








-
Ethylene, with the chemical formula C2H4, is the smallest unsaturated alkene and also the simplest known plant hormone (Fig. 1a). In agricultural applications, ethylene is widely used as a ripening agent for fruits and vegetables[1]. It is among the earliest identified endogenous plant hormones and is considered one of the five classic plant hormones. In higher plants, ethylene is produced in various tissues, including roots, stems, leaves, flowers, fruits, tubers, and seeds[2]. Throughout the entire plant life cycle—covering seed germination, leaf expansion, root hair elongation, lateral root growth, flowering, fruit ripening, leaf abscission, and senescence—ethylene plays a vital regulatory role[3]. Moreover, it is crucial for enabling plants to cope with mechanical, biotic, and abiotic stresses[1]. This review focuses on the basic mechanisms of ethylene signaling and its integration with other pathways, aiming to provide a deeper understanding of the complex functions and potential applications of this plant hormone.
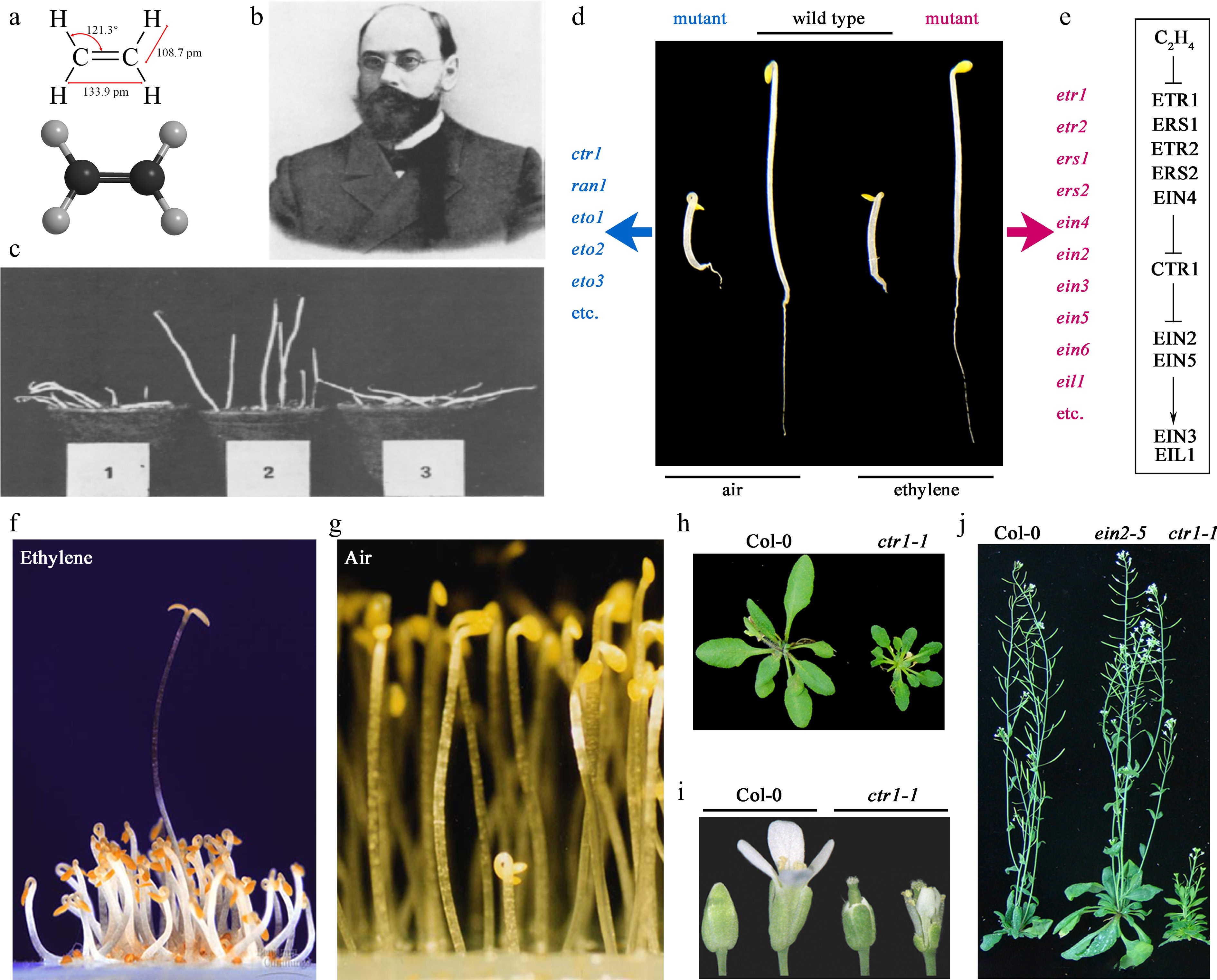
Figure 1.
Arabidopsis 'triple response' phenotype and mutant screening. (a) Chemical structure of ethylene. The upper panel shows a planar diagram with red text indicating bond angles and lengths; the lower panel illustrates a three-dimensional structure with black and gray spheres representing carbon and hydrogen atoms, respectively. (b) Russian botanist Dimitry Nikolayevich Neljubov (1876−1926)[7] (Copyright 1973, Elsevier Science & Technology Journals). (c) Pea seedlings exhibiting abnormal growth due to the influence of illuminating gas[8] (Copyright 1973, Elsevier Science & Technology Journals). Samples 1, 2, and 3 correspond to pea seedlings germinating and growing in greenhouse gas, greenhouse gas filtered through heated CuO, and greenhouse gas filtered through unheated CuO, respectively. (d) Triple response and mutant screening. Etiolated seedlings of wild-type Arabidopsis thaliana (Col-0) grown in the dark for 3 d on Murashige and Skoog (MS) medium in air and in air containing 10 μL·L−1 ethylene[17,23] (Copyright 2010, Oxford University Press). (e) A linear model of the ethylene signaling pathway. (f) Etiolated Arabidopsis seedlings grown for 3 d in the presence of 5 μL·L−1 ethylene. The tallest seedling is the etr1-1 mutant[16] (Copyright 1988, The American Association for the Advancement of Science), which shows no 'triple response'. (g) Etiolated Arabidopsis seedlings grown in air for 3 d. The shortest seedling in the middle, exhibiting the 'triple response' is the ctr1-1 mutant[19] (Copyright 2018, Oxford University Press). (h)–(j) ein2 represents the ethylene-insensitive mutant, while ctr1 represents the constitutive ethylene response mutant[23] (Copyright 2010, Oxford University Press). The mutant phenotypes are distinctly different from the wild type in various aspects such as etiolated seedlings, rosette leaves, floral organs, and mature plants. For example, the ctr1 mutant exhibits a severe constitutive ethylene response phenotype throughout its growth stages. This includes the triple response in air-grown etiolated seedling, delayed hook and cotyledons opening under light, darker green cotyledons, smaller rosette leaves, delayed bolting, smaller inflorescences, early sterility in flowers, and underdeveloped root systems.
HTML
-
Throughout history, humans have inadvertently harnessed the power of ethylene to regulate crop growth and post-harvest physiological changes, even before fully understanding its chemical nature and biological functions. In ancient China, it was observed that fruits near incense burners in temples ripened faster, leading to the development of a traditional 'incense-ripening' technique[4]. Although the mechanism—ethylene release from incomplete incense combustion—remained unknown to practitioners, the method's efficacy ensured its continued use. Modern analysis reveals that smoldering herbal incense generates substantial olefins, with ethylene emissions reaching 1,350 ± 150 μg·g−1[5]. Under typical conditions, this yields gaseous ethylene concentrations of 15–24 μL·L−1, dramatically exceeding levels released by naturally ripening fruits (0.3–0.6 μL·L−1). The ancient Greek philosopher and botanist Theophrastus (c. 371—c. 288 BC) documented a 'wounding-ripening' technique in his work 'Enquiry into Plants'. He noted that sycamore figs were difficult to ripen naturally, but if a wound were made on the fruit, it would soften and ripen within 4 d[6]. In 1901, Dimitry Nikolayevich Neljubov (Fig. 1b)[7], a young researcher at the Botanical Institute of St Petersburg, identified atypical growth in pea seedlings near greenhouse gas pipes[8]: stems reoriented horizontally with reduced elongation and radial thickening under dark conditions (Fig. 1c). Through controlled experiments excluding light/temperature variables, he established that illuminating gas components caused this response. His 1901 breakthrough revealed that ethylene—even at 0.06 μL·L−1—induced dose-dependent, reversible horizontal stem growth, while CuO combustion tube treatments neutralizing hydrocarbons restored normal vertical growth. Neljubov systematically excluded interfering toxins (e.g., SO2, phenols) and demonstrated ethylene's unique ability to override gravitropic responses. At 12.5 μL·L−1, ethylene enforced horizontal stem alignment regardless of initial orientation, proving its dominance over apical growth regulation. Crucially, he established the reversibility of ethylene effects—seedlings resumed vertical growth upon gas removal[8]. This work pioneered gas-phase plant signaling studies. Crocker's team later confirmed ethylene as the active component responsible for gas-mediated growth modulation and floral senescence[9]. Neljubov's 'dark growth-ethylene response' model evolved into the standardized 'triple response' bioassay[10], enabling 20th-century ethylene detection. His experimental rigor—controlled environments, dose-response analysis, and interference exclusion—established foundational methodologies for plant hormone research. These principles not only revealed ethylene as the first gaseous phytohormone but also shaped modern approaches to studying hormonal crosstalk and agricultural applications, cementing ethylene's pivotal role in plant physiology.
In 1934, Richard Gane from the Low Temperature Research Laboratory at the University of Cambridge (Cambridge, England) collected metabolic gas products from about 60 pounds of Worcester Pearmain apples using a bromine-containing Newth collection tube at −65 °C over four weeks. After distillation, he obtained 0.65 g of a substance with a melting point of 62.5 °C, which remained at 62.5 °C when mixed with N,N'-diphenyl ethylene diamine. This was a standard method for determining ethylene at the time, and Gane's discovery was the first time that plants' ability to produce ethylene was confirmed through chemical analysis, providing decisive evidence for later establishing ethylene as a plant hormone[11]. In 1959, Burg & Stolwijk[12] in the United States, as well as Huelin & Kennett[13] in the United Kingdom, introduced gas chromatography into the field of plant hormones, which allowed for precise determination of ethylene at the pmol level, bringing revolutionary changes to ethylene research. Using this advanced technology, it was found that the production rate of ethylene in unripe tomato fruits is low, and the production rate of ethylene gradually increases with fruit ripening, with a peak of ethylene release occurring before the respiratory peak. The increase in ethylene concentration before fruit ripening suggests its role in regulating fruit ripening, providing strong evidence for the hormonal function of ethylene. In light of the multifaceted impact of ethylene on plant growth and development, in 1965, at Burg's suggestion, the scientific community officially recognized ethylene as a plant hormone[14]. This recognition marked an important milestone in ethylene research, laying the foundation for subsequent studies in plant physiology and molecular biology.
-
Arabidopsis (Arabidopsis thaliana) is a small herbaceous plant belonging to the Brassicaceae family. Due to its short life cycle, small genome, and ease of genetic manipulation, it is widely used as a model organism in plant biology research[15]. When Arabidopsis was introduced into the field of ethylene research[16,17], it was found that its etiolated seedlings exhibit a phenomenon similar to what Neljubov observed, known as the 'triple response': in the presence of exogenous ethylene, the apical hook of etiolated seedlings grown for 3–4 d becomes excessively curved, the hypocotyl shortens and thickens, and the root also becomes shorter and thicker (Fig. 1d). Because of the specificity and prominence of this triple response, it has been used as a criterion for determining whether ethylene signaling is activated and is widely applied in studies of ethylene signal transduction pathways and related research[10].
Early-stage investigations on the plant hormone ethylene primarily focused on using physiological and biochemical experimental methods to explore the biochemical pathways of ethylene production in plants and its biological effects[18]. As research progressed, genetic and molecular biology approaches were employed to identify a series of ethylene-related genes (Fig. 1d), gradually revealing the outline of the ethylene biosynthesis and signaling pathways. This included the identification of ethylene-insensitive mutants and constitutive ethylene response mutants (Fig. 1d, f–j)[16,17,19]. In 1995, Roman and colleagues[20] used genetic analysis methods to determine the upstream and downstream relationships of the genes corresponding to these mutants, thereby proposing a nearly linear ethylene signaling pathway (Fig. 1e). Since then, this pathway has been continuously supplemented and refined using various forward and reverse genetic methods. In recent years, with the development of various omics technologies, transcriptomics and proteomics methods have been introduced into the study of the ethylene response pathway, leading to the discovery and elucidation of additional components and regulatory mechanisms involved in ethylene signaling[21,22].
-
Ethylene receptors are transmembrane proteins localized to the endoplasmic reticulum (ER) that negatively regulate ethylene signal transduction, including ETHYLENE RESPONSE 1 (ETR1), ETR2, ETHYLENE RESPONSE SENSOR 1 (ERS1), ERS2, and ETHYLENE INSENSITIVE 4 (EIN4)[24,25]. In the absence of ethylene, these receptors activate the Raf-like kinase CONSTITUTIVE TRIPLE RESPONSE 1 (CTR1), which in turn phosphorylates EIN2[26,27], a key positive component of ethylene signaling. The phosphorylated EIN2 is sequestered and inactivated at the ER membrane[27,28], leading to the degradation of core transcription factors EIN3/EIN3-LIKE 1 (EIL1) via the EIN3 BINDING F-BOX PROTEIN 1 (EBF1)/EBF2-mediated 26S proteasome pathway[23,29−31], thereby suppressing downstream ethylene responses. Upon binding of ethylene to the receptors, the activities of both receptors and CTR1 are repressed, preventing the phosphorylation of EIN2[27,28]. Dephosphorylated EIN2 is subsequently cleaved, releasing its C-terminal fragment from the ER membrane to accumulate in the cytoplasm and nucleus[27,28,32−34]. In the cytoplasm, EIN2 relocates to processing bodies (P-bodies), where it inhibits the translation of EBF1/EBF2 mRNA, thus promoting EIN3/EIL1 accumulation[33,34]. In the nucleus, EIN2 interacts with the histone-binding protein EIN2 NUCLEAR ASSOCIATED PROTEIN 1 (ENAP1) to promote acetylation of histone H3 lysine 14 (H3K14) and lysine 23 (H3K23)[35,36], which may facilitate EIN3/EIL1-mediated transcriptional regulation. Additionally, ethylene enhances EIN2 accumulation by interfering with the EIN2 TARGETING PROTEIN 1 (ETP1)/ETP2-mediated destabilization of EIN2[37]. Ethylene also promotes the proteasome-dependent degradation of EBF1/EBF2 in an EIN2-dependent manner, further stabilizing EIN3/EIL1[23]. In summary, ethylene is initially perceived at the ER, and EIN2 protein transmits the signal from the ER to the nucleus. This results in the accumulation and activation of EIN3/EIL1, which initiates transcriptional regulation and mediates the majority of ethylene-induced biological effects[38] (Fig. 2).
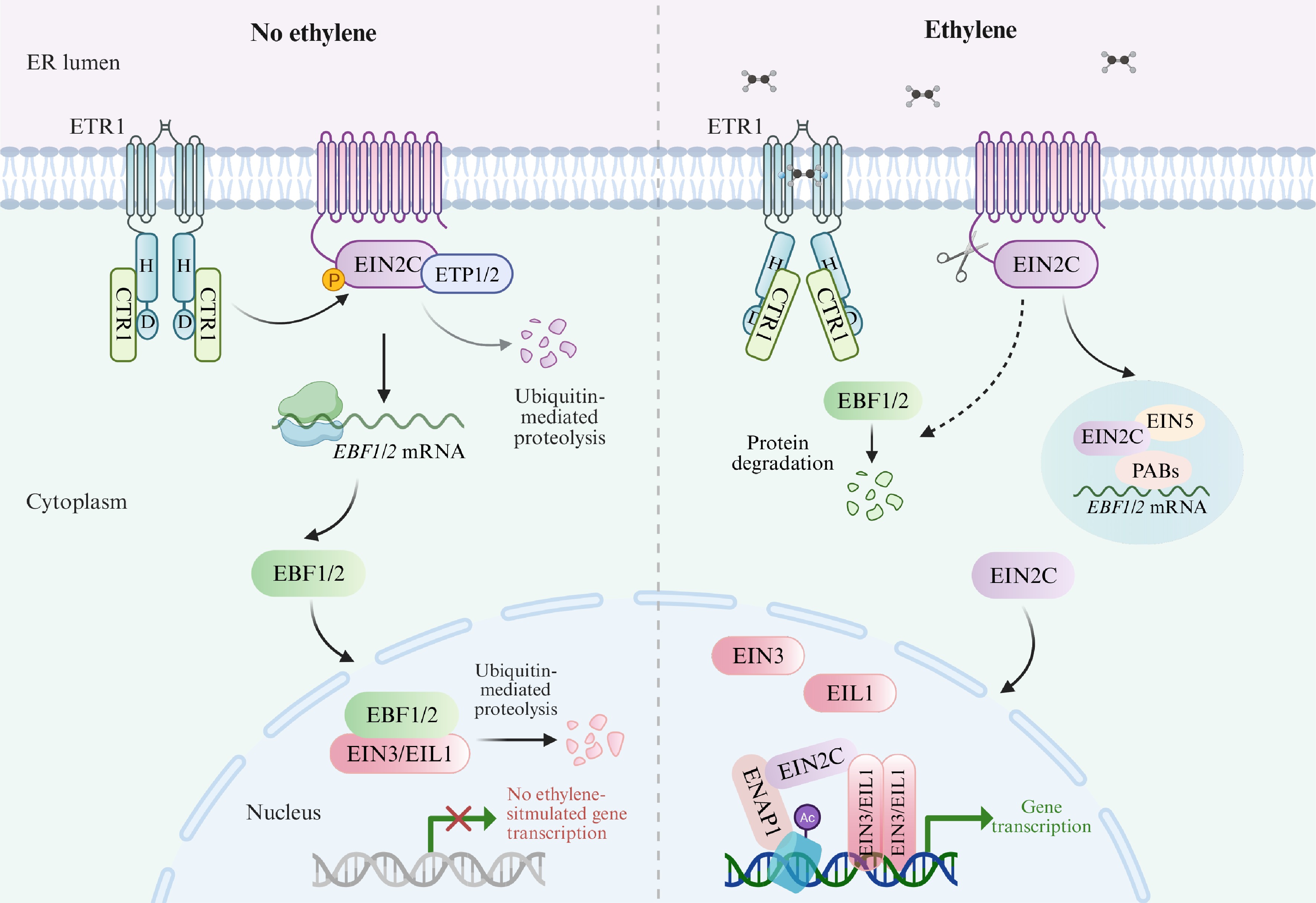
Figure 2.
Schematic diagram of the ethylene signaling pathway in Arabidopsis. In the absence of ethylene, EIN2 is phosphorylated by CTR1, resulting in degradation of EIN3/EIL1 via the EBF1/EBF2-mediated 26S proteasome pathway. Upon ethylene binding, the activities of the receptors and CTR1 are inhibited. Dephosphorylated EIN2 is then cleaved, and the resulting C-terminal fragment is released from the ER membrane to function in the cytoplasm and nucleus. In the cytoplasm, EIN2 targets EBF1/EBF2 mRNAs to P-bodies, suppressing their translation and thereby promoting EIN3/EIL1 accumulation. In the nucleus, EIN2 interacts with ENAP1 to mediate ethylene-induced changes in histone acetylation, regulating expression of EIN3/EIL1 target genes. Additionally, ethylene increases EIN2 abundance by inhibiting ETP1/ETP2-mediated EIN2 destabilization, and also induces proteasomal degradation of EBF1/EBF2 in an EIN2-dependent manner. This figure was created using the BioRender online tool (
BioRender.com ).Notably, it is worth mentioning that certain physiological effects of ethylene are independent of EIN3/EIL1. By utilizing dynamic growth platforms to monitor seedling growth rates in real time, researchers have found that the effects of ethylene occur in two phases[39]. After ethylene treatment, there is an initial rapid decline in growth rate within minutes—this constitutes the first phase. Following a brief plateau, ethylene induces a further gradual decrease in growth rate until a lower, stable state is reached—this is the second phase[39]. Studies have shown that EIN2 is essential for both phases, while EIN3/EIL1 are only required for the second phase[40], suggesting the existence of an EIN2-mediated, EIN3/EIL1-independent rapid ethylene response pathway. Further studies are anticipated to elucidate the mechanisms underlying the rapid response to ethylene, hence providing a comprehensive understanding of how ethylene, as an important gaseous hormone, influences plant growth and stress responses.
-
Plant hormone receptors often function as allosteric switches that translate ligand binding into conformational changes and derepression of signaling repressors. Despite their structural diversity, this mechanism is conserved across various hormone pathways[41]. Ethylene receptors, homologous to cyanobacterial two-component systems, exemplify this model through copper-dependent ethylene binding and complex formation at the ER. Accessory proteins further modulate receptor activity, potentially balancing redundancy with specificity in ethylene signaling.
Biochemical characteristics of ethylene receptors
-
Ethylene receptors share sequence similarity with components of two-component systems found in bacteria[42]. Recent phylogenetic analyses indicate that the ethylene-binding domains present in both cyanobacteria and plants originated from a common ancestor[43]. Accordingly, it is proposed that ethylene receptors have been acquired by plants from the cyanobacterial endosymbiont that eventually evolved into the chloroplast[43]. In Arabidopsis, each receptor contains an N-terminal ER transmembrane domain, a cGMP-specific phosphodiesterases, adenylyl cyclases, and FhIA (GAF) domain, and a C-terminal protein-kinase domain located in the cytoplasm. Additionally, the C-termini of ETR1, ETR2, and EIN4 possess an additional receiver domain[44]. Based on their kinase characteristics, ethylene receptors are divided into two subfamilies. Subfamily I includes ETR1 and ERS1, whereas ETR2, ERS2, and EIN4 belong to subfamily II[44]. The subfamily I receptor ETR1 exhibits histidine kinase activity[45], while ERS1 displays dual activity as both a histidine and serine/threonine kinase in vitro; however, it is generally believed that ERS1 only functions as a serine/threonine kinase in vivo[46]. Subfamily II receptors lack the complete histidine kinase domain and likely function as serine/threonine protein kinases[46]. Subsequent studies have shown that the kinase activities of both subfamily I and II receptors are not necessary for ethylene signaling[25,47]. Thus, although ethylene receptors retain a kinase scaffold resembling that of bacterial two-component systems, plants employ them in a signaling capacity that is largely independent of their intrinsic kinase activity.
Ethylene-binding properties
-
All five ethylene receptors can bind ethylene with high affinity[48−50]. Specific amino acids located in the first and second transmembrane helices of ETR1 are essential for ethylene binding; mutations at these sites cause complete ethylene insensitivity. These residues are arranged along a single face of their respective helices, potentially clustering together to form an ethylene-binding pocket[51]. Another class of mutations, found at the cytoplasmic ends of the first and third TM helices of ETR1, does not impair ethylene binding but renders ETR1 constitutively active, suggesting that these residues may help switch off receptor signaling following ethylene binding[51]. Building on this genetic evidence, a computational model proposed by Schott-Verdugo et al.[52] suggests that ethylene binding within the first and second helices perturbs the third helix, triggering conformational changes that inhibit ETR1 activity and initiate downstream signaling[52]. In particular, residues Ser98 and Pro110 are identified as critical for transmitting the inhibitory signal within ETR1[52].
Ethylene binding requires a copper cofactor, which is coordinated primarily by Cys65 and His69 in the second helix to form a high-affinity ethylene-binding site[52,53]. The ETR1C65Y (etr1-1) cannot bind copper or ethylene, resulting in complete ethylene insensitivity[53]. The ER-localized RESPONSIVE TO ANTAGONIST 1 (RAN1) delivers copper ions to ethylene receptors, a process crucial for receptor maturation and ethylene binding[54−56]. It has long been believed that one receptor dimer binds a single copper ion[53]; however, Schott-Verdugo et al. demonstrated that each ETR1 monomer binds a copper ion[52]. A more recent study by Azhar et al.[57] supports this one-copper-per-monomer model, and further reveals that the conserved residue Asp25 in the first helix is crucial for copper and ethylene binding. Rather than directly coordinating the metal, Asp25 likely stabilizes copper binding through a hydrogen bond with His69. Additionally, Asp25 forms a polar bridge with Lys91, mediating structural changes in response to ethylene binding[57]. Although biochemical and genetic studies have greatly advanced our understanding of ethylene recognition, high-resolution structural data are still needed to fully elucidate the molecular mechanisms that govern receptor-mediated ethylene signaling.
Overlapping but distinct functions of ethylene receptors
-
Initially identified receptor mutants (e.g., etr1-1, etr2-1, and ein4-1), are all dominant, ethylene-insensitive alleles[16,20,58]. Transgenic plants harboring ERS1 or ERS2 missense mutations that correspond to these lesions show the same ethylene-insensitive phenotype[59,60]. Further screening in these backgrounds identified a series of loss-of-function mutations in ethylene receptors that could restore ethylene sensitivity[24]. While etr1 null alleles show hypersensitivity to ethylene, single loss-of-function mutations in the other receptors typically have negligible effects on ethylene response[24,61]. By contrast, higher-order receptor knockout mutants exhibit varying degrees of constitutive ethylene responses[24], revealing functional redundancy among receptors and their overall negative regulation of ethylene signaling.
Despite this redundancy, the functions of the receptors do not entirely overlap. For example, the constitutive triple response phenotype and late-stage growth defects of the etr1 ers1 double loss-of-function mutant are far more pronounced than those of the etr2 ers2 ein4 triple loss-of-function mutant[62]. Therefore, subfamily I receptors are generally considered to play more important roles in ethylene response. Transgenic expression of ETR1 or ERS1 can restore the etr1 ers1 phenotype, whereas expression of subfamily II receptors cannot complement it[25], indicating functional specificity among different receptors. Moreover, loss-of-function mutations in receptors with receiver domains, such as ETR1, ETR2, and EIN4, impair the normal recovery of growth rate after ethylene removal, whereas loss of ERS1 or ERS2 function has no apparent effect on this process[39]. In addition, ethylene receptors also display functional divergence beyond ethylene signaling. For instance, etr1 and ein4 loss-of-function mutants show accelerated seed germination under salt stress conditions. In contrast, the etr2 null allele exhibits delayed germination, whereas ers1 and ers2 single knockout mutants have no detectable effect[63]. Together, these findings show that while the five ethylene receptors act cooperatively to repress the ethylene signaling, each receptor also executes unique roles that fine-tune ethylene response and other developmental processes.
Complex interactions among ethylene receptors
-
Early studies found that the N-termini of the receptors can form two intermolecular disulfide bonds within the ER lumen[49,64]. Whether or not disulfide bonds are formed does not affect the ethylene-binding capacity of ETR1[65]; however, studies suggest that disulfide bonds may stabilize ETR1 dimer during translation, thus increasing the proportion of functional ETR1[66]. Structural analysis reveals that the dimerization histidine phosphotransfer (DHp) domain of ERS1, a subdomain of its kinase region, is also capable of dimerization[67]. Moreover, each receptor can form homodimers or heterodimers with other receptors via their GAF domains in vivo[68,69]. These complex interactions imply that ethylene receptors exist in clusters, and a conformational change in one receptor is likely to influence the conformation of others, thereby amplifying signal output. For example, plants can respond to ethylene concentrations as low as 0.2 nL·L−1, which is at least 300-fold lower than the ethylene-binding constants detected in vitro[38,48]. This high sensitivity and broad response range are likely achieved through interactions among receptors.
REVERSION-TO-ETHYLENE SENSITIVITY 1 (RTE1) was identified in a screen for suppressors of the ethylene-insensitive allele etr1-2[70], and was later shown to encode an ETR1-interacting protein[71]. In addition to reversing the phenotype of etr1-2, the rte1 mutation can also suppress certain other dominant etr1 mutations[72]. However, it does not influence the phenotypes of etr1-1 or other receptor-dominant mutants[70,73]. These observations suggest that the activity of ethylene receptors is subject to very precise regulation, and that RTE1 may specifically affect ETR1 function. RTE1 is primarily localized to the Golgi apparatus and is also distributed on the ER[71,74]. Although its mode of action remains unclear, studies suggest that RTE1 likely stabilizes the conformation of ETR1 by acting through the N-terminus of ETR1[72,75]. The RTE1 homolog, RTE1-HOMOLOG (RTH), directly interacts with RTE1 and modulates the ethylene signaling through it, but functions in opposition to RTE1 in regulating ETR1 activity[76]. Further screening of RTE1-interacting proteins led to the identification of ER-localized cytochrome b5 (Cb5) isoforms[77] and cytoplasmic LIPID TRANSFER PROTEIN 1 (LTP1), which potentially act upstream of RTE1 to enhance ETR1 function[78]. AUXIN-REGULATED GENE INVOLVED IN ORGAN SIZE (ARGOS) proteins in Arabidopsis and maize have also been reported to negatively regulate the ethylene response and interact with RTE1[79,80]. In contrast, ARGOS proteins appear to act downstream of RTE1 to modulate ETR1 function[79].
Furthermore, a separate suppressor screen of etr1-2 identified CONSTITUTIVE EXPRESSOR OF PATHOGENESIS-RELATED GENES 5 (CPR5), a gene encoding a protein that interacts with the N-terminal portion of ETR1[81]. Unlike rte1, the cpr5 mutations suppress the ethylene-insensitive phenotypes of both etr1-1 and etr1-2, yet still do not affect other dominant receptor mutants[81]. Additionally, CPR5 could act as a nucleoporin to regulate the nucleocytoplasmic transport of ethylene-related mRNAs[82]. Apart from TETRATRICOPEPTIDE REPEAT PROTEIN 1 (TRP1), an ERS1-interacting protein reported to be involved in ethylene response[83], interacting proteins of other receptors and their roles in the ethylene signaling remain largely unexplored. In summary, these findings reveal a multilayered regulatory network where receptor clustering and diverse accessory proteins, particularly those associated with ETR1, underscore ethylene perception as a highly integrated and dynamic process. Defining the full repertoire of receptor-interacting proteins and clarifying how their interactions mediate various physiological responses will be crucial for a profound understanding of ethylene perception in plants.
-
Ethylene signaling is orchestrated through multifaceted regulation, beginning with receptor-mediated inhibition of CTR1 and activation of EIN2. The cleaved EIN2 fragment translocates to the nucleus to facilitate transcriptional reprogramming, while concurrently repressing EBF1/EBF2 mRNA translation in the cytoplasm—potentially via phase-separated P-bodies. Emerging evidence of CTR1 nuclear localization and its diverse interactors further underscores the compartmental complexity of ethylene signaling in coordinating development and stress responses.
Negative component CTR1
-
CTR1 is the first gene cloned in the ethylene signaling pathway[19,84]. Loss-of-function mutations in CTR1 lead to constitutive ethylene responses, indicating that CTR1 acts as a negative regulator of ethylene signaling[19]. The C-terminal domain of CTR1 resembles the serine/threonine kinase domain of Raf family proteins, which initiate mitogen-activated protein (MAP)-kinase signaling cascades in mammals[44]. In vitro phosphorylation assays demonstrate that the kinase domain of CTR1 possesses serine/threonine kinase activity[85]. In the ctr1-1 mutant, a highly conserved aspartic acid in the kinase domain is mutated to glutamic acid, resulting in the loss of CTR1 kinase activity. Similar to other complete ctr1 loss-of-function mutants, ctr1-1 exhibits a constitutive ethylene response, suggesting that kinase activity is crucial for CTR1 function[85].
Genetic analyses place CTR1 downstream of ethylene receptors[20,86]. Although CTR1 itself lacks transmembrane domains, it is likely anchored to the ER membrane through interaction with ethylene receptors[26]. A mutation in the ETR1-interaction region of ctr1-8 abolishes CTR1 function, highlighting that ETR1 binding is essential for CTR1 activity[26]. However, how the receptors activate CTR1 and how ethylene inhibits the activity of the receptor-CTR1 complex remain unclear. Additionally, high-order receptor loss-of-function mutants exhibit stronger constitutive ethylene responses and more severe growth defects than ctr1 mutants, suggesting that ethylene receptors possess additional CTR1-independent functions[24,87]. AT4G24480 is thought to be a paralog of CTR1, encoding the RAF6 protein that interacts with both ETR1 and ERS1. However, unlike CTR1, RAF6 plays only a minor, positive role in ethylene signaling[88]. These observations suggest that RAF6 is unlikely to be a major contributor to the CTR1-independent signaling outputs of the ethylene receptors.
Core positive regulator EIN2
-
EIN2 is a single-copy gene in the Arabidopsis thaliana genome whose loss of function causes near-complete insensitivity to ethylene, demonstrating the pivotal role of EIN2 in ethylene response[89]. Further genetic analyses indicate that EIN2 functions downstream of CTR1 and upstream of EIN3/EIL1 in the ethylene signaling pathway[20,29]. EIN2 is an ER-localized integral membrane protein with 12 transmembrane helices at its N terminus, sharing 21% amino acid sequence similarity with the Nramp family of metal transport proteins[89]. However, studies suggest that EIN2 may not function as an ion transporter in ethylene signaling[89]. The hydrophilic C terminus of EIN2 is located in the cytoplasm and shows no homology to any known proteins. Overexpression of the C-terminal end of EIN2 (CEND) does not rescue the ethylene-insensitive phenotype of ein2-5 under dark conditions but induces constitutive ethylene responses in the light[89]. In addition, the C terminus of EIN2 contains a conserved nuclear localization sequence that is essential for both its nuclear localization and its function[28].
EIN2 co-localizes with all ethylene receptors at the ER and directly interacts with them through its C-terminal domain. The kinase domain of receptors is necessary for the interaction, and their phosphorylation state affects the formation of the receptor/EIN2 complex[90,91]. Since the kinase activity of receptors is not required for the classical ethylene signaling[25,47], the biological significance of the receptor-EIN2 interaction remains to be further investigated. Moreover, EIN2 protein is targeted for degradation via the 26S proteasome pathway mediated by two F-box proteins ETP1 and ETP2, while ethylene stabilizes EIN2 protein by promoting the degradation of ETP1/ETP2 and interfering with their interaction with EIN2[37]. However, ethylene induces the accumulation of EIN3 protein more rapidly than that of EIN2[30,37]. This suggests that the regulation of EIN2 protein stability may not be directly involved in ethylene signal transduction, but instead functions as an auxiliary mechanism to amplify ethylene response.
CTR1-EIN2 transmits the signal from the ER to the nucleus
-
For a long time, identifying the substrates of CTR1 kinase remained an unresolved challenge. It was not until 2012 that studies discovered that CTR1 directly interacts with and phosphorylates EIN2, resulting in its retention and inactivation on the ER membrane[27]. Upon ethylene binding to the receptors, the activity of CTR1 is inhibited, leading to EIN2 dephosphorylation and subsequent cleavage. The free C terminus of EIN2 is then translocated from the ER to the nucleus, where it may stabilize EIN3/EIL1 proteins and consequently activate the downstream ethylene response[27,28,32]. Mutation of the nuclear localization sequence prevents EIN2 from entering the nucleus and fails to rescue the phenotype of ein2-5; similarly, sequestering the C terminus of EIN2 in the cytoplasm impedes downstream signaling, underscoring the necessity of EIN2 nuclear targeting[27,32]. Of the multiple phosphorylation sites identified on EIN2, S645 and S924 appear to be critical, as mutations of these residues abolish the CTR1-mediated phosphorylation and impair EIN2 nuclear localization, leading to constitutive ethylene responses[27,28]. Nonetheless, considerable discrepancies remain regarding the cleavage pattern of EIN2 and the functional relevance of its various phosphorylation sites across different studies[27,28,32]. While CTR1-mediated phosphorylation and subsequent cleavage-driven nuclear translocation of EIN2 mark significant advances in elucidating ethylene signaling, the precise molecular mechanisms controlling EIN2 cleavage are still a matter of ongoing debate.
Importantly, ethylene also triggers the localization of EIN2 to cytoplasmic punctate structures that partially co-localize with P-bodies[33,34]. EIN2 directs EBF1/EBF2 mRNAs to P-bodies by associating with their 3' untranslated regions (3'UTRs), repressing their translation and thereby preventing the EBF1/EBF2-mediated degradation of EIN3/EIL1 proteins[33,34]. Hence, overexpression of the EBF1 3' UTR alone can substantially reduce ethylene sensitivity by alleviating the translational inhibition of endogenous genes, leading to the overaccumulation of EBF1 protein[33]. The translational control of EIN2 is achieved through its interaction with other P-body components, such as EIN5 and poly(A)-binding (PAB) proteins. Mutants of these components also display pronounced ethylene-insensitive phenotypes[33,34]. Recent findings highlight that EIN2 harbors intrinsic sequence features favoring liquid-liquid phase separation (LLPS), and loss of EIN2 function perturbs the translation of multiple LLPS-related gene sets[92]. In addition to P-bodies, EIN2 localizes to stress granules, another type of membraneless condensate, in response to heat stress[92]. These observations raise the possibility that phase separation may represent an additional layer in EIN2-mediated ethylene signaling, although the direct mechanistic role of LLPS remains to be elucidated.
Recent studies reveal that ethylene induces the translocation of CTR1 to the nucleus in a manner independent of its kinase activity[93]. Once in the nucleus, CTR1 interacts with EBF1/EBF2 to promote EIN3 protein accumulation, thus positively regulating ethylene signaling. Elevated CTR1 levels in the nucleus further enhance plant tolerance to drought and salt stresses[93]. Moreover, proximity labeling analyses of the CTR1 interactome indicate that, beyond the classical ethylene signaling pathway, CTR1-interacting proteins are involved in various biological processes, including mitochondrial respiration, mRNA metabolism, and organelle biogenesis[94]. Overall, these findings reveal that multi-compartmental localization of CTR1 and EIN2 is central to the precise transduction of ethylene signals and their integration into broader cellular processes. A thorough elucidation of the molecular logic behind these dynamic localizations is fundamental to advancing our understanding of ethylene signaling across diverse environmental conditions.
-
Nuclear events in ethylene signaling are primarily governed by the transcription factors EIN3 and EIL1, whose accumulation is tightly regulated by the F-box proteins EBF1 and EBF2. This regulatory module forms the core of ethylene signal transduction in the nucleus and is subject to multilayered control, including transcriptional feedback, mRNA decay, translational repression, and proteasomal turnover. The divergent spatial and temporal activities of EBF1 and EBF2 further enable dynamic and context-specific modulation of EIN3/EIL1 protein levels.
Key transcription factors EIN3 and EIL1
-
The ein3 mutants were initially identified through genetic screens for ethylene-insensitive phenotypes[20,29]. EIN3 encodes a nuclear-localized transcription factor unique to plants, comprising at least three functional domains: a DNA-binding domain (DBD; amino acids 80–359), a dimerization domain (amino acids 113–257), and a C-terminal regulatory domain responsible for interactions with other proteins[29,95,96]. Further studies have refined the core DBD to residues 174–306, and its high-resolution crystal structure reveals a novel α-helical fold and key residues critical for DNA binding[97]. This work also shows that EIN3 binds DNA as a homodimer, with high-affinity binding dependent on the presence of two inverted EIN3-binding sites[97]. In Arabidopsis thaliana, EIN3 has five homologs, EIL1–EIL5, among which EIL1 shares the highest sequence similarity with EIN3[29]. Although the eil1 single mutant exhibits only mild ethylene insensitivity, the ein3 eil1 double mutant is almost completely insensitive to ethylene, resembling ein2 in many traits[23,29]. Moreover, overexpression of EIL1 in the ein3 mutant background restores ethylene responsiveness[29], indicating that EIN3 and EIL1 are the primary transcription factors in the ethylene signaling pathway with functional redundancy.
Controlling of EIN3/EIL1 protein stability by EBF1/EBF2
-
Unlike the aforementioned ethylene signaling components, EBF1 and EBF2 were not identified in early genetic screens but were instead discovered by screening for EIN3-interacting proteins[30,31]. Both EBF1 and EBF2 are nuclear-localized proteins that contain a conserved F-box domain at their N termini, enabling them to form SCF-type E3 ubiquitin ligases together with ARABIDOPSIS SKP1 HOMOLOGUE 1(ASK1) and CULLIN 1 (CUL1). Their C-terminal regions comprise 18 tandem leucine-rich repeats (LRRs), which mediate substrate recognition, specifically targeting EIN3/EIL1 for proteasomal degradation[30,31]. Due to functional redundancy, ebf1 and ebf2 single loss-of-function mutants display only mild ethylene hypersensitivity, while the ebf1 ebf2 double mutant shows overaccumulation of EIN3/EIL1 proteins and a strong constitutive ethylene response, which can be fully rescued by mutations in EIN3 and EIL1[23,30,31]. Importantly, ethylene can no longer influence EIN3 abundance in the ebf1 ebf2 double mutant, highlighting the critical role of EBF1 and EBF2 in mediating the ethylene-induced turnover of EIN3[23].
Despite their redundancy, EBF1 and EBF2 also exhibit distinct roles. Specifically, ebf2 mutants display a more pronounced ethylene hypersensitivity during seedling development, whereas ebf1 mutants show greater growth inhibition at later, mature stages[30,31,98]. Moreover, time-course analyses of hypocotyl growth dynamics have revealed that EBF1 primarily functions in the absence of ethylene and during the early stages of signaling, while EBF2 acts predominantly in the later stages of the ethylene response and is critical for the recovery of growth following ethylene removal[99].
Additionally, studies have shown that phosphorylation of EIN3 at different residues, mediated by MAP KINASE KINASE 9 (MKK9) – MAP KINASE 3 (MPK3)/MPK6[100], can affect EIN3 protein stability. However, it is generally accepted that the MAPK cascade primarily regulates ethylene biosynthesis under stress conditions and does not directly control EIN3 stability[23,101]. Therefore, the biological significance of EIN3 phosphorylation remains to be further elucidated. Overall, these findings underscore the pivotal roles of EBF1 and EBF2 in fine-tuning ethylene signaling by targeting EIN3/EIL1 for proteasomal degradation. Their functional divergence also facilitates the delicate regulation of ethylene responses across developmental stages and signaling phases.
Multilayered regulation of EBF1/EBF2 governs EIN3-mediated ethylene response
-
Effective activation of ethylene signaling is essential for plants to appropriately respond to developmental and environmental cues; however, sustained overactivation of ethylene response has obvious deleterious effects on plant development, including dwarfism, reduced fertility, and even lethality. Consequently, tight regulation of the ethylene signaling pathway is crucial. Studies have shown that the expression of both EBF1 and EBF2 genes is induced by ethylene, with EBF2 showing a more pronounced upregulation[98,102]. EIN3 directly binds to the promoter of EBF2 and activates its expression, establishing a negative feedback loop to maintain optimal levels of EIN3/EIL1 proteins, hence preventing the overactivation of ethylene response[102]. In addition to cooperating with EIN2 to repress the translation of EBF1/EBF2 mRNAs, the 5'–3' exoribonuclease EIN5 was initially identified as promoting the degradation of EBF1/EBF2 transcripts. This likely represents an additional regulatory layer in ethylene signaling, potentially counteracting the negative feedback exerted by EBF1/EBF2 on EIN3[103,104]. Recent findings reveal that the DCP1-ASSOCIATED NYN ENDORIBONUCLEASE 1 (DNE1) promotes ethylene signaling by cleaving the 3'UTRs of EBF1/EBF2 mRNAs, thereby destabilizing the transcripts and suppressing their translation[105]. Additionally, EBF1/EBF2 proteins are targeted for proteasome degradation in an EIN2-dependent manner upon ethylene treatment[23]; however, the underlying mechanism remains elusive. These findings collectively highlight a tightly coordinated regulatory framework governing EBF1/EBF2, spanning from transcriptional regulation to protein turnover. This multilayered control allows dynamic modulation of EIN3 abundance, thus fine-tuning ethylene responses.
-
Early studies revealed that members of the conserved histone acetyltransferase (HAC) family, specifically HAC1 and HAC5, act as repressors of ethylene response. The hac1 hac5 double mutant exhibits a pronounced constitutive triple response phenotype, accompanied by elevated expression levels of ethylene-responsive genes[106]. In addition, the histone acetyltransferase GENERAL CONTROL NON-REPRESSIBLE 5 (GCN5) synergizes with the receptor kinase CLAVATA 1 (CLV1) to suppress ethylene response, as evidenced by the strong constitutive triple response observed in the gcn5 clv1 double mutant[107]. In this mutant, acetylation levels of histone H3K9 and H3K14 in the promoter regions of ethylene-responsive genes are increased, which may contribute to the upregulation of these genes[107]. Moreover, overexpression of HISTONE DEACETYLASE 19 (HDA19) upregulates the expression of ethylene-regulated genes, including ETHYLENE RESPONSE FACTOR 1 (ERF1)[108]. Taken together, these results suggest that histone acetyltransferases inhibit, while histone deacetylases activate, the expression of ethylene-responsive genes, which seemingly contradicts the conventional model where histone acetylation is generally associated with gene activation.
It has been reported that ethylene treatment significantly increases H3K14 and H3K23 acetylation to promote transcription. This elevation depends on EIN2 and its interacting protein ENAP1, which possesses histone-binding activity[35,36]. Although ethylene treatment does not alter global H3K9 acetylation levels, promoters of ethylene-induced genes display significantly higher H3K9 acetylation compared to ethylene-repressed genes[35,36], suggesting that H3K9 acetylation may serve as a pre-existing molecular mark distinguishing transcriptional activation from repression. Furthermore, histone deacetylases SIRTUIN 1 (SRT1) and SRT2 interact with ENAP1 to reduce H3K9 acetylation, thereby mediating ethylene-induced transcriptional repression[109]. More recent findings reveal that ethylene triggers the nuclear accumulation of the pyruvate dehydrogenase complex (PDC) in an EIN2-CEND-dependent manner to synthesize acetyl coenzyme A in the nucleus, which facilitates EIN2-CEND-directed histone acetylation and transcription regulation[110]. Additionally, upon temperature elevation, EIN2-CEND and EIN3 accumulate and cooperate with LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), a reader of H3K27me3, to target specific bivalent genes. The chromatin remodeler INOSITOL AUXOTROPHY 80 (INO80) is concurrently recruited via interaction with EIN2-CEND and EIN3 to promote H3K4me3 deposition, counteracting the repressive effects of H3K27me3 and enabling rapid gene activation in response to warm temperatures[111]. These studies advance our understanding of EIN2-mediated histone acetylation in ethylene-related transcriptional regulation. Nonetheless, the mechanisms by which epigenetic modifications directly influence EIN3 transcriptional activity remain to be thoroughly investigated.
-
Ethylene signaling forms an intricate nexus with numerous hormonal and environmental pathways, steering plant growth, development, and stress adaptation. This network centers on three hubs: the receptor ETR1, the transducer EIN2, and the transcription factor EIN3. ETR1 extends beyond canonical ethylene perception, intersecting with cytokinin via AHP–ARR phosphotransfer and influencing redox homeostasis and translational control. EIN2 integrates signals such as salt, flooding, senescence, defense, and glucose through phosphorylation-controlled nuclear trafficking, and regulates abscisic acid (ABA) signaling independently of EIN3/EIL1. The stability and activity of EIN3 are shaped by a broad range of stimuli. In turn, EIN3 reprograms transcriptional networks through interactions with other transcription factors. Together, these modules underscore the versatility and centrality of ethylene signaling in plant adaptation.
Ethylene receptor ETR1-mediated crosstalk
-
In Arabidopsis, the receptors of phytohormone cytokinin also belong to the two-component receptor family[112]. However, unlike ethylene signaling, cytokinin signaling relies on a conserved histidine-to-aspartate phosphorelay. Upon cytokinin binding, the receptor autophosphorylates on a histidine residue and subsequently transfers the phosphate to an aspartate residue in its receiver domain[112]. The phosphate is then relayed to ARABIDOPSIS HISTIDINE-CONTAINING PHOSPHOTRANSMITTER (AHP) proteins, which translocate to the nucleus and transfer the phosphate to ARABIDOPSIS RESPONSE REGULATOR (ARR) transcription factors, initiating the regulation of cytokinin-responsive genes[112].
Although the kinase activity of ETR1 is dispensable for classical ethylene signaling, emerging evidence indicates that it contributes to processes such as the recovery of growth rates after ethylene removal, root apical meristem growth, and seed germination under stress conditions[38], suggesting that ETR1 may also function via a phosphotransfer mechanism. Interestingly, ETR1 has been shown to interact with multiple AHP proteins through its receiver domain[113], and the phosphorylation state influences its interaction with AHP1[114]. ETR1 affects the expression of ARR3 by interacting with AHP proteins, thereby mediating the regulation of root apical meristem by both ethylene and cytokinin[113]. Furthermore, mutants harboring multiple AHP and ARR mutations exhibit delayed growth recovery after ethylene withdrawal, a process dependent on the kinase activity of ETR1 but independent of cytokinin biosynthesis or receptor function[115]. These findings suggest that the ETR1–AHP–ARR module represents a non-canonical branch of ethylene signaling. However, under certain conditions, this module may also contribute to crosstalk between ethylene and cytokinin signaling pathways.
Additionally, it is worth noting that ETR1, but not EIN2, is indispensable for the generation of H2O2 in guard cells upon stimulation by ethylene, salicylic acid, and UV-B[116−118]. Different ethylene receptors exhibit distinct roles in this process[116]. Further investigation into the molecular basis of ETR1-mediated H2O2 production may uncover a unique link between ethylene receptor and redox homeostasis. Moreover, the flooding-induced phosphorylation of EUKARYOTIC TRANSLATION INITIATION FACTOR 2α (eIF2α) is entirely dependent on ETR1 but not on EIN2, suggesting a potential role for ETR1 in translational regulation[119]. Promisingly, further dissecting the non-canonical functions of ETR1 will unveil new dimensions of receptor functional versatility.
Signal interplays through CTR1 and EIN2
-
In addition to ethylene, salt stress inhibits CTR1 kinase activity through direct phosphorylation of its Ser87 residue by SALT OVERLY SENSITIVE 2 (SOS2), hence activating downstream ethylene signaling events, including EIN2 cleavage, nuclear translocation, and EIN3 accumulation[120]. A similar mechanism has been observed under flooding stress, where elevated levels of unsaturated very-long-chain ceramides, a class of bioactive sphingolipids, directly bind to CTR1. This interaction inhibits CTR1 kinase activity, thus promoting EIN2 nuclear entry and EIN3 accumulation[121]. Moreover, MPK3/MPK6 interacts with the C-terminus of EIN2, facilitating its translocation from the ER to the nucleus in guard cells upon ethylene treatment, leading to EIN3 accumulation and subsequent nitric oxide production and stomatal closure[122]. In the regulation of leaf senescence, MPK6 mediates jasmonic acid (JA)-induced nuclear translocation of EIN2, promoting EIN3 accumulation and activating the expression of ORESARA9 (ORE9)[123]. Recent studies also show that IMPORTIN β1 (IMPβ1) interacts with EIN2-CEND and mediates its nuclear translocation in response to ethylene or aphid infestation, activating phloem-based defense responses in Arabidopsis[124].
Although the above signals modulate EIN2 activity, they ultimately regulate plant responses primarily through modulating EIN3/EIL1 accumulation. However, emerging evidence suggests that EIN2 also functions independently of EIN3/EIL1. Glucose-activated TARGET OF RAPAMYCIN (TOR) kinase directly phosphorylates EIN2 at T657, preventing its nuclear localization[125]. Under TOR repression, EIN2 is not cleaved but instead translocates to the nucleus in its full-length form. Notably, the glucose-TOR-EIN2T657 and ethylene-CTR1-EIN2S645 pathways operate independently to regulate distinct biological processes[125]. Downstream of EIN2, TOR influences root apical meristem cell proliferation in light-grown seedlings through the transcription factor E2Fa, independently of EIN3/EIL1[125]. Additionally, EIN2 directly inhibits the N-acetyltransferase HOOKLESS (HLS1)-promoted histone H3 acetylation at key loci involved in the signaling of phytohormone ABA, such as ABA INSENSITIVE 3 (ABI3) and ABI5, thereby suppressing ABA signaling in an EIN3/EIL1-independent manner[126].
Collectively, these studies suggest that the phosphorylation of EIN2 serves as a critical node integrating ethylene signaling with other pathways. Distinct phosphorylation states of EIN2 may relay different upstream signals. Elucidating the phosphorylation dynamics of EIN2, particularly the identification of novel phosphorylation sites and kinases beyond CTR1 under specific environmental conditions, will be crucial for deepening our understanding of the multifaced regulatory roles of this key signaling hub.
Signaling networks centered on EIN3 protein stability
-
It is well established that EIN3/EIL1 play critical roles in integrating ethylene response with various other stimuli (Fig. 3). In addition to ethylene, glucose was the first reported regulator of EIN3 protein stability. Conversely, glucose promotes EIN3 degradation, and the ein3 mutant exhibits hypersensitivity to glucose[127]. In leaf senescence regulation, the energy-sensing protein ARABIDOPSIS SNF1 KINASE HOMOLOG 10 (AKIN10) directly phosphorylates EIN3, enhancing its degradation and hence delaying senescence[128]. Auxin has been shown to increase EIN3 accumulation, potentially through an EBF1/EBF2-independent pathway[129], whereas cold and salt stresses stabilize EIN3 protein by promoting the degradation of EBF1/EBF2[130,131]. Additionally, JA upregulates EBF1 mRNA level through MYC2, while brassinosteroids (BR) downregulate EBF1/EBF2 expression via BRASSINOZALE-RESISTANT 1 (BZR1), leading to opposite effects on EIN3 accumulation and apical hook formation[132,133].
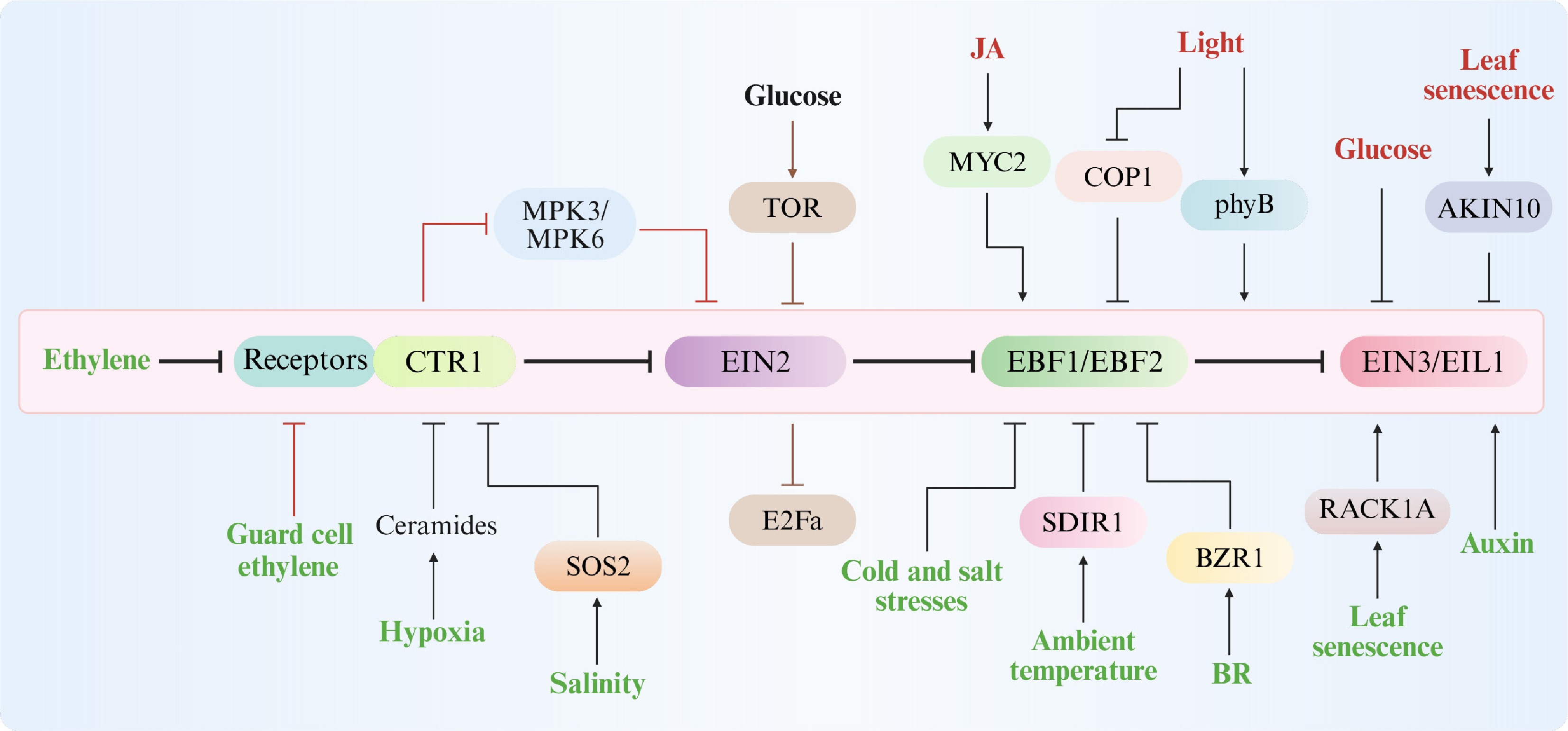
Figure 3.
Signaling networks centered on EIN3 protein stability. This figure illustrates the regulatory network of diverse signals modulating EIN3 protein stability, excluding the TOR-mediated pathway. Most signals regulate EIN3 accumulation by targeting EBF1/EBF2, while others act through upstream ethylene signaling components, such as CTR1 and EIN2, ultimately influencing EIN3 protein levels. The final effects on EIN3 accumulation are indicated in green and red, representing promotion and inhibition, respectively. The red lines represent the stomatal-specific pathway mediated by MPK3/MPK6, while the brown lines indicate the TOR-mediated regulation of EIN2, utilizing E2Fa rather than EIN3/EIL1 as downstream signaling components. This figure was created using the BioRender online tool (
BioRender.com ).CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) was the first identified EBF1/EBF2 ubiquitin ligase targeting EBF1/EBF2 for degradation via the 26S proteasome pathway. During seedling emergence, increasing light intensity represses COP1 activity, thus promoting EIN3 degradation[134]. Moreover, following the transition from dark to light, the red-light receptor PHYTOCHROME B (PhyB) translocates into the nucleus and enhances the interaction between EBF1/EBF2 and EIN3, leading to rapid EIN3 degradation[135]. Beyond targeting EIN3/EIL1, EBF1/EBF2 also mediate the ubiquitination and proteasomal degradation of PHYTOCHROME INTERACTING FACTOR 3 (PIF3), another mechanism contributing to photomorphogenesis[136]. Additionally, the zinc-finger E3 ligase SALT- AND DROUGHT-INDUCED RING FINGER 1 (SDIR1) has been identified as another regulator of EBF1/EBF2. SDIR1 directly ubiquitinates EBF1/EBF2 and promotes their degradation, thereby modulating EIN3 protein levels and ethylene responses under varying ambient temperatures[137].
In summary, these findings reveal that EIN3 protein stability is tightly regulated by a complex network of hormonal, environmental, and energy-related signals, largely mediated through the modulation of EBF1/EBF2 activity. Future studies aimed at unraveling the precise mechanisms coordinating these diverse signaling pathways will shed light on how EIN3 coordinates the trade-off between plant growth and stress tolerance.
Transcriptional activity regulatory networks mediated by EIN3
-
In addition to regulating protein stability, EIN3 transcriptional activity is intricately involved in various biological processes (Fig. 4). This regulatory complexity was first revealed through the interplay between ethylene and JA during root hair development[138]. JASMONATE-ZIM-DOMAIN (JAZ) proteins recruit HDA6 to prevent EIN3 from binding to its target genes, while JA promotes JAZ protein degradation, hence releasing the inhibition on EIN3 activity[138]. A similar mechanism operates in gibberellin acid (GA) signaling, where DELLA proteins directly repress EIN3-mediated activation of HLS1. GA promotes apical hook formation by inducing DELLA degradation, thus alleviating the suppression of EIN3[139]. Interestingly, although JA and ethylene act synergistically in root hair development, JA antagonizes ethylene-induced apical hook formation by inhibiting EIN3 transcriptional activity via MYC2[132,140]. In a reciprocal manner, EIN3 restricts MYC2 activity to negatively regulate JA-mediated defense responses against herbivores[140]. Further complexity arises in salicylic acid (SA) signaling: NONEXPRESSER OF PR GENES 1 (NPR1), a key component of SA pathway, directly suppresses EIN3 transcriptional activity to inhibit hook formation[141]. However, subsequent studies reveal that NPR1 can also enhance EIN3 activity, hence coordinating the promotive effects of SA and ethylene on leaf senescence[142,143]. Together, these findings highlight the critical role of EIN3 transcriptional activity in orchestrating diverse hormone signaling pathways.
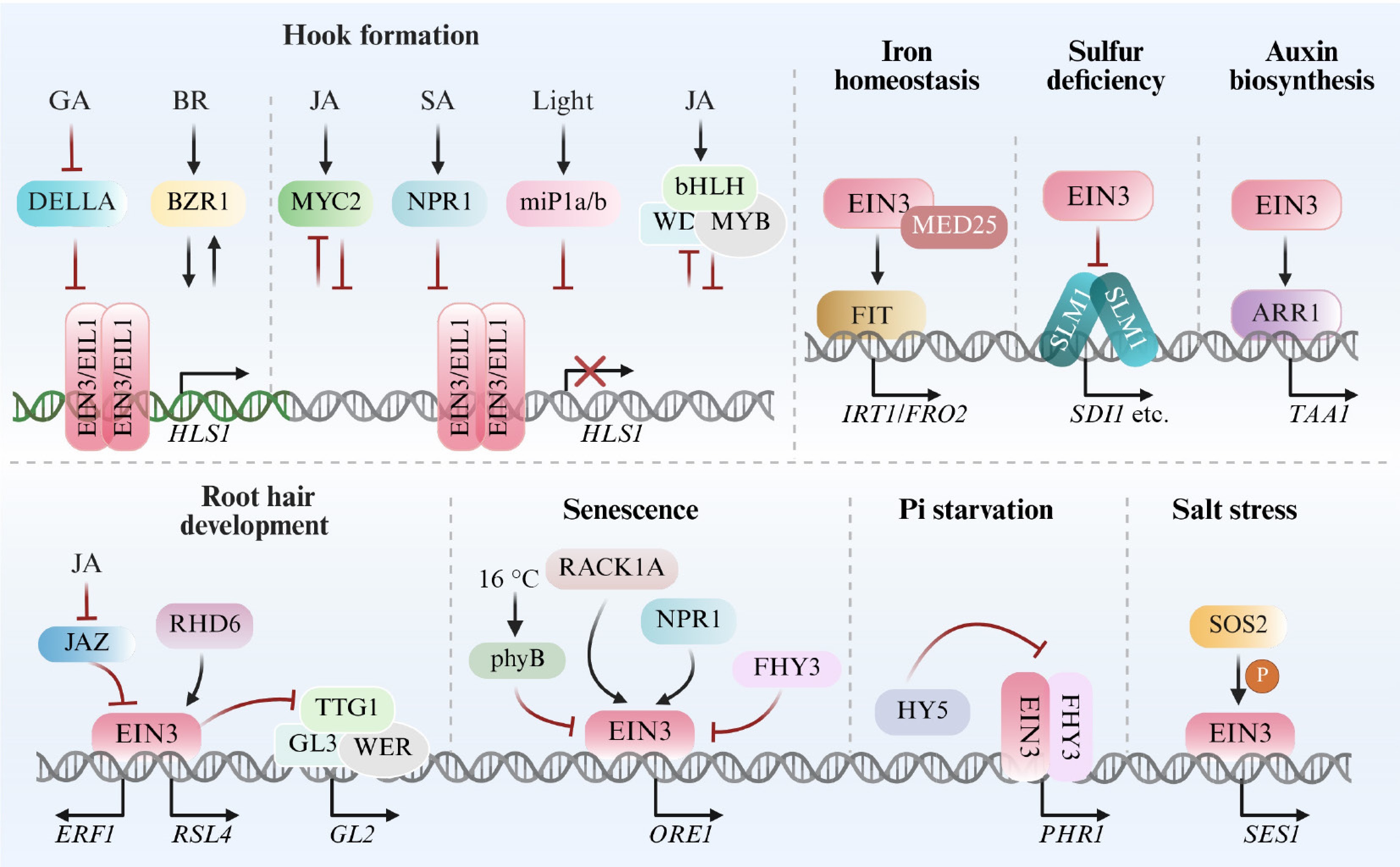
Figure 4.
Transcriptional activity regulatory networks mediated by EIN3. These signaling networks are involved in apical hook formation, root hair development, leaf senescence, and various stress responses. In addition to the upstream regulation of EIN3 activity, the figure also illustrates the influences of EIN3 on other transcription factors. Abbreviations: IRT1, IRON-REGULATED TRANSPORTER 1; FRO2, FERRIC REDUCTION OXIDASE 2; SDI1, SULPHUR DEFICIENCY-INDUCED 1. This figure was created using the BioRender online tool (BioRender.com).
Additionally, EIN3 transcriptional regulation also integrates multiple photomorphogenesis-related components, contributing to apical hook development, stress adaptation, and senescence. For example, a group of microproteins miP1a/1b, has been shown to interact with EIN3, disrupting its self-interaction and interfering with its oligomerization with PIFs, ultimately repressing EIN3 transcriptional activity during photomorphogenesis[144]. FAR-RED ELONGATED HYPOCOTYL 3 (FHY3) interacts with EIN3 to synergistically activate the expression of PHOSPHATE STARVATION RESPONSE 1 (PHR1), a key regulator in phosphate starvation responses, whereas ELONGATED HYPOCOTYL 5 (HY5) antagonizes this activation[145]. Conversely, FHY3 can also interfere with EIN3 binding to ORESARA 1 (ORE1), repressing its expression and delaying leaf senescence[146].
Apart from affecting EIN3 protein stability, phyB suppresses EIN3 transcriptional activity under low environmental temperatures (16 °C) to regulate senescence[147]. Another senescence-related regulator, RECEPTOR FOR ACTIVATED C KINASE 1A (RACK1A), promotes EIN3 protein accumulation and enhances its DNA binding ability, which in turn accelerates leaf senescence[148]. Furthermore, post-translational modifications of EIN3 have been shown to affect its activity. For instance, SOS2 directly phosphorylates EIN3 at Ser325, enhancing its transcriptional activation of ETHYLENE AND SALT-INDUCIBLE ERF1 (ESE1) and hence improving plant salt tolerance[149]. Collectively, these regulatory layers underscore the multifaceted mechanisms controlling EIN3 activity, positioning it as a central hub that integrates environmental cues and endogenous signals. Notably, the modulation of EIN3 transcriptional activity is highly context-dependent, with individual regulators exerting distinct or even opposing effects under different conditions. This dynamic and flexible regulation is critical for enhancing plant adaptability to fluctuating environments.
Building upon the regulation of EIN3 activity by diverse upstream factors, EIN3 itself also exerts control over the activity of other transcription factors through direct interactions. In the context of root hair elongation, EIN3 interacts with ROOT HAIR DEFECTIVE 6 (RHD6) to synergistically induce the expression of RHD6-LIKE 4 (RSL4), a key determinant of root hair length[150]. Meanwhile, EIN3 interferes with the formation of the WERWOLF (WER)-GLABRA 3 (GL3) – TRANSPARENT TESTA GLABRA 1 (TTG1) ternary complex[151], which is crucial for the transcriptional activation of the root hair fate-determining gene GL2. Additionally, EIN3 can directly bind to the GL2 promoter to further repress its expression[151]. Subsequent studies also reveal that EIN3 and EIL1 interact with several components of the MYB-bHLH-WD40 (MBW) complex, disturbing its complex formation and suppressing its transcriptional activity. This leads to altered anthocyanin biosynthesis, trichome development, and reduced resistance to herbivorous insects[152]. Conversely, MBW complex components can inhibit EIN3-mediated activation of HLS1, thus influencing apical hook formation[152].
Beyond these interactions, EIN3 modulates the activity of key transcription factors involved in hormonal and nutrient signaling pathways. For instance, EIN3 collaborates with BZR1 to co-upregulate HLS1 gene expression[153] and enhances BZR1's binding and transcriptional activation of SMALL AUXIN UP RNA 17 (SAUR17)[133]. Similarly, EIN3 promotes the transcriptional activity of ARR1, strengthening its induction of the auxin biosynthesis gene TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1 (TAA1)[154]. In nutrient regulation, EIN3 facilitates the protein accumulation of FE-DEFICIENCY INDUCED TRANSCRIPTION FACTOR (FIT), the core transcription factor involved in iron acquisition. EIN3 interacts with FIT and MEDIATOR 25 (MED25) to cooperatively maintain iron homeostasis in plants[155,156]. Furthermore, EIN3 interacts with SULFUR LIMITATION 1 (SLIM1)/EIL3, a member of the EIN3 family involved in sulfur deficiency responses, interfering with its DNA-binding ability and modulating sulfur-responsive gene expression[157].
Altogether, while upstream regulators fine-tune EIN3 transcriptional activity to modulate plant responses, EIN3, in turn, actively reprograms transcriptional networks by modulating the activity of other transcription factors. This bidirectional regulatory capacity underscores the pivotal role of EIN3 in integrating signaling pathways and orchestrating plant homeostasis.
-
This review comprehensively explores ethylene's roles in Arabidopsis, with a focus on the fundamental mechanisms of its signal transduction pathway. A deeper understanding of these mechanisms is essential for grasping the basic principles that govern plant growth and development. Continued progress will rest on answering key mechanistic questions while translating basic insights into climate-resilient applications. The most pressing avenues for future research are outlined below:
(1) Structural basis of ethylene perception. High-resolution structures of ethylene receptors remain essential. Cryo-EM can reveal how copper-coordinated ethylene binding drives receptor conformational shifts, whereas single-molecule tracking in living cells should map real-time receptor clustering and CTR1 activation.
(2) Functional dissection of EIN2. The Nramp-homology domain of EIN2 is predicted to function as a divalent cation channel, with AlphaFold2 structural modeling supporting its potential role as a calcium transporter (https://alphafold.com/). However, direct experimental evidence is still lacking to confirm its ion channel activity. In addition, the phase separation behavior of EIN2 and its molecular role in translational regulation require further elucidation. Comprehensive phosphoproteomic maps are also needed to clarify the context-specific phosphorylation and cleavage patterns of EIN2.
(3) Epigenetic regulation of ethylene signaling. Beyond histone acetylation, the roles of DNA methylation, SWItch/Sucrose Non-Fermentable (SWI/SNF) chromatin remodelers, RNA modifications (e.g., m6A), and non-coding RNAs in ethylene responses remain largely unexplored. Ethylene-induced long non-coding RNAs (lncRNAs) could potentially act as scaffolds that recruit EIN3 to chromatin[158].
(4) Resolving ethylene response at single-cell resolution. Traditional studies of ethylene response have largely relied on whole-organ or bulk-tissue analyses, masking the spatiotemporal and cell-type-specific complexity of ethylene responses. The advent of single-cell and spatial transcriptomic technologies now offers unprecedented opportunities to interrogate ethylene-mediated response at cellular resolution. Leveraging these technologies will clarify how ethylene shapes plant development through cell-specific, time-dynamic, and environment-responsive networks. This knowledge is crucial for decoding ethylene's control of apical-hook formation, floral organogenesis, leaf senescence, fruit ripening, and the underlying cell-type-specific hormonal crosstalk[159].
(5) Evolutionary and ecological perspectives. Comparative studies across land plants—including bryophytes with relatively primitive ethylene responses—may help elucidate how ethylene signaling evolved and diversified during plant terrestrialization[160]. In parallel, exploring ethylene's role in plant–microbiome interactions, such as rhizobial symbiosis and pathogen resistance[161], could potentially reveal novel receptor-mediated communication mechanisms. Ethylene emission may also function as a volatile ecological cue that influences neighboring plants or pollinators, although this role remains largely underexplored. With the advancement of portable laser-based spectroscopy, real-time monitoring of ethylene plumes in the field may be feasible, opening up opportunities to investigate ecological phenomena such as interplant signaling and weed–crop 'eavesdropping' [162].
(6) Harnessing ethylene signaling for crop trait improvement. Although the core components of ethylene signaling are broadly conserved across plant species, species-specific variations also exist. In rice, several unique ethylene-associated regulators—such as MAO HU ZI 3 (MHZ3), MHZ9, and MHZ11—have been identified[22]. Ethylene plays pivotal roles in regulating key agronomic traits, including fruit ripening, petal senescence, and organ abscission[163,164]. Advances in genomic resources and molecular tools have accelerated ethylene-related studies in fruit and ornamental crops such as tomato, carnation, and rose[163,165,166]. More recently, maize research identified a Zea genus-specific micropeptide that modulates ZmEIN3/ZmEIL1 expression and thereby affects kernel dehydration, highlighting ethylene's additional role in regulating grain dehydration for timely harvest[167]. Ongoing research across diverse species is expected to uncover additional regulatory modules, informing targeted strategies for trait optimization in crop breeding.
In conclusion, ethylene biology stands at the threshold of a new era—one shaped by the integration of structural biology, systems-level ecology, and climate-resilient biotechnology. By bridging fundamental molecular mechanisms with translational applications, future research will not only deepen our understanding of plant development but also accelerate innovations urgently needed for sustainable and adaptive agriculture.
This work was supported by the National Natural Science Foundation of China (Grant No. W2521010 to Guo H; Grant No. 32300275 to Hao D; Grant No. 31870254 to Li W), New Cornerstone Science Foundation (Grant No. NCI202235 to Guo H), and Shenzhen Science and Technology Program (Grant Nos KQTD20190929173906742, ZDSYS20230626091659010 to Guo H; Grant No. JCYJ20170817105503416 to Li W). Figures 2 to 4 were created in
https://BioRender.com .-
The authors confirm their contributions to the paper as follows: figure preparation, writing − draft manuscript preparation: Hao D, Li W; conceptual framework design, writing − manuscript revision: Hao D, Li W, Guo H. All authors reviewed and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Dongdong Hao, Wenyang Li
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Hao D, Li W, Guo H. 2025. Ethylene signaling in Arabidopsis: a journey from historical discoveries to modern insights. Plant Hormones 1: e014 doi: 10.48130/ph-0025-0015


|