










-
Diabetes mellitus (DM) has evolved into a global pandemic with an estimated 1.31 billion people projected to be affected by 2050, imposing an unprecedented burden on healthcare systems worldwide[1]. Diabetic retinopathy (DR) and diabetic nephropathy (DN), as common microvascular complications, are respectively the main causes of irreversible blindness and end-stage renal disease[2−4]. Despite advances in glucose-lowering therapies, patient outcomes in DR and DN remain poor. This clinical challenge is rooted in two interrelated difficulties. First, disease progression often continues independently of glycemic control, compounded by a critical lack of therapies that target the shared pathogenic pathways of DR and DN[5−7]. Second, because DR and DN often have no symptoms in the early stage, the current screening methods (such as fundus examination and urine albumin detection) lack sensitivity, convenience, and the ability to jointly provide early warning, usually resulting in delayed diagnosis[8]. This dual shortfall in early detection and mechanism-based therapy highlights the urgent need to both identify novel molecular targets for combined treatment and to develop innovative strategies for the early co-screening of DR and DN.
The pathogenesis of DR and DN is characterized by intricate interplay between metabolic dysregulation, systemic inflammation, and target organ damage[9−11]. Persistent hyperglycemia leads to ischemia and hypoxia in the retinal and renal tissues, triggering excessive production of pro-inflammatory stimuli such as reactive oxygen species (ROS) and activating the pro-inflammatory signal cascade[12,13]. During this process, macrophages, as key innate immune effector cells, have a functional polarization state that is of vital importance. Macrophages typically have two phenotypes: the classic pro-inflammatory M1 polarization phenotype or the alternative anti-inflammatory M2 polarization phenotype. The ultimate realization of function depends on the combination of different polarization markers from M1 to M2[14−17]. The chronic hyperglycemic environment induces macrophages to polarize to the M1 type, resulting in an imbalance in the dynamic balance of M1/M2 macrophages, releasing excessive pro-inflammatory cytokines, promoting the formation of an inflammatory environment in retinal and renal tissues, and ultimately damaging microvessels[18,19]. Therefore, regulating macrophage polarization has become a highly promising key strategy for treating diabetic microvascular lesions such as DR and DN.
G-protein-coupled receptors (GPCRs) are the largest family of membrane receptors in the human body and are currently the targets of over 30% of clinical drugs. They are key molecular switches for cells to sense external environments (such as hormones and neurotransmitters) and convert them into physiological responses within the cells[20,21]. Emerging evidence highlights the role of G-protein-coupled receptors (GPCRs) in integrating metabolic and immune signals, making them promising therapeutic targets for metabolic diseases[22−24]. Multiple chemokine receptors (belonging to the GPCR family) are upregulated in DN kidneys. By recruiting and activating inflammatory cells such as monocytes and macrophages, they infiltrate the renal tissue, leading to glomerular damage and fibrosis[25]. In addition, GPCRs are widely involved in the dialogue between systemic inflammation and local vascular lesions: gut microbiota metabolites can regulate systemic inflammation and immune responses through GPCRs, influencing diseases of the skeletal vascular system[26]. The GPCRs of the nervous system can mediate mechanisms such as microglia/macrophage polarization and cytokine release, driving chronic inflammation, which is highly similar to the inflammatory process in blood vessels[21].
These pieces of evidence suggest that GPCRs may be a key hub connecting disorders of glucose metabolism, inflammatory responses, and microvascular damage in the retina and kidneys, representing a common target for DR and DN. To identify such a potential target, we screened out GPR83, which was significantly upregulated in both tissues, through transcriptomic analysis of the retinal and renal tissues of diabetic rats. Although GPR83 is still an orphan receptor at present, emerging clues suggest its potential significance: it is associated with thym-derived regulatory T cell function and inflammation[27], and has been reported to interact with the AKT signaling pathway in drug-induced ototoxicity models[28]. Research has found that adrenomedullin 2 (ADM2) can affect macrophage polarization[29−32], and it is closely related to eye and kidney diseases[33,34]; downstream signaling pathways include the AKT signaling pathway[35]. Given that the AKT pathway is a core node of microvascular complications in diabetes[36,37], these clues lead to a compelling yet unverified hypothesis: GPR83 may activate AKT through ADM2, driving macrophage polarization and thereby leading to the comorbidity of DR and DN.
Therefore, this study was designed to: (1) establish GPR83 as a common pathogenic hub for DR and DN, and describe its functional connection with the ADM2-AKT pathway; (2) clarify the causal mechanism by which the GPR83-ADM2-AKT signaling axis drives M1-type polarization in macrophages, and evaluate the efficacy of ZJHY targeting this axis; and (3) construct an AI diagnostic model for early DR and DN combined screening based on key genes including GPR83, and transform mechanism insights into clinical applications.
-
Male Sprague-Dawley rats weighing 200–250 g (aged 4–6 weeks) were obtained from the GuangDong Medical Laboratory Animal Center, China. All animals were housed under a standard 12-h light/dark cycle with unrestricted access to food and tap water. All experimental procedures were reviewed and approved by the Animal Ethics Committee of Guangzhou University of Chinese Medicine (Approval No. 20240105013; dated January 5, 2024).
Rats were randomly allocated into three groups using a computer-generated randomization sequence: a normal control group (Control), a diabetes mellitus group (DM), and a ZJHY-treated diabetic group (TX). Animals in the Control group received a standard maintenance diet throughout the experimental period. Rats assigned to the DM and TX groups were placed on a high-fat diet (HFD) for 4 weeks to provoke insulin resistance. Following the HFD induction phase, animals were subjected to an overnight fast (12 h) with continued access to water and then administered a single intraperitoneal (i.p.) injection of streptozotocin (STZ; 40 mg/kg body weight; Sigma, St. Louis, MO, USA) dissolved in citrate buffer (pH 4.5) to induce diabetes. Seventy-two hours after STZ administration, fasting blood glucose levels were determined using a glucometer; only animals with blood glucose exceeding 16.7 mmol/L were considered successfully diabetic and retained in the study. Commencing 4 weeks post-STZ injection, rats in the TX group received daily i.p. injections of ZJHY at a dose of 2.16 g/kg body weight, whereas the DM and Control groups received equivalent volumes of sterile normal saline via the same route. After 4 weeks of treatment, all rats were deeply anesthetized by i.p. injection of 1% pentobarbital sodium, and the eyes and kidneys were rapidly excised for subsequent histological, biochemical, and molecular analyses.
Cell culture and treatment
-
The BV2 murine microglial cell line was procured from Wuhan University, Wuhan, China. Cells were cultured in low-glucose (5 mmol/L) Dulbecco's Modified Eagle Medium (DMEM) enriched with 10% (v/v) heat-inactivated fetal bovine serum (FBS; catalog no. 10091148; Gibco, Grand Island, NY, USA) and 1% (v/v) penicillin (100 U/mL)/streptomycin (100 μg/mL; catalog no. 15140122; Invitrogen, Carlsbad, CA, USA). Cultures were maintained at 37 °C in a humidified incubator with 5% CO2. When cultures reached approximately 70% confluence, cells were passaged and seeded into sterile flat-bottom 6-well plates for experiments.
Primary peritoneal macrophages (PEMs) were isolated from rats using a standard peritoneal lavage technique. In brief, rats were euthanized by carbon dioxide asphyxiation followed by cervical dislocation. The ventral abdominal surface was disinfected with 70% (v/v) ethanol, and a shallow midline incision was made through the skin to expose the underlying peritoneal wall without perforating the intestinal serosa. Ice-cold sterile phosphate-buffered saline (PBS, 10 mL) supplemented with 2 mM EDTA was slowly instilled into the peritoneal cavity via a 25-gauge needle. The abdomen was gently agitated for 30–60 s to dislodge resident peritoneal cells, after which the lavage fluid was carefully aspirated using a sterile pipette. The recovered cell suspension was centrifuged at 300 × g for 5 min at 4 °C. The resulting cell pellet was resuspended in complete DMEM medium (low-glucose DMEM containing 10% heat-inactivated FBS and 1% penicillin/streptomycin) and plated at a density of 1 × 106 cells per well in 6-well plates. After a 2-h incubation at 37 °C in a 5% CO2 atmosphere to permit macrophage attachment, non-adherent cells were removed by gentle rinsing with warm PBS. The remaining adherent macrophage population was employed for all subsequent experiments.
Hematoxylin and Eosin (H&E) staining and Immunohistochemistry (IHC) staining
-
Retinal and renal tissue specimens were fixed by overnight immersion in 4% (w/v) paraformaldehyde in PBS at 4 °C. Fixed tissues were dehydrated stepwise through ascending concentrations of ethanol, cleared in xylene, and infiltrated with molten paraffin wax. Paraffin-embedded blocks were trimmed and cut at a thickness of 5 μm using a rotary microtome (Leica Microsystems, Wetzlar, Germany). Tissue sections were deparaffinized in xylene, rehydrated through a descending ethanol series to distilled water, and stained with hematoxylin and eosin (H&E) following routine protocols. Histological examination and photomicrograph acquisition were performed using a light microscope equipped with a digital camera. For immunohistochemistry (IHC), sections were processed with UltraSensitive™ SP Kits (Servicebio, Wuhan, China) together with DAB chromogenic detection reagents (Servicebio, Wuhan, China) in strict accordance with the manufacturer's instructions. After antigen retrieval and blocking of endogenous peroxidase activity, sections were probed overnight at 4 °C with a primary antibody against GPR83 (1:500 dilution; ProMab Biotechnologies Inc., Richmond, CA, USA; Cat. No. 32243). Following incubation with biotinylated secondary antibody and streptavidin-horseradish peroxidase, immunoreactivity was visualized with DAB, and sections were counterstained with hematoxylin.
Immunofluorescence staining assay
-
Paraffin-embedded retinal and kidney sections were first subjected to deparaffinization by xylene immersion (10 min) and rehydrated through decreasing ethanol concentrations. Following washes with distilled water and PBS, non-specific binding was blocked by applying goat serum to the tissue sections. For occludin detection, retinas were incubated overnight at 4 °C with anti-occludin primary antibody, followed by Cy3-conjugated goat anti-rabbit IgG secondary antibody (1:200 dilution) for 1 h at room temperature. Nuclei were counterstained with DAPI for 10 min, and images were captured using a BX53 fluorescence microscope (Olympus, Tokyo, Japan). Dual-label immunofluorescence was performed on paraffin sections processed as above. Sections were co-incubated overnight at 4 °C with anti-IBA1 (1:300; Proteintech Group, Inc., 10904-1-AP) and anti-Arg-1 (1:200; Proteintech Group, Inc., 16001-1-AP) primary antibodies. After PBS washes, sections were labeled with Cy3-conjugated goat anti-mouse IgG (1:200) and FITC-conjugated goat anti-rabbit IgG (1:200) for 90 min at room temperature. Nuclei were visualized with DAPI counterstaining, and fluorescent images were acquired on a BX53 microscope (Olympus, Tokyo, Japan).
Western blot
-
Protein concentrations in tissue and cell lysates were quantified using the BCA Protein Assay Kit (Beyotime Biotechnology, Shanghai, China), and all samples were normalized to equal protein loading. Equal amounts of denatured protein (25 μg per lane) were resolved by electrophoresis on 10% sodium dodecyl sulfate-polyacrylamide gels (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA, USA). Membranes were blocked with 5% (w/v) nonfat dry milk dissolved in Tris-buffered saline containing 0.1% Tween-20 (TBST) for 1 h at room temperature and then incubated overnight at 4 °C with the following primary antibodies: anti-BCL2 (Proteintech Group Inc.; Cat. No. 68103-1-Ig), anti-BAX (Proteintech Group Inc.; Cat. No. 60267-1-Ig), anti-GPR83 (1:500; ProMab Biotechnologies Inc.; Cat. No. 32243), anti-AKT (1:500; Proteintech Group Inc.; Cat. No. 10176-2-AP), anti-phospho-AKT (Ser473) (1:1,000; Proteintech Group Inc.; Cat. No. 66444-1-Ig), anti-PI3K (1:200; ABclonal Technology; Cat. No. A19742), anti-phospho-PI3K (1:1,000; ABclonal Technology; Cat. No. AP1463), anti-ADM2 (1:2,000; Hubei Ipodix Biotechnology Co., Ltd., Wuhan, China; Cat. No. IPDX9645-A), and anti-GAPDH (loading control; 1:10,000; Proteintech Group Inc.). After extensive washing with TBST, the membranes were probed with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG or goat anti-mouse IgG secondary antibodies for 1 h at ambient temperature. Immunoreactive bands were detected by enhanced chemiluminescence (ECL) and imaged using a gel documentation system (WD-9413B; Beijing Liuyi Biotechnology Co., Beijing, China).
Quantitative real-time polymerase chain reaction (qPCR)
-
Total RNA was extracted separately from retinal tissues, renal tissues, and cultured cells using TRIzol™ reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's recommended protocol. RNA purity and concentration were assessed by spectrophotometry (NanoDrop, Thermo Fisher Scientific). Complementary DNA (cDNA) was synthesized from 1 μg of total RNA using a reverse transcription kit (Takara Bio, Shiga, Japan). Quantitative real-time PCR (qRT-PCR) was conducted to measure the mRNA expression levels of Gpr83, Adm2, Pi3k, Akt, Tnf-α, Il-6, Il-10, Il-1β, Cd86, Cd206, Bax, and Bcl2. Gene-specific primers were synthesized by GENEWIZ Inc. (Suzhou, China) and Servicebio (Wuhan, China), and the complete primer sequences are provided in Supplementary Table S1. Each 25-μL reaction mixture consisted of 1 μL template cDNA, 1 μL each of forward and reverse primers (10 μM), 12.5 μL of TB Green® Premix Ex Taq™ II (Tli RNaseH Plus; Takara Bio, Japan), and 8.5 μL of nuclease-free water. Amplification was carried out over 40 cycles with the following thermal cycling conditions: denaturation at 95 °C for 5 s and annealing/extension at 60 °C for 30 s. A melt curve analysis was appended to each run to verify the specificity of amplification. All reactions were run in triplicate, and relative gene expression was calculated by the 2−ΔΔCt method with Gapdh serving as the endogenous reference gene.
Transfection of siRNA and plasmid DNA
-
For gene silencing experiments, small interfering RNAs (siRNAs) specifically targeting the genes of interest, along with a scrambled non-targeting siRNA as the negative control, were designed and synthesized by Servicebio Biotechnology Co., Ltd. (Wuhan, China). Cells were seeded in antibiotic-free medium and allowed to reach 70%–80% confluence before transfection. siRNA-lipid complexes were formed using Lipofectamine™ 3000 transfection reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer's standard protocol and applied to the cell monolayers. The transfection medium was replaced with fresh complete culture medium after 6 h. Knockdown efficiency was assessed 24 h post-transfection by qRT-PCR for mRNA-level validation and by Western blotting for protein-level confirmation.
For ectopic overexpression of Gpr83, a plasmid construct harboring the full-length coding sequence of rat Gpr83 under a CMV promoter, together with a matching empty vector control, was transfected into cells using Lipofectamine™ 3000 reagent (Invitrogen) as described above. Cells were harvested 48 h after transfection to ensure sufficient accumulation of recombinant protein. Overexpression was verified at the transcript level by qRT-PCR and at the protein level by Western blot analysis.
The oligonucleotide primer sequences used for qRT-PCR are detailed in Supplementary Table S1. All transfection experiments were performed in biological triplicate.
Elisa assay
-
The concentrations of inflammatory cytokines in rat serum samples were determined using commercially available enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturer's protocols. The following rat-specific kits were employed: TNF-α (Cat. No. RX2DW31063), IL-6 (Cat. No. RXG60009), IL-1β (Cat. No. RX2DW30206), and IL-10 (Cat. No. RX2DW30194), all sourced from RUIXIN Biotech (Fujian, China). Optical absorbance was measured at 450 nm on an automated microplate reader (Molecular Devices, Sunnyvale, CA, USA). Each sample and standard was assayed in triplicate, and cytokine concentrations were interpolated from standard curves generated with recombinant protein standards.
RNA sequencing and data analysis
RNA extraction, library construction, and sequencing
-
Total RNA was isolated from rat retinal and kidney specimens as detailed above. The integrity and purity of each RNA preparation were verified by agarose gel electrophoresis and spectrophotometric analysis. Only RNA samples that met stringent quality criteria (RNA integrity number ≥ 7.0) were advanced to library construction. Polyadenylated messenger RNA was selectively enaboveriched using oligo(dT)-conjugated magnetic beads and then fragmented into short segments by incubation at elevated temperature in the presence of divalent cations. First-strand cDNA synthesis was primed with random hexamers using the fragmented mRNA as a template, followed by second-strand cDNA generation employing DNA Polymerase I and RNase H. The resulting double-stranded cDNA was purified with a commercial PCR purification kit. Purified cDNA ends were repaired to produce blunt ends, followed by the addition of a single adenosine (A) overhang at the 3′ termini and ligation of indexed sequencing adapters. Adapter-ligated fragments were size-selected by gel electrophoresis and enriched through limited-cycle PCR amplification to produce the final cDNA library. Library quality and concentration were assessed on a Bioanalyzer, and high-throughput paired-end sequencing was carried out on the Illumina NovaSeq 6000 platform (Illumina, San Diego, CA, USA).
Data preprocessing and differential expression gene analysis
-
Raw FASTQ sequencing files were preprocessed to remove adaptor contaminants, poly-N reads, and low-quality bases using fastp. The resulting high-quality clean reads were aligned to the rat reference genome (Rnor_6.0) using HISAT2, and gene-level read counts were quantified with featureCounts. Identification of differentially expressed genes (DEGs) was performed with the DESeq2 package in the R statistical environment. Genes satisfying the combined thresholds of |log2 fold change| > 1 and Benjamini-Hochberg adjusted p-value < 0.05 were designated as significantly differentially expressed in the pairwise contrasts: DM vs. Control, and TX vs. DM.
Gene expression pattern classification and core gene screening
-
Differentially expressed genes were systematically categorized on the basis of their directional expression changes in the diabetic state relative to the control and in response to ZJHY treatment. Two expression patterns were of primary interest: (1) genes whose expression was elevated in the DM group compared with the Control and subsequently suppressed in the TX group vs. DM; and (2) genes whose expression was decreased in DM vs. Control and restored in TX vs. DM. By intersecting these two patterns across retinal and renal tissues, we identified a core set of genes displaying a consistent, reversible expression profile upon ZJHY administration in both target organs.
Functional enrichment analysis
-
To gain insight into the biological roles of the core gene set, functional enrichment analysis was conducted using Gene Ontology (GO) categories (biological process, cellular component, and molecular function) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway mapping. Enrichment analysis was implemented with the clusterProfiler R package, and terms or pathways with a Benjamini–Hochberg corrected p-value below 0.05 were considered statistically significantly enriched.
Construction of a predictive model
Data acquisition and preprocessing
-
Publicly archived transcriptomic datasets relevant to diabetic retinopathy (DR) and diabetic nephropathy (DN) were retrieved from the Gene Expression Omnibus (GEO) repository of the National Center for Biotechnology Information (NCBI). Two DR datasets (GSE102485 and GSE160306) and three DN datasets (GSE262793, GSE166239, and GSE47183), all generated from human clinical specimens, were selected for this study. Raw count matrices were normalized and preprocessed using the DESeq2 package in R. Inter-study batch effects across the pooled datasets were identified and corrected using the sva (surrogate variable analysis) package to enable integrated, cross-cohort analyses.
Identification of differentially expressed genes (DEGs)
-
Differential gene expression analysis of the integrated GEO datasets was performed using the DESeq2 R package. Transcripts with an absolute log2 fold change (|log2FC|) exceeding 1 and a Benjamini–Hochberg adjusted p-value below 0.05 were classified as differentially expressed genes (DEGs). Separate gene lists were generated for DR (DR-DEGs) and DN (DN-DEGs), comparing disease samples with their respective controls.
Weighted gene co-expression network analysis (WGCNA)
-
Weighted gene co-expression network analysis (WGCNA) was conducted using the WGCNA R package. Eighty percent of genes exhibiting the highest expression variance across samples were retained for network construction. A soft-thresholding power (β) was selected based on the criterion of approximate scale-free topology (R2 > 0.8). Modules displaying significant Pearson correlation with DR or DN clinical phenotypes (|correlation coefficient| > 0.3) were identified as trait-relevant modules. Hub genes within these modules—defined by high intramodular connectivity—were extracted to compile disease-specific gene signatures (DR-Sig and DN-Sig).
Screening of immune-related candidate genes
-
A comprehensive catalog of 2,483 immune-related genes (IRGs) was downloaded from the Immunology Database and Analysis Portal (ImmPort;
www.immport.org ). The four gene sets derived from the preceding analyses—namely, DR-DEGs, DN-DEGs, DR-Sig, and DN-Sig—were each intersected with the ImmPort IRG list. This intersection yielded a total of 11 immune-linked candidate genes that were advanced to the machine learning-based feature selection stage.Machine learning-based feature selection
-
To distill the most robust and predictive biomarkers from the 11-candidate-gene pool, we deployed a multi-algorithm feature selection framework encompassing three complementary machine learning approaches: Least Absolute Shrinkage and Selection Operator (LASSO) logistic regression, Support Vector Machine (SVM) with recursive feature elimination, and Random Forest (RF) analysis. Each algorithm independently evaluated the association between individual genes and disease status (DR or DN) in the merged and batch-corrected dataset. Genes that were consistently ranked within the top eight most influential features by all three algorithms were retained as the final immune-related gene signature.
Predictive model construction and validation
-
Using the five-gene immune signature as input features, we constructed a series of predictive classifiers for DR and DN employing seven distinct machine learning algorithms: Naive Bayes (NB), Random Forest (RF), Support Vector Machine (SVM), AdaBoost Classification Trees (AdaBoost), Boosted Logistic Regression (LogitBoost), K-Nearest Neighbors (KNN), and the Cancerclass algorithm. Each model was trained on the designated discovery cohort (GSE160306 for DR; GSE262793 for DN) and subsequently evaluated on an independent validation cohort (GSE102485 for DR; GSE47183 for DN) to assess generalizability to unseen data. For all parametric models, hyperparameters were systematically optimized through grid search combined with 10 repetitions of 5-fold cross-validation on the training set; the non-parametric Cancerclass algorithm was fitted directly to the entire training partition without tuning. Model performance was quantified by the area under the receiver operating characteristic curve (AUC), and the top-performing model for each disease was identified.
Chromosomal localization and pathway analysis
-
The genomic coordinates of the five signature genes (Acvr1c, Tnfsf9, Gpr83, Shc4, and Ednra) were retrieved from the Ensembl genome browser and visualized as a circular karyotype ideogram using the RCircos R package. To further explore the functional significance of the gene panel, pathway enrichment analysis was performed as described previously.
Preparation of ZJHY
-
The ZJHY standardized decoction was prepared from the following medicinal herbs (all obtained from the Outpatient Pharmacy, First Clinical Medical College of Guangzhou University of Chinese Medicine, Guangzhou, China): Astragalus membranaceus (Huangqi) 15 g, Pueraria lobata (Gegen/Kudzu root) 7.5 g, Rehmannia glutinosa (Dihuang) 7.5 g, Prunus persica (Taoren/Peach kernel) 6 g, Cornus officinalis (Shanzhuyu) 7.5 g, Panax notoginseng (Sanqi) 6 g, Rheum officinale (Dahuang/Rhubarb) 3 g, Crataegus pinnatifida (Shanzha/Hawthorn) 5 g, Spatholobus suberectus (Jixueteng) 10 g, Haliotis diversicolor (Shijueming/Abalone shell) 10 g, Bombyx batryticatus (Jiangcan) 5 g, and Boswellia carterii (Ruxiang/Frankincense) 3 g. The crude herbal mixture was placed in a ceramic decoction vessel with 1,000 mL of purified water and heated to a boil. Once boiling was achieved, the heat was reduced, and the mixture was simmered for 50 min. The liquid was then separated from the herbal residue by filtration through sterile gauze, affording approximately 500 mL of the primary decoction. The retained herbal dregs were combined with 800 mL of fresh purified water, brought to a boil a second time, and simmered for an additional 30 min. This secondary filtrate (~400 mL) was combined with the primary decoction to yield a pooled extract of approximately 900 mL. The pooled liquid was further concentrated by gentle heating until the final volume was reduced to 100 mL. After passage through a 0.22-μm sterile filter, the concentrated decoction was lyophilized to obtain a dry powder, which was stored in a desiccator at −20 °C until use.
For in vitro applications, the lyophilized ZJHY powder was freshly reconstituted in sterile phosphate-buffered saline (PBS, pH 7.4) to prepare a 100 mg/mL stock solution on the day of each experiment. Serial dilutions of this stock were made in complete culture medium to obtain the desired final working concentrations. The cytotoxicity profile of ZJHY was evaluated using the Cell Counting Kit-8 (CCK-8) assay. BV2 microglial cells or primary peritoneal macrophages were exposed to a range of ZJHY concentrations for 24 h, after which CCK-8 reagent was added, and absorbance was recorded according to the kit protocol. The highest concentration that preserved cell viability above 90% relative to untreated controls was designated as non-cytotoxic and selected for all subsequent anti-inflammatory activity and functional assays.
UPLC-Q-TOF/MS analysis
-
For sample preparation, a 200-μL aliquot of the herbal decoction was mixed thoroughly with 800 μL of methanol (LC-MS grade). The mixture was homogenized by bead-milling for 5 min, vortexed vigorously for 10 min, and centrifuged at 13,000 rpm for 10 min at room temperature. The clarified supernatant was carefully transferred to an autosampler vial for instrumental analysis.
Qualitative phytochemical profiling was carried out on an ultra-performance liquid chromatography system interfaced with a quadrupole time-of-flight mass spectrometer (UPLC-Q-TOF/MS) equipped with an Agilent 6546 QTOF mass analyzer (Agilent Technologies, Santa Clara, CA, USA). Chromatographic separation was accomplished on an AQ-C18 reversed-phase analytical column (150 mm × 2.1 mm inner diameter, 1.8 μm particle size; Welch Materials, Shanghai, China) thermostated at 35 °C. The mobile phase was composed of 0.1% (v/v) formic acid in water (solvent A) and acetonitrile (solvent B), delivered at a constant flow rate of 0.5 mL/min. The gradient elution program was set as follows: 0–1 min, 2% B; 1–5 min, 2%–20% B; 5–10 min, 20%–50% B; 10–15 min, 50%–80% B; 15–20 min, 80%–95% B; 20–27 min, 95% B (isocratic hold); 27–28 min, 95%–2% B (return to initial conditions); 28–30 min, 2% B (column re-equilibration). The sample injection volume was 5 μL. Mass spectrometric detection was performed in both positive and negative electrospray ionization modes, with data acquired over the mass range of 50–1,700 m/z.
Statistical analysis
-
All statistical computations were executed using GraphPad Prism v10.0 (GraphPad Software Inc., San Diego, CA, USA). Data from independent biological replicates are presented as the mean ± standard deviation (SD). The Shapiro–Wilk test was first applied to assess whether each dataset conformed to a normal (Gaussian) distribution. For normally distributed variables with equal variance (verified by Bartlett's test), differences between two groups were examined by two-tailed unpaired Student's t-test, and differences among three or more groups were evaluated by one-way or two-way analysis of variance (ANOVA) followed by Tukey's honestly significant difference (HSD) post-hoc test for multiple pairwise comparisons. When the normality assumption was violated, nonparametric methods were employed: the Mann–Whitney U test for two-group comparisons and the Kruskal–Wallis test with Dunn's multiple-comparison correction for multi-group analyses. A two-tailed p-value of less than 0.05 was taken as the threshold for statistical significance. All experiments were repeated at least three times independently.
-
We used a streptozotocin-induced diabetic rat model to evaluate the in vivo effects of ZJHY on diabetic retinopathy (DR) and nephropathy (DN) (Fig. 1a). Fasting blood glucose (FBG) and body weight serve as key indicators of diabetes progression. Throughout the experiment, diabetic (DM) rats displayed sustained hyperglycemia and weight loss relative to controls, consistent with typical diabetic pathology (Fig. 1b, c). ZJHY treatment did not significantly improve FBG or body weight in the treated (TX) group, but it markedly reduced serum levels of pro-inflammatory cytokines in diabetic rats (Fig. 1d). In addition, ZJHY ameliorated dyslipidemia and renal dysfunction, lowering serum TG, TC, UREA, and CREA levels (Fig. 1e). Hematoxylin and eosin (H&E) staining was performed to further assess ZJHY's effects on target organs. The DM group exhibited pronounced pathological changes in the retina and kidney, whereas ZJHY treatment substantially alleviated these injuries, yielding tissue architecture closer to that of the control group (Fig. 1f–i). These findings suggest that ZJHY can simultaneously ameliorate both DR and DN, possibly through an as-yet-unknown mechanism independent of glycemic control. To identify the active constituents responsible for these co-protective effects, we performed UPLC-Q-TOF/MS analysis. A Base Peak Chromatogram (BPC) of the ZJHY extract was acquired, and component identification was conducted using UNIFI 1.9.4.0 software. A total of 85 compounds were characterized, including phenolic acids, flavonoids, terpenoids, and ginsenosides, among others (Fig. 1j), establishing a chemical foundation for ZJHY's pharmacological activity. Detailed data are provided in Supplementary Table S2.
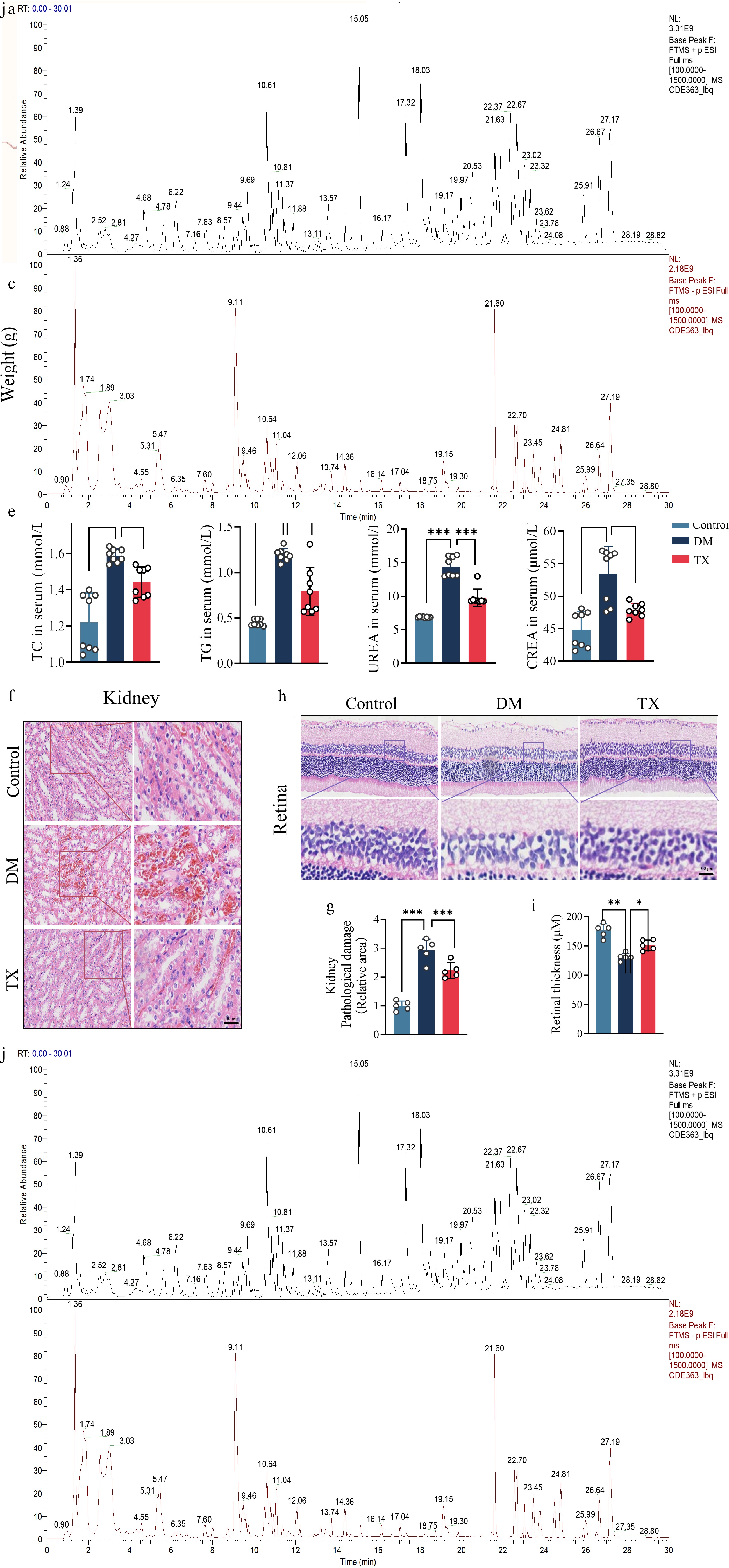
Figure 1.
ZJHY ameliorates metabolic disturbance, retinal and renal injury, and renal dysfunction in diabetic rats. (a) Schematic of the animal experimental design: Sprague-Dawley rats were randomized into the control group (Control), the diabetes mellitus group (DM), and the ZJHY-treated diabetic groups (TX). DM was induced by HFD and streptozotocin; ZJHY was administered 4 weeks post-STZ for 4 weeks. Time-course analysis of (b) fasting blood glucose (FBG), and (c) body weight during the experimental period (n = 7 per group). (d) Serum levels of inflammatory factors (IL-6, IL-1β) by ELISA or biochemical assays (n = 7 per group). (e) Serum levels of lipid metabolites (triglycerides [TG], total cholesterol [TC]), and renal function markers (urea [UREA], creatinine [CREA]) measured by ELISA or biochemical assays (n = 8 per group). Hematoxylin and Eosin (H&E) staining of (f) retinal, and (h) kidney tissues (scale bar: 100 μm). Quantitative histological scoring of (g) kidney, and (i) retinal damage (n = 5 per group). (j) Base Peak Chromatogram (BPC) of ZJHY extract analyzed by UPLC-Q-TOF/MS; 85 compounds were identified (Supplementary Table S2). Data are presented as mean ± SD. Statistical significance was determined by one-way ANOVA with Tukey's post-hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
In summary, ZJHY exerts co-protective effects on both retinal and renal tissues under diabetic conditions without disrupting systemic glucose metabolism. This finding indicates that ZJHY protects against DR and DN through a yet-uncharacterized mechanism independent of glycemic control, warranting further investigation.
ZJHY attenuates M1 macrophage polarization and inflammation in diabetic retina and kidney
-
Building on the observed protective effects of ZJHY, we next examined its influence on macrophage polarization in both tissues. ZJHY treatment significantly increased the Arg1+/Iba1+ ratio, reflecting reduced M1-type macrophage polarization (Fig. 2a, b). In the DM group, transcript levels of CD86 (an M1 marker) were elevated in the retina and kidney compared to controls, whereas those of CD206 (an M2 marker) were diminished. ZJHY treatment reversed these changes (Fig. 2c, d). Gene expression profiles of inflammation-related mediators, including Il-6, Il-1β, Tnf-α, and Il-10, further corroborated these observations, demonstrating downregulation of pro-inflammatory factors and upregulation of the anti-inflammatory mediator in both tissues (Fig. 2e, f). Moreover, the ratio of the apoptosis regulators BAX to BCL-2 was significantly decreased in both tissues, suggesting that ZJHY also attenuated cellular apoptosis (Supplementary Fig. S1a–S1c).
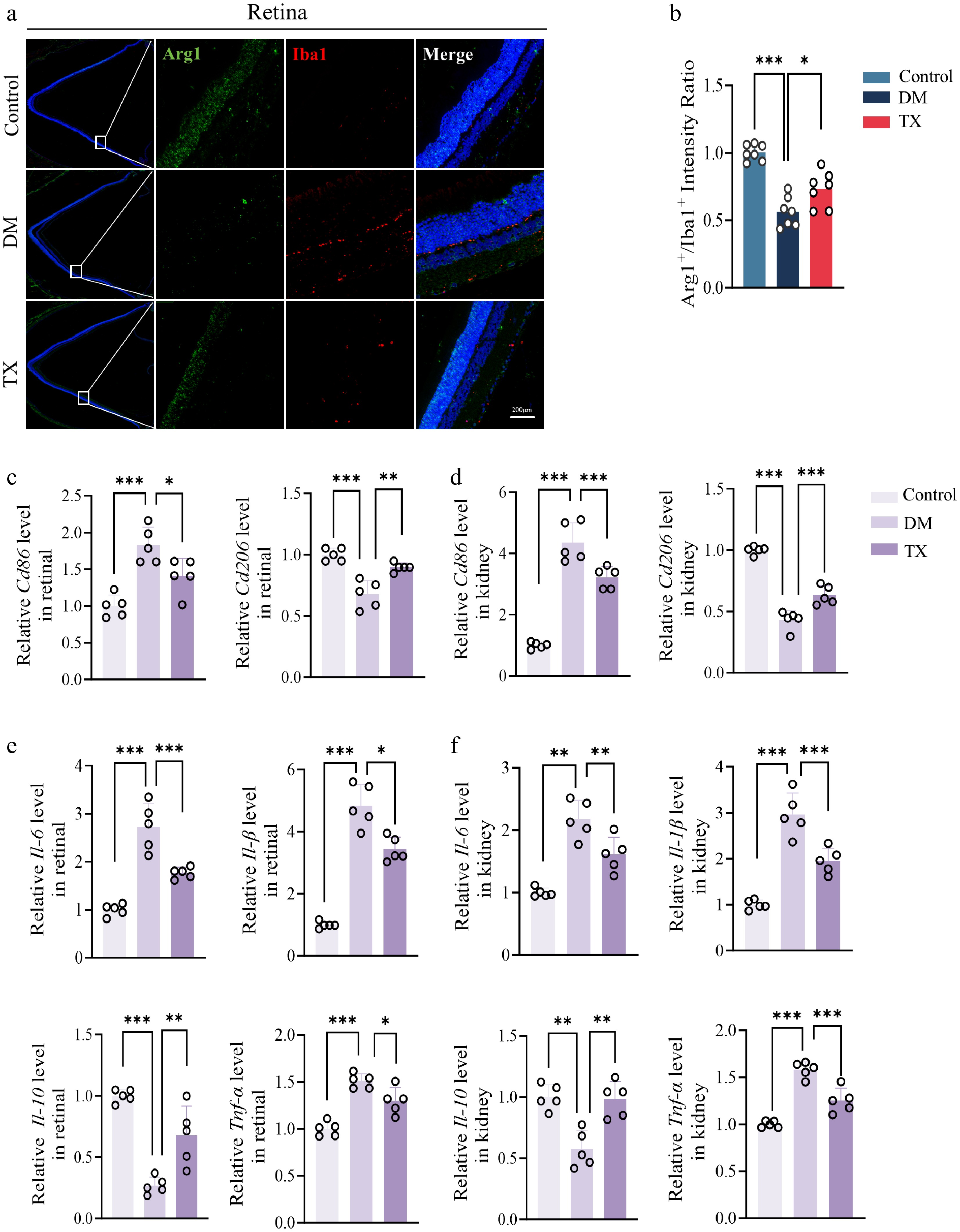
Figure 2.
ZJHY suppresses macrophage-mediated inflammation and cellular apoptosis in diabetic retinal and kidney. (a), (b) Immunofluorescence staining of retinal for (a) Iba-1 and Arg-1 (scale bar: 100 μm), and (b) quantification of the Arg1+ /Iba1+ratio (n = 7 per group). qPCR test for (c) Cd68, and (d) Cd206 in retinal and kidney tissues (n = 5 per group). (e), (f) Relative mRNA expression of pro-inflammatory genes (Tnf-α, Il-6, Il-1β) and anti-inflammatory gene (Il-10) in retinal and kidney measured by qPCR (n = 5 per group). Data are presented as mean ± SD. Statistical significance was determined by one-way ANOVA with Tukey's post-hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
In conclusion, these data indicate that the ZJHY formula protects the retina and kidneys from diabetic damage by inhibiting M1 polarization of macrophages, reducing the secretion of inflammatory factors, and suppressing the apoptotic pathway.
GPR83 is identified as a shared pathogenic target for diabetic retinopathy and nephropathy
-
To elucidate the molecular mechanisms underlying the therapeutic effects of ZJHY on diabetic retinal and nephropathy-related inflammatory lesions, transcriptomic analysis was performed on renal and retinal tissues. After RNA extraction and quality control, libraries were sequenced, and the resulting data were processed to identify differentially expressed genes (DEGs). We analyzed the upregulated and downregulated genes in retinal and renal tissues separately by comparing the DM group with the Control group, as well as the TX group with the DM group (Fig. 3a). Genes were then categorized based on their expression patterns: those upregulated by diabetes and subsequently downregulated by ZJHY treatment, or conversely, downregulated by diabetes and upregulated by treatment (Fig. 3b). This classification identified a set of genes exhibiting consistent and reversible expression dynamics across both tissues.
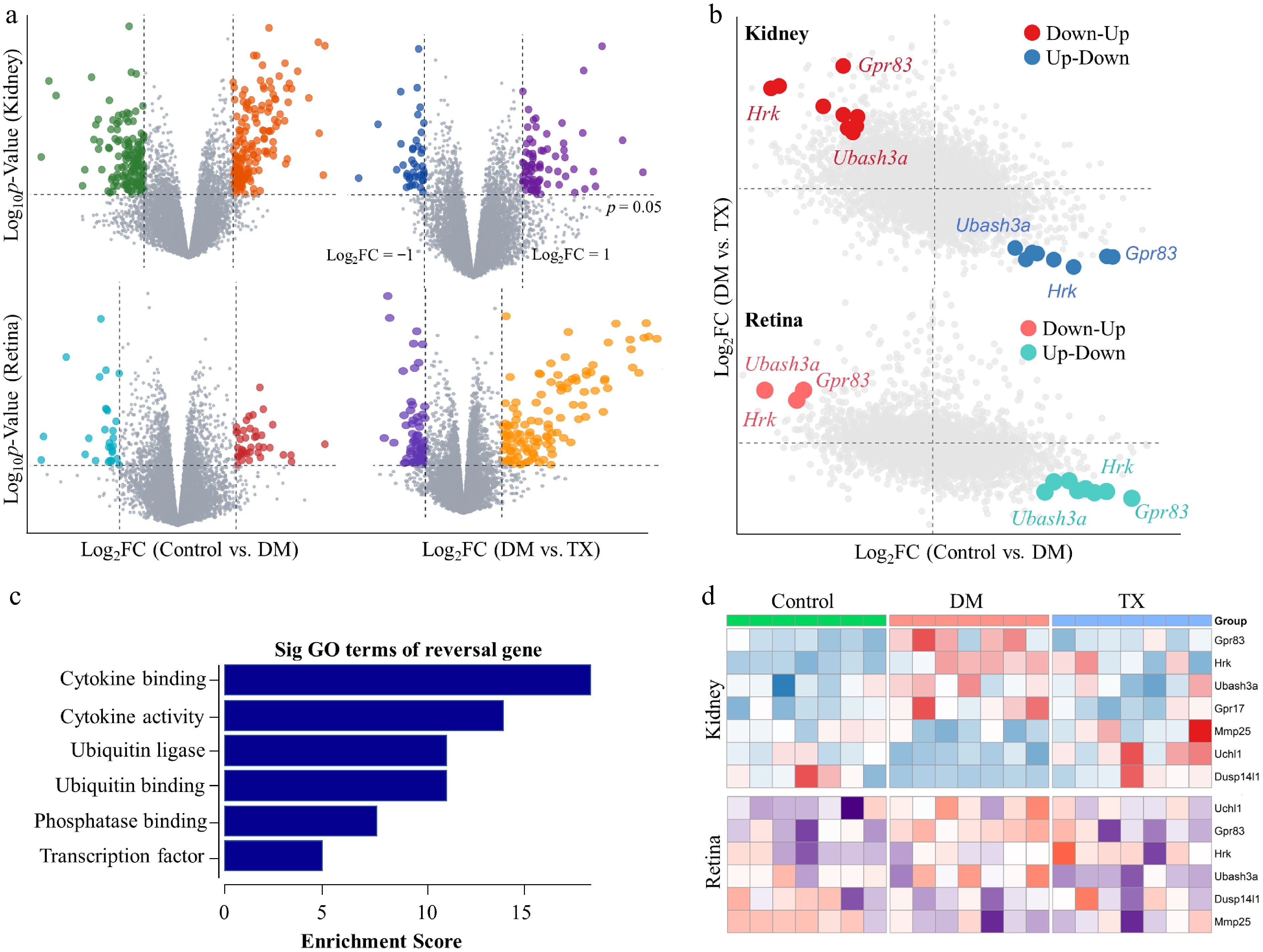
Figure 3.
Transcriptomic analysis identifies GPR83 as a key target of ZJHY in DR and DN. (a) Volcano plots of differentially expressed genes (DEGs) in the kidney and retina: left panels show DEGs between DM and Control groups, right panels show DEGs between the TX and DM groups (|log2 Fold Change| > 1, adj. p < 0.05). (b) Venn diagrams illustrating the overlap of DEGs with consistent expression patterns: upregulated in diabetes and downregulated by ZJHY treatment (top), or downregulated in diabetes and upregulated by ZJHY treatment (bottom) in both tissues. (c) Gene Ontology (GO) biological process and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of core co-regulated DEGs (adj. p < 0.05). (d) Heatmaps showing the expression of top candidate genes (Gpr83, Hrk, Ubash3a) in retinal and kidney tissues across groups.
Functional enrichment analysis was performed on these coregulated genes. The results showed significant enrichment in terms associated with cytokine activity and cytokine-mediated signaling pathways, highlighting the importance of inflammatory regulation in the therapeutic effect of ZJHY (Fig. 3c). Focusing on the most substantially altered pathway, we generated heatmaps to visualize the expression patterns of its constituent genes in both the kidney and retina. This analysis identified three genes—Gpr83, Hrk, and Ubash3a—that displayed a consistent trend: they were significantly upregulated under diabetic conditions and markedly downregulated following ZJHY treatment in both organs (Fig. 3d). This coherent pattern suggested that these genes may serve as key targets of the ZJHY formula.
Among these candidates, GPR83, a G-protein-coupled receptor, has emerged from the literature as potentially playing a pivotal role in metabolic and immune regulation, warranting further investigation[27,28,38].
ZJHY reverses diabetes-induced upregulation of GPR83 in retinal and renal tissues
-
To further clarify GPR83 expression in retinal and kidney tissues, we performed IHC experiments. Immunohistochemical staining revealed consistent patterns of GPR83 in the retinas and kidneys. Positive staining increased in retinal and kidney tissues of DM group rats, but decreased markedly following ZJHY intervention (Fig. 4a, b). We next assessed Gpr83 mRNA expression in retinal and renal tissues using quantitative RT-qPCR. The results showed that Gpr83 mRNA levels were significantly elevated under diabetic conditions and were effectively restored by ZJHY treatment, consistent with the sequencing data (Fig. 4c). Moreover, Western blot analysis confirmed that GPR83 protein expression followed the same trends as mRNA, with ZJHY reversing the diabetes-induced upregulation in both tissues (Fig. 4d, e).
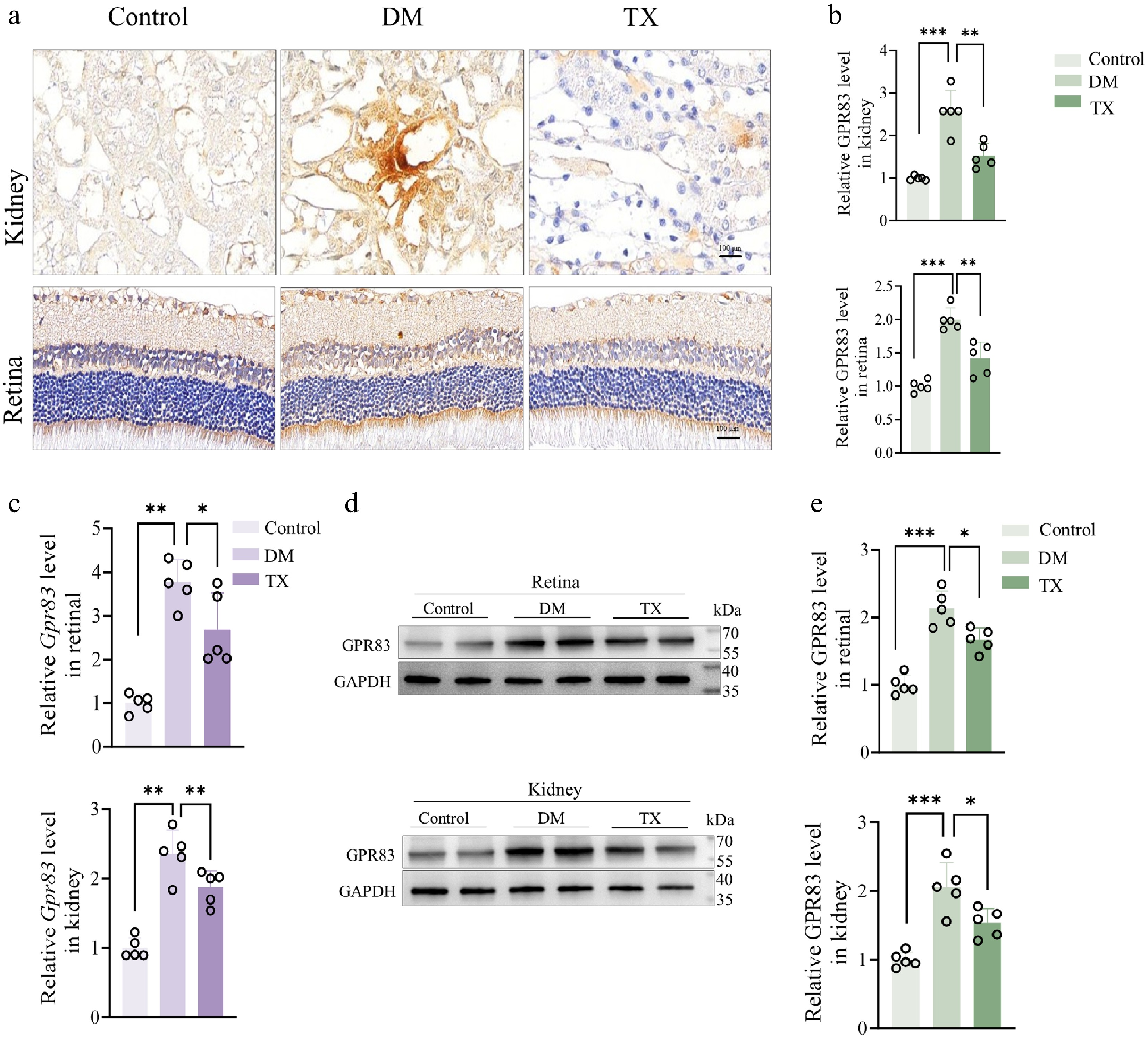
Figure 4.
ZJHY regulates the expression of GPR83 in the kidneys and retinal tissues in vivo. (a), (b) IHC staining of GPR83 in kidney and retinal tissues (scale bar: 50 μm) and quantification of positive staining intensity (n = 5 per group). (c) qPCR validation of Gpr83 mRNA expression in retina and kidney (n = 5 per group). (d), (e) Western blot analysis of GPR83 protein levels in retinal and kidney tissues (n = 5 per group). Data are presented as mean ± SD. Statistical significance was determined by one-way ANOVA with Tukey's post-hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
These findings collectively suggest that GPR83 may serve as a key therapeutic target mediating the ameliorative effects of ZJHY on diabetic retinal and renal lesions, though it remains unclear whether these effects are mediated through the ADM2-AKT downstream signaling mechanism.
ZJHY reverses diabetes-induced upregulation of ADM2 and AKT in retinal and renal tissues
-
Building on our observation that ZJHY treatment downregulated Gpr83 expression and that GPR83 may act through the ADM2-AKT axis, we next examined whether the protective effects of ZJHY in the diabetic retina and kidney were mediated via this pathway. To test this, as shown in Fig. 5a, diabetic conditions significantly upregulated Pi3k and Akt mRNA levels in both tissues compared to the control group, and this effect was notably reversed by ZJHY treatment. Interestingly, Adm2 expression, which was reduced under diabetic conditions, was also restored following ZJHY administration, suggesting that ADM2 may play a modulatory role in the AKT pathway. Consistent with the transcriptomic data, PI3K and phosphorylated AKT protein levels, as well as ADM2 expression, were elevated in the retina and kidneys of diabetic rats, and these increases were normalized after ZJHY treatment (Fig. 5b, c).
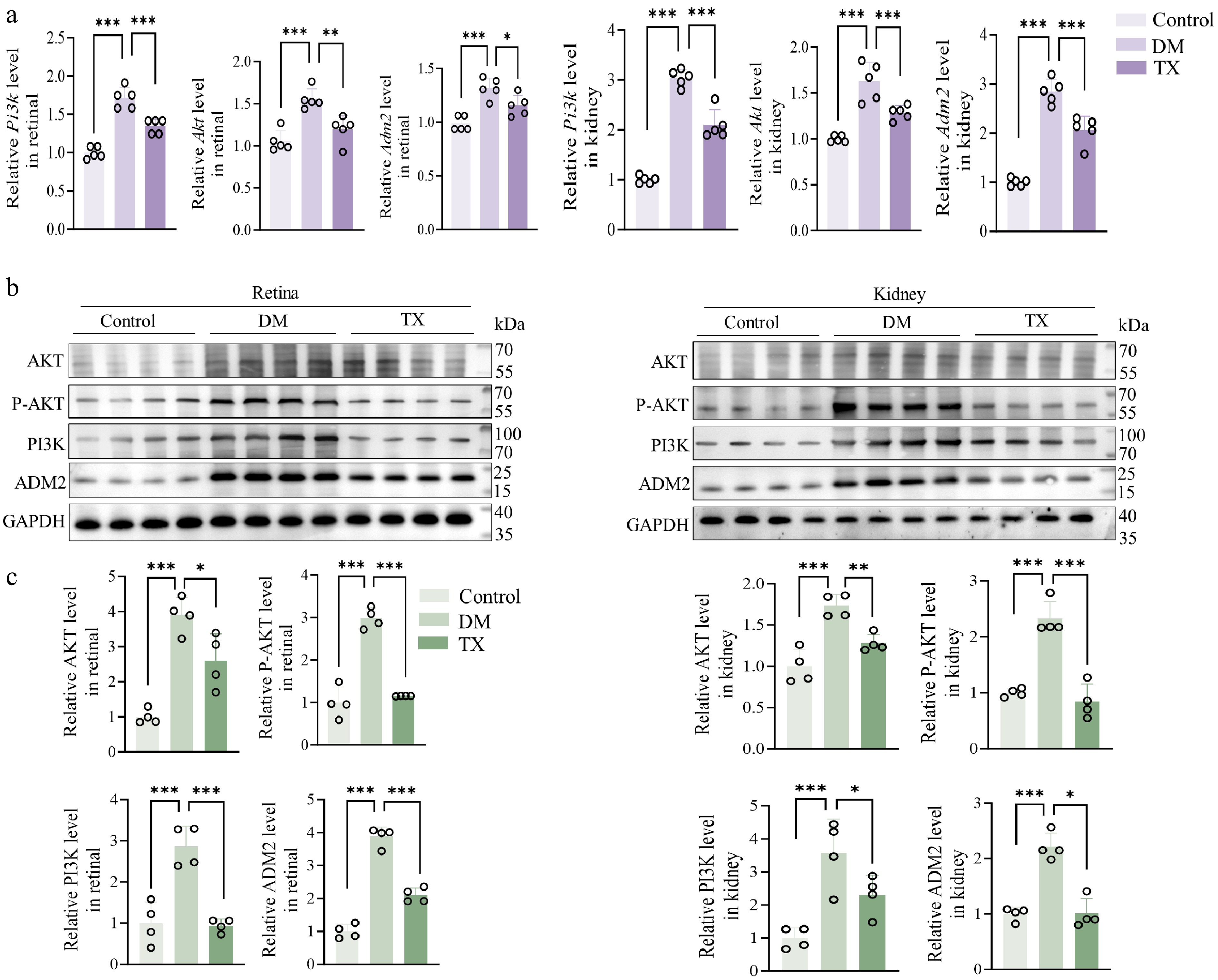
Figure 5.
ZJHY reverses diabetes-induced upregulation of ADM2 and AKT in retinal and renal tissues. (a) Relative mRNA expression of Pi3k, Akt, and Adm2 in retinal and kidney tissue measured by qPCR (n = 5 per group). (b), (c) Western blot analysis of PI3K, p-AKT, AKT, and ADM2 protein levels in retinal and kidney tissues (n = 4 per group). Data are presented as mean ± SD. Statistical significance was determined by one-way ANOVA with Tukey's post-hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
Overall, the expression of GPR83, ADM2, and AKT was co-upregulated in the retinal and renal tissues of diabetic rats, as shown in Figs. 4 and 5, and this increase was reversed by ZJHY treatment. These findings strongly suggest that ZJHY treats DR and DN through a mechanism involving GPR83, ADM2, and AKT. Nonetheless, the specific upstream and downstream relationships among these three factors require further validation.
ZJHY inhibits the secretion of pro-inflammatory factors and apoptosis of macrophages through the GPR83–ADM2–Akt axis
-
To further validate the functional role of the GPR83–ADM2–AKT axis, we conducted a series of in vitro experiments using BV2 cells and primary peritoneal macrophages. In BV2 cells, we first constructed a Gpr83 overexpression plasmid (Gpr83 OE) and synthesized small interfering RNAs (siRNAs) targeting Gpr83 (siGpr83) and Adm2 (siAdm2). Protein-level analysis confirmed successful overexpression and knockdown of the respective targets (Fig. 6a, b). The ZJHY extract was lyophilized and reconstituted for cell-based assays. To determine non-toxic working concentrations, we performed CCK-8 cytotoxicity assays. Based on the results, a concentration of 40 μg/ml was selected for subsequent experiments in microglial cells, while 30 μg/ml was used for primary peritoneal macrophages, as these concentrations exhibited no significant effect on cell viability (Supplementary Fig. S1d). We next examined whether GPR83 modulates the AKT pathway and ADM2 expression under high-glucose conditions in BV2 cells. Cells were treated with high glucose (Glu, 30 mmol/L) in the presence or absence of ZJHY, combined with Gpr83 overexpression or silencing. Western blot analysis showed that high glucose significantly upregulated AKT and ADM2 expression. Overexpression of Gpr83 further enhanced this upregulation, an effect attenuated by ZJHY treatment. Conversely, silencing Gpr83 suppressed the high glucose-induced upregulation of AKT and ADM2, and ZJHY exerted no additional effects under these conditions (Fig. 6c, d). These results indicate that GPR83 acts upstream of AKT and ADM2, positively regulating their expression, and that ZJHY specifically counteracts the effect of GPR83 overexpression. To further elucidate the role of ADM2 in this pathway, we knocked down Adm2 under high-glucose conditions and assessed the expression of the pro-apoptotic marker BAX and key inflammatory cytokines. Silencing Adm2 significantly reduced the protein expression of apoptotic markers (BAX, caspase-3) (Supplementary Fig. S1e) and the mRNA transcription of pro-inflammatory factors (Il-6, Il-1β) (Fig. 6e), suggesting that ADM2 contributes to high glucose-induced apoptosis and inflammation.
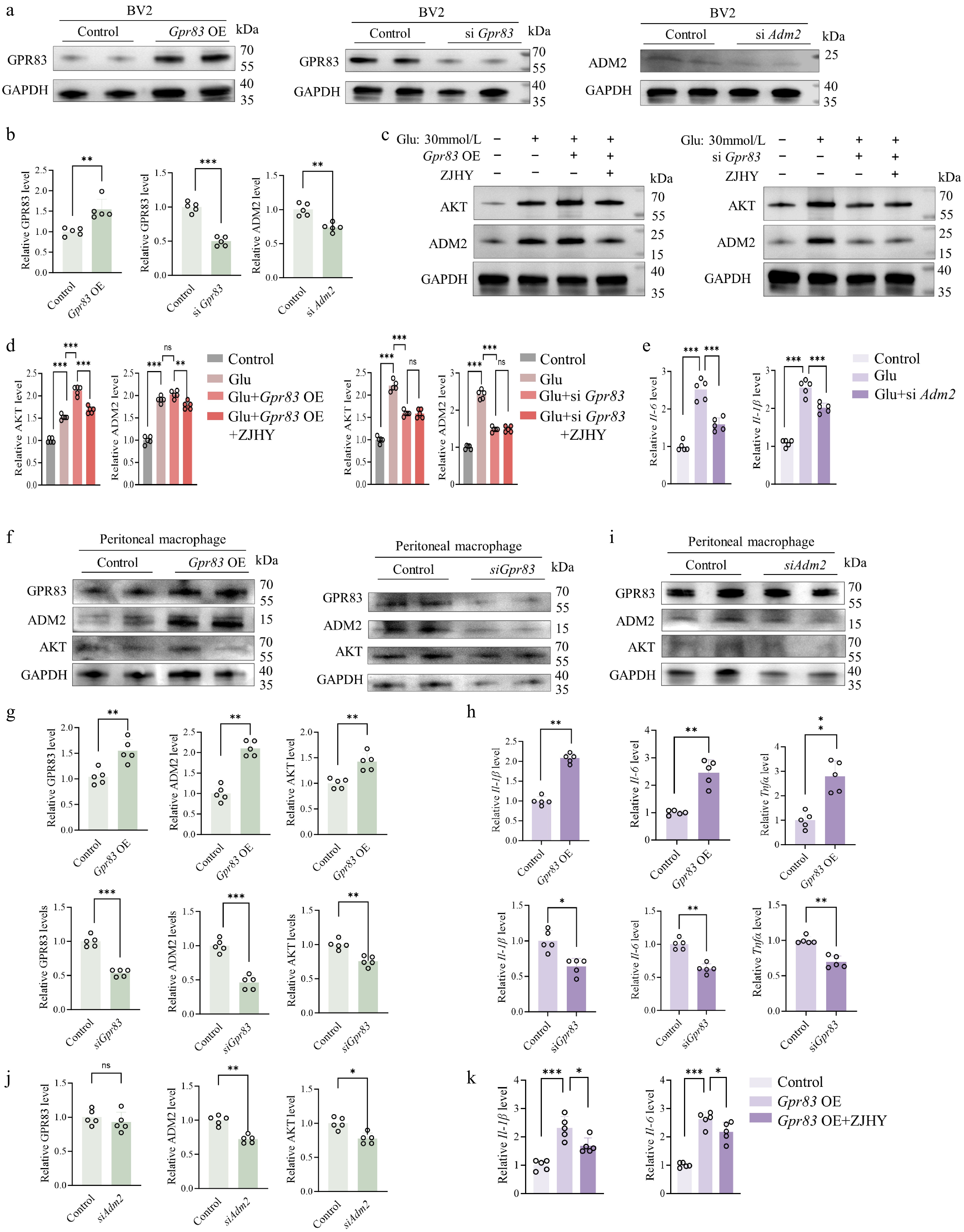
Figure 6.
ZJHY regulates the expression of the GPR83–ADM2–Akt axis in macrophages and inhibits the secretion of pro-inflammatory factors. (a), (b) Validation of Gpr83 overexpression and knockdown (siGpr83, siAdm2) efficiency in BV2 cells by Western blot (n = 5 per group). (c), (d) Western blot analysis of AKT and ADM2 protein levels in BV2 cells under ZJHY treatment, Gpr83 OE, or siGpr83 (n = 5 per group). (e) Relative mRNA expression of Il-6 and Il-1β in BV2 cells under Glu with siAdm2 transfection (n = 5 per group). (f), (g) Western blot analysis of GPR83, ADM2, and AKT protein levels in Peritoneal macrophage cells under Gpr83 OE, siGpr83, or siAdm2 (n = 5 per group). (h) Relative mRNA expression of Il-6, Il-1β, and Tnf-α in Peritoneal macrophage cells under Gpr83 OE, siGpr83 (n = 5 per group). (i), (j) Western blot analysis of GPR83, ADM2, and AKT protein levels in Peritoneal macrophage cells under siAdm2 (n = 5 per group). (k) Relative mRNA expression of Il-1β and Il-6 in Peritoneal macrophage cells under Gpr83 OE with ZJHY (n = 5 per group). Data are presented as mean ± SD. Statistical significance was determined by one-way ANOVA with Tukey's post-hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
To elucidate the functional significance of GPR83 in diabetes-associated inflammation and define the hierarchy of the GPR83–ADM2–AKT signaling cascade, we conducted a series of functional assays in primary peritoneal macrophages. Overexpression of Gpr83 significantly increased the protein levels of GPR83, ADM2, and AKT, as well as the mRNA levels of the pro-inflammatory cytokines Il-6, Il-1β, and Tnf-α, further supporting GPR83 as an upstream regulator of inflammatory responses and pathway activation; these findings indicate that GPR83 overexpression activates the GPR83–ADM2–AKT pathway and promotes pro-inflammatory cytokine release. Conversely, siRNA-mediated knockdown of Gpr83 markedly suppressed the protein levels of GPR83, ADM2, and AKT, along with the mRNA levels of Il-6, Il-1β, and Tnf-α (Fig. 6f–h). To determine the directional relationship within this axis, we transfected primary peritoneal macrophages with siRNA targeting Adm2. Western blot analysis revealed that siAdm2 significantly reduced ADM2 protein levels and concurrently decreased AKT expression, whereas GPR83 expression remained unchanged (Fig. 6i, j). These results confirm that ADM2 acts downstream of GPR83 and that GPR83 regulates AKT activation through ADM2, establishing a GPR83–ADM2–AKT signaling network that drives macrophage polarization and pro-inflammatory cytokine secretion. In Gpr83-overexpressing macrophages, ZJHY treatment effectively reduced the mRNA levels of Il-6 and Il-1β (Fig. 6k), restored anti-apoptotic BCL2 expression, and decreased pro-apoptotic BAX levels (Supplementary Fig. S1f), indicating that ZJHY counteracts GPR83-driven inflammatory and apoptotic signaling.
To further validate the functional significance of the GPR83–ADM2–AKT axis in inflammation, we examined the activation status of this signaling axis and cytokine expression at both the molecular and functional levels in primary peritoneal macrophages and BV2 microglial cells. In primary peritoneal macrophages, high glucose significantly upregulated the mRNA levels of Gpr83, Adm2, and Akt, an effect that was effectively reversed by ZJHY treatment. Notably, Adm2 knockdown did not significantly affect Gpr83 mRNA levels but markedly reduced Akt mRNA expression (Supplementary Fig. S2a–S2c), consistent with the protein-level observations in Fig. 6 and supporting the notion that ADM2 functions downstream of GPR83. Meanwhile, high glucose strongly induced the expression of the pro-inflammatory cytokines tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β), and this response was effectively suppressed by ZJHY treatment or transfection with siGpr83 or siAdm2. Under conditions of high glucose combined with Gpr83 overexpression, ZJHY reduced pro-inflammatory cytokine levels to below those observed with high glucose alone (Supplementary Fig. S2d–S2f), indicating a potent anti-inflammatory effect of ZJHY in a pathological context. In BV2 microglial cells, a similar functional pattern emerged: ZJHY treatment significantly attenuated the high-glucose-induced upregulation of TNF-α, IL-6, and IL-1β while upregulating anti-inflammatory cytokine IL-10 at the transcriptional level (Supplementary Fig. S2h–S2k), consistent with the findings in primary macrophages. Moreover, the molecular regulatory relationships within this signaling axis were thoroughly validated through gene overexpression and knockdown experiments in Fig. 6.
Collectively, these data demonstrate that ZJHY exerts anti-inflammatory effects by modulating the GPR83–ADM2–AKT axis in immune cell types relevant to retinal and renal pathology.
Together, these findings demonstrate that ZJHY mitigates inflammation and apoptosis by modulating the GPR83–ADM2–AKT axis (Fig. 7).
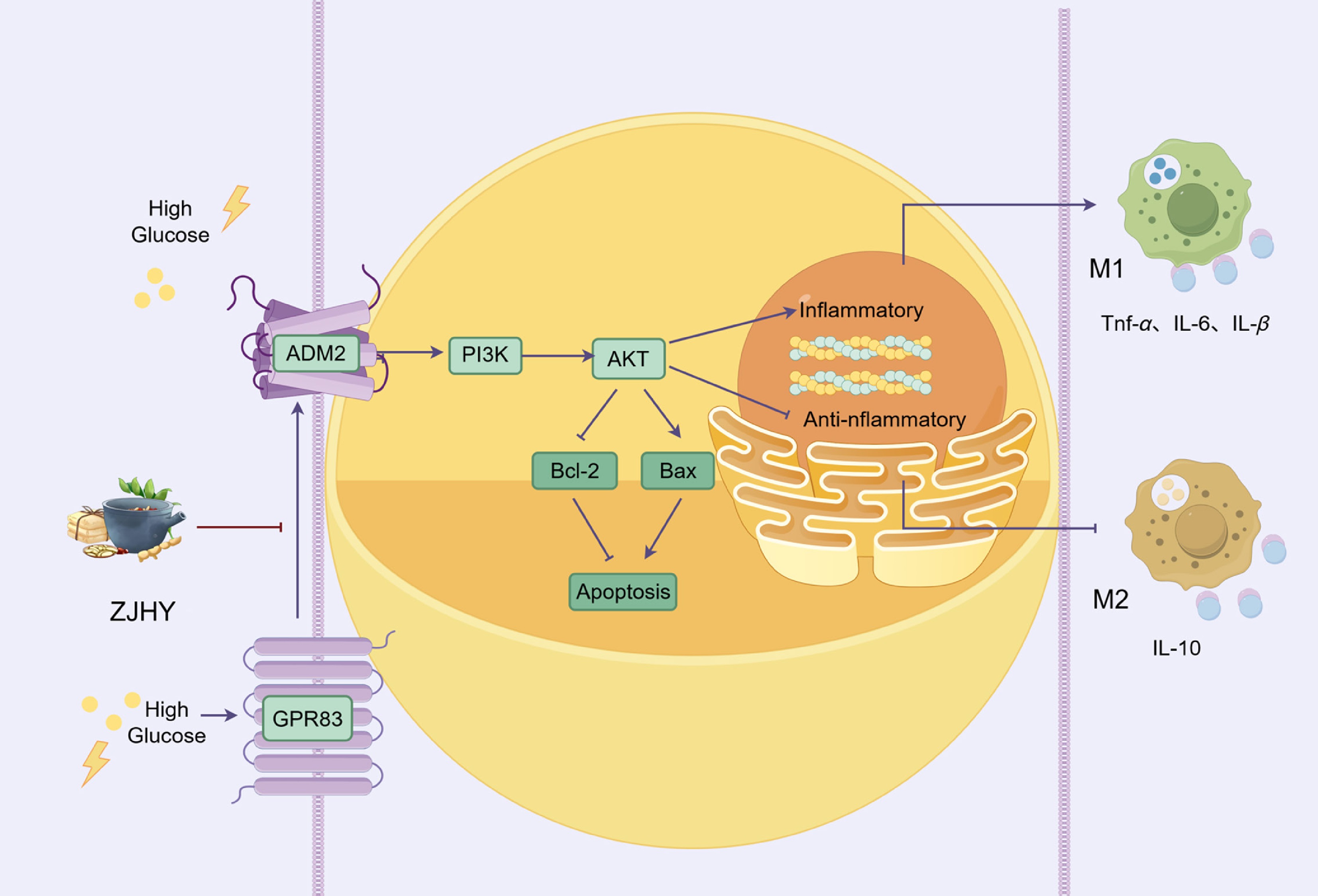
Figure 7.
ZJHY alleviates diabetic microvascular injury via ADM2–PI3K–AKT-mediated macrophage polarization and apoptosis suppression. The ZJHY formula mitigates high glucose-induced kidney and retinal damage by targeting the ADM2–PI3K–AKT signaling axis, which regulates macrophage polarization balance by shifting towards the anti-inflammatory M2 phenotype and suppressing pro-inflammatory M1-related cytokines like TNF-α, IL-6, and IL-1β, and modulating the Bcl-2/Bax ratio to inhibit excessive cellular apoptosis, ultimately alleviating tissue injury in diabetic microvascular complications.
Construction of a predictive model for diabetic retinopathy and nephropathy
-
To validate our findings in a human context, we integrated publicly available gene expression datasets for diabetic retinopathy (DR; GSE102485, GSE160306) and diabetic nephropathy (DN; GSE262793, GSE166239, GSE47183). After batch effect correction and data merging, principal component analysis (PCA) confirmed effective integration of the datasets. Differential expression analysis was then performed, with results shown in the volcano plot (Fig. 8a, b). To further explore co-expression patterns and identify disease-relevant modules, we applied Weighted Gene Co-expression Network Analysis (WGCNA) to the merged datasets. Modules significantly associated with DN and DR phenotypes were selected based on module-trait relationships. From these modules, hub genes were extracted to yield two refined gene sets: DR-Sig (diabetic retinopathy signature genes) and DN-Sig (diabetic nephropathy signature genes) (Fig. 8c, d). Subsequently, we intersected DR-DEGs, DN-DEGs, DR-Sig, and DN-Sig with a comprehensive immune-related gene set from the ImmPort database, identifying 11 immune-related candidate genes: Ednra, Ackr2, Trbc2, Ptgdr2, Glp2r, Ackr1, Gdf1, Acvr1c, Tnfsf9, Gpr83, and Shc4 (Fig. 8e).
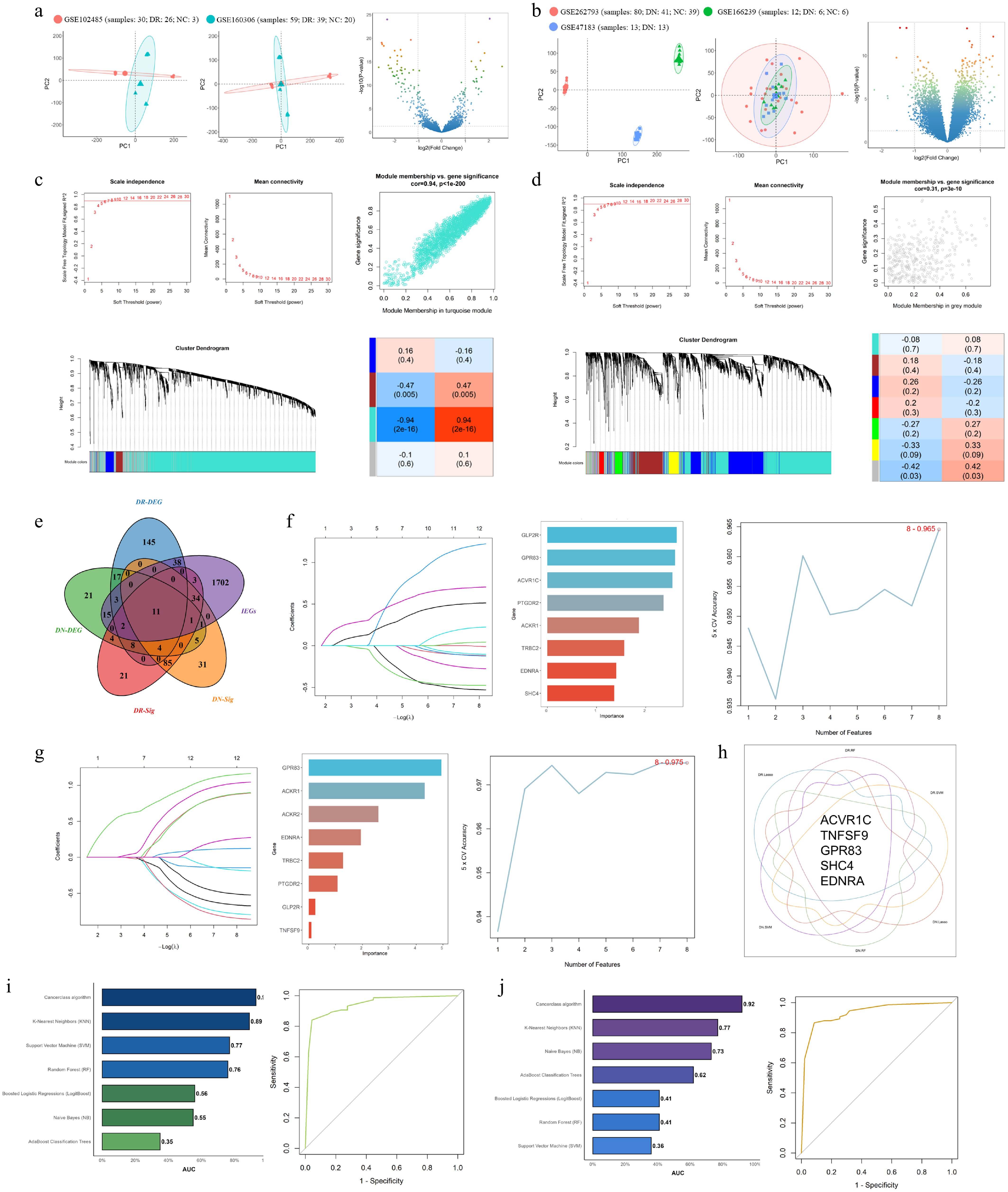
Figure 8.
A five-gene immune signature predicts diabetic retinopathy (DR) and diabetic nephropathy (DN) with high accuracy. (a), (b) Principal component analysis (PCA) plots demonstrating the integration of multiple transcriptomic datasets from patients with (a) diabetic retinopathy (DR), and (b) diabetic nephropathy (DN) after batch effect correction, and a Volcano plot displaying the differentially expressed genes. (c), (d) Weighted Gene Co-expression Network Analysis (WGCNA) showing module-trait relationships for DR and DN (|r| > 0.3). (e) Venn diagram illustrating the intersection of DR-DEGs, DN-DEGs, DR-Sig (hub genes from WGCNA), DN-Sig (hub genes from WGCNA), and immune-related genes (IRGs from ImmPort database) to identify 11 candidate genes. (f)–(h) Feature selection results from seven ML algorithms, including Naive Bayes (NB), Random Forest (RF), Support Vector Machine (SVM), AdaBoost Classification Trees (AdaBoost), Boosted Logistic Regression (Logit Boost), K-Nearest Neighbors (KNN), and the Cancerclass algorithm. (i), (j) Receiver Operating Characteristic (ROC) curves of seven machine learning models for DR (left, training: GSE160306; validation: GSE102485) and DN (right, training: GSE262793; validation: GSE47183). The Cancer class algorithm achieved the highest AUC (0.93 for DR, 0.92 for DN).
To prioritize the most predictive immune-related biomarkers, we applied three machine learning algorithms—Least Absolute Shrinkage and Selection Operator (LASSO), Support Vector Machine (SVM), and Random Forest (RF)—to evaluate associations between the 11 genes and disease status in the merged dataset. Genes consistently ranked among the top eight across all three algorithms were designated as core feature genes. This approach yielded five robust immune-related signature genes: Ackr1, Tnfsf9, Gpr83, Shc4, and Ednra (Fig. 8f–h). Using these five genes, we then constructed predictive models for both DR and DN. Seven ML algorithms were employed: NB, RF, SVM, AdaBoost, Logit Boost, KNN, and the Cancerclass algorithm. Models were trained separately on the DR dataset (GSE160306) and the DN dataset (GSE262793), then validated on independent datasets (DR: GSE102485; DN: GSE47183). Each model underwent 10 repetitions of 5-fold cross-validation for hyperparameter optimization, and predictive performance was assessed using the area under the AUC. The Cancerclass algorithm achieved the highest diagnostic accuracy for both diseases, with AUCs of 0.93 for DR and 0.92 for DN, surpassing all other algorithms (Fig. 8i, j). These findings underscore the robustness and generalizability of the five-gene immune signature for predicting diabetic retinopathy and nephropathy.
The five-gene immune signature (Acvr1c, Tnfsf9, Gpr83, Shc4, and Ednra) accurately predicts DN and DR. This predictive model offers new insights for the early screening and precise diagnosis of diabetic nephropathy and retinopathy, highlighting the central role of immune metabolic mechanisms in diabetic microvascular complications. Upon full validation, it may serve as a useful tool to support clinical decision-making.
-
This study identified a novel GPR83–ADM2–AKT signaling axis within the diabetic microenvironment that drives the shared inflammatory pathogenesis of diabetic retinopathy (DR) and diabetic nephropathy (DN) by regulating macrophage polarization. Animal experiments revealed that GPR83 and its downstream ADM2–AKT pathway were co-upregulated in the retinal and renal tissues of diabetic rats, and the multi-target protective effects of ZJHY on the retina and kidney were closely associated with downregulation of this axis, suggesting that targeting the GPR83–ADM2–AKT cascade may underlie the synergistic therapeutic efficacy of ZJHY against both DR and DN. Additionally, we constructed an AI-based diagnostic model incorporating five immune-related gene markers, including GPR83, for the early combined screening of DR and DN.
G-protein-coupled receptors (GPCRs) are key integrators of metabolic and immune signals, making them major targets for metabolic diseases[27,28,38]. GPR83 is a member of the GPCR family, yet its role in diabetic microvascular injury has not been investigated. This study is the first to demonstrate that GPR83 plays a core driving role in the pathological processes of diabetic retinopathy (DR) and diabetic nephropathy (DN). Transcriptomic, qPCR, Western blot, and immunohistochemical data consistently show that GPR83 is significantly upregulated in the retina and kidneys of diabetic rats, whereas ZJHY treatment normalizes its expression. In vitro experiments further confirmed that GPR83 overexpression aggravates high glucose-induced inflammation and apoptosis, whereas GPR83 silencing mimics the protective effect of ZJHY. These findings extend the function of GPR83 from being related to immune cell activation and metabolic homeostasis[27,28,38] to the fact that GPR83 is a key mediator linking chronic hyperglycemia with microvascular inflammation and organ damage, and its mode of action may be independent of blood glucose mechanisms. More importantly, the coordinated upregulation of GPR83 in both retinal and renal tissues—two distinct target organs—suggests it acts as a common driver of diabetic microvascular injury rather than an organ-specific mediator. Therefore, developing a broad-spectrum protective therapy targeting GPR83 for multiple diabetic microvascular complications aims to intervene in shared upstream mechanisms, addressing the unmet need for treatments directed at common pathological pathways.
Dysregulation of lactate metabolism elevates total lactate levels, alters macrophage polarization, impairs mitochondrial function, and worsens fibrosis and angiogenic abnormalities. These findings strongly suggest that modulating macrophages and improving the immune environment may serve as effective strategies for the combined prevention and treatment of microvascular complications such as diabetic DR and DKD. On this basis, we reveal for the first time that GPR83, an orphan receptor in the GPCR family, acts as a key hub regulating M1 macrophage polarization in diabetic retinal and renal tissues through the ADM2–AKT axis, supporting the broader concept of 'regulating macrophage immune metabolism'. It provides a brand-new and specific signaling pathway and drug target. Compared with PKM2 and the lactate-lactose axis[39], GPR83, as a membrane receptor, has greater potential for drug development, offering a new direction for the development of precise immunomodulatory therapies.
To verify the relationship between GPR83 and the downstream ADM2–AKT signaling pathway, we examined the gene transcription and protein expression levels of ADM2 and AKT in vivo, and these were consistent with the expression pattern of GPR83. They were co-upregulated in retinal and renal tissues from diabetic rats, and this effect was reversed by ZJHY treatment. This evidence provides the first link between GPR83 and ADM2–AKT in a specific pathological environment. In vitro functional experiments further supported the GPR83–ADM2–AKT signaling network. Overexpression or inhibition of GPR83 simultaneously amplified or attenuated the expression of ADM2 and AKT in BV2 microglial cells and primary peritoneal macrophages, as well as the expression of pro-inflammatory cytokines (IL-6, IL-1β, Tnf-α) in primary peritoneal macrophages, indicating that GPR83 acts upstream of ADM2–AKT. However, ZJHY reversed both the GPR83-induced enhancement of ADM2–AKT signaling in BV2 cells and the secretion of inflammatory factors in primary peritoneal macrophages caused by GPR83 overexpression. Meanwhile, in vitro experiments demonstrated that inhibiting ADM2 reversed the high-glucose-induced upregulation of AKT, production of pro-inflammatory cytokines (IL-6, IL-1β), and apoptosis in BV2 microglial cells. In primary peritoneal macrophages, ADM2 inhibition downregulated AKT expression but did not affect GPR83. These findings suggest that, under diabetic conditions, the GPR83–ADM2–AKT signaling network cooperatively drives macrophage polarization and promotes inflammatory cytokine secretion. GPR83 likely acts as an upstream component of this network, driving M1-type macrophage polarization via the ADM2–mediated AKT pathway, leading to the secretion of pro-inflammatory factors and contributing to pathological damage in diabetic retinopathy and nephropathy.
Consistent with these findings, transcriptional analysis confirmed that Adm2 knockdown selectively suppressed AKT expression without altering Gpr83 mRNA levels, further supporting the hierarchical organization of this signaling axis. ZJHY treatment also attenuated high glucose-induced pro-inflammatory cytokine expression in primary peritoneal macrophages and BV2 microglial cells, indicating an anti-inflammatory effect mediated through this pathway. Under GPR83 overexpression, ZJHY reduced pro-inflammatory cytokine levels below those observed with high glucose alone, suggesting a potent anti-inflammatory capacity that may be especially beneficial in pathological states involving heightened GPR83 signaling.
In BV2 cells, overexpression of GPR83 produced only a modest upward trend in ADM2 upregulation induced by high glucose, whereas GPR83 knockdown significantly reversed this effect, consistent with findings in primary macrophages. This result suggests that the regulatory relationship between GPR83 and ADM2 may involve cell-type-specific or saturation effects. Nevertheless, a clear upstream–downstream relationship was evident in both macrophage types, and the underlying mechanism warrants further study. This study also found that ADM2 exerts a pro-inflammatory effect in a chronic hyperglycemic microenvironment, which contrasts sharply with previous reports of ADM2 inhibiting M1 polarization in acute inflammation models[40,41]. In this study, the upregulation of endogenous ADM2 is directly involved in pro-inflammatory signal transduction, and silencing ADM2 can significantly inhibit the release of inflammatory factors and apoptosis induced by high glucose. This functional difference may be related to the different pathological microenvironments: the abnormal activation of GPR83 may cause the downstream signal of ADM2 to shift from the protective cAMP-PKA pathway to the pro-inflammatory PI3K–AKT–NF–κB pathway, suggesting that ADM2 is bidirectional in inflammatory regulation[42]. ZJHY treatment can effectively reverse the abnormal activation of this axis, further confirming its protective effect on DR/DN.
A striking observation from our study is that ZJHY does not directly modulate FBG or body weight in diabetic rats, yet it exerts important protective effects on target organs. These effects are reflected in improved serum lipid metabolism (reduced TG and TC), restored renal function (decreased UREA and CREA), and attenuated pathological damage in both the kidneys and retinas. Concomitantly, ZJHY effectively suppresses systemic inflammation by reducing serum IL-6 and IL-1β levels, which is consistent with its documented anti-inflammatory properties[43−46]. TCM formulas are distinguished by their ability to modulate multiple pathways simultaneously, and our results reinforce this paradigm. Unlike single-target drugs that often fail to halt the complex progression of diabetic complications, ZJHY acts on interconnected pathological processes to achieve multi-organ protection, including macrophage polarization, apoptotic signaling, and metabolic homeostasis[45,46]. Previous preclinical studies have reported the efficacy of TCM formulas in DR or DN individually, but our work is the first to demonstrate that ZJHY confers synchronous protection against both complications[43,44]. This broad-spectrum activity highlights the potential of ZJHY as a combined treatment drug for patients with DR and DN, independent of blood glucose control.
In addition to these experimental observations, it is worth noting that ZJHY has already demonstrated clinical efficacy. The ZJHY formula is not merely an experimental agent but a clinically established preparation developed by Professor Yanggui Yu, a renowned Traditional Chinese Medicine specialist in Guangdong Province. It has been approved as an in-hospital preparation (Z20071182) and has been widely used in the First Affiliated Hospital of Guangzhou University of Chinese Medicine for decades. Previous clinical studies have demonstrated that ZJHY combined with laser therapy significantly reduces macular retinal thickness and lowers the levels of inflammatory factors such as NOS, VEGF, and IL-6 in the aqueous humor of patients with diabetic macular edema[43]. Another trial reported that ZJHY combined with anti-VEGF intravitreal injection further decreases intraocular cytokine levels, reduces the frequency of injections, and improves visual outcomes[44]. Moreover, a randomized controlled trial showed that ZJHY combined with conventional treatment significantly improved renal function parameters (e.g., UACR, Ccr) and lipid metabolism in patients with early diabetic nephropathy[47]. Preclinical studies have also confirmed that ZJHY downregulates TNF-α and IL-1β expression in retinal tissues, alleviates blood-retinal barrier damage, and reduces retinal edema[48]. Network pharmacology analysis suggests that multiple active ingredients in ZJHY, such as quercetin and kaempferol, may exert synergistic anti-inflammatory effects, potentially through the AKT signaling pathway[45]. Collectively, these clinical and pharmacological findings support the translational relevance of our mechanistic discoveries and indicate that ZJHY is a promising multi-target therapy for both diabetic retinopathy and nephropathy, acting independently of glycemic control.
Together, these studies provide evidence from both mechanistic and genetic perspectives, supporting the existence of shared pathogenic pathways between DR and DN.
Although screening programs for diabetic retinopathy (DR) and diabetic nephropathy (DN) have enhanced early detection, current clinical markers frequently capture established structural or functional damage rather than the initial molecular events that initiate microvascular injury. Consequently, there remains an unmet need for non-invasive, mechanism-based biomarkers capable of identifying at-risk individuals before irreversible pathology develops[49,50]. Using WGCNA and ML algorithms, we identified a five-gene immune signature (Acvr1c, Tnfsf9, Gpr83, Shc4, Ednra) that predicts DN and DR with high accuracy (AUC = 0.92 for DN, AUC = 0.93 for DR). This signature captures early immune-metabolic dysregulation underlying microvascular complications, offering predictive value beyond conventional markers that primarily indicate end-organ damage[51−54]. Notably, GPR83 is a component of this diagnostic signature, establishing a direct link between disease pathogenesis and diagnosis. The other signature genes have well-documented roles in immune regulation and vascular biology: Acvr1c modulates fat metabolism and cell proliferation[55,56], Tnfsf9 is involved in T cell activation and macrophage polarization[57,58], Shc4 regulates intercellular communication through extracellular vesicles (EVs)[59], and Ednra is involved in regulating immune responses and inflammatory processes and is associated with immune cell infiltration, such as macrophages and T cells[60,61]. Collectively, this AI model, grounded in pathogenic mechanisms, represents a promising non-invasive tool for the early joint screening of DR and DN in clinical practice.
There are several key limitations in this study that warrant discussion. First, regarding the completeness of mechanism validation, a gap exists in our evidence chain: while cell-based experiments have demonstrated a functional association at this pathway node, in vivo genetic or pharmacological rescue experiments are lacking. Additionally, in vitro studies have yet to include a definitive rescue experiment to establish the upstream–downstream relationship, such as supplementing ADM2 in the context of GPR83 overexpression. This experiment will be completed in future work to rigorously confirm the causal sequence among the three components. Second, owing to the complex composition of the traditional Chinese medicine compound ZJHY, the direct active ingredients responsible for its protective effects remain unidentified. In the future, through methods such as chemical proteomics, components that directly interact with GPR83 can be screened out from the identified compounds[28], further providing a basis for the development of more precise therapeutic drugs. Finally, the five-gene diagnostic tags discovered based on bioinformatics in this study, although demonstrating excellent predictive performance in existing datasets, still need to be further verified in prospective, multi-center, large-scale real-world clinical cohorts[62] to evaluate their universality and stability in different populations. Additionally, while the functional effects of ZJHY on cytokine expression were consistent across primary macrophages and BV2 microglial cells, molecular readouts exhibited some variability between the two cell types, potentially reflecting cell-type-specific differences in post-transcriptional regulation or experimental conditions. Nonetheless, the core mechanistic conclusions are supported by multiple lines of evidence from both molecular and functional assays.
In conclusion, this study systematically explored the pathogenic role and therapeutic potential of the novel GPR83–ADM2–AKT signaling axis in diabetic complications. ZJHY may target this axis to treat DR and DN, addressing the current gap in therapies for diabetic microvascular complications independent of glycemic control. An AI diagnostic model based on a five-gene immune signature was also developed for the combined early screening of DR and DN. These findings deepen the understanding of the molecular mechanisms underlying diabetic microvascular injury and provide a theoretical foundation for translating GPR83-targeted strategies and ZJHY into clinical settings.
-
Animal care and experiments were approved by the Animal Ethics Committee of Guangzhou University of Chinese Medicine (Approval No. 20240105013, approved on January 5, 2024). The research followed the 'Replacement, Reduction, and Refinement' principles to minimize harm to animals.
-
The authors confirm their contributions to the paper as follows: conceived and led the study, performed biochemical experiments and animal studies, and wrote the original draft: Liu B; performed cell culture experiments, contributed to RNA sequencing, and participated in animal studies: Huang Z, Xiong Y; responsible for data statistics and typesetting: Lu C, Luo Y, Huang Y; supervised the study, provided resources, and critically revised the manuscript: Yu X. All authors reviewed the results and approved the final version of the manuscript.
-
This work was supported by the National Natural Science Foundation of China (Grant No. 82174441 and 82575149), the Scientific Research Project of the Administration of Traditional Chinese Medicine of Guangdong Province (Grant No. 20241092), and the Hunan Provincial Natural Science Foundation of China (Enterprise Joint Fund, Grant No. 2025JJ90254). We would like to sincerely thank the staff for maintaining the public database freely available to all users, which is undoubtedly helpful for the development of medicine.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/vns-0026-0026.
- Supplementary Table S1 Primer sequences used for quantitative real-time PCR in rats and mice.
- Supplementary Table S2 List of 85 characterized compounds.
- Supplementary Fig. S1 Effects of ZJHY on apoptosis and cell viability.
- Supplementary Fig. S2 ZJHY regulates the GPR83–ADM2–AKT axis and suppresses inflammatory responses in primary peritoneal macrophages and BV2 microglial cells.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu B, Huang Z, Xiong Y, Lu C, Luo Y, et al. 2026. ZJHY formula protects against diabetic retinopathy and nephropathy via the GPR83–ADM2–AKT signaling axis. Visual Neuroscience 43: e027 doi: 10.48130/vns-0026-0026 

ZJHY formula protects against diabetic retinopathy and nephropathy via the GPR83–ADM2–AKT signaling axis
- Received: 21 April 2026
- Revised: 18 May 2026
- Accepted: 27 May 2026
- Published online: 29 June 2026
Abstract: Diabetic retinopathy (DR) and diabetic nephropathy (DN) continue to develop even under optimal glycemic control, highlighting the urgent need for non-glucose-dependent protective strategies. Traditional Chinese Medicine offers a unique solution through its overall regulatory advantages of multiple components and multiple targets. This study explored the protective effect and mechanism of the 'Zi-Shen-Jian-Pi-Hua-Yu' (ZJHY) formula on DR and DN. In a diabetic rat model, ZJHY significantly alleviated retinal and renal damage independently of blood glucose control, improved lipid metabolism and renal function, and reduced systemic inflammation. This protective effect is related to reversing the upregulation of the orphan receptor Gpr83 induced by diabetes and restoring the function of the GPR83-ADM2-AKT signaling axis. In vitro mechanism verification indicates that under high glucose conditions, activation of the GPR83-ADM2-AKT axis can induce pro-inflammatory macrophage (M1) polarization and the secretion of inflammatory cytokines such as TNF-α, IL-6, and IL-1β, while knockdown of GPR83 can inhibit this process. In addition, based on hub genes including GPR83, we integrated multiple human datasets and applied machine learning methods to identify a robust five-gene immune marker (Acvr1c, Tnfsf9, GPR83, Shc4, Ednra), which can serve as an accurate tool for the joint screening and prediction of DR and DN.