









-
Pogostemon cablin, commonly known as patchouli or guang huo xiang, is a regionally significant medicinal plant predominantly found in tropical and subtropical areas. In China, patchouli is primarily cultivated in the provinces of Guangdong, Guangxi, and Hainan, spanning latitudes from 23° N to 18° S[1]. The optimal growth of P. cablin is heavily dependent on specific ecological conditions characterized by abundant sunlight, warmth, and adequate precipitation, all of which are essential for enhancing their growth. Patchouli serves as the primary source of patchouli oil, a valuable product extensively utilized in the fragrance, cosmetics, and pharmaceutical industries, functioning both as a raw material and a fixative[2,3]. Among its constituents, patchoulol is particularly noteworthy, playing a crucial role in defining its characteristic aroma[4]. Additionally, patchoulol has been shown to exhibit a range of biological activities, including immunomodulatory, anti-inflammatory, antioxidative, antimicrobial, and insecticidal effects[5]. As such, the cultivation of patchouli not only contributes significantly to regional economies but also holds substantial relevance in the fields of traditional and modern medicine due to its diverse therapeutic properties.
The growth and development of plants, including patchouli, are profoundly influenced by ecological factors, particularly light and temperature, which are critical determinants of photosynthetic efficiency. Research has established that light plays a pivotal role in various physiological and developmental processes in plants[6]. Over time, plants have evolved sophisticated sensory mechanisms to perceive and respond to light based on its intensity, direction, duration, and spectral quality[7]. These adaptations allow plants to optimize their growth strategies according to the prevailing light conditions, enabling them to harness solar energy effectively. Photosynthesis, the process by which plants convert light energy into chemical energy, is fundamental to their growth and development. This biochemical process not only generates essential carbohydrates for energy but also fuels the biosynthesis of secondary metabolites, which are crucial for a plant's survival and interaction with its environment[8]. Understanding these processes can be conceptualized through the lens of energy flow mechanisms within plant systems[9]. The interplay between photosynthetic source and sink dynamics further influences photosynthetic efficiency and the availability of energy and metabolites for growth, especially under varying light conditions, emphasizing the complex interactions within plant physiological processes[10].
The effects of light on plant growth and secondary metabolite production are often mediated through hormonal responses. Plant hormones, or phytohormones, such as auxins and jasmonates, play pivotal roles in regulating growth, development, and stress responses, with many often acting downstream of light signaling cascades[11]. The interaction between light signals and hormonal pathways has become a focal point in plant physiology research, especially in relation to the biosynthesis of economically and medicinally valuable compounds, such as patchouli oil. The accumulation of patchoulol has been shown to be significantly affected by light conditions during patchouli cultivation[12,13]. This highlights the importance of environmental management in optimizing patchouli oil yield and quality. Over the past century, research methodologies for unraveling the complexities of secondary metabolite biosynthesis have evolved from classical biochemistry approaches to more integrated molecular biology and omics techniques[14]. The application of techniques such as RNA sequencing (RNA-seq) has allowed for more detailed investigations into the transcriptional profiles associated with specific metabolic pathways. Recent studies have revealed that the expression levels of patchoulol synthase genes exhibit diurnal dynamics, with transcriptional abundance peaking in the evening[15]. This diurnal regulation underscores the significance of circadian rhythms in the context of secondary metabolite biosynthesis. Furthermore, exogenous application of methyl jasmonate (MeJA) has been reported to enhance patchoulol accumulation, indicating that jasmonate (JA) signaling plays a role in the regulation of patchouli oil biosynthesis[16,17]. Recent studies also suggest that light can stimulate JA biosynthesis, subsequently triggering signaling pathways that activate the expression of essential developmental genes, such as HY5, which is crucial for photomorphogenesis in Arabidopsis[18]. This intertwining of light perception and hormonal responses underscores the complexity of the regulatory networks governing plant metabolic processes.
Despite these advances, the intricate crosstalk between light signaling and JA signaling in relation to patchoulol biosynthesis within patchouli remains poorly understood. To bridge this knowledge gap, we systematically investigated the effects of light on patchoulol production and its coordination with JA signaling. This study aims to address this knowledge gap by examining patchoulol levels in response to various light conditions, including white, blue, red, far-red light, and darkness. Through an integrated approach combining RNA-seq analysis of samples cultivated under full-spectrum light and darkness, we explored the molecular mechanisms underlying light-induced patchoulol biosynthesis and its interaction with JA signaling. Our findings not only enhance the understanding of plant metabolic regulation but also provide practical insights for optimizing patchouli cultivation. This research ultimately contributes to improving both the yield and quality of patchouli oil, offering valuable strategies for the sustainable cultivation of this economically and medicinally significant plant.
-
Pogostemon cablin used in this work were the same species as that for genome sequencing, and originated in Yangchun City, Guangdong Province, China[19]. After the tissue culture state, patchouli plants are transferred to an artificial climate chamber (25 ± 2 °C). Two-month-old plants with similar growth states were used for experiments and treated in darkness and in white (90~120 μmol·m−2·s−1), blue light (40~60 μmol·m−2·s−1), red light (40~60 μmol·m−2·s−1), far-red light (7~9 μmol·m−2·s−1), for 1~2 weeks (12 h light/12 h dark).
RNA sequencing
-
RNA sequencing in this work was conducted by Biomarker Technologies (
www.biomarker.com.cn ). Total RNA was extracted from patchouli leaves. RNA concentration and purity were measured using NanoDrop 2000 (Thermo Fisher Scientific, Wilmington, DE, USA). RNA integrity was assessed using the RNA Nano 6000 Assay Kit of the Agilent Bioanalyzer 2100 system (Agilent Technologies, CA, USA). RNA sequencing libraries were prepared using NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB, USA) according to the manufacturer's instructions. Sequencing was performed on an Illumina platform (Illumina NovaSeq 6000) to generate paired-end reads. Raw sequencing data were processed using FastQC for quality control and adapter trimming. Clean reads were aligned to the reference genome (Accession No. GWHBAZF00000000)[19] obtained from China National Center for Bioinformation (www.cncb.ac.cn ) using HISAT2. Gene expression levels were estimated by fragments per kilobase of transcript per million fragments mapped, and differential expression analysis was performed using DESeq2. Genes with an adjusted p-value < 0.01 found by DESeq2 were assigned as differentially expressed. Differential expression analysis of two samples was performed using the edgeR. The FDR < 0.01 and fold change ≥ 2 was set as the threshold for significantly differential expression.Patchoulol extraction and GC-MS analysis
-
Approximately 0.2 g flesh samples from the second pair of fully expanded leaves were collected and immediately flash-frozen in liquid nitrogen. Then pulverized using a Retsch Mixer Mill MM400 cryogenic grinder with the following parameters: liquid nitrogen pre-cooling for 5~10 min, grinding parameters set to 25~28 Hz frequency, three cycles of 1 min grinding with 2 min intermediate cooling in liquid nitrogen. Subsequent steps followed standardized protocols: Addition of 1.5 mL ethyl acetate; ultrasonic extraction (40 kHz, 30 min), and centrifugation (12,000 × g, 10 min). Post-processing included dehydration with activated sodium sulfate, 0.22 μm membrane filtration, and storage in GC-MS vials. GC-MS analysis was conducted as previously described[19]. The extracts were detected using an Agilent 8890B gas chromatograph equipped with Agilent 5977B mass spectrometer. A HP-5MS column (0.25 mm × 30 m, 0.25 μm) was used to separate volatile components of 1.0 μL injection volume at a 20:1 split ratio. High-purity (99.999%) helium was taken as the carrier gas at a flow rate of 1.0 mL/min. The initial column temperature was 60 °C, holding for 2 min, then raised to 250 °C at a rate of 6 °C/min and held for 3 min. Full mass spectra were generated for metabolite identification by scanning within the m/z range of 40 ~ 550. Compounds were identified by comparing their retention times and mass spectra with those of standards. Electron impact ionisation at 70 eV was used for every measurement. The ion source and quadrupole analyser temperatures were fixed at 230 and 150 °C, respectively.
Jasmonates extraction and UPLC-MS/MS analysis
-
Some modifications were made to previous methods[20−22]. UPLC-MS/MS analysis was conducted by Wuhan MetWare Biotechnology Co. Ltd. (
www.metware.cn ). Firstly, fresh patchouli leaves were sampled, immediately frozen in liquid nitrogen, and ground into powder; 50 mg of them was transferred into 2 mL centrifuge tubes, and dissolved in 1 mL methanol/water/formic acid (15:4:1, V/V/V). Then, 10 μL internal standard (100 ng/mL), which was mixed by deuterium-labeled jasmonic acid (d5-JA), deuterium-labeled methyl jasmonate (d5-MeJA), deuterium-labeled gibberellin A4 (d2-GA4), deuterium-labeled gibberellin A19 (d2-GA19), deuterium-labeled gibberellin A20 (d2-GA20), deuterium-labeled gibberellin A53 (d2-GA53) and deuterium-labeled gibberellin A9 (d2-GA9), was added for quantification, as well as for error elimination to a certain extent. The mixture was vortexed for 10 min, then centrifugation for 5 min (12,000 rpm, 4 °C), the supernatant was transferred to clean microtubes, followed by evaporation to dryness and dissolved in 100 μL 80% methanol (V/V), and filtered through a 0.22 μm membrane filter for further UPLC-MS/MS analysis. The sample extracts were analyzed using an UPLC-ESI-MS/MS system equipped with Waters ACQUITY UPLC HSS T3 C18 column (2.1 × 100 mm, 1.8 μm). The solvent system consisted with water with 0.04% acetic acid (A) and acetonitrile with 0.04% acetic acid (B); gradient program started at 5% B (0−1 min), increased to 95% B (1−8 min), 95% B (8−9 min), finally ramped back to 5% B (9.1−12 min); flow rate, 0.35 mL/min; temperature, 40 °C; injection volume, 2 μL. Linear ion trap and triple quadrupole scans were acquired on a triple quadrupole-linear ion trap mass spectrometer (QTRAP), equipped with an ESI Turbo Ion-Spray interface. The ESI source operation parameters were as follows: ion source, ESI+/−; source temperature 550 °C; ion spray voltage, 5500 V (Positive), −4,500 V (Negative); curtain gas was set at 35 psi. Jasmonates were analyzed using scheduled multiple reaction monitoring (MRM). Data acquisitions were performed using Analyst 1.6.3 software (Sciex). Multiquant 3.0.3 software (Sciex) was used to quantify all metabolites. Mass spectrometer parameters including the declustering potentials (DP) and collision energies (CE) for individual MRM transitions were done with further DP and CE optimization. A specific set of MRM transitions were monitored for each period according to the metabolites eluted within this period.Chlorophyll fluorescence assay
-
Chlorophyll fluorescence parameters, as well as the light‐response curve were measured as per a previous method[23] using Imaging‐PAM M‐Series chlorophyll fluorescence system (Heinz‐Walz Instruments, Germany).
Quantitative real-time PCR
-
qRT-PCR assay was performed using 2 × SYBR Green Pro Taq HS Premix II (AG, China) on a LightCycler 480 system (Roche, Switzerland). The PCR procedure was set at 95 °C for 30 s, following 40 cycles of 95 °C for 10 s, 60 °C for 30 s. Pat18S was taken as the internal gene, and the relative gene expressions were calculated using the 2−ΔΔCᴛ method.
-
To investigate the relationship between patchoulol biosynthesis, plant hormone levels, and light exposure, we conducted experiments with patchouli leaves grown under various light conditions, including white light, blue light, red light, far-red light, and darkness (Fig. 1a). Volatile and non-volatile compounds were extracted and analyzed through gas chromatography and high-performance liquid chromatography. Our results indicated that the patchoulol content was maximized under white light conditions, surpassing levels observed in blue, red, and far-red light, as well as in darkness (Fig. 1b). This finding suggests that light is a critical factor influencing the synthesis of patchoulol in patchouli. Additionally, we employed liquid chromatography to assess the levels of eight different plant hormones (abscisic acid, Auxin, cytokinin, ethyne, gibberellin, jasmonates, salicylic acid, strigolactone) in patchouli leaves subjected to these varying light treatments (Supplementary Fig. S1). The data showed that exposure to light, compared to a dark environment, led to enhanced biosynthesis of jasmonates and many of their precursors (Fig. 1c; Supplementary Fig. S2). Notably, both patchoulol and JA accumulations exhibited significant increases in light, particularly under white light conditions (Fig. 1b, c). Light may play a significant role in modulating metabolic processes, as evidenced by the changes in patchoulol production under different light conditions. These findings provide a foundation for further investigation into the effects of environmental factors on the biosynthesis of economically important compounds in patchouli.
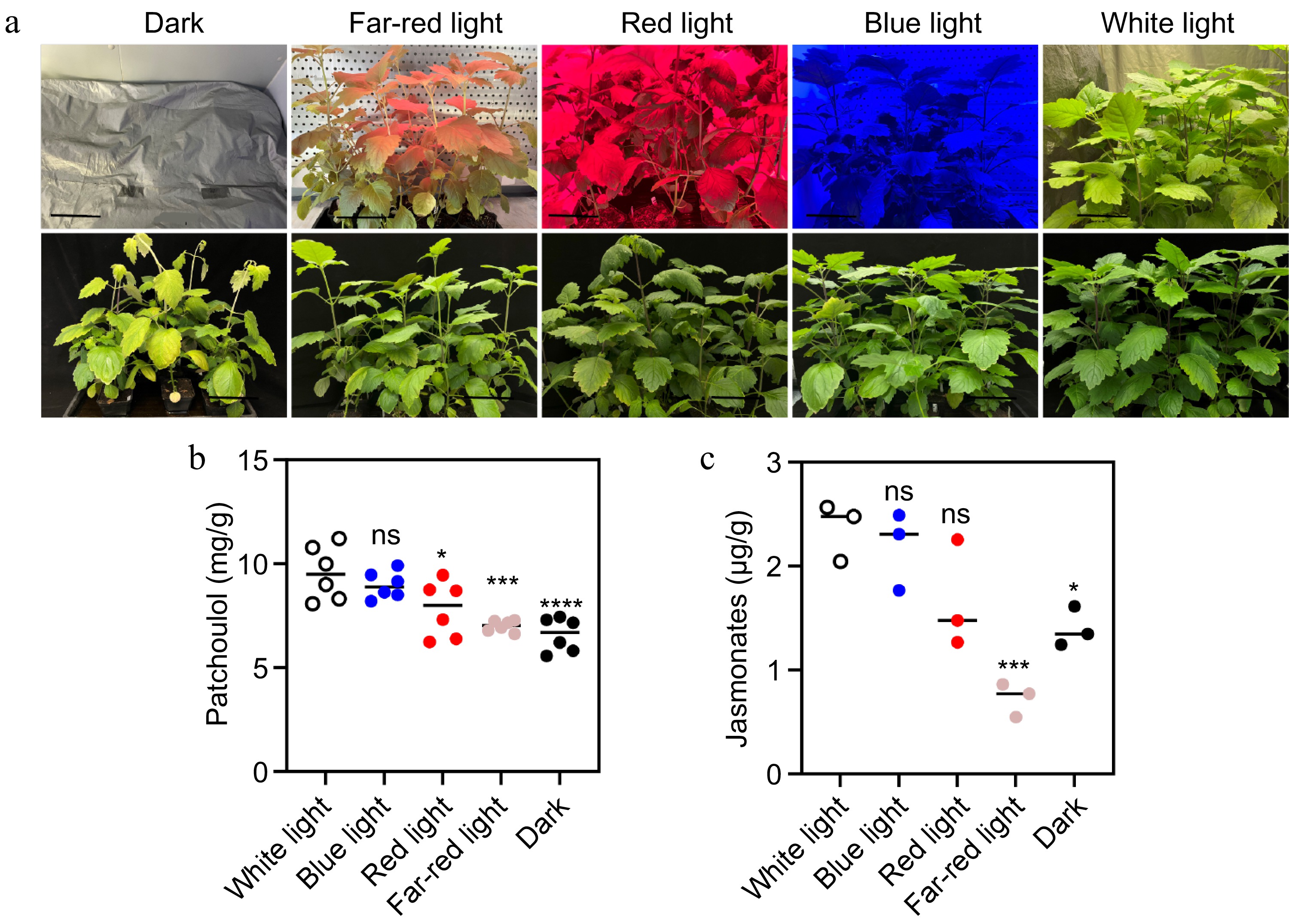
Figure 1.
Effect of light on patchoulol and JA accumulation. (a) Photographs of patchouli plants under different light conditions: dark, far-red light, red light, blue light, and white light. The top row shows the plants after two weeks of treatment. (b) Patchoulol accumulation across different light treatments. (c) JA levels under the same conditions. Black line length: 10 cm. * p < 0.05; ** p < 0.01; *** p < 0.001; ns, not significant. Student's t-test.
Gene expression analysis reveals the roles of light and JA signaling in regulating patchoulol biosynthesis
-
The synthesis of plant secondary metabolites is often regulated by plant hormones, with light acting as a crucial mediator in this process[11,24]. Based on our findings and previous research, we hypothesize that patchoulol biosynthesis is primarily regulated by the crosstalk of light signaling and JA signaling. To investigate this hypothesis, RNA-seq was conducted on patchouli leaves exposed to both light (full spectrum) and darkness. To account for potential circadian rhythm effects on gene expression, samples were collected at three time points: 0, 4, and 12 h post-exposure to the experimental conditions. The sequencing results indicated that 4,752 genes were differentially expressed in the 4-hour (4 h) group, while 6,195 genes were differentially expressed in the 12-hour (12 h) group (Fig. 2a). Functional enrichment analysis of these differentially expressed genes (DEGs) revealed significant associations with processes such as rhythmic processes, cell junctions, transporter activity, nucleic acid binding transcription factor activity, and electron carrier activity, with notable changes observed in both time points (Fig. 2a). In the 4-h group, a total of 2,244 DEGs were annotated using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database, with significant enrichment in pathways related to plant hormone signal transduction (178 genes), starch and sucrose metabolism (117 genes), circadian rhythm—plant (81 genes), photosynthesis (69 genes), terpenoid backbone biosynthesis (30 genes), and sesquiterpenoid and triterpenoid biosynthesis (10 genes) (Fig. 2b). Further analysis of DEGs in pathways associated with light, plant hormones, and terpenoid biosynthesis—specifically plant hormone signal transduction, circadian rhythm—plant, terpenoid backbone biosynthesis, and sesquiterpenoid and triterpenoid biosynthesis—revealed distinct expression trends. The light-exposed group exhibited a more rhythmic expression pattern across these pathways (Fig. 2c). This comprehensive analysis enhances our understanding of the regulatory mechanisms governing gene expression in response to light, providing valuable insights into the biosynthesis of secondary metabolites in patchouli. Specifically, our findings reveal that light signaling interacts with JA signaling to coordinately regulate patchoulol biosynthesis, as evidenced by the rhythmic expression patterns of genes involved in terpenoid biosynthesis and plant hormone signal transduction. These results suggest that light acts as a key environmental cue to synchronize metabolic pathways with circadian rhythms, optimizing the production of secondary metabolites. Furthermore, the identification of key genes and pathways, such as those related to terpenoid backbone biosynthesis and sesquiterpenoid biosynthesis, provides a foundation for future studies aiming to enhance the production of economically important compounds in patchouli and other medicinal plants.
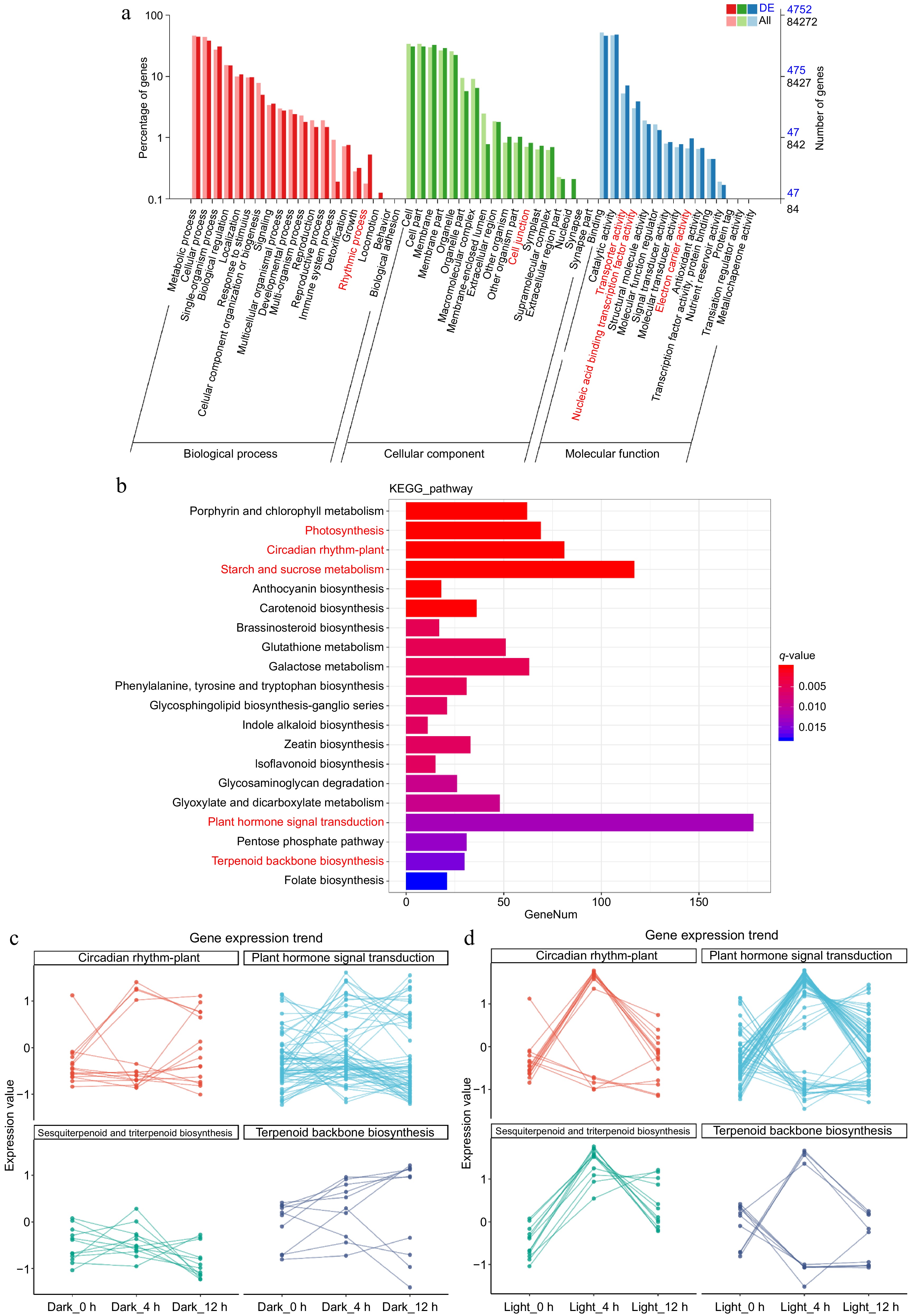
Figure 2.
Gene expression profile and function enrichment statistics of patchouli under dark and light conditions. (a) The bar chart displays the percentage of genes associated with various Gene Ontology (GO) terms across three categories: biological process (red), cellular component (green), and molecular function (blue). Notably, second-level function terms that are significantly enriched are highlighted in red. (b) The histogram presents the results of KEGG pathway analysis, showing the number of genes involved in various metabolic pathways. Key pathways, such as 'Photosynthesis' and 'Circadian rhythm—plant', are marked in red, indicating their significant enrichment. (c) The line graphs depict the gene expression trends for selected pathways, including 'Plant hormone signal transduction', 'Circadian rhythm—plant', 'Sesquiterpenoid and triterpenoid biosynthesis', and 'Terpenoid backbone biosynthesis'. The expression values are plotted across different time points under dark and light conditions, illustrating the dynamic changes in gene expression in response to light exposure.
Effect of light on photosynthetic performance
-
As mentioned above, we found a strong correlation between light, patchoulol and jasmonates. Considering the importance of photosynthesis in plants, the photosynthetic efficiency under different light conditions were analyzed. Based on chlorophyll fluorescence imaging technology, we detected the photosynthetic parameters of photosystem II in patchouli leaves. These parameters can help us monitor the health status of the plant photosynthetic system, the state of energy conversion, and the state of light damage to plant leaves. It can be seen from the light curves of patchouli leaves with different light treatments that F0, the minimal fluorescence in the dark adaptation state, varied between 0.05 and 0.10 — small difference between groups (Fig. 3a). While the maximum quantum yield of PS II photochemistry Fm, of all groups, decreased abruptly at the moment the light started, and continued to decrease, then the rate of reduction decreased significantly and became smaller and smaller; white light with full spectrum stayed at the top during this process (Fig. 3b). Y(II) values were highest in dark conditions but dropped under all light treatments, especially in blue light, indicating photoinhibition; It was also observed that the actual photosynthetic efficiency did not exactly coincide with the potential maximum photosynthetic efficiency, even though their general trends were similar (Fig. 3c). In terms of energy utilization, it seems that white, blue, and far-red light had higher photosynthetic efficiency. Y(NPQ) increased under light exposure, with the highest values observed in blue light, reflecting enhanced photoprotection (Fig. 3d). Overall, these findings highlight that while light promotes photosynthesis, specific conditions, particularly blue light, can detrimentally affect photosynthetic efficiency compared to dark conditions.
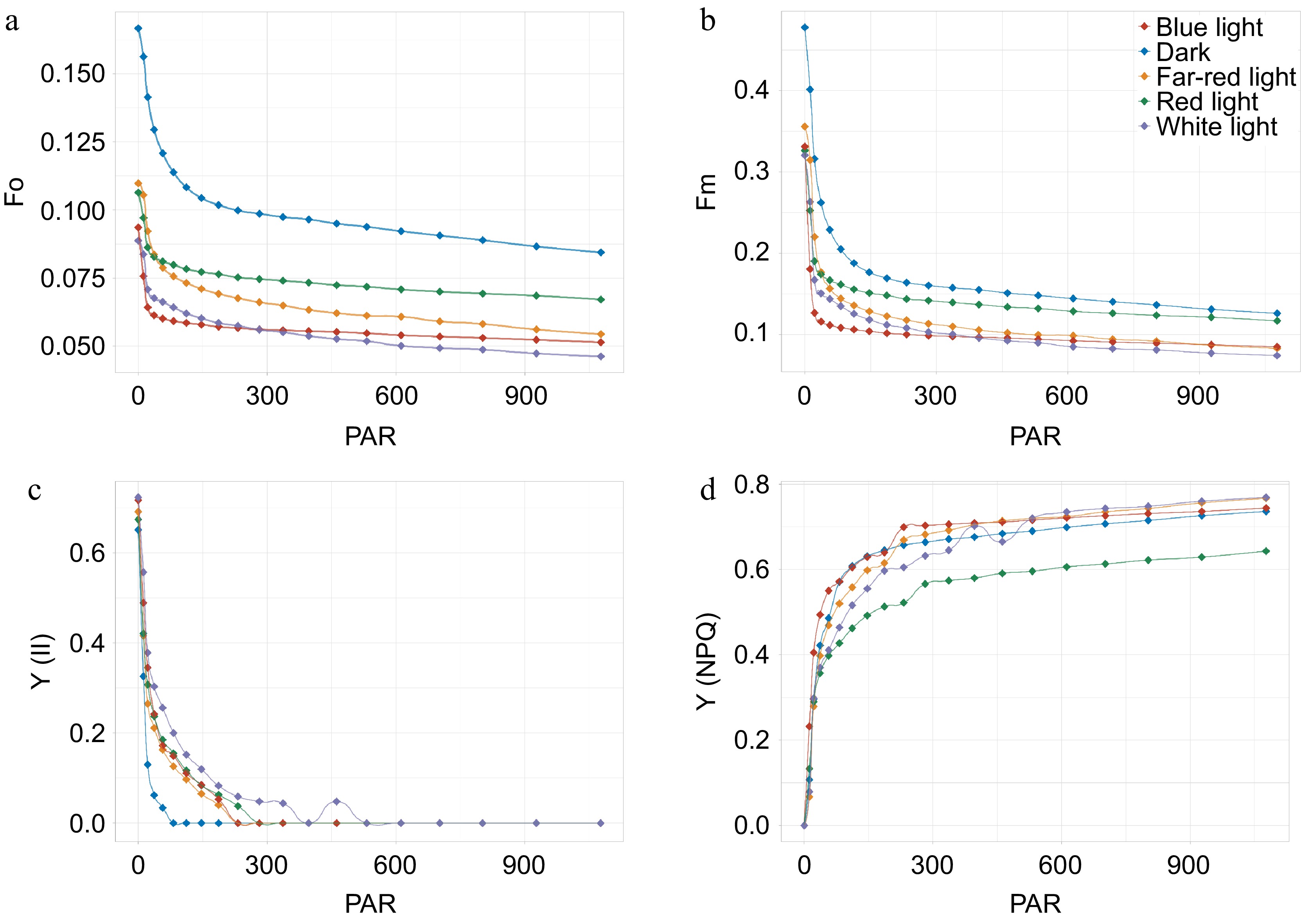
Figure 3.
Photosynthetic performance of patchouli plant in different light conditions. (a)−(d) Illustrate the light response curves for different photosynthetic parameters. F0, minimal fluorescence in the dark adaptation state; Fm, the maximum quantum yield of PS II photochemistry measured in the dark adaptation state; Y(II), PS II actual photosynthetic efficiency, effective quantum yield of photochemical energy conversion in PS II; Y(NPQ), Quantum yield of regulated non-photochemical quenching. X-axis refer to photosynthetically active radiation (PAR).
Light modulates the expression of enzyme genes related to patchoulol biosynthesis
-
Mevalonate (MVA) and methylerythritol phosphate (MEP) are main pathways of terpenoid backbone biosynthesis. The expression of key enzyme genes involved in MVA and MEP pathways, as well as patchoulol biosynthesis, was assessed in patchouli under varying light conditions. The results demonstrated no effect or repression of light on the expressions of acetoacetyl-CoA thiolase (AACT), 3-hydroxy-3-methylglutaryl-CoA synthase (HMGS), 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), mevalonate kinase (MVK), phosphomevalonate kinase (PMK), mevalonate diphosphate decarboxylase (MVD), isopentenyl diphosphate isomerase (IPPI) and farnesyl diphosphate synthase (FPPS); while under the same conditions, it was activated when it came to 1-Deoxy-D-xylulose-5-phosphate synthase (DXS), 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR), 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase (MCT), 4-(cytidine 5'-diphospho)-2-C-methyl-D-erythritol kinase (CMK), 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MDS), 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate synthase (HDS), 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase (HDR) and patchoulol synthase (PTS) (Fig. 4). For enzymes in the MEP pathway, it might be interpreted that plastid is the compartment which these enzymes work (Fig. 4). Notably, light exposure exerted divergent regulatory effects on the two terpenoid precursor pathways: the cytosolic MVA pathway genes remained either unaffected or showed mild suppression (Fig. 4), whereas their plastid-localized MEP pathway counterparts, particularly DXS and PTS, exhibited potent activation under white light. Consistent with the transcriptional activation PTS in the MEP pathway (Fig. 4), GC-MS analysis revealed a higher patchoulol level under white light (Fig. 1b), demonstrating the close link between gene expression and metabolism. The biosynthesis of patchoulol is catalyzed by PTS, and the MVA and MEP pathways may exert effects via redox-mediated regulation and metabolic flux compensation.
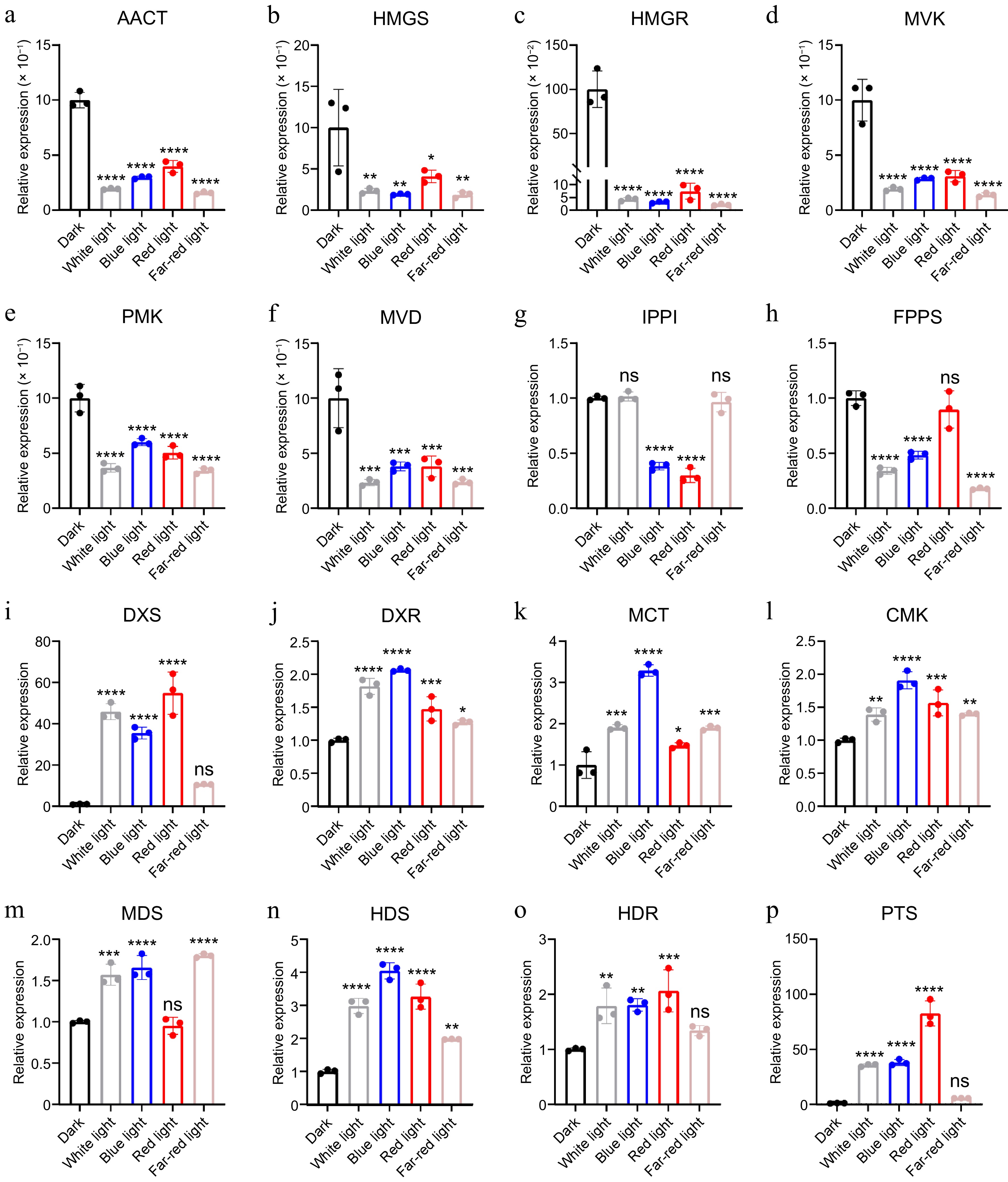
Figure 4.
Relative expression of enzyme genes in patchoulol biosynthesis. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; ns, not significant. Student's t-test.
Light promotes the biosynthesis of JA precursors
-
JA and its derivatives, collectively called jasmonates (JAs), are lipid-derived signaling compounds; JAs are initially formed from α-linolenic acid (α-LeA) esterified in chloroplast membranes[25]. To further investigate the relationship between light signals and jasmonates biosynthesis, we assessed the levels of JA precursors and other jasmonates within the JA biosynthesis pathway. Our results demonstrated that exposure to light significantly enhanced the biosynthesis of these JA precursors. Notably, the levels of 12-oxo-phytodienoic acid (OPDA) were markedly higher in light-treated patchouli compared to those maintained in the dark (Fig. 5a). Additionally, we observed slight increases in the levels of both OPC-4 and OPC-6 in the light-exposed groups; however, these differences did not achieve statistical significance (Fig. 5b). Moreover, presence of light was found to stimulate the biosynthesis of MeJA and jasmonoyl-isoleucine (JA-Ile), both of which are critical components in the JA signaling pathway. Although both compounds were detected at low concentrations (less than 5 ng/g), their biosynthesis was significantly enhanced by light exposure (Fig. 5c). Collectively, these findings underscore the intricate role of light in regulating JA biosynthesis and its associated signaling pathways, emphasizing the importance of gene expression regulation in these processes.
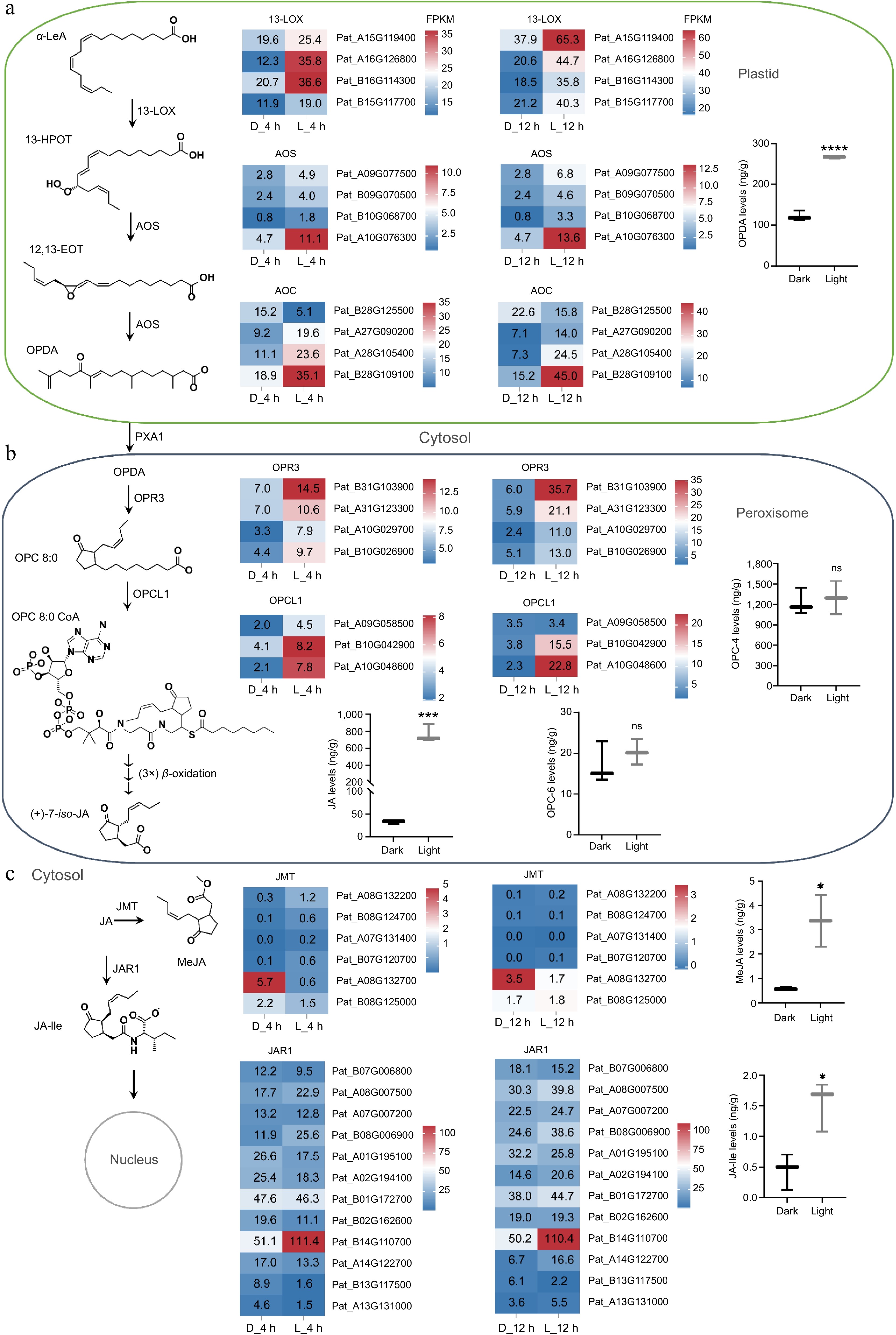
Figure 5.
Gene expression profiles and product accumulations in the jasmonates biosynthesis pathway. (a) Accumulation of OPDA and expression of related enzyme genes in plastids. (b) Accumulation levels of OPC-4 and OPC-6 and their synthetases gene expression in peroxisomes. (c) Accumulation levels of MeJA and JA-Ile and expressions of their synthetases in the cytosol. * p < 0.05; **** p < 0.0001; ns, not significant. Student's t-test.
Identification of candidate genes associated with light-induced patchoulol biosynthesis
-
To explore the correlation between light and patchoulol biosynthesis, we performed weighted gene co-expression network analysis (WGCNA) and identified several co-expression modules. The numerous regions in the network heatmap illustrated differences in gene expressions (Fig. 6a). All genes were clustered into two groups, dark and light, with a minimum height value of 0.25 and a minimum module size of 30 (Fig. 6b). Among these identified modules, the cyan module, which is characterized by a strong positive correlation with light conditions (0.93) and a strong negative correlation with dark conditions (−0.72), was selected for further analysis due to its relevance to light signaling and the presence of key genes such as PatHY5(Fig. 6c, d). Further analysis found 2,509 genes in this module. Genes in six expression patterns were proved with higher credibility (p > 0.05), and HY5 was in one of them (Fig. 6e). Finally, fewer genes were identified, assisted with gene function annotation. These potential genes were further used for correlation analysis with HY5, and the result showed negative correlations between HY5 and CRY1, PHYA, PIF4, PIF7, TCP14, IPPI, etc., while positive correlations between HY5 and TIFY6A, NAC002, NAC083, NES, GES, etc. (Fig. 6f). This detailed analysis enhances our understanding of the intricate relationships between gene expression, light conditions, and the biosynthesis of secondary metabolites in patchouli.
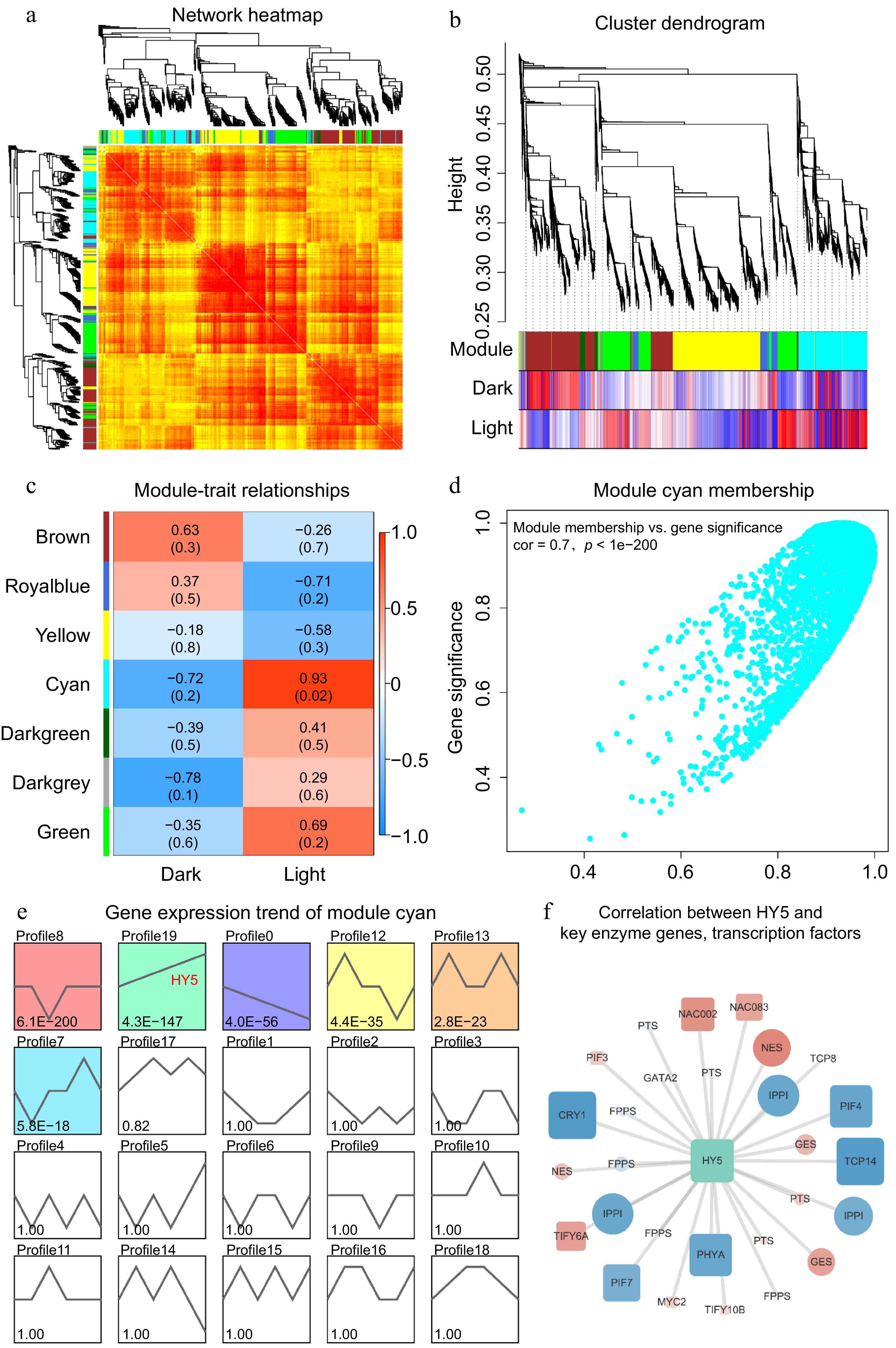
Figure 6.
(a) The network heatmap illustrates the gene expression profiles of samples from darkness and light groups; not all genes are displayed. Darker red shades indicate smaller dissimilarities between genes. Symmetrical phylogenetic clustering trees are shown on the left and top sides. (b) The clustering dendrogram provides a visual representation of the hierarchical clustering of genes, with a minimum height value of 0.25 and a minimum module size of 30. This dendrogram further emphasizes the separation of genes into dark and light groups. (c) The module-trait plot demonstrates the correlations between different gene modules and traits. The intensity of the colors indicates the strength of the correlation, with darker red representing stronger positive correlations and darker blue indicating stronger negative correlations. (d) The scatter plot reveals the relationship between module membership and gene significance for the cyan module. A higher correlation coefficient and smaller P value indicate that the module's gene members are more representative of the module's characteristics. (e) The gene expression trends of module cyan members are displayed, depicting various expression profiles. The smaller the P value in these profiles, the more reliable the trends are. Notably, the profile associated with HY5 is highlighted. (f) The correlation network illustrates the relationships between HY5 and key enzyme genes as well as transcription factors. Positive correlations are marked in red, while negative correlations are marked in blue. The size of the nodes reflects the strength of the correlation with HY5, providing insights into the regulatory network involved in patchoulol biosynthesis.
HY5 functions as a key signal integration point that coordinates the light and hormone signaling pathways[26]. For further illuminating the roles PatHY5 played in patchoulol biosynthesis, PatHY5 sequence was analyzed. Firstly, amino acid sequences of PatHY5 were aligned with that of AtHY5, which has been studied pretty clearly so far. They had a common bZIP HY5-like domain, being the members of the bZIP superfamily, and high homogeneity in other functional fragments (Supplementary Fig. S3a). Then its tertiary structure model was built using SWISS-MODEL tool, and one of these predicted models built on AlphaFold DB model of HY5_SOLLC, transcription factor HY5 from Solanum lycopersicum, showed 83.97% sequence identity (Supplementary Fig. S3c). It was also compared with the AtHY5 (At5g11260) structure from AlphaFold (Supplementary Fig. S3b). Evolutionarily, PatHY5 was proved to be a closer genetic relationship to Andrographis paniculata and Salvia miltiorrhiza than other medicinal plant species, such as Andrographis paniculata, Artemisia annua, Cannabis sativa, Carthamus tinctorius, Dendrobium catenatum, Gentiana rigescens, Isatis tinctoria, Salvia miltiorrhiza, and so on (Supplementary Fig. S3d). Moreover, position-specific ultra-short motifs from above aligned homologous sequences were found using kpLogo tools. The analysis reveals that at position 1, glutamine (Q) exhibits a high −log10(P) value of 12.6, indicating its significant role as a functional residue. Additionally, histidine (H) at position 12 shows a notable significance, suggesting its potential involvement in enzymatic activity or ligand-binding interactions. The lower panel of the kpLogo indicates that proline (P) and alanine (A) at positions 3 and 4, respectively, are highly represented, suggesting structural conservation. Conversely, leucine (L) and glycine (G) at positions 15 and 20 are frequently observed, indicating their importance in maintaining protein stability and folding. The clustering of specific amino acids may indicate critical binding sites or active regions within the protein structure. Understanding these motifs is vital for deciphering the molecular mechanisms underlying protein function and interaction networks.
-
In this study, we observed a correlation between light exposure, JA levels, and patchoulol biosynthesis. However, the specifics of the crosstalk between light and JA signaling remain unclear, as does the precise function of HY5 within this interaction. These unresolved questions are the focus of our ongoing research. Previous studies have shown that the PatJAZ protein interacts with the PatMYC2 transcription factor and that overexpression of PatJAZ can inhibit patchoulol accumulation[16]. In our research, we utilized a dual-luciferase reporter gene assay to assess the effects of PatHY5 as a transcription factor on the promoters of patchoulol synthetase (Supplementary Fig. S4). However, it remains uncertain whether these effects are direct. Additionally, while we speculated on a potential protein interaction between PatHY5 and PatMYC2, this was not validated through yeast two-hybrid assays. Neither did that between PatHY5 and PTS promoters. A plausible hypothesis is that PatMYC2 may first regulate the promoter of PatHY5 after being released from the PatJAZ protein complex in response to JA signaling triggered by light-induced JA accumulation. Once activated, PatHY5 could then influence patchoulol synthetase, either directly or indirectly, thereby promoting patchoulol synthesis. While it appears that PatHY5 plays a central role in the light-JA crosstalk that drives patchoulol biosynthesis, it is important to note that this intricate regulation likely involves multiple factors within the complex metabolic network of plants.
Role of jasmonates in light-induced patchoulol biosynthesis: exploring alternative signal molecules in JA signaling
-
In this study, we observed that light significantly promoted patchoulol accumulation, a process that correlates with enhanced levels of jasmonates. Previous research has indicated that the exogenous application of MeJA can stimulate patchoulol production in patchouli[16], suggesting that the light-mediated regulation of patchoulol biosynthesis may indeed involve JA signaling pathways. One key signal in this pathway is jasmonoyl-isoleucine (JA-Ile), which facilitates the binding of the JAZ protein to the SCFCOI1 complex; this interaction leads to the degradation of JAZ proteins and the subsequent release of MYC2[27]. Our data indicate considerable variability in the total JA levels between dark- and light-treated patchouli (Fig. 1c). However, the difference in JA-Ile levels was relatively minor (Fig. 5c), raising questions about the specific role JA-Ile plays in this signaling mechanism. While it is plausible that JA-Ile could still be a contributing factor, given that plant hormones can exert significant physiological effects even at low concentrations[28]. Recent research manifests that other compounds within the JA signaling cascade may also be influential. For instance, there is emerging evidence that other JA derivatives, such as OPDA-Ile[29], and different forms of JA like jasmonic acid or MeJA[30]. Thus, the specific JA signaling molecules implicated in the regulation of patchoulol biosynthesis warrant further investigation. Understanding the full scope of JA involvement—particularly the potential contributions of these alternative signals—will be critical for delineating the complex regulatory networks that govern patchoulol production in response to light. Future studies should aim to clarify these relationships, paving the way for more targeted approaches in enhancing patchoulol biosynthesis through light and JA signaling interactions.
Transcriptional regulation of terpenoid metabolism: the role of HY5
-
Transcriptional regulators play a critical role in the biosynthesis of patchoulol, as evidenced by numerous studies that have identified key regulatory genes involved in this process[31]. The successful assembly of the patchouli genome at the chromosome level has significantly advanced our understanding of these regulatory mechanisms the assembly of patchouli genome at the chromosome level contributes to this research process[19]. Photoreceptors in plants detect light conditions, activating light signaling pathways that orchestrate a multitude of biological processes. These processes encompass protein-protein interactions, transcriptional regulation of various transcription factors, and the expression of light-responsive genes, all of which collectively influence plant growth, development, and the biosynthesis of secondary metabolites[6]. One of the pivotal regulators in this context is HY5, which operates at the intersection of multiple signaling pathways, including those governing terpenoid biosynthesis[32]. HY5 is known to bind to several genes implicated in the terpene biosynthetic pathway, mediating their transcription in response to light signals. For instance, in Artemisia annua, the transcription factor AaHY5 displays distinct regulatory patterns for artemisinin biosynthesis under varying light conditions, directly activating key biosynthetic genes like ADS and CYP71AV1[33]. In contrast, our study reveals that PatHY5 in patchouli likely functions through an indirect regulatory cascade, as evidenced by HY5-overexpression experiment. This mechanistic divergence highlights species-specific evolutionary adaptations in light-hormone crosstalk, where PatHY5 may integrate upstream signals to coordinate metabolic flux redistribution rather than directly activating terminal synthases. Furthermore, we observed that the expression levels of HY5 across different treatment groups did not exhibit significant variation (approximately two-fold change), suggesting that post-translational modifications or protein interaction networks may dominate its regulatory activity in patchouli.
Photodynamic: linking photosynthetic efficiency to patchoulol biosynthesis
-
An intriguing observation in our research is the decrease of auxins under light conditions—contrary to the trend of JA (Supplementary Fig. S1). Light plays a crucial role in influencing both primary and secondary metabolism in plants, predominantly through the process of photosynthesis[34]. Photosynthetic efficiency can be regulated by stomatal size and movement rate[35]. During light exposure, stomatal movement is driven by energy derived from photosynthetically active radiation (PAR), facilitating photosynthetic phosphorylation; while in darkness, oxidative phosphorylation occurring in guard-cell mitochondria fuels stomatal movements[36]. In the natural environment, plants frequently face the challenge of adapting to fluctuating light conditions[37]; their light-harvesting systems function not only as collectors of light energy but also as dissipators of excess energy during these adaptive responses[38,39]. This adaptation involves intricate energy transfer mechanisms between photosystem I (PSI) and photosystem II (PSII)[40], with the balance of light energy between the two systems maintained through state transitions[38]. Optimizing light harvesting can help to improve plant photosynthetic efficiency, as indicated by recent studies[41−43]. In cyanobacteria, glycogen synthesis has been shown to be dependent on energy inputs, peaking as they enter a rapid growth phase, and metabolite overflow contributes to energy balancing[44]. A similar trend is observed in the accumulation of fatty acids in microalgae[45]. Following the law of conservation of energy, when photosynthetic efficiency reaches a relatively stable state, an increase in light energy within the plant's environment correlates with heightened chemical energy production through photosynthesis. This, in turn, facilitates a more abundant carbon supply for metabolic processes, thereby supporting the synthesis of secondary metabolites in plants. This relationship may provide an explanation for the observed correlation between light wavelength and patchoulol biosynthesis in our study (Fig. 1b, c). Another observation is that patchouli plants under light conditions exhibit enhanced oxidation-reduction potential (Fig. 3), which may promote the expression of certain enzyme genes in the MVA pathway. This phenomenon could be attributed to the light-induced modulation of redox homeostasis, as supported by previous studies on the interplay between light signaling and metabolic pathways[46]. As plants harness greater amounts of light energy, the resultant increases in photosynthetic efficiency and carbon assimilation likely create a more favorable environment for the production of secondary metabolites, including patchoulol (Fig. 7). Understanding these dynamics can offer valuable insights into optimizing patchoulol production through targeted manipulation of light conditions, ultimately enhancing agricultural practices and outcomes in patchouli cultivation.
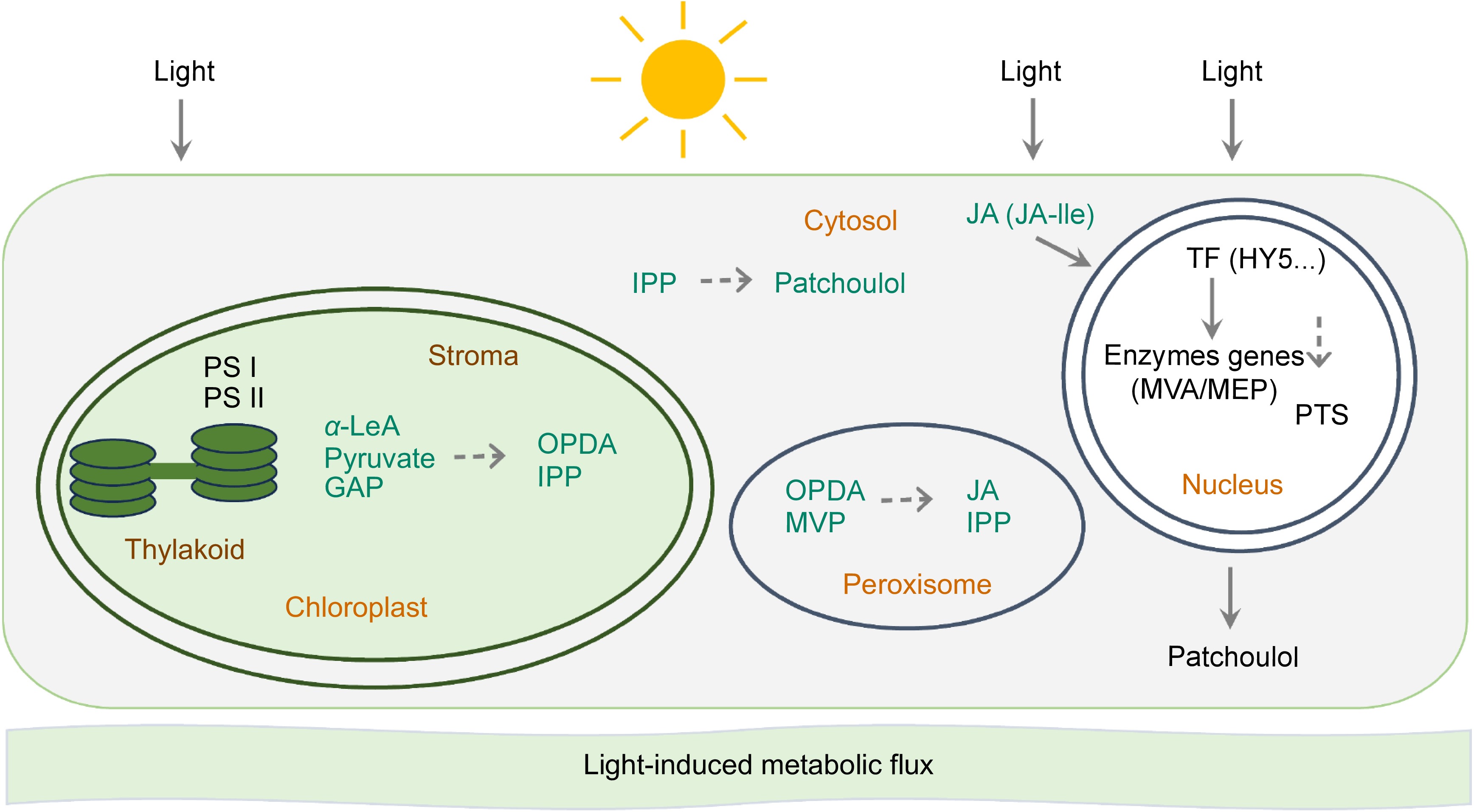
Figure 7.
Jasmonates regulate light-induced patchoulol biosynthesis in P. cablin. Light may initiate a multi-layered regulatory network in P. cablin. Chloroplast-MEP axis: light induces MEP pathway genes, enhancing IPP production; plastid transporters export precursors to cytosol for patchoulol synthesis. Photosynthetic coupling: PSI/II-generated redox signals activate MEP enzymes while diverting carbon flux from competing pathways. JA-mediated regulation: light-stimulated OPDA synthesis in chloroplasts triggers JA-Ile signaling, activating MYC2 transcription factors that upregulate PTS and MEP genes. Photoreceptor cascades: phytochrome/cryptochrome-HY5 modules directly activate terpenoid biosynthetic genes through G-box promoter elements.
-
In conclusion, this study provides preliminary evidence for the pivotal roles of jasmonates in light-induced patchoulol biosynthesis. The data suggest that light enhances patchoulol production through a complex interplay between light signaling and JA pathways. Chemically, within a specific intensity, and at least in the visible light range, light promotes jasmonates biosynthesis and patchoulol biosynthesis, with a strong correlation observed between jasmonates level and patchoulol accumulation. Physiologically, patchoulol biosynthesis seems to be related to the activity of photosynthetic apparatus. Genetically, light influences the expression of genes in patchoulol and jasmonates biosynthesis pathways. Collectively, JA and photosynthesis play a role in light-induced patchoulol biosynthesis, and it may involve metabolic compensation and oxidation-reduction. Nevertheless, further research is needed to elucidate the specific mechanisms by which HY5 or other key genes modulate these pathways and to explore the temporal dynamics between light-induced patchoulol biosynthesis and JA accumulation. This enhanced understanding could potentially revolutionize the optimization of patchoulol production in patchouli, thereby amplifying its industrial utility and economic value.
This work was supported by the National Natural Science Foundation of China (U22A20446, 32322007, 32100192), and the Basic and Applied Basic Research Foundation of Guangdong Province (2022A1515110510, SL2023A04J02655).
-
The authors confirm contribution to the paper as follows: study conception and design, supervision, review and editing: Wang H, Jin H; review and editing: Duan S; funding acquisition: Wang H, Jin H, Duan S, Chang Z; resources: Shen Y; methodology: Wang M, Hong L, Chen Z, Zeng Y, Huang H; data collection: Ou D, Wang Y; analysis and interpretation of results: Wang Y; draft manuscript preparation: Wang Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated in this study are available in the paper or online Supplementary Information.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yanhua Wang, Dafeng Ou
- Supplementary Fig. S1 Impact of light on plant hormones in patchouli. These plant hormones include ABA (abscisic acid), Auxin, CK (cytokinin), ETH (ethyne), GA (gibberellin), JAs (jasmonates), SA (salicylic acid), SL (strigolactone), in patchouli grown in darkness or under four different light conditions. Statistical analysis was performed using Student’s t test. *p < 0.05; ns, no significant.
- Supplementary Fig. S2 Impact of light on jasmonates and their precursors in patchouli. Compounds in the JA signaling pathway in patchouli grown under above experimental conditions. Statistical analysis was performed using Student's t test. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; ns, no significant.
- Supplementary Fig. S3 HY5 sequence homogeneity.
- Supplementary Fig. S4 Regulation of PTSp1 and PTSp2 promoter activity by HY5.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Ou D, Huang H, Zeng Y, Chen Z, et al. 2025. Jasmonates regulate light-induced patchoulol biosynthesis in Pogostemon cablin. Medicinal Plant Biology 4: e015 doi: 10.48130/mpb-0025-0012 

Jasmonates regulate light-induced patchoulol biosynthesis in Pogostemon cablin
- Received: 27 December 2024
- Revised: 10 March 2025
- Accepted: 24 March 2025
- Published online: 23 May 2025
Abstract: Patchoulol, a primary component of patchouli oil which is extensively utilized in the fields of medicine, flavoring, and fragrances. Notably, previous research has indicated that light conditions significantly influence the accumulation of patchoulol in patchouli plants. In this study, we further investigated the hormone levels in patchouli under various light treatments, revealing a strong correlation between jasmonates and patchoulol biosynthesis. Both patchoulol and jasmonate biosynthesis were found to be light-dependent, exhibiting wavelength-specific responses within certain ranges showing similar trends. Additionally, gene expression analysis under light and dark conditions identified several candidate genes within the light and jasmonates signaling pathways. These findings provide a theoretical foundation for understanding the light-induced mechanisms of patchoulol biosynthesis and offer insights for advancing the patchouli industry.
-
Key words:
- Jasmonates /
- Pogostemon cablin /
- Patchouli biosynthesis /
- Light signal /
- Photosynthesis /
- Plant metabolism