









-
Chinese bayberry (Morella rubra Sieb. et Zucc), is an evergreen fruit tree native to southern China, belonging to the Myrica genus, a member of the Myriceae family[1]. The Chinese bayberry has a long history of cultivation, and is renowned for its distinctive flavor, vibrant coloration, and nutritional benefits conferred by abundant bioactive compounds including flavonols, phenolic acids, and anthocyanins[2,3]. The fruit of Chinese bayberry has a unique morphology. The outside of the seed exhibits many dense saccular structures radiating from the seed surface. During fruit development, these saccular bodies undergo progressive expansion and thickening, ultimately forming the edible sarcocarp[2]. However, during the expansion of these saccular bodies, they are susceptible to infection by Roucongbing disease that leads to abnormal fruit development in M. rubra. The main symptoms of the disease include excessive growth of fruit saccular bodies, which are separated from the fruit pit at the base and raised significantly. These separated and elevated saccular bodies swiftly wither and necrose, impeding normal fruit expansion and potentially leading to the cessation of fruit development. Currently, Roucongbing disease has been found in several cultivars of Chinese bayberry, such as Dongkui, Biqi, Ruansi, and Yingsi, with a higher incidence in Dongkui. The disease usually occurs in clusters and spreads quickly in orchards, severely incurring substantial economic losses. The unclear etiology and the mechanisms of response to Roucongbing disease in bayberry present a formidable challenge to its effective prevention and management. Consequently, there is an urgent to explore the effects and response mechanisms of the disease at the physiological and molecular levels to enhance our understanding and control of this pernicious condition.
Both biological and abiotic stresses affect plant growth and development. Plant responses to different stresses are highly complex and involve changes at the transcriptome, cellular, and physiological levels[4]. Under natural conditions, plants face the threat of infection by pathogens (including bacteria, fungi, and viruses) and attack by herbivore pests[5]. In addition to biotic stresses, extreme temperatures, drought, and nutrient limitations are major abiotic factors that reduce plant growth, productivity, and fruit quality[6]. Previous studies have confirmed the roles of the major plant hormones in regulating abiotic and biotic stress responses with a special focus on the significance of crosstalk between different hormones in generating a sophisticated and efficient stress response[7]. Some producers considered that Roucongbing disease was a physiological disease caused by nutritional deficiency or inconsistent growth due to hormonal imbalances, therefore, it was necessary to investigate through integrated transcriptomic and physiological analyses.
Nutrient deficiency, as one of the major abiotic stress factors, not only directly limits plant growth and fruit development[8,9] but also affects the ability to resist biotic stress[10−13]. Nitrogen (N) is an essential nutrient affecting many parameters in fruit productivity. N deficiency affects vegetative growth, flowering, and fruit quality[14], also N negatively affects the plant defense, N addition generally reduces the amount of defensive compounds and affects plant disease resistance[13]. Potassium (K) regulates multiple plant stress responses, including in concert with jasmonic acid (JA), ethylene (ET), and auxins[15]. Various contemporary studies revealed that K+-homeostasis plays a substantial role in plant responses and tolerance to abiotic stresses[16]. K-deficient plants tend to be more susceptible to infection than those with an adequate supply of K[17]. In addition to being a vital component of cell structure, calcium (Ca) also functions as the most prominent second messenger in the cascade reaction of plant defense signals[18,19]. Calcium deficiency in apple fruits causes bitter pit[20], and exogenous Ca treatment of papaya can improve the resistance of anthracnose[21]. In recent years, increasing research is focusing on the function of manganese (Mn) in plant defense against pests and pathogens, especially in woody plants. As a necessary micronutrient for enzymatic reactions and defense, manganese is involved in the creation of lignin and phenolic compounds, which help plants resistance to disease[13,22].
Plant hormones play an important role in the regulation of fruit development and ripening, and they also emerged as signaling compounds with key functions in responses to disease resistance[23]. The major hormones produced by plants are auxins, gibberellins (GA), cytokinins (CK), abscisic acid (ABA), ethylene (ET), salicylic acid (SA), and jasmonates (JA). Among them, auxins and GAs regulate the progress of cell division and cell expansion in fruit development. ABA, SA, JA, and ET are known to play major roles in mediating plant defense response against pathogens and abiotic stresses. SA signaling positively triggers plant immunity against biotrophic pathogens, while the JA pathway is mostly required for resistance against necrotrophic pathogens[24,25]. Jasmonic acids (JAs) are important injury signaling molecules and play an important role in regulating plant growth and defense responses, which participate in the process of wound healing in apple fruit[26,27]. Various enzymes are involved in JA synthesis, such as allene oxide synthase (AOS), allene oxide cyclase (AOC), and oxo-phytodienoic acid reductase (OPR3). The Arabidopsis mutants of JA biosynthesis genes (such as aos and opr3) are significantly more susceptible to insects when compared with wild-type plants[28]. As a plant defense hormone required for immunity, exogenous salicylic acid (SA) treatment can improve the resistance of fruits[29,30], however, there was reciprocal antagonism between SA and JA signals in shaping the outcome of plant-pathogen interactions[31,32].
To study the cause and response mechanism of Roucongbing disease in bayberry fruits, the overall nutritional level of diseased and healthy trees and endogenous hormones in healthy and diseased fruits from the same tree were detected quantitatively. Through integrated transcriptomic and targeted hormonal analyses, this study reveals multilevel response mechanisms to Roucongbing disease, advancing our understanding of this economically significant pathology. Overall, this study advances our knowledge of Roucongbing disease, which provides a theoretical foundation for its prevention and control, and the future breeding of resistant cultivars.
-
Fifteen-year-old 'Dongkui' (M. rubra) were used in this study, which were all cultivated in the same orchard in Taizhou City, Zhejiang Province, China. This region experiences a subtropical monsoon climate with slightly acidic red soil. Three disease-free, healthy trees were chosen as control trees, while three trees with severe Roucongbing disease symptoms were designated as diseased trees. All the trees were in the same facility to ensure consistency in environment and agricultural management. Among them, the trees with more than 20% of diseased fruit were selected as diseased trees, while those with no diseased fruit were selected as control trees. In all four directions (east, west, south, and north) of the control trees, the fruits and surrounding branches containing stems and leaves comprised the control group (CK). The diseased fruits and surrounding branches in diseased trees were described as the diseased group (D), and the healthy group (H) was made up of no diseased fruits and surrounding branches from the same tree. A portion of fruits, leaves, and stems from each treatment were collected and used for element content determination, and the other parts were immediately frozen in liquid nitrogen after being collected and stored in a −80 °C ultra-low temperature refrigerator for future use. To investigate overall nutrient levels, control trees (CK), and diseased trees (D and H) were systematically compared for differential nutrient profiling. Concurrently, a focused analysis was conducted on endogenous hormone quantification and transcriptome sequence analysis between diesease (D) and healthy (H) fruits within the same trees, with control for genetic and environmental variability.
Element analysis
-
The leaves and fruits were heated at 105 °C for 30 min and then dried at 65 °C for 3 d after cleaning with ultra-pure water. The contents of potassium (K), phosphorus (P), magnesium (Mg), calcium (Ca), iron (Fe), manganese (Mn), zinc (Zn), and boron (B) contents were quantified by inductively coupled plasma-optical emission spectrometry (ICP-OES iCAP series 7000 Plus Thermo Scientific) and slightly improved[33,34]. This method included: weighing 1.0 g of the samples into the microwave digestion inner tank, adding 5 mL of nitric acid (65%), and 2 mL of hydrogen peroxide (30%), placed for a night and digested at 180 °C until complete resolution. After the digestion was completed, the digestive solution was then transferred with deionized water to a constant volume of 50 mL, mixed well for testing, and then the sample blank test was conducted. The nitrogen (N) content used sulfuric acid for digestion and determined using the total Kjeldahl digestion method[35].
Hormonal analysis
-
The quantification of different endogenous hormones was performed by Beijing Biomarker Biotechnology Co., Ltd. (Beijing, China) using an LC-MS/MS platform[36]. The procedure involved freeze-drying and grinding the plant fresh samples into powder, followed by extraction using methanol. Following centrifugation and filtration, the samples were examined using an AB Sciex 6500 Q TRAP triple quadrupole linear ion trap mass spectrometer. Based on the local metabolite database, qualitative and quantitative analyses of metabolites in the samples were performed. Characteristic ions for each substance were screened using a triple quadrupole filter, and the signal intensity of characteristic ions was obtained from the detector. The chromatographic peaks were integrated and calibrated using MultiaQuant software, where the relative quantity of each drug was represented by the peak area (Area) of that particular chromatographic peak. All the data from the chromatographic peak integration were then exported and saved.
Transcriptome sequencing
-
Transcriptomics was performed by Beijing Biomarker Biotechnology Co., Ltd. (Beijing, China). Total RNA was extracted from fruit pulp using the CTAB method, and Oligo (dT) magnetic beads were employed to enrich mRNA. After being reverse-transcribed into single-stranded cDNA, the enriched fragments were synthesized into double-stranded cDNA. After purification and blunt-end modification, sequencing was performed using the Illumina HiSeq X Ten platform. Trinity software (version 2.4.0) was used for assembly and identification. The longest sequence for each gene was selected based on similarity and length. CD-HIT software (version 4.6) was used to cluster the sequences and eliminate redundancy.
RNA-seq analysis
-
Raw data were filtered to remove low-quality data, eliminate adapter sequences, and obtain high-quality clean data for analysis. The clean data was aligned to the reference sequence, the Bayberry T2T genome, using the Hisat2 program[37,38]. Fragments Per Kilobase of transcript per Million mapped reads (FPKM), which is equal to mapped cDNA fragments (million) × transcript length (kb), were used to quantify the levels of gene expression. Using the DESeq program, differential expression analysis was carried out, and genes with FPKM values greater than 1 were classified as DEGs. Gene Ontology (GO) enrichment analysis was performed using the GOseq R package based on the Wallenius non-central hyper-geometric distribution. KEGG pathway enrichment analysis was conducted using KOBAS 3.0. The heatmap of DEGs and the correlation network diagram of transcriptome and targeted hormones were generated using the R heatmap package on the BMK Cloud platform (
www.biocloud.net ).RNA isolation and qRT-PCR
-
Total RNA from fruits was extracted using the Trizol method. cDNA was synthesized in a 10 μL reaction using the HiScript III RT SuperMix with gDNA wiper (Vazyme, China). The qRT-PCR reaction was performed on a LightCycler 96 real-time PCR instrument (Roche, China) referring to previous research methods[37]. Each sample was analyzed with three technical replicates. Quantitative primers were designed based on gene sequences, with MrACT used as the housekeeping gene. Data were processed using the 2−ΔΔCᴛ method. All primers were listed in Supplementary Table S1.
-
Roucongbing disease in M. rubra often occurs in large areas, especially abundant in the lower and middle strata of the canopy, with a higher incidence in humid and warm environments, particularly during the early stage of fruit development (fruit enlargement period) (Fig. 1a). The early symptoms of this disease are abnormal growth and clustered protrusions of saccular bodies (Fig. 1b). With fruit growth and development, the saccular bodies become detached from the fruit core and gradually wither into a brownish-yellow color (Fig. 1c), either remaining attached to the fruit surface or falling off. Microscopic examination of diseased and healthy fruits revealed distinct differences: the diseased fruits exhibited a blackened seed epidermis and a dark brown base of the pulp, which was absent in the healthy controls (Fig. 1d, e). The diseased fruits exhibited morphological deformities severely, ultimately failing to form normal mature fruits. Fruits with milder early symptoms also tend to show uneven surface textures at the mature stage, significantly affecting their commercial value.

Figure 1.
The phenotype of Roucongbing disease in bayberry fruit. (a) The disease occurs in large areas, (b) morphology of diseased fruits, and (c) healthy fruits, the anatomical observation of (d) diseased fruit, and (e) control fruit.
The imbalance of nutrient levels can lead to plant physiological diseases and also affect the resistance to plant disease. To examine the hypothesis that this disease is associated with element deficiency, the whole nutrient levels of the control tree and diseased tree were compared. The mineral element contents of each group (CK, H, and D) fruits, as well as the surrounding leaves, were quantified (Table 1). Our results showed that the magnesium (Mg), potassium (K), manganese (Mn), and zinc (Zn) concentrations in the leaves of the diseased trees changed overall when compared to the control trees. The K content of diseased leaves decreased but that of fruits increased significantly, which suggested that more K was transferred from the leaves to the fruit when diseased. The contents of Mg and Zn in diseased leaves were significantly higher than those in the control and healthy groups. Interestingly, our data revealed that the concentration of Mn, a pivotal element in plant defense mechanisms, was substantially reduced in both the leaves and fruits of diseased trees compared to the CK group. In healthy and diseased fruits from the same tree, there were no significant differences in other elements except for a change in N content, which implied that Roucongbing disease might not be directly caused by nutritional deficiencies.
Table 1. Mineral element contents of leaves and fruits in different groups.
Leave Fruit CK H D CK H D N (g/kg) 13.51 ± 0.39 b 14.75 ± 0.38 a 13.37 ± 0.34 b 18.89 ± 0.03 c 21.04 ± 0.20 b 22.49 ± 0.07 a K (g/kg) 5.84 ± 0.10 a 5.28 ± 0.11 c 5.57 ± 0.02 b 10.12 ± 0.17 b 12.35 ± 0.43 a 13.56 ± 0.69 a Ca (g/kg) 9.74 ± 0.31 b 10.47 ± 0.36 b 14.13 ± 0.34 a 2.33 ± 0.03 b 2.68 ± 0.09 a 2.76 ± 0.12 a P (g/kg) 0.44 ± 0.01 b 0.40 ± 0.01 c 0.47 ± 0.02 a 1.09 ± 0.04 b 1.30 ± 0.06 a 1.33 ± 0.03 a Mg (g/kg) 1.15 ± 0.05 c 1.27 ± 0.03 b 1.54 ± 0.04 a 0.98 ± 0.04 b 1.13 ± 0.09 a 1.15 ± 0.08 a Mn (mg/kg) 1,256.21 ± 14.70 a 495.29 ± 5.18 b 488.18 ± 7.87 b 197.15 ± 5.63 a 74.75 ± 2.36 b 66.72 ± 0.15 b Fe (mg/kg) 253.88 ± 10.41 a 276.57 ± 12.43 a 252.80 ± 10.04 a 432.43 ± 18.17 a 392.91 ± 21.62 a 400.75 ± 18.47 a Zn (mg/kg) 60.31 ± 0.73 c 88.79 ± 1.40 a 77.30 ± 2.25 b 35.35 ± 0.67 a 35.31 ± 0.93 a 36.63 ± 0.34 a B (mg/kg) 38.92 ± 3.08 a 32.77 ± 1.90 a 35.99 ± 4.15 a 0.75 ± 0.13 a 0.72 ± 0.13 a 0.66 ± 0.09 a Data in the table are expressed as mean ± SE. CK: the leaves and fruits from the trees with no diseased fruits; D: the diseased fruits and surrounding leaves in diseased trees (with more than 20% of diseased fruit); H: the healthy fruits and surrounding leaves from the same trees as D group. Different letters indicate statistically significant differences in one-way ANOVA (p < 0.05). Plant endogenous hormones associated with Roucongbing disease
-
Plant hormones are the main signal substances that regulate fruit growth, and development and respond to stress. To explore the main hormone signaling pathways in response to Roucongbing disease, the endogenous hormones in bayberry fruit were quantified and revealed a total of 13 endogenous hormones (Supplementary Table S2), most of which were significantly different between healthy and diseased fruits (Fig. 2a). Our results showed that gibberellin A3 (GA3) (Fig. 2g), abscisic acid (ABA) (Fig. 2f), jasmonic acid (JA) (Fig. 2e), 1-Aminocyclopropanecarboxylic acid (ACC) (Fig. 2b), salicylic acid (SA) (Fig. 2d), and indole-3-acetic acid (IAA) (Fig. 2c) are several major hormones with high content in bayberry fruits. GA3 is the main hormone in the expanded stage of fruit, in our results, the content of GA3 was the most abundant in bayberry fruits. In addition to GA3, the contents of ABA, JA, and IAA were also significantly increased in diseased fruits, among them, JA was the most increased hormone and the content in diseased fruits was about seven times that of healthy fruits. On the contrary, the ethylene synthesis precursor ACC was significantly decreased in diseased fruits. While SA, as the main hormone in plant response to disease, had no significant difference between healthy and diseased fruits.

Figure 2.
Hormone content analysis of healthy and diseased fruits. (a) Heatmap and cluster analysis of all endogenous hormones, and the quantitative comparison of (b) 1-aminocyclopropanecarboxylic acid, (c) indole-3-acetic acid, (d) salicylic acid, (e) jasmonic acid, (f) abscisic acid, and (g) gibberellin A3 were analyzed in healthy and diseased fruits. Error bars indicate standard error from three biological replicates (n = 3). Different lowercase letters indicate significant differences (Student's t-test; p < 0.05).
Analysis of DEGs
-
To elucidate the response mechanism in bayberry fruit, healthy and diseased fruit pulp tissue samples from the same tree underwent transcriptome sequencing by the Illumina NovaSeq 6000 high-throughput sequencing platform. The data are as follows in Table 2: 124.67 million reads were included in the clean data that was produced following filtering. The Q30 base ratio exceeded 93.46%, whereas the GC base ratio reached 47.96%−48.64%, suggesting a comparatively high sequencing quality of the samples that complied with bioinformatics standards.
Table 2. Statistical analysis of transcriptome sequencing data.
#Sample ID ReadSum BaseSum GC (%) N (%) Q20 (%) Q30 (%) H-1 20031820 5981473950 48.12 0 97.92 94.57 H-2 20965481 6269407182 47.99 0 98.04 94.84 H-3 20704915 6179849928 47.96 0 97.87 94.45 D-1 20425378 6102338718 48.64 0 97.29 93.46 D-2 21524247 6421989748 48.15 0 97.66 94.19 D-3 21018045 6268115810 48.17 0 97.59 94 To confirm the relationship between all samples, correlation, and principal component analysis (PCA) analysis were performed. As shown in Fig. 3, the R2 values among the three replicates of the same treatment were all larger than 0.942, while the correlation among the two groups was significantly different (Fig. 3a). For the PCA analysis, three replicates of the same treatment clustered together, indicating high reliability among the samples (Fig. 3b). The samples from the different groups were distant from each other, indicating significant differences in transcriptome samples between different treatments, with PC1 and PC2 contributing rates of 52.99% and 16.31%, respectively. Gene expression in different samples was assessed using the FPKM method, and differential analysis was carried out. Transcriptomic profiling revealed 16,155 genes co-expressed across H and D groups (Fig. 3e), and there were 2,582 genes expressed differentially (2,582 DEGs) (Fig. 3c), with 1,603 genes upregulated and 979 down-regulated in the D group (Fig. 3d).

Figure 3.
Transcriptome analysis of healthy and diseased fruits. (a) Heatmap of Pearson correlation analysis. (b) Principal component analysis of all samples. (c) Heatmap clustering, and (d) volcano plot analysis of all differentially expressed genes. (e) Venn diagrams of all genes.
GO and KEGG pathway analysis
-
All of the annotated genes were used as background and GO enrichment analysis was performed to characterize the biological functions of DEGs and further classified into three categories: the cellular component, biological process, as well as molecular function process (Fig. 4a). The results showed that in the biological process (BP), the DEGs between H vs D were mainly involved in the cellular process, metabolic process, and biological regulatory activities. In the cellular component (CC), the DEGs were mainly linked to the cellular anatomical entity, intracellular, and protein-containing complex. The results showed that in the molecular function (MF), the DEGs were mainly involved in the catalytic activity, binding, and transporter activity.
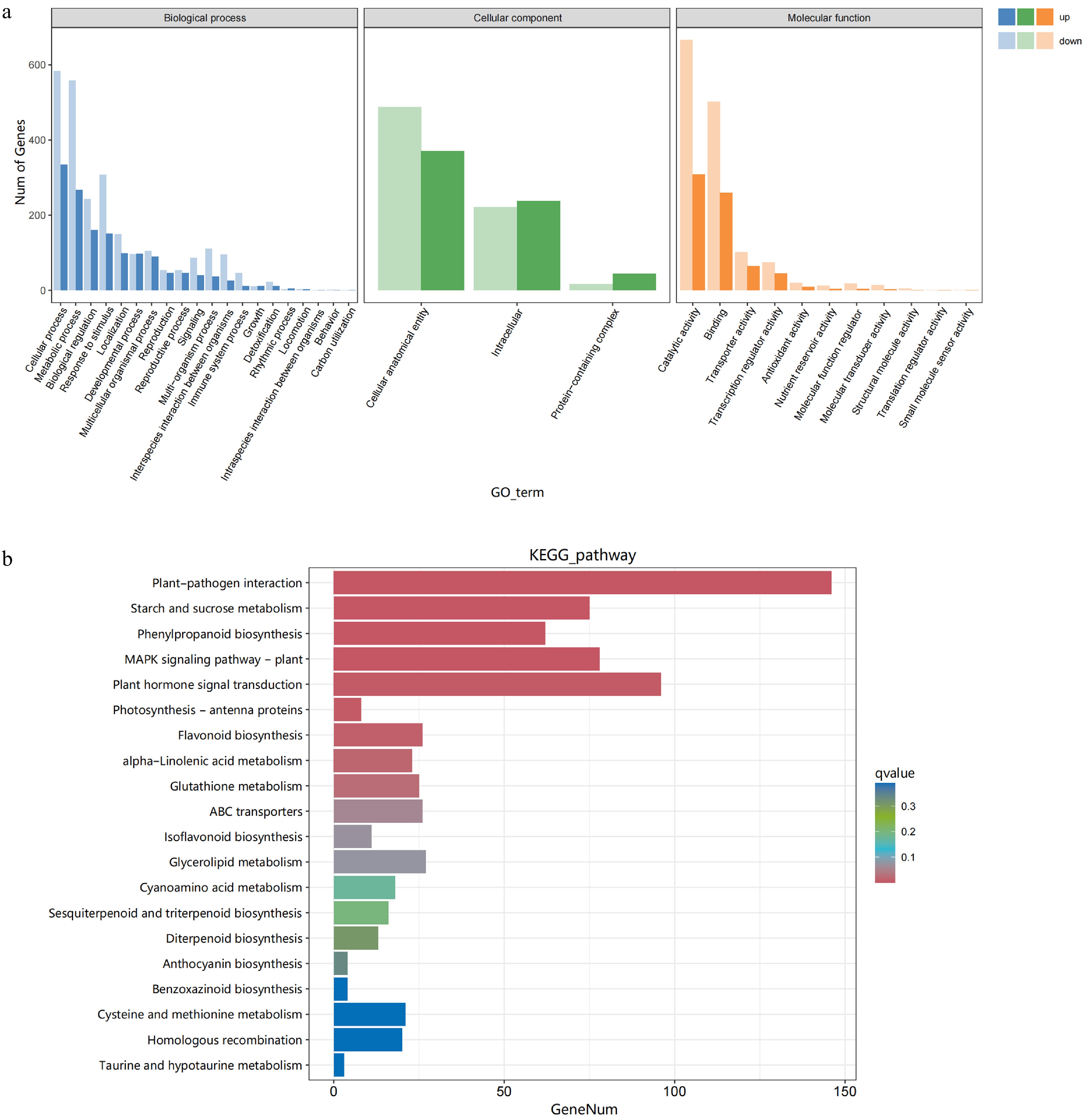
Figure 4.
GO and KEGG analysis. (a) GO enrichment analysis, and (b) the top 20 KEGG pathways of DEGs were analyzed in healthy and diseased fruits.
KEGG enrichment analysis of metabolic pathways was performed for DEGs in each group. The DEGs between healthy and diseased fruits were assigned to 126 pathways (Fig. 4b). Notably, the plant-pathogen interaction pathway was the most abundant among these pathways, containing 146 DEGs in total (134 up-regulated and 12 down-regulated). Furthermore, the pathways of plant hormone signal transduction, the MAPK signaling pathway in plants, starch, and sucrose metabolism, and phenylpropanoid biosynthesis were also prominently enriched, with 96, 78, 75, and 63 DEGs, respectively. These pathways are recognized as pivotal in the plant's response to disease, indicating fruits of bayberry are involved in the defense against pathogens.
Analysis of DEGs in major pathways
-
The plant-pathogen interaction pathway is the major pathway for the enriching of DEGs between diseased and healthy bayberry fruits. Therefore, we conducted further analysis of this pathway (Fig. 5a), among them, there were a total of 20 transcription factors. Two of them belonged to the Pti transcription factor family, four belonged to the bHLH transcription factor family, and the WRKY transcription factor family had the largest number, with 13 DEGs. Importantly, all WRKY transcription factors exhibit significantly upregulated expression levels in diseased fruits. In the plant-pathogen interaction, the DEGs also include 20 Receptor-Like Kinases (RLKs). Except for two downregulated RLKs, the expression levels of other MrRLK genes were significantly upregulated in diseased bayberry fruits. Pathogenesis-related proteins (PRs), TIR domains, and NB-ARC domains play crucial roles in plant disease resistance. Among these 146 DEGs, there were five DEGs encoding PR proteins, four DEGs encoding TIR domain proteins, and seven DEGs encoding NB-ARC domain proteins (Fig. 5b). The expression levels of these genes in diseased fruits were significantly higher than those in healthy fruits of the same plant.
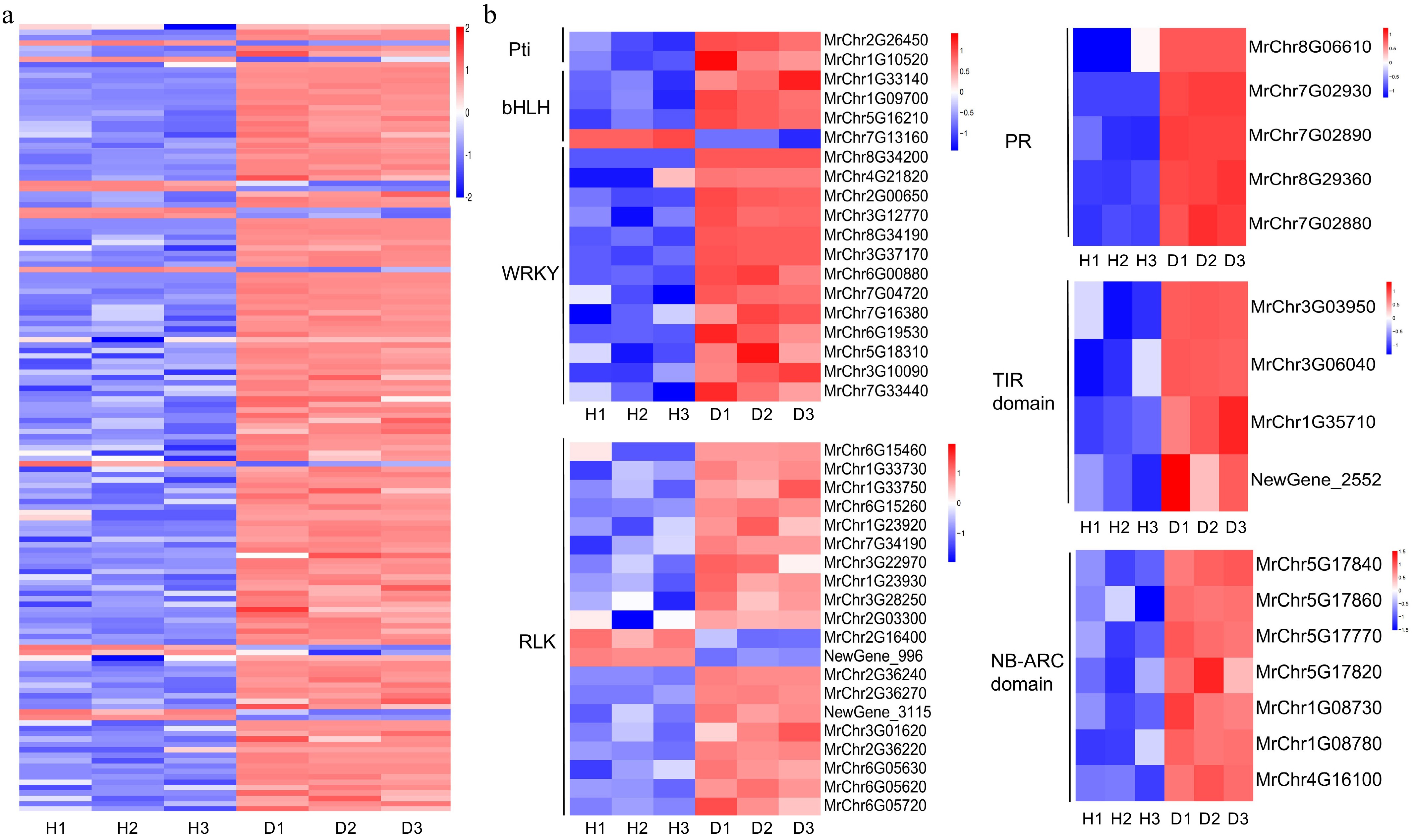
Figure 5.
Heatmap analysis of plant-pathogen interaction pathway. (a) Heatmap of 146 DEGs of the plant-pathogen interaction pathway. (b) Gene expression analysis of some transcription factors families and disease resistance related genes in this pathway.
Conjoint analysis of transcriptome and targeted hormones
-
By conjoint analysis of transcriptome and major hormones, 124 DEGs in the plant-pathogen interaction pathway were identified that correlated with the six hormones, JA was in the center role of the correlation network diagram, which suggested that JA played a key role in Roucongbing disease resistance (Fig. 6a). Next, we focused on the genes related to JA synthesis in the transcriptome. We found that the expression levels of all genes in the JA synthesis pathway, including MrLOX, MrAOS, MrAOC, MrOPR3, and MrJMT were all significantly increased in the diseased fruits (Fig. 6b). These results were consistent with the determination of JA hormone content, indicating that the JA pathway played an important role in disease defense.
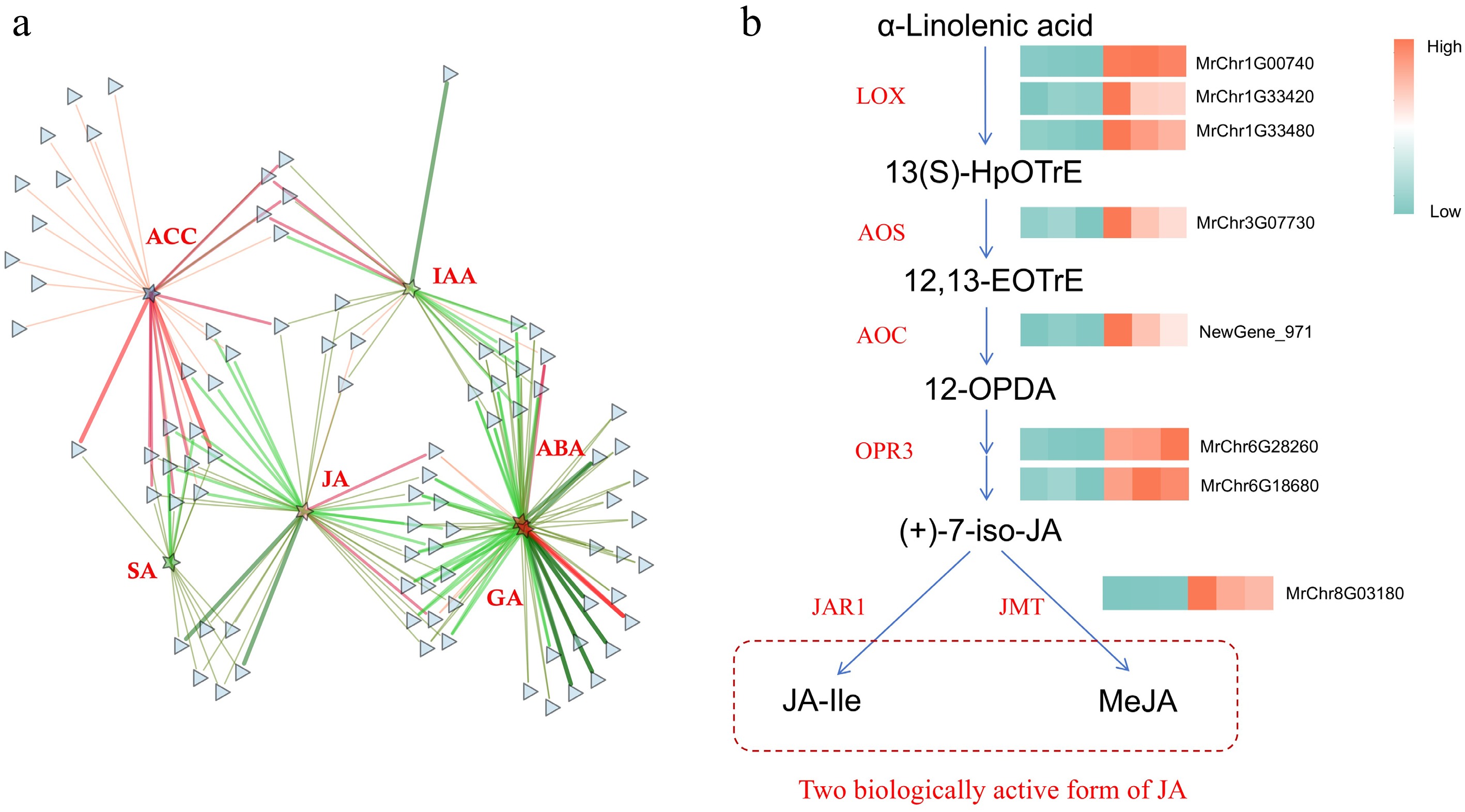
Figure 6.
The role of JA in Roucongbing disease resistance. (a) Correlation network diagram of the plant-pathogen interaction pathway and six major hormones. (b) Transcriptome analysis of the expression patterns of JA synthesis related genes. Green to red color indicates the gene expression increasing in abundance.
Validation of sequencing data by quantitative PCR
-
To validate the RNA-Seq data, we conducted qRT-PCR analysis. We selected four hormone related genes, such as the JA synthesis gene MrAOS (MrChr3G07730) (Fig. 7a), MrOPR3 (MrChr6G28260) (Fig. 7b), MrLOX2.1 (MrChr1G00740) (Fig. 7c), the ABA synthesis related gene MrNCED2 (MrChr2G18590) (Fig. 7d), and several genes in the plant-pathogen interaction pathway, including WRKY70 (MrChr2G34190) (Fig. 7e), WRKY18 (MrChr8G34190) (Fig. 7f), MrPTI6-like (MrChr1G10520) (Fig. 7g), and MrPR1 (MrChr7G02890) (Fig. 7h). All the qRT-PCR results were consistent with the transcriptome, providing robust support for our transcriptomic observations.
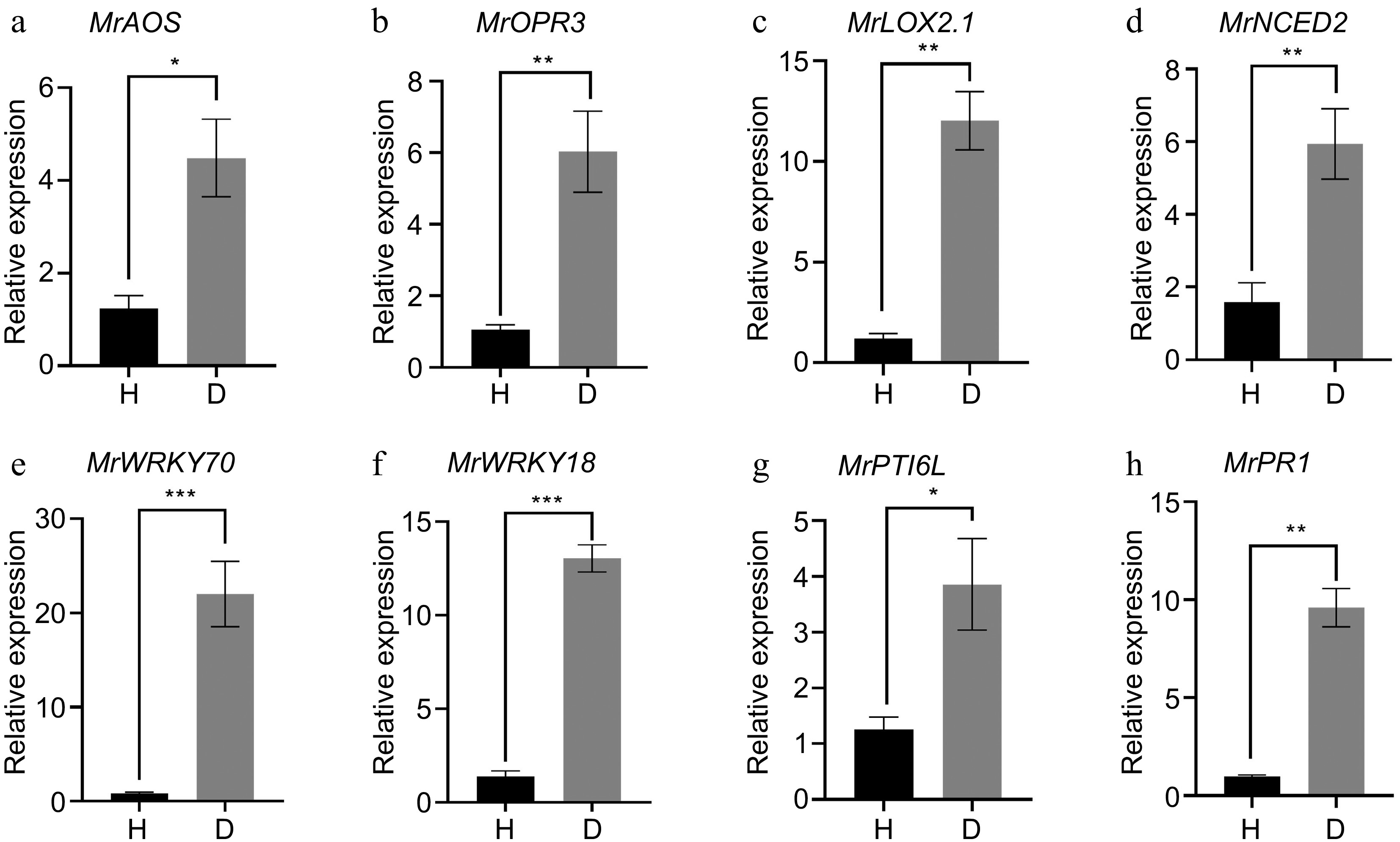
Figure 7.
qRT-PCR analysis of DEGs identified from RNA-Seq. (a) MrAOS: allene oxide synthase 1; (b) MrOPR3: 12-oxophytodienoate reductase 3; (c) MrLOX2.1: Linoleate 13S-lipoxygenase 2-1; (d) MrNCED2: 9-cis-epoxycarotenoid dioxygenase 2; (e) MrWRKY70: transcripdion factor WRKY70; (f) MrWRKY18: transcripdion factor WRKY18; (g) MrPTI6L: pathogenesis-related genes transcriptional activator PTI6-like; (h) MrPR1: Pathogenesis-related protein PR1. Error bars indicate standard error from three biological replicates (n = 3). An asterisk indicates statistically significant differences compared with the control (Student's t-test; * p < 0.05, ** p < 0.01, *** p < 0.001).
-
At present, there are many speculations on the causes of Roucongbing disease, and no pathogenic microorganism has yet been isolated. In this study, a comprehensive analysis was conducted at the physiological and transcriptome levels. The results of nutrient element content in both diseased and healthy fruits within the same tree did not show significant differences, which implied the nutritional level of the same tree remained consistent. However, the nutritional state of diseased and control trees differed, such as the content of N, Ca, and Mg were increased and the Mn content decreased in the diseased trees. The plants with high N content were more susceptible to disease[13], and our results were consistent with this. Interestingly, Mn was shown to be the most significant difference in content. Our results showed that the leaves and fruits in diseased trees had lower concentrations of Mn than those of the control trees significantly, however, no Mn deficiency phenotype was observed in the diseased trees. Mn is an antioxidant that might lessen oxidative damage[39]. Additionally, Mn plays a key role in plant defense. The immune system is typically triggered by pathogen infection, which causes a significant build-up of reactive oxygen species (ROS), and significant damage to plant cells. Under abiotic stress, ROS also accumulates in large quantities in plants. Mn is a cofactor of Mn-SOD that is present in peroxisomes and mitochondria and is involved in lignin synthesis in plants. Previous studies have shown that exogenous Mn treatment for leaves can lessen pathogen-induced lignification and ROS generation, hence improving plant resistance.[40]. In our study, the significant decrease in Mn content in diseased bayberry trees suggests a potential role of Mn in the defense against Roucongbing disease and a higher Mn content might contribute to strengthen the resistance in M. rubra, providing a theoretical basis for improving disease resistance through cultivation techniques.
Hormonal signaling controls the conflict between defense and development in plants. In our research, hormone levels in healthy and diseased fruits differed significantly. In diseased fruits, there was a considerable rise in JA, ABA, IAA, and GA3, among them, GA3 was the most abundant hormone and JA was the most significantly increased within them. Previous studies have revealed that the JA pathway plays an essential role in plant defense responses against necrotrophic and hemibiotrophic pathogens[41]. Tissue injury or local pathogen infection causes a rise in the concentration of JA[42,43]. In strawberry, terpinen-4-ol induced the expression of genes in jasmonic acid (JA) biosynthesis (FaLOX, FaAOC, and FaOPR3) and increased JA content to enhance disease resistance, which demonstrates the effect of JA in the induction of disease resistance[44]. In our study, it was observed that Roucongbing disease promoted a significant accumulation of JA locally. The combined analysis of transcriptome and targeted hormones showed that JA played a central role in response to Roucongbing disease and the key genes in the JA synthesis pathway were significantly upregulated in diseased fruits, consistent with the observed trends in hormonal changes, which showed a potentially important role for JA in defense against this disease. It has been confirmed that exogenous GA3 treatment can promote early enlargement of grape berries[45], indicating that GA plays an important role in fruit expansion. In the results of this study, the high content of GA was consistent with the fact that the fruit deformity mainly occurs in the fruit expansion stage, which was significantly increased in diseased fruit with the disease symptoms of fruit overexpansion, this suggests that the accumulation of GA3 might be the cause of abnormal enlargement in diseased fruits. The primary plant defense hormone, SA, did not significantly alter between unhealthy and diseased fruits, indicating that it may not be directly involved in defending against Roucongbing disease in M. rubra fruits.
At the transcriptional level, several transcription factors exhibited differential expression. It has been consistently demonstrated that WRKY transcription factors are essential for plants to respond to biotic and abiotic stressors[46]. In this study, it was noted that the WRKY family contained the most differentially expressed transcription factors in the plant-pathogen interaction pathway. Analysis of all DEGs in healthy and diseased groups showed that all differentially expressed WRKY members (22 in total) were significantly upregulated in diseased fruits (Fig. 2). Pathogens induce WRKY70 expression in Arabidopsis[47], and overexpression of WRKY70 strengthens plant resistance in poplars[48,49]. PbWRKY18 expression was induced by pathogens and MeJA, and overexpression of PbWRKY18 positively regulated the expression of PbCHS3 to enhance pear resistance to pathogens[50]. WRKY40 directly binds to the chitinase ChilV3 promoter in peppers to improve defense against pepper bacterial wilt[51]. The most highly activated transcription factors in Roucongbing diseased fruits were MrWRKY70, MrWRKY40, and MrWRKY18, indicating their critical roles in the resistance to Roucongbing disease. Several genes linked to the disease were shown to be highly elevated in infected fruits, in addition to transcription factors. In addition to WRKY transcription factors, the MrPti gene also increased significantly. In Arabidopsis, Pti4/5/6 could activate the expression of the AtPR1 and AtPR2[52], and in our results, a member of the Pti family, the MrPTI6-like gene was increased significantly in diseased fruit. In plant defense, the TIR domain and NB-ARC, have less substantial expression changes than pathogenesis-related genes (PRs), especially those in the PR1 protein family, however, whether it was activated by MrPti6-like remains to be further investigated. PR proteins play a critical role among the accumulated resistance genes, as evidenced by the increased expression levels of these genes in sick fruits. Subsequent investigation reveals that the majority of the differentially expressed MrPR genes found in this study are members of the PR1 family. Previous studies showed that PR1 basic protein has antifungal activity in tobacco and tomato[53]. According to our study, MrPR1 plays an important role in the defense mechanism of M. rubra fruit. Consequently, the differentially expressed MrWRKY70, MrWRKY18, and MrPR1 genes identified in this study can serve as candidate genes for improving resistance to Roucongbing disease in M. rubra and for the breeding of new varieties.
-
This study showed that there were alterations in the nutrient levels of leaves and fruits in diseased trees compared with the control trees, the Mn content in diseased trees decreased significantly. Meanwhile, there was a noticeable increase in JA content in diseased fruits, along with a significant elevation in the expression levels of JA synthesis-related genes. Furthermore, the transcriptome analysis results revealed that the expression of disease-resistance-related genes such as WRKY, Pti, and PR were significantly increased in diseased fruits. The findings enrich our comprehension of Roucongbing disease, which provides a scientific basis for the resistance strategy and breeding of resistant varieties.
The present work was supported by grants from the Natural Science Foundation of China (32402531), the special breeding program for new varieties in Zhejiang (2021C02066-2), the Agricultural Project of Huangyan district, Taizhou City, Zhejiang Province (JBGS2022-HYNY01), and the 'Lingyan' R&D in Zhejiang (2023C02031).
-
The authors confirm their contribution to the paper as follows: study conception and design: Sun L; analysis and interpretation of results: Qi Y, Zhang Y; data collection: Zheng X, Yu Z; funding management: Qi X, Zhang S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 List of primers used for qRT-PCR analysis.
- Supplementary Table S2 Endogenous hormone contents of fruits in different samples.
- Copyright: © 2025 by the author(s). Published byMaximum Academic Press, Fayetteville, GA. This articleis an open access article distributed under Creative CommonsAttribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun L, Qi Y, Zhang Y, Yu Z, Zheng X, et al. 2025. Physiological and transcriptome analysis of Roucongbing disease in Chinese bayberry (Morella rubra Sieb. et Zucc). Fruit Research 5: e027 doi: 10.48130/frures-0025-0019 

Physiological and transcriptome analysis of Roucongbing disease in Chinese bayberry (Morella rubra Sieb. et Zucc)
- Received: 08 January 2025
- Revised: 30 March 2025
- Accepted: 01 April 2025
- Published online: 08 July 2025
Abstract: Chinese bayberry (Morella rubra Sieb. et Zucc) is a characteristic fruit of China with high nutritional value. The frequent occurrence of Roucongbing disease in bayberry fruits has been a major problem affecting production and marketing in recent years. Unfortunately, the causes and the response mechanisms of this disease are still unclear, which makes it extremely difficult to prevent and treat. In this study, the overall nutrient content between diseased trees and control (non-diseased) trees were compared, which suggested that Mn might be associated with resistance to Roucongbing disease. A comparative analysis of the endogenous hormone profiles between healthy and diseased fruits from the same tree revealed significant differences, with jasmonic acid (JA), gibberellin (GA), abscisic acid (ABA), and auxin (IAA) exhibiting pronounced increments in the diseased fruits. The transcriptome analysis showed that the differentially expressed genes (DEGs) were mainly enriched in the plant-pathogen interaction pathway, which included multiple genes encoding some WRKY transcription factor family members, and the pathogenesis-related protein PR1. Our results indicated a strong correlation between the DEGs of these pathways and JA signaling, implying that the upregulation of genes involved in JA biosynthesis might serve as a pivotal defensive response against Roucongbing disease in bayberry fruits. Furthermore, the expression patterns of select DEGs were validated by quantitative Real-time PCR (qRT-PCR). This study explored the response mechanism at physiological and molecular levels, providing a theoretical foundation for the prevention and control of Roucongbing disease and the breeding of resistant cultivars.
-
Key words:
- Chinese bayberry /
- Roucongbing disease /
- Physiological /
- Transcriptome /
- Response mechanism