39]. -
In the MR analyses, the IVW method was applied as the primary analytical strategy. The results provided evidence of a significant causal relationship between TRAb levels and the risk of GO. Specifically, analyses based on the VA Million Veteran Program dataset indicated that genetically predicted TRAb levels were associated with a markedly elevated risk of GO (odds ratio [OR] = 2.42, p = 0.008). Consistent results were observed in the UK Biobank cohort, where higher genetically inferred TRAb levels were similarly linked to an increased likelihood of GO (OR = 7.96, p = 0.005) (Table 1).
Table 1. Association between genetically predicted thyrotropin receptor antibody and the risk of Graves' ophthalmopathy.
Exposure Outcome study Method Number of SNPs OR (95% CI) p-Value Thyrotropin receptor antibody VA Million Veteran Program Inverse variance weighted 43 2.42 (1.26–4.66) 0.008 Thyrotropin receptor antibody UK Biobank Inverse variance weighted 33 7.96 (1.85–34.28) 0.005 SNP, single-nucleotide polymorphism; OR, odds ratio; CI, confidence interval. Causal effects of inflammatory proteins on GO
-
In the VA Million Veteran Program dataset, we identified that 19 out of 91 circulating inflammatory proteins were significantly causally associated with GO (p < 0.05). After false discovery rate (FDR) correction, six circulating inflammatory proteins remained significant (pFDR < 0.05). The results showed that the genetically predicted higher fibroblast growth factor 21 levels (OR: 1.60; p < 0.001; pFDR = 0.002), fractalkine levels (OR: 1.44; p = 0.002; pFDR = 0.038), and monocyte chemoattractant protein 2 levels (OR: 1.14; p = 0.003; pFDR = 0.048) showed a positive association with GO, suggesting that genetically predicted higher levels of these proteins are associated with an increased risk of GO. In contrast, GO was inversely associated with C-C motif chemokine 25 levels (OR: 0.85; p < 0.001; pFDR = 0.006), C-X-C motif chemokine 1 levels (OR: 0.70; p = 0.001; pFDR = 0.012), C-X-C motif chemokine 5 levels (OR: 0.76; p < 0.001; pFDR = 0.006) (Fig. 1a).
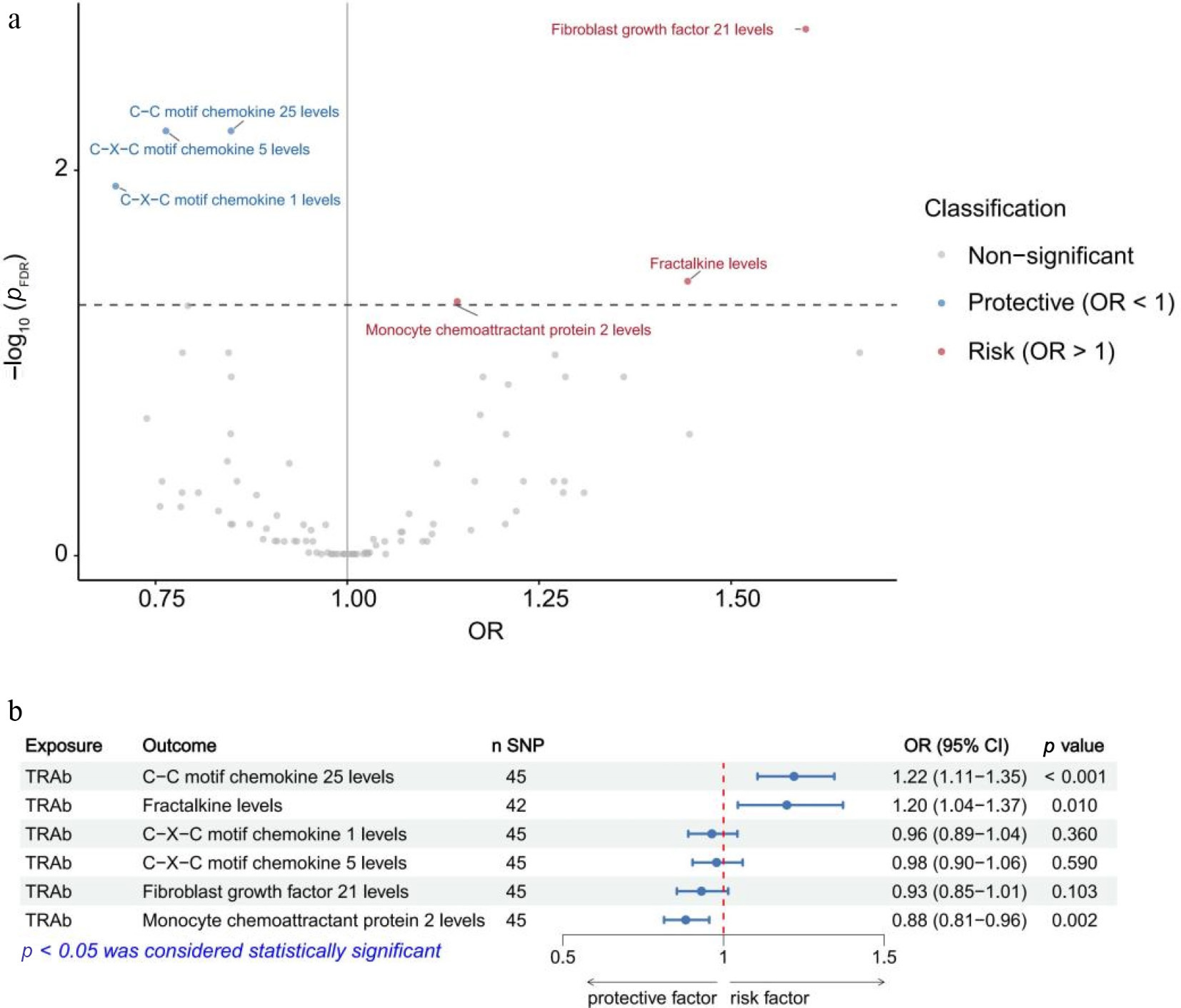
Figure 1.
Mendelian randomization analyses of inflammatory proteins on Graves' ophthalmopathy (a) and of thyrotropin receptor antibodies on inflammatory proteins (b) identified in the VA Million Veteran Program. FDR, false discovery rate; OR, odds ratio; CI, confidence interval; TRAb, thyrotropin receptor antibody; SNP, single-nucleotide polymorphism.
In the UK Biobank dataset, we identified that 20 out of 91 circulating inflammatory proteins were significantly causally associated with GO (p < 0.05). After FDR correction, nine circulating inflammatory proteins remained significant (pFDR < 0.05). The results showed that the genetically predicted higher tumor necrosis factor receptor superfamily member 9 levels (OR: 3.59; p < 0.001; pFDR = 0.001), C-X-C motif chemokine 10 levels (OR: 3.42; p < 0.001; pFDR < 0.001), fractalkine levels (OR: 3.14; p < 0.001; pFDR = 0.001), C-X-C motif chemokine 9 levels (OR: 2.69; p < 0.001; pFDR = 0.005), CD40L receptor levels (OR: 2.02; p < 0.001; pFDR < 0.001), tumor necrosis factor beta (TNF-β) levels (OR: 1.61; p < 0.001; pFDR < 0.001), and C-X-C motif chemokine 6 levels (OR: 1.44; p = 0.004; pDR = 0.041) were positively associated with GO, indicating that elevated genetically predicted these proteins may contribute to an increased susceptibility to GO. In contrast, GO was inversely associated with C-C motif chemokine 4 levels (OR: 0.66; p < 0.001; pFDR < 0.001) and IL-18 levels (OR: 0.43; p < 0.001; pFDR < 0.001) (Fig. 2a).
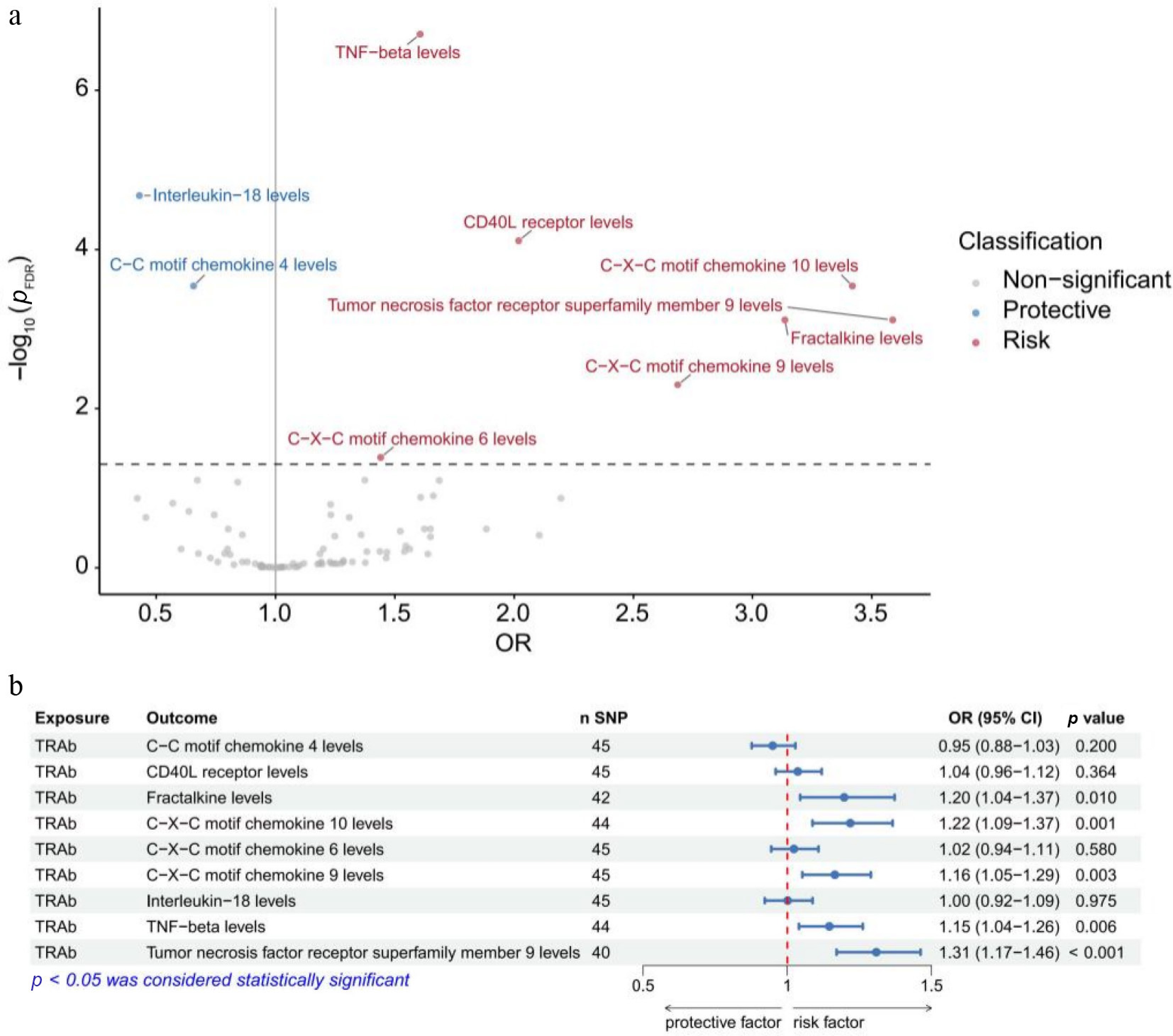
Figure 2.
Mendelian randomization analyses of inflammatory proteins on Graves' ophthalmopathy (a) and of thyrotropin receptor antibodies on inflammatory proteins (b) identified in the UK Biobank. FDR, false discovery rate; OR, odds ratio; CI, confidence interval; TRAb, thyrotropin receptor antibody; SNP, single-nucleotide polymorphism.
Causal effects of TRAb on inflammatory proteins identified
-
In the VA Million Veteran Program dataset, among the inflammatory proteins examined, genetically predicted higher levels of TRAb were positively associated with increased levels of C-C motif chemokine 25 (OR = 1.22, p < 0.001) and fractalkine (OR = 1.20, p = 0.010) but were inversely associated with monocyte chemoattractant protein 2 levels (OR = 0.88, p = 0.002) (Fig. 1b). Considering both the statistical significance and the direction of the effect, fractalkine was selected for inclusion in the subsequent mediation MR analysis of the VA Million Veteran Program sample.
In the UK Biobank dataset, genetically predicted higher TRAb levels were associated with increased levels of fractalkine (OR = 1.20, p = 0.010), C-X-C motif chemokine 10 (OR = 1.22, p = 0.001), C-X-C motif chemokine 9 (OR = 1.16, p = 0.003), TNF-β (OR = 1.15, p = 0.006), and TNF receptor superfamily member 9 (OR = 1.31, p < 0.001) (Fig. 2b). Given the consistency in the direction of the effect, all of these inflammatory proteins were included in the mediation MR analysis of the UK Biobank sample.
Mediated MR analysis
-
Among the potential inflammatory mediators, fractalkine exhibited significant mediation effects in both the VA Million Veteran Program dataset (mediation proportion = 0.07, p = 0.048) and the UK Biobank dataset (mediation proportion = 0.10, p = 0.029). In addition, C-X-C motif chemokine 10 (mediation proportion = 0.12, p = 0.008), C-X-C motif chemokine 9 (mediation proportion = 0.07, p = 0.024), TNF-β (mediation proportion = 0.03, p = 0.012), and TNF receptor superfamily member 9 (mediation proportion = 0.17, p = 0.002) also demonstrated significant mediation effects in the UK Biobank dataset (Table 2). The relationships between CX3CL1, CXCL10, and TNFRSF9 and GO in the UK Biobank dataset showed evidence of pleiotropy (Table 2). The radial plot and radial regression identified 13 outlines (rs1049709, rs16904321, rs184401, rs2517572, rs2518028, rs2523531, rs3093988, rs3131006, rs36067349, rs73137770, rs8050713, rs9271588, and rs4713400) for CX3CL1, 10 outlines (rs1077965, rs11066188, rs11066301, rs11513729, rs11645285, rs2523495, rs3093975, rs3132510, rs72651343, and rs115140093) for CXCL10, and 7 outlines (rs118083884, rs144141891, rs1793894, rs3130063, rs3130976, rs3131617, and rs9669611) for TNFRSF9 (Fig. 3). After excluding the outliers and reanalyzing the data, the mediation effects remained significant for CX3CL1 (mediation proportion = 0.09, p = 0.028) and CXCL10 (mediation proportion = 0.09, p = 0.019), with no indication of pleiotropy (all MR-Egger intercept tests had p > 0.05). In contrast, pleiotropy for TNFRSF9 persisted, and its result was therefore excluded.
Table 2. Mediation analysis of inflammatory proteins in the causal pathway linking thyrotropin receptor antibody to Graves' ophthalmopathy.
Dataset Mediator β overall β1 β2 Mediation proportion p-Value Exposure-outcome pleiotropy test Exposure-mediator pleiotropy test Mediator-outcome pleiotropy test VA Million Veteran Program CX3CL1 0.88 (0.23–1.54) 0.18 (0.04–0.32) 0.37 (0.13–0.60) 0.07 (0–0.15) 0.048 0.212 0.430 0.102 UK Biobank CX3CL1 2.07 (0.61–3.53) 0.18 (0.04–0.32) 1.14 (0.59–1.70) 0.10 (0.01–0.19) 0.029 0.230 0.430 0.012 CXCL10 0.20 (0.08–0.31) 1.23 (0.67–1.79) 0.12 (0.03–0.20) 0.008 0.230 0.205 0.030 CXCL9 0.15 (0.05–0.25) 0.99 (0.44–1.54) 0.07 (0.01–0.14) 0.024 0.230 0.610 0.178 TNF-β 0.14 (0.04–0.23) 0.47 (0.32–0.63) 0.03 (0.01–0.06) 0.012 0.230 0.291 0.093 TNFRSF9 0.27 (0.16–0.38) 1.28 (0.65–1.90) 0.17 (0.06–0.27) 0.002 0.230 0.782 < 0.001 The values of β overall, β1, and β2 represent the overall causal effect of TRAb on GO, the effect of TRAb on inflammatory proteins, and the effect of inflammatory proteins on GO, respectively. The indirect (mediated) effect is calculated as the product of β1 and β2, quantifying the portion of the TRAb–GO association mediated by inflammatory proteins. 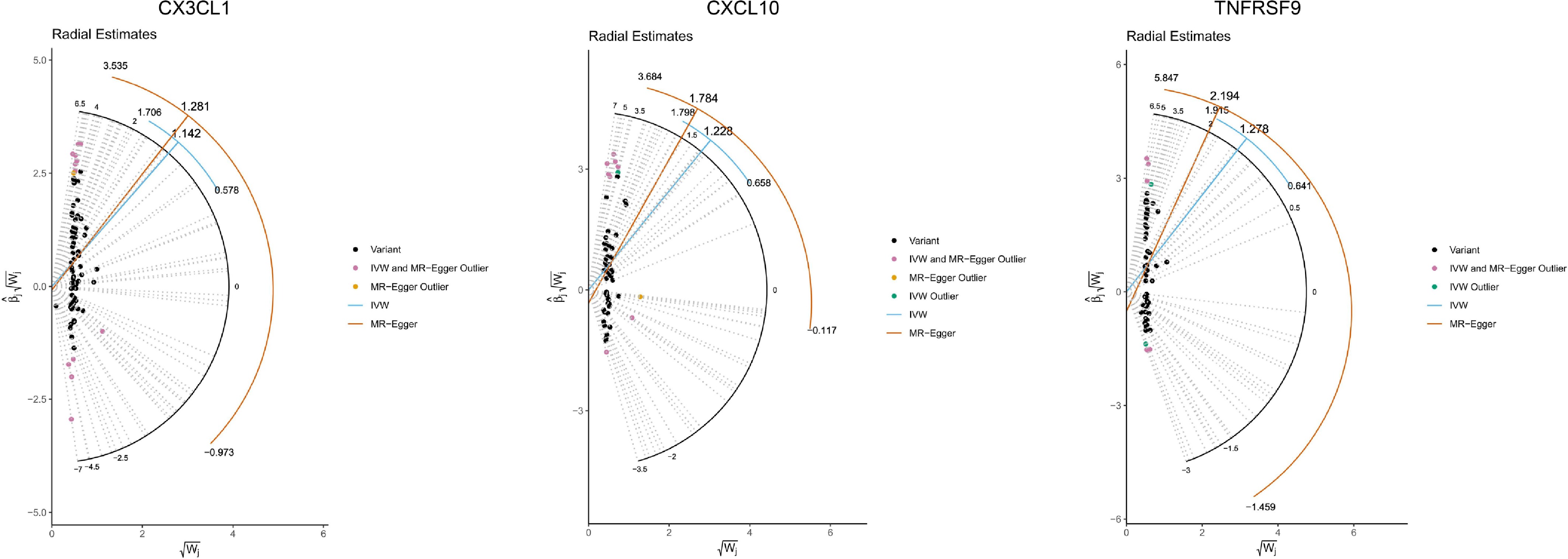
Figure 3.
Radial plots and corresponding radial regression analyses for CX3CL1, CXCL10, and TNFRSF9.
Interaction network analysis
-
The interaction network illustrated in Fig. 4 depicts the functional relationships among the identified mediating proteins and an additional 20 genes with which they are predicted to interact. Within this network, CX3CL1 (fractalkine) is connected through 301 interaction links, CXCL9 (C-X-C motif chemokine 9) through 1061 links, CXCL10 (C-X-C motif chemokine 10) through 1015 links, and lymphotoxin alpha (LTA) (TNF-β) through 350 links, highlighting the extensive interaction landscapes of these mediators. Functional network analysis revealed that CX3CL1, CXCL9, CXCL10, and LTA were significantly involved in 81, 153, 130, and 59 functional categories, respectively (Supplementary Tables S1–S4). For CX3CL1, CXCL9, and CXCL10, the analyses consistently highlighted pathways related to chemokine-mediated signaling, as well as leukocyte migration and chemotaxis, underscoring their central roles in immune cell recruitment. In contrast, LTA was predominantly associated with T-cell activation and NF-κB signaling, reflecting their involvement in immune activation and inflammatory amplification.
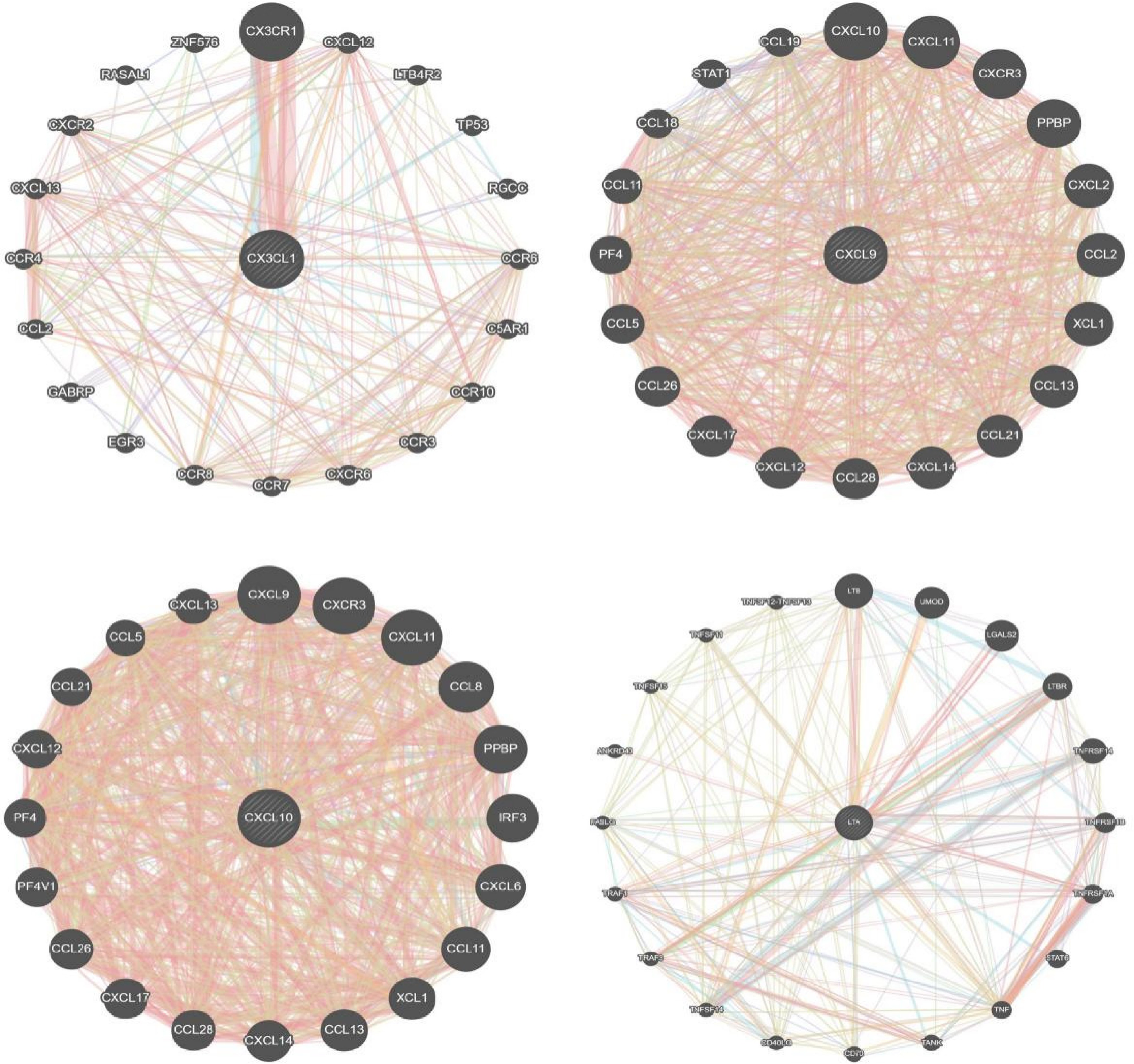
Figure 4.
Interaction network analysis of the identified inflammatory proteins.
-
In this study, we used a comprehensive analytical framework to investigate the causal relationship between TRAb and GO and to identify inflammatory proteins that may mediate this association. Mediation analyses indicated that several inflammatory proteins, including fractalkine (CX3CL1), CXCL9, CXCL10, and TNF-β, partially mediated the effect of TRAb on GO. Notably, CX3CL1 showed a consistent mediating effect in both the VA Million Veteran Program and UK Biobank datasets, whereas CXCL9, CXCL10, and TNF-β were identified as mediators in the UK Biobank dataset. Interaction network analysis further supported the biological relevance of these findings, highlighting chemokine signaling, leukocyte migration, T-cell activation, and NF-κB signaling as key pathways linking TRAb-driven immune dysregulation to the development of GO.
Over the past few decades, three functional categories of TRAbs have been described: Stimulating antibodies (TSAbs), blocking antibodies (TBAbs), and neutral antibodies[40]. Although all three forms may be present in patients with GD, stimulating antibodies are the predominant subtype and are widely considered a key biomarker of the disease[41]. TSAbs are particularly common in individuals with GO and demonstrate strong clinical relevance as well as predictive value[42]. When TRAbs bind to the thyrotropin receptor (TSHR), they promote the production of glycosaminoglycans (GAGs) through activation of intracellular signaling pathways involving cyclic adenosine monophosphate (cAMP) and phosphoinositide 3-kinase/protein kinase B signaling pathway (PI3K/AKT). These signaling cascades partially overlap with those triggered by IGF-1R. Recent research has shown that TSAbs can stimulate the phosphorylation of IGF-1R and activate signaling pathways associated with both TSHR and IGF-1R in human and murine fibroblasts. These observations suggest that TSAbs may amplify IGF-1R's activity, thereby promoting retro-orbital cell proliferation and inflammatory responses[43].
Our analyses indicate that CX3CL1 acts as an important mediator in the downstream inflammatory processes associated with TRAb, as this signal consistently emerged across both human datasets. CX3CL1 exists in two biologically distinct forms: A membrane-bound form (mFKN) and a soluble form (sFKN). Although both forms interact with the same receptor, CX3CR1, they exert different biological effects[44]. The soluble form is generated when metalloproteinases cleave the membrane-anchored protein through proteolytic processing, releasing a fragment that contains the chemokine domain[45]. Functionally, sFKN behaves similarly to classical chemokines and plays a prominent role in directing cell migration. Like many chemotactic signaling systems, cell motility is initiated when sFKN binds to its receptor CX3CR1 on the cell's surface. CX3CR1 belongs to the G-protein-coupled receptor (GPCR) family, and activation of this receptor triggers intracellular signaling pathways that regulate cell movement[46−48]. In general, chemokines coordinate immune responses by establishing concentration gradients that guide leukocytes toward sites of inflammation[49−52]. The broad chemotactic influence of sFKN is largely due to the widespread expression of CX3CR1[45]. This receptor is expressed in a variety of hematopoietic and non-hematopoietic cells. For example, CX3CR1 is present on circulating immune cells, including CD4+ and CD8+ T-lymphocytes[45]. Consequently, CX3CL1-CX3CR1 signaling may facilitate the recruitment and infiltration of CD4+ and CD8+ T-cells into orbital tissues during the development of GO[53].
In addition to CX3CL1, several other molecules, including CXCL9, CXCL10, and TNF-β, were identified as potential mediators in the UK Biobank dataset. Among these, CXCL9 and CXCL10 are chemokines that bind to the CXC chemokine receptor CXCR3[54], and their involvement in GO has been documented previously[16]. Elevated levels of these chemokines have been closely associated with disease activity[16]. Experimental studies have shown that interferon-γ (IFN-γ) can stimulate orbital fibroblasts and preadipocytes to produce CXCL9 and CXCL10, whereas TNF-α can further enhance this response through synergistic interactions with IFN-γ[16,55]. CXCL9 and CXCL10 function to recruit Th1 lymphocytes to sites of inflammation[56−58]. Once recruited, Th1 cells produce additional IFN-γ and TNF-α, which further stimulate the surrounding cells to secrete more Th1-associated chemokines. This process establishes a positive feedback loop that amplifies the inflammatory response[59,60]. Because of their central role in Th1-mediated immune responses, CXCR3 and its ligands have been proposed as promising therapeutic targets[54]. In particular, interventions that inhibit IFN-γ–dependent chemokine production or block the CXCR3 receptor may be especially relevant during the early or active stages of diseases such as GO[54]. The detection of TNF-β indicates activation of TNF superfamily-related inflammatory signaling, although in GO, it likely represents a broader upstream immune context rather than a direct disease-specific effector.
This study has several notable strengths. First, we applied a MR framework to investigate the causal mediators between TRAb and GO, which reduces confounding and reverse causality compared with conventional observational studies. By using genetic variants as instrumental variables, MR provides a more robust approach for causal inference. Second, the analyses incorporated two independent large-scale GWAS datasets, including the VA Million Veteran Program and the UK Biobank, to perform cross-cohort validation. The replication of the key mediator CX3CL1 across these independent populations enhances the reliability and generalizability of the results. Finally, the integration of interaction network analysis provided additional biological context for the identified mediators, further supporting the plausibility of the proposed immunological mechanisms involved in GO's pathogenesis.
However, some limitations should also be considered when interpreting the findings of this study. First, the inflammatory protein data were derived from circulating plasma measurements rather than from orbital tissues. Circulating protein levels may not fully reflect tissue-specific inflammatory activity. Therefore, the identified mediators should be interpreted as systemic biomarkers that may be associated with, but not necessarily identical to, local pathogenic processes. Future studies could utilize techniques such as flow cytometry or in vitro orbital fibroblast models to validate the tissue-specific expression and functional mechanisms of the identified mediators, such as CX3CL1. These approaches would help confirm the localized roles of these inflammatory proteins in the pathogenesis of GO, providing a more detailed understanding of their involvement in tissue-specific immune responses and further refining the therapeutic potential of targeting these mediators. Second, only one independent dataset (FinnGen) was available for TRAb at the time of this analysis. Future research incorporating additional datasets for TRAb would be valuable in further validating and strengthening these findings. Moreover, the GWAS for TRAb captured overall circulating TRAb levels and did not distinguish between functional antibody subtypes, such as stimulating, blocking, or neutral TRAb. Because stimulating TRAb is considered the major pathogenic antibody in GD, future studies incorporating subtype-specific measurements may help refine the mechanistic interpretation of the observed associations. Third, the datasets used in this study include a limited number of GO cases, which may reduce the statistical power to detect modest effect sizes. Therefore, we focus on the intersection of findings from both the UKB and VA Million Veteran Program datasets, while also separately highlighting the unique findings from each dataset. Future studies utilizing larger datasets will be valuable for further validating these results. Fourth, given the nature of MR analysis, the results should be interpreted as qualitative rather than quantitative. This is because the genetic instruments used typically explain only a small proportion of the variance in the traits of interest, and the primary aim of MR is to identify causal relationships rather than estimate the exact magnitude of the effects. Moreover, several methodological limitations related to the mediation framework should be acknowledged. The mediation analysis was based on a Sobel test-based approach applied to summary-level MR data, and its underlying assumptions cannot be fully verified in the present setting. As such, the results may be partly influenced by potential violations of these assumptions. In addition, the observed mediation proportions were relatively modest, indicating that the identified inflammatory proteins explain only a small fraction of the overall TRAb–GO association. Therefore, these mediators should be interpreted as partial contributors rather than dominant or exclusive biological pathways.
-
In conclusion, this study provides genetic evidence supporting a potential mediating role of circulating inflammatory proteins in the pathway linking circulating TRAb to the risk of GO, with CX3CL1 (fractalkine) emerging as a particularly important mediator. These mediators are functionally associated with chemokine signaling, immune cell recruitment, T-cell activation, and NF-κB-related inflammatory pathways. Collectively, our findings further strengthen the evidence that TRAb-associated immune dysregulation may contribute to the development of GO through chemokine-driven leukocyte migration and adaptive immune activation. Further experimental and clinical studies are warranted to clarify the tissue-specific roles of these mediators and to explore their potential as therapeutic targets for GO.
-
All data utilized in this analysis were derived from previously published studies in which informed consent had been obtained from all participants. As a result, no additional ethical approval was required for the present study.
-
The authors confirm contribution to the paper as follows: study conception and design, analysis and interpretation of results: Liang J, Chen J, Zheng Y; data collection: Liang J, Chen J; draft manuscript preparation: Liang J, Chen J, Wu X, Li J, Wang D, Ling J, Mei C, Zheng Y. All authors reviewed the results and approved the final version of the manuscript.
-
The data supporting the findings of this study are publicly available from the FinnGen website for TRAb (https://labvalues.finngen.fi/), the GWAS Catalog for inflammatory proteins (www.ebi.ac.uk/gwas/publications/37563310), and for GO from the VA Million Veteran Program (www.ebi.ac.uk/gwas/studies/GCST90477300) and the UK Biobank (www.ebi.ac.uk/gwas/studies/GCST90435691).
-
We thank the participants and investigators of the FinnGen, UK Biobank, and the VA Million Veteran Program studies. We also thank the staff of Core Facilities at State Key Laboratory of Ophthalmology, Zhongshan Ophthalmic Center for technical support.This publication does not represent the views of the Department of Veterans' Affairs or the United States Government. This study was supported by the Guangzhou Municipal University Joint Funding Project (2025A03J3979), the Research Funds of the State Key Laboratory of Ophthalmology (2025QZLH03; 2025QZSPT35), Guangdong Basic Research Center of Excellence for Major Blinding Eye Diseases Prevention and Treatment, and State Key Laboratory of Ophthalmology, Zhongshan Ophthalmic Center at the Sun Yat-Sen University.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/vns-0026-0022.
-
Authors contributed equally: Jiangbo Liang, Jianqi Chen
- Supplementary Tables S1 Functional enrichment analysis of the CX3CL1-Centered interaction network.
- Supplementary Table S2 Functional enrichment analysis of the CXCL9-Centered interaction network.
- Supplementary Table S3 Functional enrichment analysis of the CXCL10-Centered interaction network.
- Supplementary Table S4 Functional enrichment analysis of the LTA-Centered interaction network.
- Supplementary File 1 The detailed criteria and procedures used for selecting instrumental variables.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liang J, Chen J, Wu X, Li J, Wang D, et al. 2026. Inflammatory proteins as mediators in the causal pathway from thyrotropin receptor antibody to Graves' ophthalmopathy. Visual Neuroscience 43: e028 doi: 10.48130/vns-0026-0022 

Inflammatory proteins as mediators in the causal pathway from thyrotropin receptor antibody to Graves' ophthalmopathy
- Received: 22 March 2026
- Revised: 22 April 2026
- Accepted: 29 April 2026
- Published online: 30 June 2026
Abstract: Graves' ophthalmopathy (GO) is a debilitating autoimmune inflammatory disorder triggered by thyrotropin receptor antibody (TRAb). This study aimed to identify circulating inflammatory proteins mediating the causal pathway from TRAb to GO through a mediation Mendelian randomization (MR) framework. Genome-wide association study (GWAS) summary statistics for TRAb were obtained from the FinnGen project (n = 3,690). Data on circulating inflammatory proteins were derived from a meta-analysis of 11 population-based GWAS cohorts of European ancestry (n = 14,824). GWAS datasets for GO were obtained from the VA Million Veteran Program and UK Biobank for cross-validation. Genetically predicted TRAb levels were associated with an increased risk of GO in both the VA Million Veteran Program (odds ratio [OR] = 2.42, p = 0.008) and the UK Biobank (OR = 7.96, p = 0.005). CX3CL1 showed consistent mediation effects in both datasets. Additionally, CXCL10, CXCL9, and TNF-β exhibited significant mediation effects in the UK Biobank. The identified mediators showed directionally consistent effects, supporting their potential roles in the TRAb–GO pathway. Network analysis highlighted chemokine signaling, leukocyte migration, T-cell activation, and NF-κB signaling as key pathways. These findings provide genetically supported evidence suggesting that TRAb may influence the GO risk through specific inflammatory mediators.
-
Key words:
- Graves' ophthalmopathy /
- Thyrotropin receptor antibody /
- Inflammatory proteins /
- CX3CL1 /
- CXCL10 /
- CXCL9 /
- TNF-β