










-
Sleep occupies approximately one-third of a person's life and is a vital physiological function that maintains balance in the circadian rhythm and overall health. Adequate sleep is crucial for physical recovery, immune system function, cognitive performance, emotional regulation, cardiovascular health, metabolism, and weight control[1]. Despite its recognized importance, sleep is often neglected. Young people frequently sacrifice sleep due to work pressures and screen exposure, while older adults experience age-related sleep disturbances[2]. Recent studies suggest that sleep not only coexists with various health issues but may also play a causal role in their development, elevating sleep health to a significant public health challenge[3]. Suboptimal sleep quality, manifested by insufficient sleep duration, nocturnal sleep fragmentation, and circadian misalignment, is associated with a spectrum of systemic pathologies. Chronic sleep deprivation (SD) has been linked to elevated cardiovascular disease risk due to heightened sympathetic activity and elevated BP[4]. Sleep fragmentation disrupts glucose metabolism and insulin sensitivity, thereby disrupting the balance of glucose and insulin and elevating susceptibility to diabetes mellitus and obesity[5]. Furthermore, circadian rhythm disturbances prevalent in shift workers demonstrate associations with neurodegenerative disorders such as Alzheimer's and Parkinson's diseases, attributed to heightened oxidative stress and impaired clearance of cerebral metabolic waste[6]. We have compiled a list of health problems that can be caused by different types of sleep disorders (Supplementary Table S1−S4).
In particular, recent research has highlighted that disturbances in the sleep process can significantly affect overall health and are closely linked to changes in ocular physiological function, potentially exacerbating various eye diseases. For example, sleep disorders like Obstructive Sleep Apnea (OSA) have been associated with conditions such as dry eye syndrome, myopia, glaucoma, cataracts, and diabetic retinopathy (DR)[3,7,8]. This connection emphasizes the need to integrate sleep assessments into ophthalmic diagnosis and treatment, and suggests that improving sleep quality could optimize eye health and enhance patients' quality of life.
The primary objective of this review is to systematically explore the relationship between sleep disorders (such as SD and circadian rhythm disturbances), and eye health, focusing on two dimensions: ocular tissues (e.g., cornea, retina, and optic nerve), and ocular diseases (e.g., dry eye, glaucoma, and age-related macular degeneration (AMD)). The review will first examine the physiological basis of sleep, its effects on the brain and body systems, and then discuss various sleep disorders and their potential impacts on eye health. We will also explore lifestyle factors and interventions to improve sleep quality. Finally, the paper will address future research directions aimed at enhancing ophthalmologists' understanding of the importance of sleep in the prevention and treatment of ocular diseases.
-
To ensure a comprehensive and unbiased integrative literature review, we performed an extensive literature search across multiple scientific databases, including PubMed, Scopus, Web of Science, and Google Scholar. This inclusive approach enabled the capture of diverse research perspectives across both clinical and preclinical domains. Articles published in multiple languages were included, and non-English studies were translated using professional services to enhance global representation.
A broad set of keywords and their combinations was used to maximize retrieval relevance, including: 'sleep patterns', 'ocular health', 'sleep disorders', 'eye diseases', 'circadian rhythm's', 'DR', 'glaucoma', 'dry eye syndrome', 'melatonin', and 'retinal homeostasis'. Corresponding Medical Subject Headings (MeSH) were incorporated to further refine the search scope.
The literature search covered studies published from 1976 to 2023, with a cut-off date for inclusion set at February 2025 to ensure coverage of the most recent evidence. Additionally, we manually screened the reference lists of eligible articles to identify further relevant sources.
Inclusion and exclusion criteria
-
Studies were included if they directly investigated the relationship between sleep (or sleep disorders) and ocular health outcomes, whether at the physiological, molecular, or clinical level. To ensure methodological rigor, we excluded articles that lacked a clearly described methodology, presented incomplete or non-extractable data, failed to report outcomes relevant to sleep or ocular health, or were review papers without critical appraisal or original analysis. Unlike prior reviews that excluded non-English literature, we evaluated such studies based on their scientific relevance and quality, utilizing professional translation when necessary to maintain inclusivity.
Literature quality assessment
-
A total of 145 studies that met the inclusion criteria were evaluated and classified into three categories: 44 randomized controlled trials, 74 observational studies (including case-control and cohort designs), and 27 systematic reviews or meta-analyses. Collectively, these studies contributed to a complementary evidence chain, encompassing mechanistic exploration, short-term clinical validation, and long-term integration at the population level.
To assess the methodological quality of the RCTs, we applied the Cochrane Risk of Bias Tool 2.0 (RoB 2). Of the 44 RCTs, 65.9% (29/44) were judged to have a low risk of bias due to the appropriate implementation of randomization procedures, allocation concealment, and blinding protocols. An additional 29.5% (13/44) were rated as having some concerns, largely due to insufficient reporting of randomization methods or incomplete outcome data handling. The remaining 4.5% (2/44) were classified as high risk, primarily because of systematic flaws in allocation concealment or the absence of outcome assessor blinding.
For the 74 observational studies, we used the modified Newcastle-Ottawa Scale (NOS) to evaluate study quality. 56.8% (42/74) were rated as high quality, while 36.5% (27/74) were downgraded to moderate quality due to limitations such as the absence of sample size calculations or reliance on single-center datasets. A small proportion, 6.8% (5/74), were classified as low quality, typically due to small sample sizes (n < 20) or poorly defined inclusion criteria.
The quality of the 27 systematic reviews and meta-analyses was assessed using the AMSTAR-2 tool. Among these, 48.1% (13/27) met at least 12 quality indicators and were rated as high quality, with notable strengths in protocol pre-registration, comprehensive search strategies, and standardized risk of bias assessments. 40.7% (11/27) were considered moderate quality, often due to the omission of grey literature searches or inadequate heterogeneity analysis. The remaining 11.1% (3/27) were judged to be low quality, having failed to meet two or more critical criteria, such as conducting publication bias assessments.
Synthesis strategy and limitations
-
Although this review synthesized findings across a wide range of studies, we did not conduct a formal meta-analysis due to substantial heterogeneity in study designs, outcome measures, statistical reporting, and analytical methods. In particular, inconsistent reporting of effect sizes and the absence of standardized quantitative indicators precluded meaningful power calculations or effect pooling.
Instead, we adopted a systematic narrative synthesis approach to identify emerging patterns, mechanistic pathways, and translational implications. This method allows for a broader integration of both quantitative and qualitative data across diverse evidence types. Future research should aim to conduct meta-analyses using harmonized outcome measures, effect size standardization, and bias-adjusted models to quantitatively assess the association between sleep and ocular health.
-
Sleep is a complex physiological state involving multiple systems within the brain and body, encompassing two primary types: rapid eye movement (REM) sleep and non-rapid eye movement (NREM) sleep. NREM sleep is further subdivided into three stages: N1 (light sleep), N2 (moderate sleep), and N3 (deep or slow-wave sleep), with each complete sleep cycle lasting approximately 90 min[9]. During this cycle, individuals transition from wakefulness to N1, followed by deeper N2 and N3 stages. The N3 stage plays a critical role in body recovery processes, including tissue repair, energy restoration, and the release of essential hormones[10]. The REM stage is marked by increased brain activity, which is vital for memory consolidation and learning, while muscle atonia, except for the eye and respiratory muscles, is observed[11].
Sleep is regulated by several brain regions, such as the brainstem, hypothalamus, and forebrain, and is influenced by neurotransmitters like adrenaline, serotonin, and γ-aminobutyric acid[12]. It significantly impacts the endocrine system by regulating cortisol (the stress hormone) levels and promoting growth hormone (GH) release[13]. Additionally, sleep boosts immune function and regulates inflammatory responses[14]. During sleep, heart rate and blood pressure (BP) decrease, while significant changes in respiratory patterns occur, particularly during REM sleep. Body temperature also slightly decreases, helping maintain the sleep state[10]. Thus, the normal sleep process is essential for overall health.
Historically, sleep has been considered primarily in terms of sleep disorders, often viewed as a symptom of disease rather than a positive health function. For example, earlier research typically focused on sleep duration and insomnia symptoms, such as difficulty falling asleep, fragmented sleep, and early awakening, with OSA being the most extensively studied disorder[15]. Today, the concept of 'sleep health' encompasses the need for sufficient, high-quality sleep at appropriate times to meet the body's and brain's needs for recovery and repair. Key components of sleep health include: (1) Sleep Duration: Adults should aim for 7–9 h of sleep per night, while adolescents typically require more. (2) Sleep Quality: High-quality sleep is characterized by deep, continuous sleep with minimal interruptions, particularly during the NREM and REM stages. (3) Sleep Cycle: A healthy sleep cycle includes transitions between REM and NREM sleep, with NREM sleep progressively deepening to allow for optimal recovery. (4) Sleep Continuity: Sleep continuity refers to uninterrupted sleep throughout the night. Frequent awakenings can reduce sleep quality and hinder the body's recovery process. (5) Sleep Rhythm: Maintaining a regular sleep/wake cycle is crucial for aligning with the body's internal biological clock, which promotes better sleep quality. (6) Sleep Environment: A comfortable, quiet, dark, and cool environment enhances sleep quality by reducing disturbances from noise or uncomfortable temperatures. (7) Pre-Sleep Habits: Engaging in relaxing activities and avoiding stimulants before bedtime can help prepare the body for restful sleep.
The emerging concept of 'sleep health' provides a more holistic view, considering sleep not just as the absence of disorder but as an essential factor for overall well-being. The multidimensional nature of sleep health allows it to be measured as a continuous variable among individuals. The importance of sleep health is illustrated in (Fig. 1).
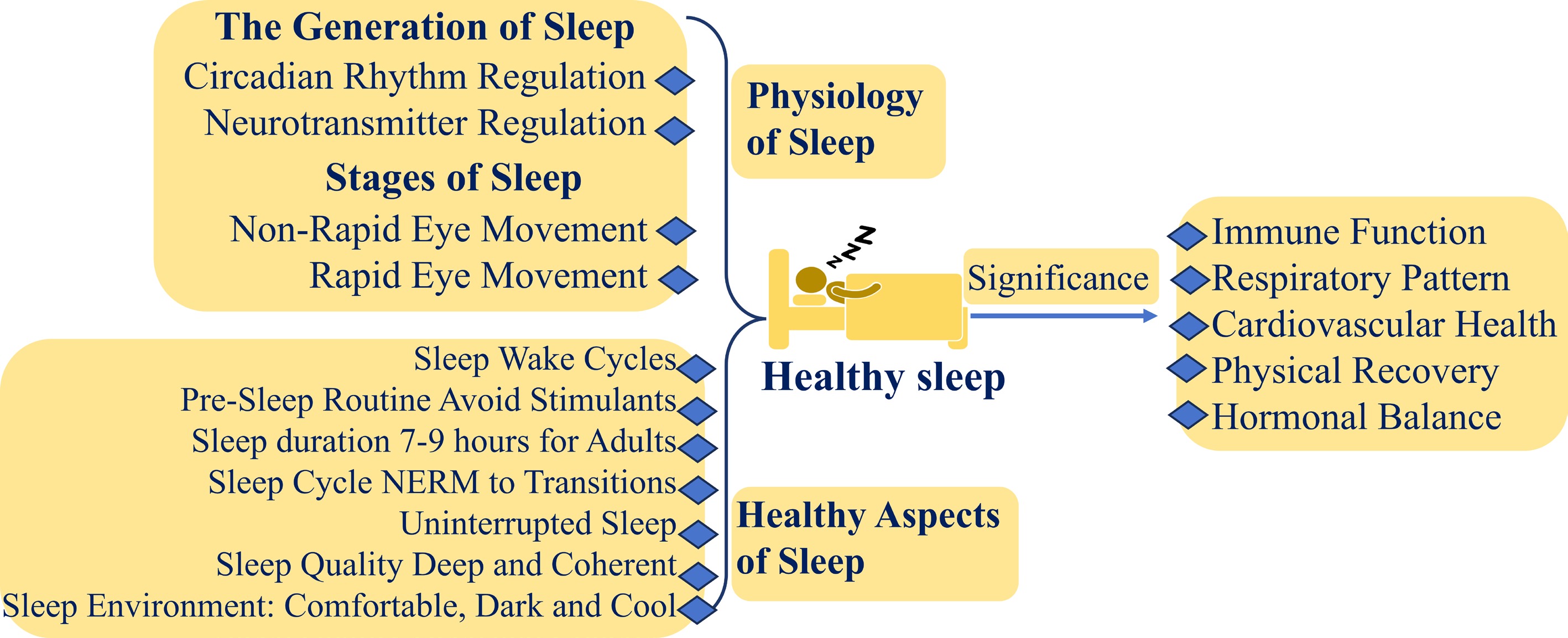
Figure 1.
The physiology of healthy sleep and its importance (created by the authors).
Current sleep recommendations suggest that adults should sleep at least 7 h per night. According to the Centers for Disease Control and Prevention, approximately 35% of adults in the US are sleep-deprived[16]. Insomnia symptoms, including difficulty falling asleep, disrupted sleep, and early waking, are common, with approximately 30% to 35% of adults experiencing them and around 10% meeting the diagnostic criteria for insomnia disorder[17]. Estimates indicate that the prevalence of OSA ranges from 3% to 10%, though this may be underreported, especially in overweight and elderly populations[18]. These statistics highlight the complexity of sleep issues and emphasize the significant public health challenges posed by poor sleep characteristics and sleep disorders (Fig. 2).
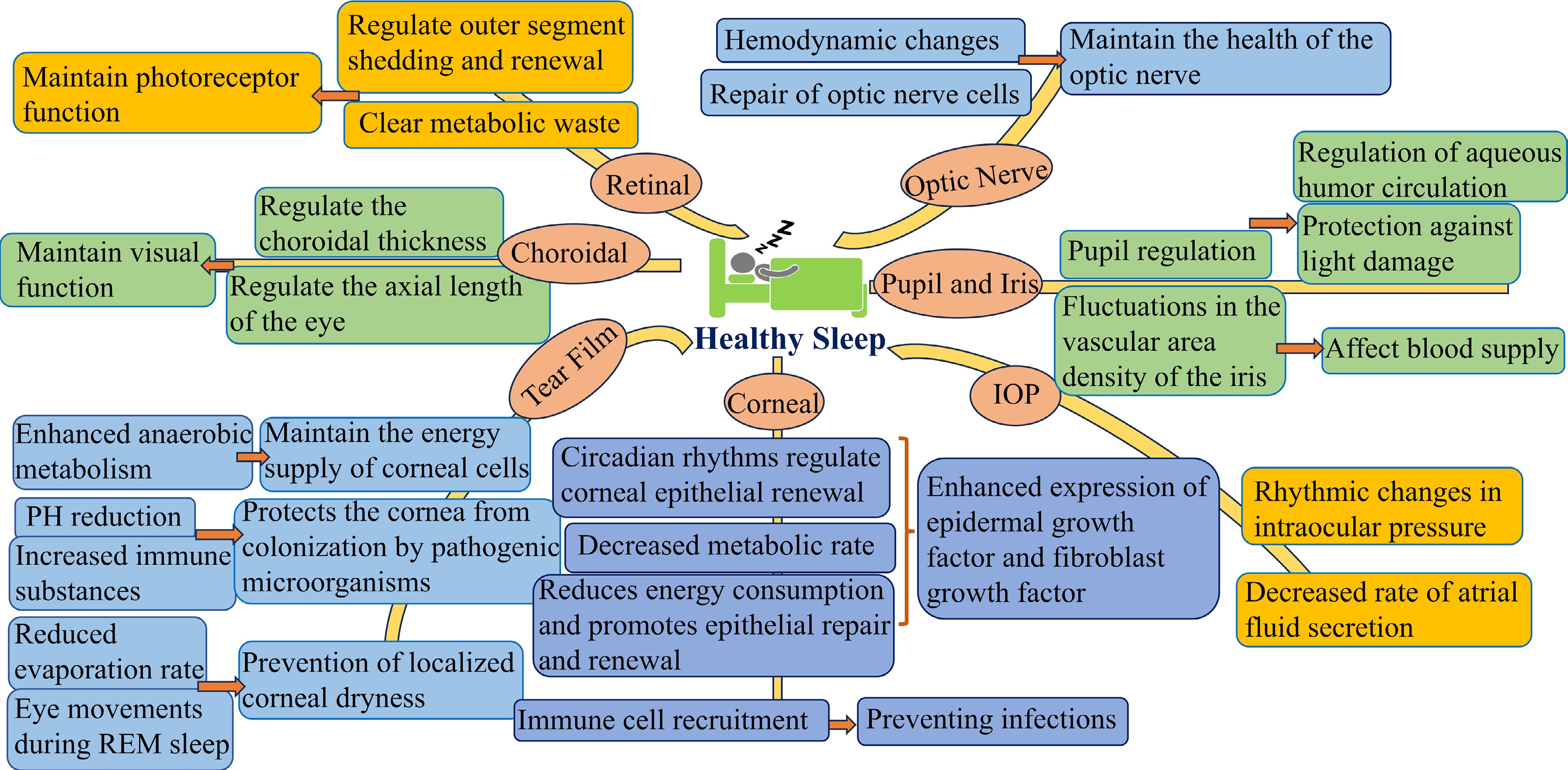
Figure 2.
The relationship between sleep and eye health (created by the authors).
-
Various biological and metabolic processes in the cornea are regulated by circadian rhythms. High-throughput RNA sequencing in mice has revealed that approximately 24% of corneal transcripts exhibit rhythmic oscillations[19,20]. The renewal of the corneal epithelium follows a circadian rhythm, which varies across species. In rats, for instance, the mitotic rate of corneal epithelial cells peaks at night and drops during the day[21]. Conversely, in mice, this mitotic rate is highest during the day and lowest at night[22]. Moreover, hyperglycemia and dietary changes, such as high-fructose or high-fat diets, can significantly disturb these circadian rhythms[20,23,24].
Interestingly, the growth and shedding of nerve fibers in the corneal epithelium also follow a circadian rhythm. In mice, corneal epithelial nerve axons elongate during the day and begin shedding post-light exposure, reflecting changes in corneal sensitivity during sleep that align with the circadian rhythm-driven nerve fiber shedding[25].
Neutrophils are crucial first-line defense cells that respond quickly to infection or injury. Studies in nocturnal animals, such as mice, have shown that the recruitment of neutrophils to the corneal limbus peaks at midnight[20,26]. Similarly, the recruitment of γδ T cells, which play an important role in immune surveillance and inflammatory responses in barrier tissues, also exhibit a circadian rhythm, with higher activity during the night[27]. This recruitment may be linked to lower cortisol levels during the night, as cortisol typically exerts anti-inflammatory effects. The nocturnal activation of neutrophils may be an adaptive mechanism to protect the cornea from external stimuli and injury during the active period, providing additional immune defense during these hours. Circadian recruitment of neutrophils and γδ T cells is influenced by various factors, including circadian clock gene expression, ambient light, hormone levels, and local cytokine and chemotactic factors[28].
Notably, SD has been shown to increase neutrophil activity and recruitment, both in the blood and the corneal limbus. This may be due to the stress response induced by SD, leading to elevated glucocorticoid and epinephrine levels that stimulate the release of more neutrophils from the bone marrow[29]. Similarly, γδ T cells show increased recruitment after SD, and these cells produce higher levels of cytokines like IL-17, leaving the cornea in a sub-inflammatory state. In sleep-deprived mice, corneal injury results in an exaggerated inflammatory response and delayed repair[26].
In summary, the circadian rhythm of the cornea results from the combined effects of circadian clock genes, local metabolism, and systemic regulation, influencing its physiological functions and disease progression.
The role of circadian rhythms in the iris and pupil
-
The human iris is an important and highly vascularized diaphragm that anatomically separates the anterior and posterior chambers of the eyeball. A study using optical coherence tomographic angiography to measure iris vessels in 15 healthy adults revealed circadian fluctuations in vascular area density and vascular skeleton density, with peaks at 18:00 and a nadir at 00:00. Notably, changes in iris vessel area density/vessel skeleton density were positively correlated with BP variations but negatively correlated with intraocular pressure (IOP) fluctuations[30]. However, the OCTA technology used in this study captured only superficial iris vasculature and failed to visualize deep iris vasculature (due to limited signal penetration).
Pupil size changes are regulated not only by external light exposure but also by intrinsic rhythms. An infrared-sensitive camera study recorded rabbit pupil diameter changes under constant darkness, showing smaller diameters during the subjective light phase and larger during the dark phase. The minimum pupil diameter occurred about 5 h after the subjective light phase, at 8.2 mm, and then gradually increased to a peak of 10.8 mm at the onset of the subjective dark phase[31]. This rhythmic oscillation of the pupil, which is independent of the external light environment, further validates the role of the autonomous biological clock in the central regulation of pupil movement.
Circadian rhythms on the iris and pupil may be related to circadian variations in IOP; however, current research is very limited, and further exploration is needed in the future.
The role of circadian rhythms in intraocular pressure
-
IOP is essential for maintaining the eye's shape and optical function. Studies have shown that IOP exhibits regular circadian rhythmic variations, which are synchronized with the body's internal rhythms. This synchronization is achieved, in particular, through the circadian clock in the suprachiasmatic nucleus[32]. In humans, IOP increases at night regardless of body posture[33]. Similarly, nocturnal or diurnal animals also exhibit elevated IOP during nighttime[34]. This nocturnal rise in IOP may be associated with circadian variations in aqueous humor dynamics and altered autonomic nervous system activity during sleep.
Circadian regulation of aqueous humor flow
-
The secretion and outflow of aqueous humor are vital physiological processes that regulate IOP. During sleep, the rate of aqueous humor secretion decreases by 50%–60%[35], resulting in a higher IOP at night compared to the daytime, highlighting the influence of circadian signals on aqueous inflow and outflow. The ciliary body plays a primary role in producing aqueous humor through active secretion, with minor contributions from diffusion and ultrafiltration. In addition to its secretion function, the ciliary body's endocrine activities, and secretion of regulatory peptides significantly influence aqueous humor dynamics and the circadian rhythm of IOP[36].
Molecular mechanisms regulating circadian fluctuations in intraocular pressure
-
The IOP rhythm transmission pathway involves two main components: adrenal glucocorticosteroids and the sympathetic nervous system[37]. Animal studies have shown that dexamethasone administration at different circadian times can shift the biological clock of the iris-ciliary body. Administration between CT8 and CT12 causes a phase delay, while treatment between CT16 and CT20 induces a phase advance, indicating that glucocorticoids (GCs) can adjust the phase of the iris-ciliary body biological clock[37]. Furthermore, adrenalectomy or superior cervical ganglionectomy in mice disrupts IOP rhythms and the biological clock, particularly reducing nocturnal IOP elevation. Reinfusion of cortisol and norepinephrine (NE) restores these disrupted rhythms[37].
In addition, β2-adrenergic receptors, glucocorticoid receptors, and clock proteins are highly expressed in the non-pigmented epithelium of the ciliary body[38]. The interaction between GCs and sympathetic pathways is critical for regulating circadian IOP rhythms. Studies have shown that the incomplete amplitude of IOP rhythms following intraocular injections of NE or GCs in mice lacking adrenal glands or superior cervical ganglia highlights the importance of glucocorticoid-sympathetic interactions in IOP regulation[37]. Collectively, circadian IOP rhythms are regulated by signals originating from the suprachiasmatic nucleus, with temporal signals transmitted through GCs and NE from the adrenal glands and sympathetic nervous system.
The role of circadian rhythms in the lens
-
Typically, ocular tissues contain a robust antioxidant defense system to maintain redox balance and minimize oxidative stress damage. The lens, in particular, contains remarkably high levels of the antioxidant glutathione (GSH). Current research suggests that the redox state of the lens is regulated by circadian rhythms[39]. As an avascular tissue, the lens relies on the aqueous humor for its GSH supply and other amino acids necessary for synthesis. Importantly, circadian rhythms regulate aqueous humor secretion by the ciliary body[35], suggesting that the lens's GSH uptake and synthesis may be influenced by the biological clock. Studies have shown diurnal variations in GSH levels in tissues, including the cerebral cortex[40], and recent research has identified the clock gene Bmal1 in lens nuclear cells. This gene is closely associated with Reactive Oxygen Species (ROS) homeostasis and antioxidant responses, linking it to circadian rhythms[41].
In conclusion, these findings highlight the intricate interplay between circadian rhythms and the redox homeostasis of the lens, which plays a crucial role in maintaining the transparency of the lens.
The role of circadian rhythms in the retina
Circadian regulation of photoreceptor outer segment shedding
-
The retinal circadian system represents a unique structure. It contains a complete circadian system. Circadian rhythm changes in the retina are essential for maintaining normal visual function. First, the shedding of photoreceptor outer segments in the retina is regulated by circadian rhythms. In mammals, retinal pigment epithelial (RPE) cells rhythmically phagocytose distal shed disc membranes at the apical microvilli daily[42]. This process is coordinately regulated by circadian clock genes (e.g., Bmal1, Per1) and local signals (e.g., melatonin, dopamine)[43], with phagocytic peaks concentrated within 2 h after light onset[44]. In contrast, rod outer segment shedding in frog retinas rapidly and fully adapts to phase-shifted light cycles, exhibiting only random, arrhythmic shedding under continuous darkness. Thus, frogs demonstrate stronger environmental dependency[45]. In goldfish, photoreceptor outer segment shedding shows no circadian or endogenous regulation and appears entirely dependent on ambient light changes[46].
Circadian regulation of melatonin and dopamine
-
In addition, the secretion of melatonin and dopamine in the retina is similarly tightly regulated by the biological clock. Specifically, dopamine promotes light-adapted physiology during the day, while melatonin supports dark-adapted physiology at night[47]. Melatonin-secreting photoreceptors and dopamine-secreting cells in the inner plexiform layer form a cellular feedback loop that regulates the circadian rhythm of retinal physiology. Melatonin has potent anti-apoptotic and antioxidant effects[48], and elevated melatonin levels at night not only protect RPE cells from oxidative stress and ischemia-induced cell death but also delay photoreceptor degeneration in mutant mice[49].
Retinal circadian rhythms are associated with various ocular diseases, and their dysfunction may contribute to photoreceptor degeneration, myopia progression, and other pathologies. Future studies should clarify the identity of true clock cells within the retina, their mechanisms of rhythmic coordination, and the impact of circadian rhythms on specific retinal circuits and functional modules. These investigations will provide critical insights into retinal physiology and advance therapeutic strategies for related disorders.
Sleep in tear film and lacrimal gland health
The role of sleep in tear film dynamics
-
The dynamics of the tear film during sleep were historically poorly understood, with initial studies on animals and anesthetized humans showing a significant decrease in tear secretion with closed eyes. These early observations highlighted the stark differences in tear dynamics between open and closed eyes[50]. More recent research, however, has used micro-capillaries to collect small tear samples immediately after prolonged eye closure or overnight sleep. These studies have found that glucose levels in the tear film remain stable after overnight closure[51]. However, significant reductions in pH[52] and osmolarity[53], along with shifts in the malate dehydrogenase to lactate dehydrogenase ratio and lactate dehydrogenase isoenzyme profiles, suggest an increase in anaerobic metabolism during sleep[54,55]. Furthermore, certain tear biomarkers, such as epidermal growth factor (EGF) and vascular endothelial growth factor (VEGF), follow circadian rhythms and peak in the evening[56]. In contrast, significant diurnal variations in cytokines like interleukin-10 (IL-10), Interferon-Gamma (IFN-γ), and tumor necrosis factor-alpha (TNF-α) have been observed, peaking in the morning and late evening[57]. Albumin concentrations in tears peak within 10 min of waking and drop to their lowest levels between 2-6 h later, gradually increasing through the latter part of the day[58].
Two hours after awakening, tear film evaporation rates increase rapidly[59], while during sleep, the closed-eye state reduces tear evaporation despite diminished lacrimal gland secretion, thereby maintaining ocular surface moisture. Notably, in the REM phase, eye movements further redistribute tears to alleviate ocular surface dryness[60]. However, certain sleeping positions, such as lying face-down, can partially open the eyelids, increasing tear evaporation and impacting ocular moisture[61]. Side-sleeping, influenced by pillow or hand pressure, may hinder eyelid closure and increase tear evaporation. While changing sleep positions can help alleviate dry eye symptoms[62], it does not address the root cause of chronic dry eye.
The role of sleep in tear film immunity
-
Recent studies have found that the composition, origin, and renewal of the tear film undergo significant changes during sleep or eye closure. For instance, studies analyzing tear samples from dogs showed substantial diurnal fluctuations in immunoglobulin concentrations[63], while human tear fluid studies revealed that lysozyme and Immunoglobulin A concentrations peak between 9 a.m. and 12 p.m. but decrease during sleep, particularly between midnight and 3 a.m.[54,64]. Immunoblot analysis revealed that eye closure activates the complement system, specifically Complement C3, while reflex tear secretion substantially decreases or ceases. Secretory Immunoglobulin A concentrations, however, increase during this period. Neutrophils also accumulate in the conjunctival sac during sleep or eye closure[65,66], where they become activated, along with an increase in Th17 lymphocytes[65]. This rise in leukocytes is accompanied by an increase in neutrophil-derived cytosolic proteases, which likely play a critical role in protecting the eye from pathogens during sleep. Therefore, during the eye-closed state, the ocular surface defense shifts from a passive mode to an active immune and sub-inflammatory state, protecting against trapped microorganisms[67].
The role of sleep in lacrimal gland structure and function
-
Recent studies have revealed distinct circadian rhythms in the secretory activity of the lacrimal glands, closely aligned with the body's biological clock[68,69]. Factors such as jet lag, shift work, and new parenthood can lead to SD or poor sleep quality, negatively affecting lacrimal gland function. These disruptions primarily result in shifted circadian rhythms and reduced tear secretion[70,71]. Such effects likely arise from sleep disorders that disturb hormonal balances, particularly hormones that regulate tear secretion. Additionally, sleep disorders may dysregulate the autonomic nervous system, which is essential for regulating lacrimal gland function[72,73].
Circadian rhythms also influence the recruitment of specific immune cell populations within the lacrimal gland. Neutrophils, T lymphocytes (including γδ T cells), and natural killer cells have all been found to exhibit circadian recruitment[68]. Conditions like hyperglycemia[68], nutritional stress[69], SD[71], and altered photoperiods[68] can significantly alter the circadian recruitment patterns of these immune cells. Furthermore, age-related changes impact both the circadian transcriptome and the recruitment patterns of immune cells in the lacrimal gland[74–75].
Sleep deprivation and lacrimal gland dysfunction
-
The negative effects of poor sleep on lacrimal gland function and the biochemical composition of the tear film can primarily be attributed to several factors. First, SD disrupts the circadian rhythm of lacrimal glands, leading to reduced tear secretion, increased epithelial cell defects, and lipid accumulation—symptoms that mimic dry eye disease (DED)[71,76]. Specifically, SD alters the circadian transcriptome of lacrimal glands, resulting in irreversible changes in immune cell trafficking and secretory functions. Additionally, OSA has been shown to disrupt signaling pathways within the lacrimal gland, exacerbating myoepithelial cell damage and inflammatory responses. OSA interferes with key metabolic signaling pathways in the lacrimal glands, leading to dry eye by reducing tear secretion and increasing cell apoptosis.
Moreover, SD induces pathological changes that are partially reversible with rest, suggesting that the lacrimal system can temporarily adapt to acute sleep loss[70]. Evidence shows that N-Palmitoylethanolamine treatment can alleviate SD-induced lacrimal dysfunction by restoring lipid metabolism and cellular morphology in the lacrimal gland[77]. Additionally, overnight sleep alters the composition of the tear film, influencing various biochemical markers and cell types involved in tear film homeostasis[65].
In conclusion, improving sleep quality, particularly by increasing deep sleep, supports lacrimal gland function and alleviates dry eye symptoms. Therefore, maintaining good sleep habits is essential for the health of the tear film and lacrimal glands, and improving sleep quality can effectively relieve the symptoms of dry eye syndrome.
Sleep in corneal health
The role of sleep in corneal repair and regeneration
-
Sleep plays a crucial role in maintaining corneal health, as the transparent cornea accounts for approximately 70% of the eye's focusing capacity, which is essential for visual clarity. During sleep, corneal cells undergo vital repair and regeneration, helping to mitigate minor damage caused by factors such as prolonged screen use, dust exposure, and dry environments. The eyelids, when closed during sleep, evenly distribute tears over the corneal surface, providing essential lubrication and nutrients. This process helps alleviate dry eye symptoms and maintain corneal health, ensuring that the cornea remains hydrated and capable of optimal function. In summary, sleep maintains the structural and functional diurnal-nocturnal balance of the cornea through dynamic repair mechanisms, serving as a critical regulatory window for corneal health.
The role of sleep in corneal metabolic and neural function
-
The cornea, being an avascular tissue, derives its oxygen mainly from the atmosphere. However, during sleep, eyelid closure reduces the oxygen concentration on the corneal surface to about one-third of its open-eye level[78]. Despite this limitation, the cornea still receives sufficient oxygen through conjunctival blood vessels and aqueous humor, maintaining adequate levels for cellular metabolism[79].
Sleep also aids in clearing metabolic wastes from the cornea, helping preserve its transparency and functionality. SD increases systemic inflammation, which affects ocular tissues, including the cornea[80]. Adequate sleep helps reduce these inflammatory responses, protecting the cornea. Sleep disorders, such as sleep apnea, can worsen corneal health by disrupting oxygen supply and increasing inflammation[81].
Contact lens wearers, in particular, experience significant metabolic changes during sleep due to hypoxia and hypercapnia[82]. These changes affect the epithelial, stromal, and endothelial layers of the cornea, leading to observable effects post-sleep. In the epithelial layer, metabolic changes result in decreased rates of metabolism, morphological alterations, microcyst formation, and reduced sensitivity. In the stromal layer, changes include edema, acidosis, neovascularization, and potential thinning of the cornea. In the endothelial layer, sleep-related changes can lead to vacuole formation, altered cell density, and potential functional impairments[70].
These findings suggest that sleep modulates the metabolic microenvironment and inflammatory homeostasis, thereby directly influencing the structural integrity and functional stability of the cornea's multilayered architecture.
The role of sleep in corneal molecular regulation
-
Sleep regulates corneal health by controlling the metabolic activity of corneal cells, facilitating repair and renewal, and modulating intercellular signaling. During sleep, the overall metabolic rate decreases, reducing energy consumption by corneal cells and promoting cell repair and regeneration. Corneal cells use this period for key biological processes such as DNA repair, protein folding, and lipid renewal. The hypoxic conditions during sleep help reduce oxidative stress, optimizing the regenerative environment for corneal cells and supporting corneal transparency[83]. Furthermore, sleep influences the expression of cytokines and growth factors that regulate intercellular communication. For example, sleep enhances the expression of EGF and fibroblast growth factor (FGF), both of which are essential for the migration and proliferation of corneal epithelial cells[84]. Additionally, sleep helps regulate inflammatory mediators, reducing cellular damage by minimizing inflammatory responses[83]. Collectively, these mechanisms indicate that sleep integrates metabolism, redox homeostasis, and signaling through multidimensional molecular networks, providing precise spatiotemporal regulation for corneal repair.
Sleep in iris and pupil health
The role of sleep in iris blood supply and nutrient transport
-
The iris, located between the cornea and the lens, plays a crucial role in regulating the amount of light that reaches the retina. It contains abundant pigment, blood vessels, and smooth muscle, with melanin serving to protect the retina from intense light exposure. Early electron microscopy studies have identified four types of close contacts between melanocytes and nerve endings in the human iris stroma, corresponding to adrenergic and cholinergic trophic innervation. This discovery not only reveals the dynamic regulatory mechanisms of iris nutrient supply during sleep but also suggests that fluctuations in autonomic nervous function during sleep cycles may influence iris physiology through neuro-melanocyte interactions[85]. In addition, circadian rhythms affect the vascular area density and vascular skeleton density of the iris. This fluctuation further affects the efficiency of blood supply and nutrient exchange in the iris, providing varying degrees of support for normal iris function at different times of the day[86].
The role of sleep in pupil size and reactivity
-
Changes in pupil size during sleep relate to sleep stages; in cats, the pupil significantly narrows during NREM sleep and dilates periodically after REM sleep begins[87]. Unlike mammals, pigeons show pupil dilation during NREM sleep and brief, rapid bursts of constriction and dilation timed with the REM phase[88]. This difference between mammals and birds may stem from different muscular controls of the iris. Recent studies indicate that sleep disorders impact pupil dynamics during sleep; video-based eye-tracking detected altered pupil responses in patients with Parkinson's disease and REM sleep behavior disorder, showing reduced constriction, dilation, and fewer blinks compared to a healthy control group[89].
Mechanisms of sleep-driven pupil regulation
-
Research conducted on head-fixed mice during NREM sleep demonstrated significant pupil constriction, which later dilated during REM sleep but remained smaller than during wakefulness. This pupil constriction during sleep is thought to protect the retina by minimizing light exposure and sensory input, thus facilitating deeper sleep. The phenomenon is mediated by the parasympathetic nervous system and likely acts as a shield against external disturbances during sleep[90]. In mammals, pupil size reflects autonomic nervous system activity; increased sympathetic activity causes pupil dilation during alert wakefulness, while increased parasympathetic activity leads to pupil constriction during sleep or sedation[91–92].
Melatonin also plays a role in regulating the pupil's circadian response to light[93]. Sleep influences hormone levels, particularly melatonin secretion, which is primarily regulated by circadian rhythms. Sleep homeostasis and circadian rhythms are interconnected and jointly regulate various physiological variables[94]. Among the core circadian clock genes, only the Cry1 gene follows the circadian rhythm, remaining largely unaffected by the sleep-wake cycle[95]. The circadian rhythm of melatonin secretion is closely related to normal sleep patterns in humans[96]. Research has shown that night shift workers, including nurses, exhibit significantly lower melatonin levels compared to day shift workers[97]. Long-term chronic SD can disrupt the biological clock and impair the pineal gland's ability to secrete melatonin. Moreover, the widespread use of indoor lighting and electronic screens before bedtime in modern life may further disrupt melatonin secretion, delaying its release and reducing its duration[97].
In summary, sleep dynamically regulates iris blood supply and pupillary motor function through complex neuro-endocrine mechanisms. The balance of the autonomic nervous system and the rhythmic secretion of melatonin form the core components of this regulatory process. Sleep disorders may disrupt these physiological mechanisms, thereby increasing the risk of ocular diseases. Therefore, maintaining good sleep habits is essential for preserving the normal physiological functions of the iris and pupil.
Effects of sleep stages on intraocular pressure
-
IOP fluctuations during sleep are closely related to sleep stages. A contact lens sensor (CLS) continuously measures IOP by detecting corneal curvature changes without requiring the subject to be awake[98]. A study using CLS to monitor 24-h IOP in 12 healthy volunteers found that nocturnal IOP was significantly lower during wakefulness compared to N1, N2, and N3 stages of NREM sleep and REM sleep. IOP was highest during REM sleep and progressively decreased with the deepening of NREM sleep[99]. However, before this, some studies measured the relationship between IOP and the sleep cycle using tonometry and found that the IOP during REM sleep was lower than that during slow-wave sleep. The discrepancies in research findings are associated with the methods used for IOP measurement. Continuous monitoring with CLS provides real-time data on IOP fluctuations, whereas tonometer-based measurements require periodic awakening of subjects, which may disrupt natural sleep cycles and thereby affect IOP readings[100]. Additionally, differences in sleep posture can influence nocturnal IOP changes, introducing confounding variables[101], which are often inadequately controlled in clinical studies. Future research should employ standardized, non-invasive continuous IOP monitoring methods to validate these findings across distinct sleep stages. Notably, sleep-related disorders such as severe OSA can prolong nocturnal IOP elevation, with the severity of IOP changes correlating with the disease progression[102].
Recent research has revealed the complex relationship between sleep and IOP regulation, highlighting circadian rhythmic variations in IOP that are synchronized with body rhythms. Nocturnal fluctuations in IOP are closely associated with sleep stages, with distinct patterns observed during different sleep phases, and disruptions in these patterns have been linked to sleep-related disorders. Physiological processes governing aqueous humor dynamics, which are modulated by intrinsic ocular circadian rhythms, contribute to elevated IOP during the night. Molecular mechanisms involving adrenal glucocorticosteroids and sympathetic nervous system signaling play key roles in regulating circadian IOP rhythms, with important clinical implications for glaucoma risk. SD disrupts these rhythms, potentially increasing nocturnal IOP and posing risks to ocular health. Understanding the interplay between sleep and IOP regulation is essential for developing comprehensive management strategies in clinical practice.
Sleep in retinal health
The impact of sleep on metabolic waste clearance in the retina
-
Sleep plays a crucial role in the removal of metabolic waste from the brain and retina. This process is facilitated by the lymphatic fluid system, which uses cerebrospinal fluid and interstitial fluid to clear toxic metabolites accumulated in the brain. Studies have shown that the clearance of potentially toxic proteins occurs twice as fast during sleep compared to wakefulness[103−105]. Recently, research has emphasized the importance of sleep in clearing metabolic waste from the retina. During REM sleep, a 'vitreous pump' helps remove metabolic waste products from the retina, functioning similarly to the lymphatic system in the brain[106]. This process is essential for maintaining retinal health and may have significant implications for various retinal diseases. Additionally, studies measuring choroidal blood flow with laser scatter flowgraphy have shown that blood flow to the optic nerve head peaks at midnight, suggesting that the optic nerve undergoes efficient metabolism and repair during the night[107]. Therefore, adequate sleep contributes to retinal function indirectly by improving systemic health factors, such as vascular regulation, which ensures sufficient blood flow and oxygenation to retinal tissues.
Immune status of the retina
-
The retina possesses a complex immune system that is essential for maintaining ocular health and responding to infection, inflammation, and injury. This system includes macroglial cells (e.g., Müller cells and astrocytes), microglial cells, and resident immune cells like lymphocytes and monocytes. Macroglial cells protect retinal neurons, while microglia serve as the primary immune cells in the central nervous system, clearing debris after neuronal injury and regulating inflammation[108].
Recent studies using adaptive optics have provided label-free imaging of dynamic changes in retinal immune cells, revealing that their activity is influenced by circadian rhythms and sleep patterns[109–110]. Microglia, for example, undergo rhythmic changes in morphology and function, with distinct activity patterns during sleep and wakefulness[111]. Chronic SD disrupts the circadian regulation of microglial activity, leading to increased oxidative stress, elevated inflammatory cytokine production, and impaired clearance of cellular debris, which contributes to retinal dysfunction and neurodegeneration[112]. These findings underscore the importance of sleep as a key strategy for preventing retinal dysfunction and protecting vision.
Sleep in optic nerve health
Hemodynamic adjustments during sleep
-
Sleep exerts significant effects on optic nerve health through various physiological mechanisms, including hemodynamic adjustments, changes in metabolic demands, and facilitation of cellular repair and neuroprotection. One of the primary changes during sleep is a reduction in systemic hemodynamics, with heart rate and BP typically decreasing throughout the night[10]. These changes influence the blood supply to the optic nerve. While blood flow may decrease during sleep, in healthy individuals, this reduction is usually not sufficient to cause tissue damage. This decrease in blood flow could help lower the metabolic demands of the optic nerve and reduce oxidative stress, potentially protecting optic nerve cells from free radical damage[113].
However, sleep-related changes in blood flow can have a more detrimental effect in certain conditions. For example, OSA has been shown to significantly reduce blood flow and oxygen supply to the optic nerve, which may compromise optic nerve health[114−116]. In populations with conditions like non-arteritic anterior ischemic optic neuropathy, nocturnal hypotension may exacerbate optic nerve damage due to insufficient perfusion[117]. While reduced BP and heart rate may offer protective benefits in some cases, these effects are not universal and may not apply to individuals at higher risk for ischemic optic neuropathy.
Cellular recovery and repair
-
In addition to hemodynamic changes, sleep plays a crucial role in cellular recovery and repair of the optic nerve. During deep sleep, the body activates processes of cellular regeneration and repair, which are essential for addressing cellular damage accumulated during the day. Nerve growth factors, such as brain-derived neurotrophic factor are released during sleep and play a vital role in the survival and regeneration of optic nerve cells[118]. The expression of these growth factors is upregulated during sleep[119], which helps maintain the health and function of optic nerve cells, contributing to their repair and regeneration. This process is vital for ensuring the long-term health of the optic nerve, particularly in individuals exposed to daily oxidative stress and other damaging factors.
Sleep in lens health
Sleep and metabolic waste clearance
-
Despite its crucial role in light transmission and refraction, the lens is vulnerable to oxidative damage from external sources, such as ultraviolet (UV) radiation, air pollutants, and environmental toxins, as well as internal sources like cellular metabolism and inflammation, all of which contribute to oxidative stress[120]. During sleep, the body enters a critical phase of self-repair and metabolic waste clearance, which is essential for maintaining lens transparency. This phase effectively removes metabolic byproducts from the lens due to UV exposure and oxidative stress. The protective mechanisms that the lens employs during sleep are summarized in Table 1.
Table 1. Protective mechanisms of the lens during sleep.
Activity Function Impact UV protection Protects against UV ray damage Minimizes oxidative stress on the lens Antioxidant increase Boosts levels of antioxidants like GSH Protects lens cells from oxidative damage ROS neutralization Neutralizes ROS Maintains lens clarity and function Nutrient supply Supplied by aqueous humor and vitreous humor Supports overall lens health Low oxygen maintenance Maintained by low oxygen in aqueous humor and vitreous humor Regulates factors like VEGF-A, crucial for lens transparency Aqueous humor reduction Reduces flow during sleep Enhances repair mechanisms during sleep The impact of sleep on aqueous humor flow and oxygen levels
-
The lens is an avascular transparent tissue, primarily obtaining its nutrients from the aqueous and vitreous humor. These humors are characterized by low oxygen levels, with oxygen content around 2% in the anterior segment of the aqueous humor and less than 1.5% in the posterior segment of the vitreous humor[121]. Unlike the cornea, the hypoxic environment surrounding the lens plays a key role in regulating its growth and maintaining its transparency. Research suggests that hypoxia regulates the expression of Vascular Endothelial Growth Factor in the lens[122]. Prolonged exposure to a high-oxygen environment can increase the risk of age-related nuclear cataracts, particularly in the elderly[123]. During sleep, the flow rate of aqueous humor slows to approximately 50% of its daytime rate, and oxygen content in both the aqueous and vitreous humor decreases further. This reduction in oxygen levels may facilitate the repair of oxidative damage accumulated during the day[35].
Melatonin and lens antioxidant response
-
Melatonin levels in the lens and atrial fluid are regulated by circadian rhythms. Research using radioimmunoassays to measure melatonin levels in the atrial fluid of male rabbits found significantly lower levels at 16:00 compared to 22:00 and 1:00[124]. Both in vivo and in vitro studies in mice have demonstrated that aralkylamine N-acetyltransferase activity and melatonin levels in the lens oscillate with a diurnal pattern, peaking at night[125]. This suggests that a biological clock controls aralkylamine N-Acetyltransferase activity in the rat lens. Elevated melatonin levels in the lens during sleep may contribute to an active antioxidant response. Additionally, estrogen levels also increase during sleep, providing further protection against oxidative damage to the lens through various mechanisms[126–127]. Therefore, prioritizing good sleep hygiene is crucial for reducing the risk of lens-related eye diseases, including cataracts, and for preserving overall visual health. Ensuring high-quality sleep allows the lens to repair and regenerate, protecting against oxidative damage and maintaining its clarity and function.
Sleep in choroidal health
-
The choroid, a highly vascularized tissue, is the primary blood supply for the retina's outer layers and the macula. Growing evidence highlights the choroid's critical role in regulating eye growth and the development of myopia. One key aspect of choroidal function is its circadian rhythm, with choroidal thickness peaking around midnight and reaching its lowest point around noon[128,129]. Studies show that myopic eyes tend to have thinner choroids, while hyperopic eyes have thicker choroids[130]. Importantly, choroidal thickness is negatively correlated with the eye's axial length[128]. During sleep, the choroid thickens, coinciding with a shortening of the axial length, a process that may significantly aid in myopia recovery.
In primate models, high-frequency A-scan ultrasonography has been used to measure diurnal changes in axial length. In younger, faster-growing rhesus monkeys, the axial length increases during the day and decreases at night, while in slower-growing adolescent monkeys, the opposite pattern is observed[131]. These findings suggest that choroidal changes during sleep contribute to the regulation of axial length, which plays a key role in myopia development and recovery.
Further clinical research indicates that sleep disorders, such as OSA, can affect choroidal thickness, potentially compromising visual health[132]. Notably, continuous positive airway pressure (CPAP) therapy for OSA has been shown to reverse some of these effects. SD, which is common among night shift workers, is also linked to changes in choroidal thickness, suggesting that inadequate sleep may adversely impact ocular health[133]. Sleep quality—including both duration and factors like insomnia—has been found to correlate with changes in choroidal structure, further emphasizing the broader relationship between sleep patterns and eye health[134].
Summary of key studies on sleep and ocular health
-
Given the extensive range of studies cited throughout the paragraphs above, a structured summary is provided below to facilitate cross-study comparison and enhance information accessibility (Supplymentary Table S5). This table presents representative research on the relationship between sleep and various aspects of ocular health, including IOP, retinal oxidative stress, corneal physiology, and lacrimal gland function. Studies are categorized by design, sample size, subject type (human vs animal), main findings, and noted methodological limitations. By synthesizing diverse evidence sources, this summary aims to support a clearer understanding of current research trends and gaps in the field of sleep–eye health interactions.
-
Recent literature has increasingly emphasized the link between sleep and major health outcomes, including cardiovascular disease, obesity, mental health issues, and neurodegenerative conditions. The association between sleep disorders and eye diseases has also begun to receive more attention[8,135,136]. Studies have shown that poor sleep can lead to ocular discomfort such as dry, painful, itchy, and stuffy eyes. Chronic SD not only exacerbates these symptoms but also alters the physiology and metabolism of the eye, potentially increasing the risk of various ocular diseases. OSA is the most common sleep disorder, with consistent evidence indicating that patients with OSA are at an increased risk of conditions such as blepharospasm, NAION, diabetic macular edema, and other retinal vasculopathies[8]. Supplementary Table S6 outlines the impact of different sleep disorders on ocular health, and the Table 2 outlines the common ways in which sleep disorders affect eye health.
Table 2. Common and uncommon pathways impacting subparts of the eye due to poor sleep.
No. Subpart of the eye Changes in tear film and components Relevant paper 1 Tear film Disrupted tear production due to immune dysregulation. Altered tear composition due to hormonal changes. Decreased tear clearance due to metabolic changes. 2 Intraocular
pressure (IOP)Immunological response affecting aqueous humor dynamics. Impact of sleep disturbances on ocular blood flow regulation. Reduced waste clearance affecting trabecular meshwork function. Sleep-related changes in scleral compliance influencing IOP. 3 Cornea Impaired epithelial regeneration due to metabolic changes. Reduced corneal sensitivity due to hemodynamic changes. Increased risk of inflammation due to immune activation. Corneal thickness variation due to sleep stages and IOP fluctuations. 4 Lens Accumulation of metabolic by-products (e.g., advanced glycation end-products). Changes in lens hydration levels due to circadian rhythm disruption. Oxidative stress leading to cataract formation. Reduced antioxidative enzyme activity linked to poor sleep. 5 Optic nerve Immunological responses leading to optic nerve inflammation. Hemodynamic changes impacting optic nerve head blood flow. Increased intraocular pressure influencing optic nerve head integrity. Altered venous pressure and autoregulation affecting optic nerve function. 6 Retina Impaired waste clearance leading to retinal cell dysfunction. Changes in retinal blood flow due to sleep-related hemodynamic shifts. Disrupted circadian rhythm affecting retinal cell metabolism. Altered neural signaling in the retina due to sleep deprivation. Eyestrain
Eyestrain: causes and mechanisms
-
Eyestrain is a common symptom of visual discomfort that results from environmental factors, eye diseases, refractive errors, or prolonged eye use. It is not a disease in itself but rather a set of symptoms caused by various factors, often related to refractive errors or excessive eye strain. The pathophysiological mechanism of eyestrain mainly involves overuse and stress on the eyes and visual system, which includes lens regulatory fatigue, eye muscle fatigue, dry eye symptoms, visual information processing stress, and neck and shoulder tension from poor posture. Chronic SD can exacerbate eye fatigue through multiple mechanisms, as illustrated in Supplementary Fig. S1.
Sleep deprivation's effect on muscle repair and metabolism
-
The repair of skeletal muscle, including extraocular muscles, is regulated by circadian rhythms. SD impairs the muscles' ability to rest and recover, contributing significantly to eyestrain. More than 2,300 genes in mammalian skeletal muscle exhibit circadian expression patterns, and these genes are involved in muscle function, including muscle formation, transcription, and metabolism[137]. SD increases the risk of muscle damage, with elevated serum levels of creatine kinase, aspartate aminotransferase, alanine aminotransferase, C-reactive protein (CRP), and myoglobin, which are indicators of muscle damage in athletes after exercise[138]. Moreover, disrupted circadian rhythms affect skeletal muscle repair and metabolism. Muscles play a key role in postprandial glucose uptake, with more than 80% of glucose being absorbed by skeletal muscle after meals. Sleep disturbances impair glucose metabolism, leading to reduced muscle repair, altered cognitive function, and mental fatigue[139], all of which contribute to eye strain. Studies in mice with deletions of the core clock genes Bmal1 and Clock have shown altered muscle pathology, including structural changes and impaired function, with reductions in mitochondrial volume and abnormal mitochondrial morphology[140]. Notably, Bmal1 has been shown to play a key role in regulating muscle stem cell expansion to influence muscle regeneration[141].
Impacts of sleep deprivation on growth hormone and muscle repair
-
GH and insulin-like growth factor-1 (IGF-1) act synergistically to inhibit protein hydrolysis and promote muscle performance, playing key roles in muscle repair and recovery[142]. GH is primarily secreted at night, with the peak of GH release occurring shortly after the first stage of slow-wave sleep[143]. Sleep regulates the secretion of GH and IGF-1, with SD resulting in minimal or no nocturnal GH release. Conversely, during recovery nights after SD, GH release increases significantly[143]. Animal studies have shown that SD impairs muscle regeneration and decreases muscle IGF-1 and plasma testosterone levels[143,144]. Additional sleep for more than a week increases circulating serum IGF-1 levels[144]. SD also increases the secretion of catabolic hormones and inflammatory responses, creating a highly protein-hydrolyzing environment that raises the risk of muscle damage[145].
Sleep deprivation and tear production: a link to eye discomfort
-
DED can lead to eye fatigue through multiple mechanisms. The cornea, one of the most richly innervated tissues in the body, is highly sensitive to external stimuli such as dust or foreign objects. This triggers reflexes like blinking to protect the corneal surface[146]. The tear film is critical for maintaining ocular surface lubrication and optical function. In individuals with DED, insufficient tear production, rapid tear evaporation, or shortened tear film breakup time expose the corneal surface to irritants[147]. Persistent dryness and friction lead to compensatory blinking, which increases the frequency of eye muscle accommodation, exacerbating eye muscle fatigue. In DED, the ocular surface lacks sufficient tear protection, making it more susceptible to oxidative damage, which triggers inflammation and further damages the ocular surface[148]. This disruption in ocular regulation contributes to eye fatigue, often accompanied by symptoms like redness, swelling, and pain.
Circulatory consequences of sleep deprivation on eye health
-
SD impairs blood circulation throughout the body, including the eyes, leading to insufficient oxygen and nutrient delivery to eye tissues, which can exacerbate eyestrain. SD hampers nitric oxide production, a key vasodilator, thereby reducing blood flow in microvessels such as those in the retina[149]. SD also increases oxidative stress, which accumulates ROS and free radicals, causing damage to cells and tissues, including ocular blood vessels and nerves. Furthermore, SD triggers an inflammatory response, releasing pro-inflammatory factors like TNF-α and interleukins, which impair ocular microcirculation and affect tissue health[150]. SD can also impair endothelial cell function, which regulates vascular tone, blood coagulation, and inflammation. Dysfunction of endothelial cells may lead to vasoconstriction, increased blood viscosity, and inflammation, further affecting the ocular blood supply. Additionally, SD disrupts neuromodulatory mechanisms by altering the balance between the sympathetic and parasympathetic nervous systems, further contributing to blood flow impairments[151].
Cognitive impairment and visual fatigue: the role of sleep
-
SD also impairs cognitive functions like attention and concentration[152]. These deficits necessitate greater brain energy and focus on visual tasks, further increasing visual fatigue. Collectively, these factors contribute to eye strain and discomfort, highlighting the importance of improving sleep quality to alleviate these burdens and reduce the risk of eyestrain. While factors like prolonged visual tasks and poor ergonomic practices contribute to eyestrain, significant gaps remain in understanding the precise pathophysiological mechanisms and the impact of external factors like lighting and screen use. Addressing these research limitations through well-designed studies is crucial to developing effective prevention and treatment strategies for eyestrain.
Critical analysis of literature limitations
-
A significant limitation in eyestrain research is sample bias, with many studies recruiting office workers or students who are more prone to visual stress due to prolonged screen use or intensive reading habits. This bias can skew results, exaggerating symptom prevalence within these groups and limiting generalizability. Additionally, measurement errors due to the subjective nature of symptoms like soreness and fatigue pose challenges for accurate assessment. Self-reported data also introduce variability, further undermining reliability. Moreover, confounding factors such as age, lifestyle, and concurrent medical conditions are often overlooked, complicating the identification of risk factors. Data interpretation bias also contributes to reinforcing pre-existing hypotheses without exploring alternative explanations, limiting the understanding of the underlying mechanisms behind eyestrain.
Dry eye disease
-
DED is one of the most prevalent ocular surface diseases worldwide. The tear film, essential for maintaining ocular surface health, consists of three layers: a lipid layer, an aqueous layer, and a mucin layer. The coordinated functioning of these layers helps to lubricate the ocular surface, prevent dryness, and protect the eye from foreign bodies and microbial invasion[153]. During normal sleep, the ocular surface is well-nourished and restored due to eyelid closure, reduced evaporation, and continued parasympathetic arousal that maintains basal tear secretion, despite tear production being reduced to approximately one-third of daytime levels[154]. However, sleep disorders disrupt tear homeostasis during sleep, altering hormonal levels and neural excitability, which increases the risk of developing dry eye. Studies have shown a causal relationship between sleep disorders and DED, creating a vicious cycle that exacerbates both conditions[70,155−157]. The possible mechanisms by which sleep disorders contribute to dry eye development are illustrated in Supplementary Fig. S2.
Sleep quality and dry eye disease
-
A study involving 71,761 participants aged 19-94 years used a dry eye questionnaire to assess dry eye symptoms. Results showed that individuals with poor sleep quality had a significantly higher incidence of dry eye than those with good sleep quality. Nearly 50% of patients with high levels of dry eye symptoms reported poor sleep quality, and they were 50% more likely to develop dry eye compared to those with good sleep quality. This relationship was observed in both younger and older adults, as well as in men and women[157]. Another survey of Korean adults found a significant association between reduced sleep duration and an increased prevalence of dry eye[158] Moreover, the sleep quality of dry eye patients improved after treatment with hyaluronic acid, mucin secretagogues, and steroids[159]. An experiment carried out at Xiamen University's Medical College Eye Research Institute (Xiamen, China) further revealed the pathological mechanism of SD-induced ocular surface damage: short-term SD triggers adaptive epithelial hyperplasia, while long-term sleep restriction leads to irreversible corneal stem cell loss and structural damage. This highlights sleep's key role in maintaining corneal integrity and suggests sleep disorders may promote dry eye development through oxidative stress-mediated pathways[83]. These studies suggest that SD plays a significant role in the development of DED, damaging the tear film through various mechanisms (Supplementary Table S7).
Damage of sleep deprivation on the function of the lacrimal gland
-
The lacrimal gland is responsible for secreting the aqueous components of the tear film, along with essential nutrients and functional proteins. Impaired lacrimal gland function can significantly affect the stability of the tear film, increasing the risk of dry eye. Studies have shown that subjects with SD exhibit a shortened tear film break-up time, hyperosmolarity of the tears, and reduced tear production compared to controls[160]. This dysfunction is an important factor in ocular surface pathology. SD impairs tear gland secretion by altering neurotransmitter levels, including acetylcholine and dopamine, both of which are essential for stimulating tear secretion. In sleep-deprived states, concentrations of these neurotransmitters in the lacrimal gland are significantly reduced[70]. Studies in mouse models have further demonstrated that SD leads to lacrimal gland dysfunction by accumulating ROS and disrupting metabolic, immune, and neural functions[71]. Additionally, SD causes pathological changes on the ocular surface that mimic dry eye symptoms, including decreased tear production, corneal sensory abnormalities, epithelial detachment, microvilli loss, corneal epithelial cell apoptosis, impaired tight junctions, and squamous metaplasia[70].
Gender differences and hormonal influence
-
DED has a gender-specific prevalence, with higher rates observed in women, particularly postmenopausal women[161]. This difference is largely attributed to sex hormones. While the role of estrogens in ocular surface health remains controversial, the protective effects of androgens are well-established[162]. Androgens, primarily derived from local tissue synthesis of androgen precursors, bind to androgen receptors in ocular surface tissues, where they help regulate meibomian gland function. This regulation increases meibomian gland lipid secretion, which is essential for maintaining tear film stability and reducing tear evaporation[163]. Studies have shown that androgen deficiency leads to lacrimal gland damage and increases the risk of dry eye[164], and androgen replacement therapy has shown therapeutic potential for aqueous-deficient and hyper-evaporative dry eye[165]. Furthermore, SD has been shown to decrease luteinizing hormone and testosterone levels by affecting the hypothalamic-pituitary-gonadal axis in mice[155]. Short-term SD also decreases serum testosterone levels[166], which may contribute to an increased risk of developing dry eye by affecting androgen levels.
Damage to the meibomian glands due to sleep deprivation
-
The secretory activity of the meibomian glands is partly regulated by circadian rhythms[167]. SD disrupts these rhythms, impairing secretion cycles and repair mechanisms. This disruption reduces the activation of hormone and growth factor pathways necessary for nocturnal repair, leading to decreased quantity and quality of the tear film lipid layer. SD not only disrupts systemic metabolic and immune homeostasis but also damages the meibomian glands through oxidative stress, inflammation, hormonal dysregulation, and neuroimmune disruption[168,169]. Excessive ROS and chronic inflammation caused by SD lead to functional impairment, apoptosis, gland atrophy, and acinar dropout, which reduce meibum production and accelerate tear evaporation[111]. Hormonal imbalances, particularly reduced androgen levels, further impairs the glands' resistance to oxidative and inflammatory damage[170].
Critical analysis of literature limitations
-
A critical analysis of the literature on the relationship between SD and DED highlights several limitations. Many studies suffer from sample bias, as they often focus on populations predisposed to DED, such as older individuals or those with preexisting ocular conditions, limiting the generalizability of the findings. Furthermore, the reliance on self-reported metrics to assess sleep patterns introduces measurement errors and undermines data reliability. The methodologies used to assess tear production and tear film integrity, such as Schirmer's test and tear break-up time, exhibit variability in sensitivity and specificity, adding ambiguity to the results. Additionally, many studies fail to adequately control for confounding factors such as age, gender, medication use, and environmental conditions, all of which independently influence both sleep quality and tear film characteristics. Finally, there is a tendency toward confirmation bias in data interpretation, with studies often reinforcing existing hypotheses without exploring alternative explanations or examining causal relationships between SD and DED. Addressing these limitations through more rigorous study designs and better control of confounding variables is essential to advance our understanding of the SD-DED relationship.
Impact of obstructive sleep apnea (OSA) on dry eye disease
-
OSA, a common sleep disorder characterized by intermittent pauses in breathing, leads to chronic intermittent hypoxemia, carbon dioxide retention, and disturbances in sleep stages. These disruptions can result in sympathetic arousal, indicating that OSA has systemic effects. Recent studies have shown an association between OSA and an increased incidence of dry eye. OSA patients have decreased Schirmer's test values, shorter tear film break-up times, higher Ocular Surface Disease Index scores, and increased meibomian gland loss, especially in severe cases of OSA[171,172].
A meta-analysis including 1,526 patients revealed that dry eye indicators were significantly worse in OSA patients compared to non-OSA patients, with a significant reduction in tear film break-up time and Schirmer's test scores and an increase in OSDI scores[173]. These findings suggest that OSA may increase the risk of developing DED through multiple mechanisms, including hypoxia and inflammation.
Potential mechanisms linking obstructive sleep apnea to dry eye disease
-
The precise mechanisms by which OSA causes dry eye are not fully understood, but several factors are likely involved. OSA is often accompanied by a systemic inflammatory response, characterized by increased levels of pro-inflammatory cytokines such as IL-6, CRP, IL-8, and TNF[174,175]. OSA is also associated with meibomian gland loss, which plays a crucial role in the pathogenesis of dry eye[176]. Additionally, eyelid ptosis, a common complication of OSA, may accelerate tear film evaporation by affecting tear film dynamics[177]. Most notably, hypoxic conditions during OSA episodes may induce several pathological changes in the lacrimal gland, leading to impaired tear production and composition. These changes are mediated by the activation of Hypoxia-inducible factor-α and Nuclear Factor-κB signaling pathways, downregulation of peroxisome proliferator-activated receptor α, myoepithelial cell damage, abnormal lipid accumulation, and increased apoptosis[76]. Finally, CPAP treatment for OSA has been associated with a significant increase in dry eye symptoms[178]. Therefore, OSA may be a potential risk factor for DED, with its severity influencing the degree of DED.
Limitations of the current research
-
Despite evidence linking OSA and DED, several limitations exist in the current research (Supplementary Table S8). A major issue is sampling bias, as studies often include OSA patients who may exhibit more severe symptoms or distinct physiological traits compared to the general population. Additionally, diagnostic measures such as the Schirmer test and tear film break-up time can be inconsistent, leading to varied results. Many studies also fail to account for confounding factors like age, obesity, and comorbidities, which can independently affect both OSA and tear film health. Furthermore, research often overlooks the indirect effects of OSA treatments, such as CPAP, which may influence tear film and ocular surface health. More comprehensive and controlled studies are necessary to better understand the relationship between OSA and DED and to develop effective interventions.
Knowledge gaps and future research needs
-
There are significant gaps in the literature regarding the relationship between OSA and DED. Longitudinal studies employing objective measures of both sleep disturbances and ocular health are urgently needed to better understand the causal pathways between OSA and DED. These studies should explore the effects of OSA treatments, such as CPAP, on dry eye symptoms and identify specific biochemical and cellular pathways affected by OSA. Research should also include a broader, more diverse patient population to assess the generalizability of findings and help develop inclusive clinical guidelines for managing patients with comorbid OSA and dry eye.
Myopia
Mechanisms of myopia development
-
Myopia, the most common refractive error in children and adults, develops through complex mechanisms influenced by both genetic and environmental factors. Two primary mechanisms drive the development of myopia: factors that affect IOP, ocular structure, development, and physiology, and those related to the central nervous system, particularly the regulation of circadian rhythms. Genetic factors are considered the major risk factors for myopia, but environmental influences, such as prolonged near work and poor lighting conditions, have also been shown to contribute to its development[179]. SD may further exacerbate myopia through multiple mechanisms (Supplementary Fig. S3).
Impact of sleep: Research over the past 15 years has established a link between insufficient sleep duration, poor sleep quality, and the development of myopia. Studies have found that children who sleep less than 7 h a night and go to bed late are more likely to develop myopia. Additionally, children with myopia tend to have shorter, more irregular sleep schedules compared to their non-myopic peers[180−182].
Circadian rhythms, which regulate sleep-wake cycles, are increasingly recognized as key factors influencing ocular growth and refractive development. Certain genes associated with refractive error are closely linked to circadian regulation, suggesting that circadian rhythm genetic modifiers may contribute to the pathophysiology of myopia and other refractive errors[179]. For example, the gene Cone-Rod Homeobox (CRX), essential for photoreceptor development, regulates genes involved in light-sensing and circadian signaling[183]. Paired box gene 4 (PAX4), involved in developmental processes, has also been associated with circadian rhythm modulation in retinal tissues[184]. Another key transcription factor, orthodenticle homeobox 2 (OTX2), regulates retinal cell differentiation and influences circadian processes through its regulation of light-sensing mechanisms[185]. These genetic interactions highlight the intricate relationship between circadian rhythm pathways and refractive development.
Animal studies further support the role of circadian rhythm disruption in myopia. For example, the deletion of the retina-specific clock gene Bmal1 in mice resulted in myopia[186]. Similarly, chicks exposed to light at night developed myopia, suggesting that disruption of the light-dark cycle adversely affects normal eye growth and leads to refractive error[187]. These findings underscore the importance of maintaining healthy sleep patterns and circadian rhythm stability in promoting normal ocular development.
Role of circadian rhythms
-
Circadian rhythms, regulated by neurohormones such as melatonin and neurotransmitters like dopamine, play a significant role in ocular growth and refractive development. Melatonin, a key regulator of the sleep-wake cycle, is synthesized and released by retinal photoreceptors under the control of retinal ganglion cells, which sense external light signals[188]. Elevated morning serum melatonin levels in myopic individuals, compared to non-myopic individuals, suggest that disruptions in melatonin signaling and circadian rhythms may contribute to the pathophysiology of refractive error[189].
Dopamine, another critical neurotransmitter, exhibits circadian variation in retinal tissues and plays a central role in modulating eye growth. Light exposure stimulates dopamine release, which inhibits axial elongation, a key factor in myopia progression[187]. Dysregulation of dopamine signaling, particularly under conditions of abnormal light exposure or circadian rhythm disruption, has been linked to uncontrolled axial elongation and increased myopia risk.
Additionally, ocular parameters such as IOP, choroidal thickness, and axial length demonstrate circadian patterns[186]. These rhythmic variations in ocular physiology interact with neurohormonal regulators like melatonin and dopamine, further supporting the hypothesis that circadian rhythms play a pivotal role in refractive development. While the evidence underscores the importance of circadian regulation, the precise interplay between melatonin, dopamine, and ocular biomechanics remains an active area of investigation.
While growing evidence points to the potential role of sleep and circadian rhythms in myopia development, current research does not conclusively establish insufficient sleep as a direct risk factor for myopia. Further studies are needed to clarify this relationship and explore whether interventions targeting sleep patterns can effectively prevent or mitigate myopia progression in susceptible populations. Such research could lead to novel preventive strategies that incorporate both ocular health and improvements in sleep quality, offering new avenues for addressing myopia in both children and adults.
Glaucoma
-
The pathologic elevation of IOP is a critical factor in the development of glaucoma; however, some patients continue to experience disease progression even when IOP is lowered[190]. Recent research suggests that insufficient sleep duration and poor sleep quality may exacerbate glaucoma progression[191]. While much of the focus has been on circadian dysregulation in glaucoma patients, particularly due to retinal ganglion cell damage, circadian disruptions may also play a role in the onset and progression of the disease. Studies have shown an association between sleep duration, sleep quality, and glaucoma risk[192]. Individuals with shorter sleep duration tend to have a higher prevalence of glaucoma, especially those with obesity[193]. Furthermore, sleep disturbances, such as shorter sleep duration and later sleep stages, have been linked to retinal ganglion cell loss and dysfunction, which are characteristic of glaucoma[194]. Although the evidence does not establish a direct causal relationship, disrupted sleep patterns may exacerbate glaucoma progression through mechanisms such as increased IOP and impaired retinal perfusion. IOP, BP, ocular perfusion pressure, and melatonin secretion, all of which are influenced by circadian rhythms, are strongly associated with the onset and progression of glaucoma (Supplementary Fig. S4).
Effect of circadian changes in intraocular pressure, blood pressure, and retinal perfusion pressure on glaucoma
-
SD influences IOP through various biochemical pathways, significantly altering circadian rhythms that are essential for eye health. The circadian regulation of IOP involves interactions between melatonin secretion and body temperature cycles, both of which are affected by sleep patterns. Key neuroendocrine signaling molecules, including NE and dopamine, regulated by the hypothalamus and the photic-sensitive suprachiasmatic nucleus, play a crucial role in this regulation.
The balance between aqueous humor production and outflow is vital for maintaining normal IOP. In humans, this process is regulated by circadian rhythms[195]. During SD, the rate of aqueous humor flow is significantly faster than during normal sleep, although it does not reach daytime levels[196], which may contribute to an increase in IOP overnight. Several factors associated with glaucoma development, including IOP, BP, and ocular perfusion pressure, exhibit circadian rhythms[32]. For instance, IOP in healthy individuals fluctuates within a range of 4-6 mmHg during the day[197], whereas in glaucoma patients, IOP fluctuates between 6-15 mmHg[198]. Larger IOP fluctuations increase the probability of glaucomatous visual field loss progression by 30%[199]. Additionally, primary glaucoma patients with greater IOP fluctuations show significantly faster disease progression[200]. Retinal ganglion cell loss in primary open-angle glaucoma has also been linked to disruptions in 24-h IOP rhythms and misalignment with circadian body temperature rhythms[201]. Interestingly, while there is no significant difference in nocturnal BP between glaucoma patients and healthy controls, significant fluctuations in nocturnal BP have been observed in patients with progressive glaucoma[202]. Diurnal variations in IOP and BP lead to an increased decline in nocturnal ocular perfusion pressure, further accelerating RNFL thinning and visual field defects in glaucoma patients[203]. These studies suggest that insufficient sleep duration or poor sleep quality leads to increased IOP and retinal perfusion pressure fluctuations, which may accelerate disease progression in glaucoma patients.
Potential mechanisms of optic nerve damage by sleep deprivation
-
SD can adversely affect optic nerve health through various biochemical pathways, including alterations in cellular metabolism, stress responses, inflammatory reactions, and hemodynamic changes. Initially, SD leads to increased oxidative stress in the body, as the majority of the body's repair and metabolic clearance processes occur during deep sleep. Normally, sleep helps to reduce the generation of free radicals and enhances antioxidant activity.
SD also disrupts the regulation of neurotransmitters, which play a key role in maintaining optic nerve health. Sleep modulates the synthesis and release of various neurotransmitters and neuromodulators, such as serotonin, NE, and dopamine, which influence the function and survival of retinal cells. Disruptions in their normal cycles due to SD can negatively impact the metabolism and function of optic nerve cells[204]. SD increases the expression of pro-inflammatory cytokines, such as TNF-α and IL-6, which are key mediators of inflammation. Studies in humans and animal models show that SD induces a systemic inflammatory response by increasing circulating levels of these cytokines, which contribute to oxidative stress and tissue damage[205,206]. Chronic elevation of TNF-α and IL-6 may disrupt homeostasis in retinal tissues, accelerating neuroinflammatory processes and increasing the risk of retinal dysfunction. While these inflammatory responses may be beneficial in the short term for fighting infections or injuries, prolonged elevation of these cytokines can lead to optic nerve damage, including apoptosis or dysfunction of optic nerve cells.
Moreover, SD affects cardiovascular regulation, potentially leading to BP fluctuations and unstable blood flow, which may compromise the optic nerve's blood supply. The optic nerve depends on a stable blood supply for oxygen and nutrients; any reduction in blood flow can result in dysfunction or even death of optic nerve cells[207]. Lastly, SD can impair cellular energy metabolism, reduce ATP production, and decrease the cell's ability to repair. The process of autophagy, responsible for clearing damaged proteins and organelles, is also regulated by the sleep cycle. Without this process, damaged and dysfunctional cells accumulate, increasing the risk of cell death[208].
Effect of reduced melatonin secretion on glaucoma
-
Melatonin, a key regulator of the sleep-wake cycle, plays an essential role in physiological regulation, immune function, and ocular health. Recent studies indicate that melatonin not only influences sleep but also plays a role in regulating IOP, protecting the retina from glaucomatous damage, and potentially reversing existing damage. Melatonin secretion is regulated by external light, with retinal ganglion cells transmitting light signals via the retinal-hypothalamic tract to the suprachiasmatic nucleus, which in turn stimulates or inhibits melatonin release from the pineal gland[209,210]. Ocular structures, including the retina, lens, and ciliary body, also secrete melatonin[211−213], adding an extra layer of regulation within the eye. However, SD and modern lifestyle factors, such as exposure to indoor light or electronic screens before bedtime, disrupt melatonin secretion. This desynchronization may delay melatonin release and shorten its maintenance, negatively impacting the biological clock and ocular health[188].
Melatonin lowers intraocular pressure
-
Recent studies have shown that melatonin levels in the eye are significantly increased in individuals with elevated IOP and in glaucoma patients[214]. Both melatonin and its analogues have been found to lower IOP significantly in both animal models and human studies[215−218]. This suggests that melatonin regulates IOP through a negative feedback mechanism, maintaining IOP within normal levels. Elevated IOP activates TRPV4 receptors in the non-pigmented epithelial cells of the ciliary body, which stimulates increased melatonin synthesis[219]. In turn, melatonin reduces aqueous humor production by decreasing chloride ion flow from the cytoplasm of nonpigmented epithelial cells to the extracellular space, thereby lowering IOP[220]. Additionally, melatonin modulates IOP indirectly by reducing β2-adrenergic receptor expression in nonpigmented epithelial cells of the ciliary body[221]. These findings highlight melatonin's role in regulating IOP and suggest its potential therapeutic value in managing glaucoma and controlling IOP.
Optic neuroprotective effects of melatonin
-
Melatonin offers various mechanisms to protect the optic nerve. However, SD or poor sleep quality can impair melatonin secretion. Studies in glaucoma patients have shown reductions in the amplitude of the P50 and N95 waves in electroretinograms, which are indicators of macular function and retinal ganglion cell health[222,223]. Oral melatonin treatment for 90 d has been shown to significantly increase P50 and N95 amplitude and shorten latency, suggesting that melatonin may help repair retinal ganglion cell dysfunction caused by primary open-angle glaucoma[224]. Additionally, in a rat glaucoma model induced by hyaluronic acid, melatonin significantly reduced retinal ganglion cell loss after 10 weeks of elevated IOP[225].
Melatonin's neuroprotective effects can be categorized into two key areas: (1) Antioxidant effect: Melatonin acts as a potent antioxidant in the retina, protecting RPE cells from oxidative damage by reducing ROS levels, inhibiting pro-oxidant enzymes, and stimulating the secretion of antioxidant enzymes[226−229]. (2) Anti-excitatory neurological damage: Melatonin mitigates excitatory nerve injury by increasing glutamate uptake, decreasing glutamate synthesis, and reducing glutamate deaminase activity, thereby lowering glutamate concentrations in the synapse[225]. These properties make melatonin a promising therapeutic agent for glaucoma and optic nerve protection.
Link between obstructive sleep apnea and glaucoma
-
Emerging research has shown a significant association between OSA and an elevated risk of developing glaucoma, particularly concerning RNFL thinning and glaucoma progression. A meta-analysis involving 48 studies and over 4.5 million patients found that OSA patients were 3.66 times more likely to develop glaucoma compared to non-OSA patients, even after adjusting for confounders such as age, sex, and comorbid conditions like hypertension and diabetes[230]. The study also found that OSA patients had significantly higher IOP and reduced RNFL thickness, with the severity of OSA correlating with the degree of IOP elevation and RNFL thinning. These findings suggest that OSA contributes to glaucoma risk through mechanisms such as hypoxic injury, vascular changes, and oxidative stress.
The precise mechanisms by which OSA causes glaucoma and DED are not fully understood, but several factors are believed to be involved, including systemic inflammation, MGD, and hypoxia. OSA is often associated with increased systemic inflammatory markers, including IL-6 and CRP, which can damage ocular tissues and contribute to the development of glaucoma and dry eye[174,175].
Additionally, OSA-induced hypoxia leads to inflammation and oxidative stress, activating signaling pathways such as HIF and NF-κB, which promote endothelial dysfunction and vascular damage, further impairing retinal function and accelerating glaucoma progression[231−234].
In conclusion, the relationship between sleep, circadian rhythms, and glaucoma is complex and requires further exploration. Evidence suggests that sleep disturbances, particularly SD and OSA, can disrupt IOP regulation, retinal perfusion, and neuroprotection, all of which contribute to glaucoma progression. Melatonin plays a crucial role in regulating IOP and protecting the optic nerve, making it a potential therapeutic target for glaucoma treatment.
Cataract
-
Cataract remains the leading cause of blindness in middle- and low-income countries. Cataractogenesis is a complex process that involves multiple environmental, metabolic, genetic, and nutritional factors affecting the lens over time. Oxidative damage from free radicals is widely recognized as a major mechanism driving cataract formation[235]. Additionally, excessive apoptosis of lens epithelial cells and abnormal lens proteins are significant contributors to cataract development. Recent studies have suggested that shorter sleep duration may increase the risk of developing cataracts, though the exact mechanisms remain unclear. It is hypothesized that SD may influence cataract formation indirectly through several pathways (Supplementary Fig. S5). For example, SD increases sympathetic nerve activity, activating the Hypothalamic-Pituitary-Adrenal axis, leading to excessive steroid hormone secretion by the adrenal cortex[236]. These endocrine changes may cause systemic oxidative stress and inflammatory responses[80], which could negatively impact lens health and increase cataract risk. Further studies are needed to clarify the relationship between SD and cataractogenesis and to develop effective preventive strategies.
Role of systemic inflammatory response in cataract
-
Studies have demonstrated that SD can lead to elevated levels of pro-inflammatory cytokines such as TNF-α, IL-10, and CRP in peripheral blood, suggesting that SD may trigger systemic inflammation[29]. Notably, a significant association has been found between serum IL-6 levels and the incidence of nuclear cataracts[237]. This finding supports the hypothesis that SD may indirectly influence cataract formation by inducing a systemic inflammatory response.
However, the relationship between systemic inflammation and cataracts has been inconsistent. For example, one study comparing urinary levels of 15-keto dihydro-PGF2a, a marker of systemic inflammation, in patients with age-related cataracts and the normal population found no significant association[238]. Similarly, another study found no correlation between serum CRP levels and cataract development in individuals aged 40 to 80 years[239]. These inconsistencies may be due to variations in participant age or study design, highlighting the need for further research using more reliable inflammatory markers, comprehensive assessments of inflammatory pathways, and more precise experimental studies in cataractogenesis. While there is growing evidence linking SD and cataract development, further studies are needed to fully understand the underlying mechanisms. The role of oxidative stress, systemic inflammation, and disrupted circadian rhythms in cataractogenesis highlights the potential impact of sleep quality on lens health.
Role of systemic oxidative stress in cataract
-
As an important refractive structure, the maintenance of transparency of the lens is critical for maintaining normal visual function. Under normal conditions, the body maintains a balance between oxidants and antioxidants, which is essential for cellular survival and health. SD has been shown to decrease GSH levels in the body[240], which is particularly concerning because GSH is a crucial antioxidant in the lens, preventing protein cross-linking and aggregation that would otherwise lead to oxidative damage[241,242].
As individuals age, GSH levels in the lens decrease, while hydrogen peroxide levels increase, contributing to the oxidative imbalance that plays a key role in the development of age-related cataracts[243]. In animal models of induced cataracts, oxidative stress has been shown to cause structural and biochemical changes in the lens, resembling those seen in age-related cataracts[244]. While oxidative stress promotes neovascularization in tissues such as the cornea and choroid, its role in cataractogenesis is primarily through inducing lens epithelial cell apoptosis, protein aggregation, and oxidative damage to lens fiber cells. These processes lead to lens opacification and degeneration, rather than neovascularization.
However, it is unclear whether oxidized substances in plasma can cross the blood-ocular barrier and damage ocular tissues such as the aqueous humor and lens during a systemic inflammatory response. Studies comparing plasma levels of antioxidants (e.g., vitamins C, E, and carotenoids) in cataract patients and healthy individuals have shown that higher blood antioxidant levels correlate with a lower risk of cataract development[245,246]. Furthermore, patients with age-related cataracts are more likely to have a history of corticosteroid use or diabetes, both of which are associated with an increased risk of cataract[247].
Critical analysis of literature limitations
-
While existing research has provided valuable insights into the potential association between sleep patterns and cataract risk, several limitations need to be addressed. A primary concern is sample bias, as many studies rely on self-reported sleep durations and symptoms, which may not accurately reflect true sleep patterns or the severity of cataract symptoms. This reliance on subjective reports can introduce potential sources of bias and measurement errors. Additionally, confounding factors such as age, smoking, UV exposure, and other health conditions are often insufficiently controlled in studies, which may independently influence both sleep quality and cataract development. Another limitation is the lack of longitudinal data, as most studies are cross-sectional or retrospective, limiting the ability to establish causality between SD and cataract onset. These methodological constraints underscore the need for better-designed studies to enhance our understanding of the mechanisms linking SD and cataracts.
Changes in circadian rhythms and sleep patterns after lens implantation
-
Recent studies have highlighted a significant association between cataract surgery and changes in circadian rhythms and sleep patterns. For example, Feng et al. demonstrated that a blue light-filtering intraocular lens improved sleep quality in patients compared to a standard intraocular, primarily through modulation of circadian rhythms[248]. Similarly, Giménez et al. reported improved circadian rhythm synchronization following cataract surgery, attributing this improvement to better light transmission and sleep-wake cycle resetting in older patients[249]. While Giménez et al. did not specifically assess sleep quality, Chellappa et al. provided further evidence by showing that cataract surgery, particularly with UV-only blocking lenses, enhanced circadian rhythm stability, cognitive function, and overall sleep quality in older adults[250]. Additionally, Turner et al. found that cataract-induced visual impairment significantly increased the prevalence of short sleep syndrome, characterized by less than five hours of sleep per night[250]. These findings suggest that cataract surgery, especially with the use of IOLs, may benefit sleep patterns and circadian rhythms, illustrating the complex relationship between ocular health and sleep quality.
Diabetic retinopathy
-
DR is a prevalent complication of diabetes mellitus, characterized by irreversible retinal damage. Alongside microvascular alterations, retinal inflammation, and neurodegenerative processes are closely linked to DR pathogenesis. Epidemiological evidence indicates that about one-third of diabetic individuals exhibit varying levels of DR severity, with its incidence increasing alongside the global rise in diabetes prevalence. DR has emerged as the leading cause of blindness in working-age adults worldwide[251,252]. SD may elevate the risk of DR complications in diabetic patients through various mechanisms (Fig. 3). For instance, a study revealed that diabetic men who slept fewer than 5 ho or more than 9 h exhibited a heightened risk of DR[253]. Additionally, in adolescents with type 1 diabetes, inadequate sleep, and poor sleep quality were associated with insulin resistance, poor glycemic control, and microvascular complications[254]. These findings suggest that SD could impact the initiation and progression of DR, potentially by disrupting circadian rhythms.
Figure 3.
Mind map diagram illustrating the mechanisms through which sleep deprivation impacts diabetic retinopathy. (Selected artwork (retina) depicted in the figure is derived from or adapted from images provided by Servier Medical Art (Servier;
https://smart.servier.com ), licensed under a Creative Commons Attribution 4.0 Unported License).Although studies linking SD to vision-threatening diabetic retinopathy (VTDR) have been inconclusive, some indicate poorer sleep quality among VTDR patients[255], while others report no significant link between SD and VTDR[253]. These inconsistencies underscore the need for further exploration to clarify the relationship between SD and DR. Recent research has also shown that SD can induce systemic pathologies, including abnormal glucose metabolism, oxidative stress, and elevated BP, which contribute significantly to the onset and progression of DR. Therefore, comprehensive investigations into the SD-DR relationship are crucial for DR prevention and management.
Recent research has revealed that SD can induce systemic pathologies, including abnormal glucose metabolism, oxidative stress, and elevated BP, contributing significantly to the onset and progression of DR. Thus, conducting comprehensive investigations into the SD-DR relationship is crucial for DR prevention and management.
Possible mechanisms of sleep deprivation causing diabetic retinopathy
-
Circadian rhythms play a vital role in regulating various physiological processes, including glucose metabolism. SD and poor sleep quality are major causes of circadian rhythm disruption, which negatively affect β-cell function and insulin sensitivity, thereby impacting glucose tolerance in diabetic patients[256]. Brief periods of SD, even as short as a single night, have been shown to reduce insulin sensitivity[257].
SD not only affects glycemic control in diabetic patients but may also worsen the progression of DR. Poor glycemic control is a primary contributor to DR onset and progression. Hyperglycemia triggers apoptosis of retinal cells and the formation of microaneurysms, leading to retinal capillary occlusion and ischemia[258−260]. In the early stages of DR, there is often an increase in retinal blood flow velocity and overall blood flow[261]. However, when this increased blood flow is coupled with hyperglycemia, it can cause capillary occlusion, vasodilation, and leakage, further exacerbating DR progression[262].
Intensive hyperglycemic treatment has been shown to significantly decrease DR incidence compared to conventional treatment methods[263]. While extensive evidence links SD to increased diabetes risk, studies specifically focusing on the effects of SD on DR remain inconclusive. These findings highlight the need for thorough investigations into how circadian rhythms, sleep quality, and glycemic control interact in the pathogenesis of DR.
Oxidative stress and inflammation
-
Sleep plays a critical role in enhancing antioxidant capacity in the brain and retina, while also regulating immune function. Insufficient sleep can lead to oxidative stress and uncontrolled inflammation, both of which are tightly linked to DR pathogenesis[8,264]. Oxidative stress and inflammation play central roles in DR, particularly during the non-proliferative and proliferative stages, where chronic low-grade inflammation of the retina is often observed[265].
Inflammation is especially critical in the early stages of DR, where the accumulation of inflammatory mediators can lead to neuronal death and the formation of new blood vessels in the retina[266]. Additionally, the excessive accumulation of ROS not only damages retinal vessels and surrounding tissues but may also impair pancreatic β-cell function, further disrupting blood glucose regulation[267]. Studies have shown that inhibiting the release of pro-inflammatory factors and limiting ROS generation is crucial for reducing vascular and neurodegenerative changes caused by diabetes, which may help reduce visual impairments and retinal neuronal apoptosis[268−270].
In summary, insufficient sleep may exacerbate local inflammatory responses in the retina, damaging retinal tissue and accelerating the progression of DR. Anti-inflammatory therapies may therefore be an effective preventive strategy for managing DR, and further research into these mechanisms is needed to develop more effective treatment strategies.
Elevated blood pressure
-
Hypertension is a significant risk factor for DR. Research indicates that SD may contribute to hypertension and increased BP fluctuations by disrupting autonomic nervous system balance and circadian rhythms[271]. A study examining the impact of BP control on DR in type 2 diabetes patients randomly assigned participants into two groups: one group-maintained BP below 150/85 mmHg, while the other group-maintained BP below 180/105 mmHg. Results showed significantly fewer retinal microaneurysms in patients with BP controlled below 150/85 mmHg compared to those with less stringent BP control[272].
Recent research also found that diabetic patients with severe DR had higher 24-h BP and greater variability in night time systolic BP compared to those without DR or with mild DR[273]. Additionally, hypertension can induce vascular morphological changes, such as hard exudates, cotton wool spots, and retinal hemorrhages, which resemble changes seen in mild to moderate non-proliferative DR[274]. These findings underscore the role of hypertension in DR development and progression.
Depression
-
Chronic SD has been shown to affect mood and trigger depressive-like behaviors[275,276]. Studies indicate a correlation between sleep quality, depression severity, and changes in RNFL and macular thickness[277]. A comprehensive analysis of National Health and Nutrition Examination Survey (NHANES) data (2005−2006 to 2017−2018) found that mild to moderate depression is an independent risk factor for increased DR prevalence in adults[278]. A prospective cohort study also found a link between depression severity and increased DR risk in diabetic patients[279]. Depression may impair retinal function, particularly initial visual processing capabilities and treatment with ketamine for depression has been shown to promote retinal neuroplasticity[280,281]. While the relationship between depression and DR is not fully understood, depression may contribute to DR development in diabetic patients.
While there is evidence linking SD and DR, the relationship remains complex and not fully understood. Understanding the mechanisms by which SD influences DR could lead to new preventive and therapeutic strategies for managing DR in diabetic patients, which is essential as the global prevalence of diabetes continues to rise.
Obstructive sleep apnea and diabetic retinopathy
-
The association between OSA and type 2 diabetes is well-established, with OSA contributing to increased insulin resistance and abnormal glucose tolerance. However, the relationship between OSA and DR remains more complex and is not fully understood. A 2009 cross-sectional case-control study indicated that a 4% oxygen desaturation index is an independent risk factor for proliferative DR (R = 0.43, p = 0.03)[282]. A longitudinal cohort study from the UK supported this, showing that OSA remained associated with vision-threatening DR and diabetic macular edema (DME) even after adjusting for confounding factors[283].
A systematic review and meta-analysis of 28 studies with 201,567 participants found that OSA patients had a 34% increased risk of developing DR compared to the control group (95% CI: 1.07−1.69). For patients under 60 years, the association between OSA and DR was even stronger (OR = 1.60, 95% CI: 1.25−2.06)[284]. Although most studies included in the meta-analysis were cross-sectional, these findings still suggest an important relationship between OSA and DR.
Future research directions
-
Several key directions for future research exist in the fields of myopia, glaucoma, and DR.
Myopia: Current evidence on the relationship between sleep and myopia remains inconsistent. It is critical to clarify how SD influences eyeball growth and refractive development, particularly by examining the roles of neurohormones like melatonin and dopamine.
Glaucoma: Longitudinal cohort studies are needed to determine how abnormal sleep patterns accelerate optic nerve damage through pathways such as IOP fluctuations and autonomic nervous system dysfunction. Research should also explore underlying mechanisms and potential interventions to understand and mitigate the impact of sleep disorders on glaucoma progression.
DR: Combining polysomnographic monitoring with retinal metabolomics analysis will help reveal how sleep fragmentation exacerbates oxidative stress and blood-retinal barrier disruption. In-depth mechanistic studies are necessary to clarify the role of sleep disorders in the pathophysiology of DR, especially the functions of melatonin and other sleep-related hormones and neurotransmitters.
-
The interdisciplinary treatment model that integrates sleep medicine and ophthalmology is grounded in the concept that sleep quality directly influences eye health. Optimizing sleep quality can lower IOP and improve retinal circulation, potentially slowing the progression of diseases such as glaucoma and DR. As in the study by Tejero-Garcés et al., an interdisciplinary approach combines the diagnostic expertise of sleep medicine specialists with the therapeutic skills of ophthalmologists, resulting in more comprehensive and effective treatment plans[285]. This collaboration helps treat both sleep and eye diseases, offering a broader, more holistic approach to patient care.
In recent years, the advancement of smart wearable technology has offered innovative approaches for the integrated management of ocular health and sleep quality. For example, sensor-equipped contact lenses enable continuous 24-h monitoring of dynamic IOP curves, providing critical data for personalized glaucoma treatment[98]. Head-mounted eye-tracking devices allow for persistent monitoring of ocular movement trajectories[286]. Additionally, wearable epidermal patches capable of simultaneously tracking hemodynamic and metabolic biomarkers enhance our understanding of physiological responses and disease mechanisms[287]. These technological innovations in wearable devices facilitate uninterrupted health monitoring, highlighting the potential of modern medical technologies in advancing disease management and prevention. By bridging theoretical insights with practical applications, this approach not only improves clinical outcomes but also elevates the quality of life for patients with ocular disorders.
Fostering interdisciplinary research collaboration between sleep medicine and ophthalmology is essential for improving treatment outcomes. By integrating expertise and resources from both fields, research teams can develop innovative treatment modalities and investigate how specific sleep interventions may improve outcomes for eye diseases. Furthermore, targeted clinical trials that assess the efficacy of integrated sleep management strategies for ophthalmic patients will help validate these interventions and provide a solid scientific foundation for their future clinical application[288].
Through these approaches, researchers can deepen our understanding of the complex interactions between sleep disorders and ocular diseases, offering more comprehensive treatment options and advancing the integration of ophthalmology and sleep medicine.
Guidelines and recommendations
Recommendations for clinicians
Routine sleep disorder screening
-
It is essential for clinicians to routinely screen patients with chronic eye diseases, such as glaucoma, DR, and AMD, for sleep disorders. The significant correlation between these eye diseases and sleep disturbances, especially sleep apnea, underscores the importance of screening. Sleep apnea can profoundly impact ocular health and overall well-being, making early detection crucial. The STOP-BANG questionnaire, a validated screening tool, is recommended for identifying high-risk patients who may require further evaluation through a sleep study. This simple, yet effective, screening method is proven to be a reliable tool for detecting OSA in ophthalmic settings.
Integrating sleep management into eye care protocols
-
It is recommended that eye care protocols incorporate sleep management. This involves collaboration among an interdisciplinary team, including sleep medicine specialists, ophthalmologists, and primary care providers. By developing a comprehensive treatment program that addresses both the management of eye diseases and the improvement of sleep quality, clinicians can provide more effective, patient-centered care. For patients diagnosed with OSA-related eye diseases, CPAP therapy is particularly recommended. CPAP therapy not only significantly improves sleep quality but may also slow the progression of eye diseases, such as glaucoma and retinopathy, by enhancing nocturnal IOP and improving ocular blood flow.
Recommendations for researchers
-
Researchers should intensify their investigations into how sleep quality impacts the progression of eye diseases, with a particular focus on the relationship between sleep disorders and ocular health. Studies should explore how various types and severities of sleep disorders, such as OSA, influence eye disease progression through key physiological factors like IOP and retinal blood flow. These studies are critical for managing chronic eye conditions like glaucoma and DR, which are significantly associated with sleep disturbances[8].
With the growing accumulation of healthcare data and advances in analytical tools, leveraging big data and artificial intelligence to analyze the complex relationship between sleep disorders and eye diseases is increasingly feasible. These technologies can enhance the precision of research and assist in developing more targeted prevention and treatment strategies, particularly for early intervention in high-risk populations. By applying advanced data analysis techniques, researchers can uncover more nuanced insights into the sleep-eye disease link and improve clinical decision-making.
-
Current research on the relationship between sleep disorders and ocular health suffers from significant knowledge gaps and methodological limitations, demanding a systematic breakthrough. A primary issue is the lack of standardization in objective measurement technologies. Inconsistent results often arise from differing measurement approaches; for instance, IOP monitoring during sleep varies methodologically. While tonometry-based methods require subject awakening, contact lens sensors enable continuous monitoring but may disrupt corneal hydration.
Establishing causality between sleep and ocular health necessitates large-scale longitudinal studies. These should track sleep patterns and ocular parameters across age groups while strictly controlling for confounding factors like socioeconomic status and comorbidities. Mechanistically, research should focus on how circadian rhythm disruptions affect the ocular surface microenvironment, diurnal IOP patterns, and lens antioxidant defense systems at the molecular level.
Many studies have small, non-diverse samples, particularly in glaucoma and dry eye research. OSA-related studies, mostly clinic-based, suffer from selection bias, limiting their generalizability. Current evidence, predominantly cross-sectional, makes causal inference difficult. There's also a lack of interventional data; for example, whether sleep improvements like OSA treatment or better sleep hygiene can slow ocular disease progression remains unverified.
Rodent models dominate circadian rhythm research, but their nocturnal physiology differs greatly from humans. Comparative studies in diurnal models (e.g., non-human primates, rabbits) are needed for a more human-relevant understanding of ocular physiology. Clinical studies often overlook key confounders such as light exposure, medication use, systemic inflammation, and cardiovascular risks, which can impact IOP regulation and retinal metabolism.
In summary, current research on the relationship between sleep disorders and ocular health has many problems. The lack of standardization in measurement techniques leads to inconsistent results. Large-scale longitudinal studies are urgently needed to establish causal relationships. Samples have limitations and causal inference is difficult. At the same time, there is a lack of interventional data. Although rodent models are widely used, there are significant physiological differences between them and humans, and clinical studies often overlook key confounding factors. To achieve a breakthrough in this field of research, systematic improvements in methodology are required. More targeted and scientific research should be carried out to fill knowledge gaps, better understand the complex relationship between sleep and ocular health, and provide a solid basis for the prevention and treatment of related diseases.
-
The relationship between sleep and ocular health is gaining increasing attention. Future research will focus on the following key areas:
Circadian rhythm disruption and ocular microcirculation: Investigate the mechanisms by which circadian misalignment impacts ocular hemodynamics, including abnormal regulation of ciliary artery blood flow, altered diurnal fluctuations in IOP, and dynamic changes in choroidal thickness.
Sleep interventions and ocular disease risk: Design multicenter randomized controlled trials to evaluate the preventive effects of standardized sleep interventions on ocular complications in high-risk populations. Assess whether exogenous melatonin supplementation reduces retinal oxidative stress and mitigates circadian-related retinal degeneration.
Clinical integration of sleep monitoring technologies: Leverage optical coherence tomography angiography to explore correlations between real-time sleep parameters and choroidal blood flow measurements. Develop sleep-based predictive models for the early detection of ocular diseases.
Personalized sleep medicine in ocular disease management: Investigate the efficacy of individualized sleep management strategies in optimizing treatment outcomes for ocular diseases. Integrate multimodal sleep monitoring and dynamic interventions into ophthalmic clinical pathways to improve patient-reported visual function and objective disease progression metrics.
Design of methodological pathways for future research
-
Longitudinal cohort studies: Aim to track the impact of chronic sleep disorders on the incidence and progression of ocular diseases. Example: A 5- to 10-year follow-up study comparing shift workers and daytime workers to assess glaucoma risk, IOP fluctuations, and RNFL thinning.
Randomized controlled trials: Evaluate whether sleep interventions (e.g., CPAP for OSA, melatonin supplementation, sleep hygiene optimization) prevent or slow ocular disease progression. Example: A placebo-controlled trial testing melatonin's ability to improve retinal microcirculation and reduce oxidative stress in AMD patients.
Circadian gene expression and molecular studies: Investigate how clock gene dysregulation (e.g., BMAL1, CLOCK, PER2) affects retinal health and IOP modulation. Example: Analyzing gene expression in aqueous humor and retinal tissues from sleep-deprived vs control animal models.
Advanced imaging and wearable monitoring technologies: Explore real-time associations between sleep patterns and ocular physiological changes. Example: Using OCTA to measure choroidal blood flow variations in individuals with irregular sleep-wake cycles.
Conclusion
-
In summary, exploring these research avenues will reveal more about the intricate relationship between sleep and ocular health. This knowledge is expected to lead to new strategies for the prevention and treatment of ocular diseases, with the potential to significantly improve patient care.
-
Circadian rhythms play a fundamental role in regulating the physiological activities of the eye, with sleep synchronized to the biological clock being essential for normal ocular function. This review has explored the relationship between sleep disorders and various eye diseases (Fig. 4), although conclusive results are still lacking. Short-term SD or poor sleep quality may lead to eye fatigue, dryness, and discomfort. Long-term sleep deficiency, however, can increase the risk of more serious eye diseases through mechanisms such as disrupted melatonin secretion, systemic oxidative stress, and inflammatory responses.
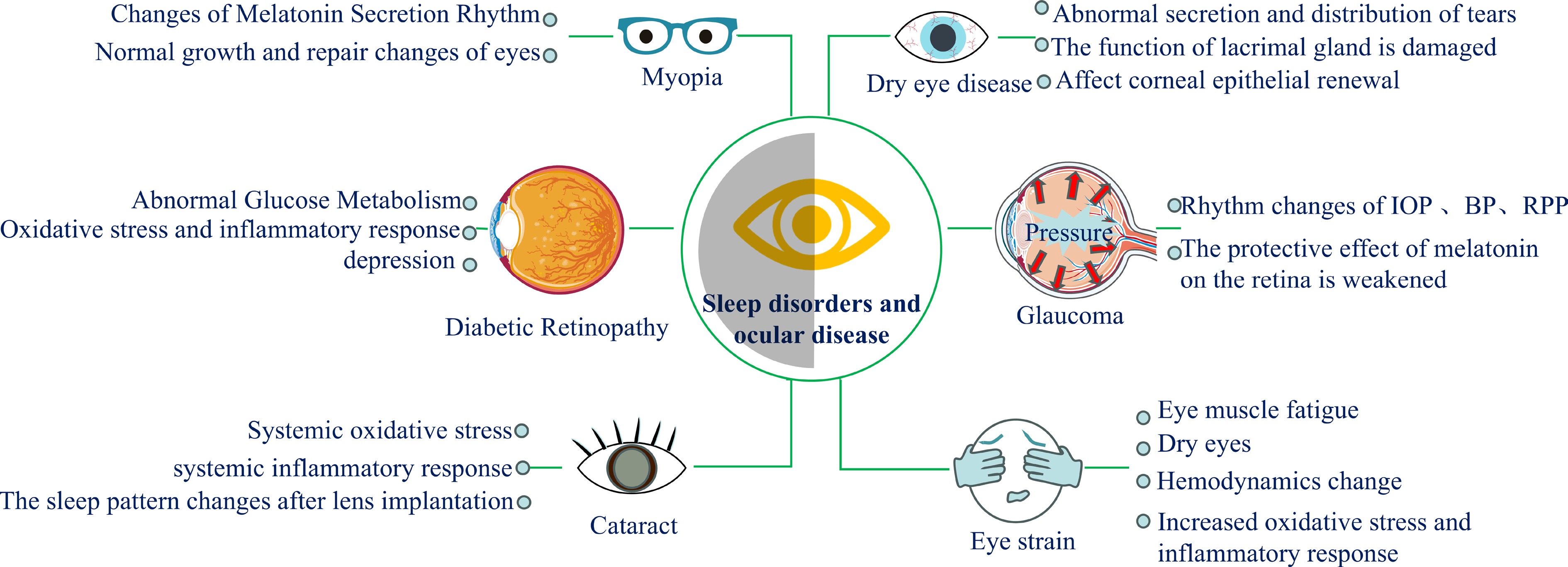
Figure 4.
Mind map diagram for sleep disorders and ocular disease. Selected artwork (the 'diabetic retinopathy' and 'glaucoma') depicted in the figure is derived from or adapted from images provided by Servier Medical Art (Servier;
https://smart.servier.com ), licensed under a Creative Commons Attribution 4.0 Unported License.Extensive research indicates a strong link between sleep disorders and ocular health, though the precise pathological mechanisms remain unclear. Disruptions in circadian rhythms likely play a significant role in these conditions. In addition to insufficient sleep duration and quality, sleep disorders such as OSA significantly impact ocular health. Further research into how sleep disorders influence eye disease risk, as well as the specific pathogenic mechanisms involved, is crucial for developing effective prevention and treatment strategies for ocular conditions.
Maintaining ocular health requires a comprehensive approach, which includes not only regular eye care practices but also the development of good sleep habits. Together, these practices can help mitigate the risks associated with sleep disorders and promote long-term ocular health.
-
This narrative review utilized publicly available literature and did not involve human or animal participants. Therefore, institutional review board approval was not required.
This work was supported by the Ministry of Science and Technology of the People's Republic of China (Grant No. 2018YFC0114500), and the National Natural Science Foundation of China (Grant Nos 82171014, 81470603, and 81770962, all awarded to Z.L.).
-
The authors confirm contribution to the paper as follows: study conception and overall study design, critical manuscript review and editing: Li Z; draft manuscript preparation, literature collection and comprehensive review : Xuan S. Both authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplymentary Table S1 Impact of Specific sleep abnormalities on cardiovascular health.
- Supplementary Table S2 Impact of specific sleep abnormalities on metabolic health.
- Supplementary Table S3 Impact of specific sleep abnormalities on neurological and mental health.
- Supplementary Table S4 Impact of specific sleep abnormalities on immune function.
- Supplementary Table S5 Key studies on sleep and ocular health: methodology and findings.
- Supplementary Table S6 Impact of specific sleep abnormalities on ocular health.
- Supplementary Table S7
- Supplementary Table S7 Effects of sleep disorders on the tear film and its components.
- Supplementary Table S8 Summary of key limitations in current evidence linking sleep and ocular health.
- Supplementary Fig. S1 Mind map diagram illustrating the mechanisms through which SD impacts eye strain. (Created by the authors).
- Supplementary Fig. S2 Possible mechanisms by which SD affects DED development. (Selected artwork (eye) depicted in the figure is derived from or adapted from images provided by Servier Medical Art (Servier; https://smart.servier.com/), licensed under a Creative Commons Attribution 4.0 Unported License).
- Supplementary Fig. S3 Possible mechanisms by which SD affects myopia development. (Created by the authors).
- Supplementary Fig. S4 Possible mechanisms by which SD affects glaucoma development. (Selected artwork (the "longitudinal section of the brain" and "glaucoma") depicted in the figure is derived from or adapted from images provided by Servier Medical Art (Servier; https://smart.servier.com/), licensed under a Creative Commons Attribution 4.0 Unported License).
- Supplementary Fig. S5 Mind map diagram illustrating the mechanisms through which SD impacts cataract development. (Created by the authors).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xuan S, Li Z. 2025. Linking sleep patterns to eye health: a review of current understanding and future directions. Visual Neuroscience 42: e010 doi: 10.48130/vns-0025-0008 

Linking sleep patterns to eye health: a review of current understanding and future directions
- Received: 07 December 2024
- Revised: 04 May 2025
- Accepted: 13 May 2025
- Published online: 27 June 2025
Abstract: Sleep plays a crucial role in maintaining ocular health. However, the relationship between sleep quality and eye diseases remains insufficiently explored. This narrative review consolidates findings from recent literature published over the past decade. It examines how sleep patterns influence the physiological stability of ocular tissues and their potential links to common eye conditions. Evidence from both animal and human studies suggests that sleep may influence retinal function, tear film homeostasis, intraocular pressure regulation, and inflammatory responses via circadian and neuroendocrine mechanisms. Particular attention is given to the associations between sleep disorders—such as obstructive sleep apnea—and ocular diseases including glaucoma, diabetic retinopathy, and dry eye syndrome. While existing research highlights the potential biological pathways, substantial heterogeneity in study designs precludes quantitative synthesis. Future interdisciplinary investigations are needed to clarify causal relationships and inform clinical strategies integrating sleep assessment into ophthalmic care.
-
Key words:
- Sleep quality /
- Ocular health /
- Sleep disorders /
- Eye diseases /
- Melatonin /
- Circadian rhythms /
- Eye strain /
- Dry eye syndrome /
- Myopia /
- Glaucoma /
- Cataracts /
- Diabetic retinopathy