









-
The phytohormone abscisic acid (ABA) functions as a master regulator of plant growth and stress adaptation, orchestrating the plant's responses to drought, salinity, and pathogen attacks, while also modulating developmental processes such as seed dormancy, stomatal closure, plant senescence, and fruit ripening[1−3]. The core of ABA signaling is the receptor Pyrabactin Resistance 1 (PYR1)/PYR1-Like (PYL)/Regulatory Component of ABA Receptor (RCAR) (PYR/PYL/RCAR) family, which function as molecular switches by binding ABA and initiating a signaling cascade via interacting with protein phosphatase type 2Cs (PP2CA) to relieve the inhibition of PP2C on Sucrose Nonfermenting 1 (Snf1)-related protein kinase 2s (SnRK2s)[4]. Since their landmark discovery in Arabidopsis, extensive studies have revealed their structural plasticity and functional diversification across plant species. PYR/PYL/RCAR receptors belong to a multi-gene family, with 14 members in Arabidopsis and ranging from eight to 23 putative members in crop species, such as soybean, rice, sweet orange, potato, and wheat[5,6]. Due to gene redundancy and difficulties in the generation of pyl multiple mutants, the functional landscape of PYR/PYL/RCARs remains incompletely illustrated. Therefore, conducting detailed phenotypic analyses across diverse biological contexts can elucidate the unique roles of PYR/PYL/RCARs, which is particularly important for comprehensively understanding ABA signaling regulatory mechanisms during normal plant growth and development, as well as stress response pathways.
PYR/PYL/RCAR receptors (PYR/PYLs) play a crucial role in ABA-dependent responses to abiotic stress. Lee et al.[7] demonstrated that AtPYL8 specifically mediates ABA-dependent drought tolerance, as indicated by PYL8-OX plants exhibited only slight damage under drought conditions, while PYL8-RNAi and WT were severely damaged. Notably, overexpression of AtPYL4 and AtPYL5 also enhanced drought resistance, antioxidant enzyme activity, and osmolyte levels[8]. In rice, OsPYL/RCAR5 positively regulated resistance to drought and salinity stress, but inhibited plant growth and reduced total seed yield[9], suggesting coupling of ABA-mediated growth suppression and stress adaptation. However, Yang et al.[10] showed that selective activation of ABA receptors such as RCAR6/PYL12 in Arabidopsis enhances drought resilience by optimizing water productivity, and maintaining high biomass accumulation via sustained CO2 assimilation gradients, without compromising intrinsic water use efficiency (WUE). Similarly, Miao et al.[11] used CRISPR/Cas9 technology to edit group I (PYL1-PYL6 and PYL12) and group II (PYL7–PYL11 and PYL13) PYL genes in rice and demonstrated that rice group I PYL receptors, particularly PYL1/4/6, play dominant roles in coordinating stomatal regulation, seed dormancy, and vegetative growth, with combinatorial mutants achieving enhanced grain yield under field conditions while preserving dormancy integrity. These findings highlight the evolutionary specialization of PYL subfamilies in balancing growth and stress adaptation, offering a targeted genetic strategy to improve crop productivity without compromising essential agronomic traits.
Emerging evidence highlights the role of PYR/PYL receptors in plant senescence, though their functional diversity across species and developmental stages remains incompletely illustrated. Zhao et al.[12] uncovered that PYL9-mediated ABA signaling accelerates drought-induced leaf senescence by activating SnRK2-dependent phosphorylation of ABF/RAV1 transcription factors, which subsequently upregulate senescence-associated genes independently of the ethylene pathway. The induced senescence generates osmotic gradients to prioritize water allocation to developing tissues, revealing an adaptive strategy where PYL9-enhanced senescence directly supports survival under extreme drought, positioning PYL receptors as key targets for engineering stress-resilient crops. Similarly, PYL8 acts as a critical modulator of glucose sensitivity and dark-induced senescence in Arabidopsis, where its overexpression amplifies Glc-dependent suppression of germination via upregulation of AtHXK1 and ABI5 while accelerating senescence through transcriptional activation of senescence-associated genes[13]. In Acer rubrum, ArNAC148 and ArPYR13 cooperatively regulate leaf senescence[14]. However, in our previous study, co-silencing of four ABA receptors SlRCAR9/11/12/13 fast-tracked fruit senescence, as indicated by accelerated water loss, softening, as well as reduced susceptibility to Botrytis cinerea in co-silenced lines during postharvest storage[15]. These findings indicate that PYR/PYL-mediated regulation of senescence displays species- and paralog-specific functional divergence, highlighting evolutionary diversification in ABA-responsiveness across plant lineages.
Tomato (Solanum lycopersicum L.), a member of the Solanaceae family, is a globally cultivated crop with significant economic value and a model system for floral and fruit development, offers a unique platform to investigate the interplay between ABA, GA, ethylene signaling, and developmental processes while addressing gaps in receptor functional characterization. Despite select PYR/PYL homologs have been functionally characterized, current investigations predominantly center on climacteric fruit ripening mechanisms, while largely overlooking PYR/PYL-mediated growth and senescence regulation in tomato, creating critical knowledge gaps in plant developmental biology. For instance, Zou et al.[15] reported that co-silencing of four PYLs suppressed fruit ripening progression by impairing key ethylene biosynthesis genes and downstream signaling components. Kai et al.[16] demonstrated that SlPYL9 accelerates fruit ripening through ABA-dependent modulation of core signaling components (SlPP2C1/2/9, SlSnRK2.8, and SlABF2), triggering transcriptional cascades that upregulate ethylene biosynthesis and cell wall remodeling enzymes. In this study, we identified and renamed 15 SlPYR/PYL homologs through evolutionary analysis. Tissue-specific expression dynamics (root, leaf, flower, fruit) were systematically characterized throughout developmental stages, with additional profiling under phytohormone treatments (ABA, ethylene) and abiotic stressors (drought, salinity). Furthermore, the functional significance of PYR1 and PYL8 in growth and senescence was elucidated. This lays a theoretical foundation for a comprehensive understanding of the roles of PYR/PYLs in vegetative and reproductive growth.
-
The nucleotide and amino acid sequence of the PYR/PYL genes in Arabidopsis were identified from NCBI (
www.ncbi.nlm.nih.gov/nucleotide ). Blastn was carried out to identify SlPYR/PYL family genes in tomato using the Tomato Genome database (http://solgenomics.net/tools/blast/ ). A phylogenetic tree based on the alignment of amino acid sequences of AtPYR/PYLs and SlPYR/PYLs was constructed using MEGAX software by the Neighbor-Joining method, with a bootstrap test replicated 1,000 times, the p-distance method, and pairwise deletion.Plant materials and growth conditions
-
Tomato plants (Solanum lycopersicum cv. Micro-Tom) were cultivated in a glasshouse at 25 °C/20 °C (day/night), a photoperiod of 16 h light and 8 h darkness, and 70% relative humidity. The plants were germinated and irrigated weekly with Hoagland's nutrient solution. For gene expression analysis, the tomato tissues, including roots (R), 4-week-old stems (S), and 4-week-old leaves (L), flowers (F) at the bud and anthesis stage, and fruits at the immature (IM), mature green (MG), breaker, yellow ripe (YF). and red ripe (RR) stages were harvested. Additionally, different floral organs, including sepals, petals, stamens, and pistils were harvested from flowers at the anthesis stage, respectively. All collected samples were immediately frozen in liquid nitrogen and stored at −80 °C until further use.
ABA, salt, and drought treatments on seedlings
-
One-month-old tomato seedlings were divided into four groups, including 18 plants in each group. Group I was used as control, group II and group III were treated with 100 μM ABA and 150 mM NaCl, respectively. The leaves were collected at 0, 24, and 72 h after treatment. Group IV was grown without watering for two weeks until wilting, and then the leaves were harvested. All collected samples were immediately frozen in liquid nitrogen and stored at −80 °C until gene expression analysis.
Plant hormone treatments on fruit development and ripening
-
Flower emasculation, artificial pollination, auxin (2,4-D), and GA3 treatments were carried out according to the methods in our previous study[17]. The ovaries were collected at 2 d ahead of anthesis (2 DAA), 4 d after artificial pollination (4 DPAP), 4 d after auxin and GA3 treatment (4 DPAT and 4 DPGT, respectively). For sample collection, three biological replicates were performed (approximately 10−20 ovaries in each replicate).
To perform ethylene treatment, the mature green tomato fruits (35 DPA) of uniform size were selected and divided into two groups. Group I fruit at harvest (day 0) without treatment was set as control. Group II fruit was put into containers and sprayed with 500 mg/L ethephon (pH 6.5). Then, the fruit samples were maintained at 25 °C, and collected at 72 h after treatment. For this experiment, three biological replicates with 10 fruit samples per replicate were performed. All samples were frozen in liquid N2 and stored at −80 °C until further analysis.
Real-time quantitative PCR (qRT-PCR) analysis
-
Total RNA from different tomato samples was extracted and purified using the RNeasy Plant Mini Kit and RNase-free DNase Set (QIAGEN, Germany) according to the manufacturer's instructions. CDNA was synthesized from 1 µg of total RNA by using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, USA). QRT-PCR was carried out using a Fast SYBR Mixture (CWBIO, Beijing, China) on a real-time thermal cycler (CFX96, Bio-Rad, USA). The program was set as follows: 95 °C incubation for 10 min, then 40 cycles of melting at 95 °C for 15 s, and annealing/extension at 60 °C for 60 s. Gene-specific primers of SlPYR/PYLs were designed using online software (
www.genscript.com.cn/tools/real-time-pcr-taqman-primer-design-tool ) and listed in Supplementary Table S1. For all experiments, three biological replicates were performed. Relative expression levels were calculated based on the 2−ΔΔCᴛ method using actin (tomato) as a reference gene.Subcellular localization of SlPYR1.1 and SlPYL8.1 proteins
-
To explore the subcellular localization, the open reading frame (ORF) sequence of SlPYR1.1 and SlPYL8.1 was fused in-frame with C-terminal of GFP into the pGreen vector under the control of CaMV 35S promoter. For transfection assay, protoplasts were obtained from suspension-cultured tobacco BY-2 cells. The cells were incubated in an orbital shaker (30−40 rpm) for 1 h at 37 °C in Tris-MES buffer containing 1% caylase, 0.2% pectolyase Y-23, and 1% bovine serum albumin (BSA). Then the protoplasts were filtered through nylon and washed by W5 buffer, and resuspended in MMg Buffer, then transfected using the PEG method as described by Xian et al.[18]. Briefly, 0.2 mL protoplast suspension (1 × 106 mL−1) was transfected with 50 μg of carrier DNA (salmon sperm DNA), 30 μg of 35S:SlPYR1.1-GFP/35S:SlPYL8.1-GFP and 40% PEG for 1 h. After centrifuging and suspending in W5, transfected protoplasts were incubated for at least 16 h at 25 °C. GFP fluorescence was observed by laser scanning confocal microscopy (Leica TCS SP5, Wetzlar, Germany). The protoplast transformed with 35S:GFP was set as control. All transient expression assays were repeated at least three times.
Overexpression of SlPYR1.1 and SlPYL8.1 in Micro-Tom tomato
-
To generate 35S:SlPYR1.1 and 35S:SlPYL8.1 transgenic plants, the coding sequence of these two genes was amplified from tomato cDNA using primers listed in Supplementary Table S1, and then cloned into the modified binary vector pLP100 carried a GUS reporter gene and kanamycin resistance gene under the CaMV 35S promoter. Then the plasmids pLP100-35S-SlPYR1.1 and pLP100-35S-SlPYL8.1 were transferred to Agrobacterium tumefaciens GV3101 for WT Micro-Tom tomato genetic transformation according to the methods in our previous study[15]. Briefly, 7-day-old cotyledons and hypocotyls were used as explants for transformation. After 1 d pre-culture and immersing in A. tumefaciens GV3101 harboring pLP100-35S-SlPYR1.1 and pLP100-35S-SlPYL8.1 (all the cultures were adjusted to OD600 = 0.1) for 20 min, the explants were successively transferred to co-culture medium, selection/differentiation medium, and root induction medium. The positive transgenic plants were identified by GUS staining and PCR detection. T2 generations of the transgenic lines were used for phenotype analysis. QRT-PCR was performed to detect the relative expression levels of SlPYR1.1 and SlPYL8.1 in transgenic plants. All plants were grown in a glasshouse under controlled conditions (16 h light/8 h dark cycle, 25 °C day/20 °C night, and RH 70%).
Phenotypic and molecular characterizations of SlPYR1.1-OX and SlPYL8.1-OX plants
Responses of transgenic plants to ABA
-
To determine the plant sensitivity to ABA, seeds of WT and transgenic tomatoes were sterilized and grown on half-strength Murashige and Skoog (MS) culture medium supplemented by 0, 1, 3, 5, and 10 μM ABA, respectively. After 7 d of cultivation under controlled conditions as mentioned, the germination rate and root length of the seedlings were illustrated and measured.
Phenotypic characterization
-
At least three independent transgenic tomato lines were used for phenotypic analysis. For SlPYR1.1-OX lines, the morphology parameters including plant height, length of stem internodes, and length of rachis were determined. For SlPYL8.1-OX lines, leaf size (width and length) was measured, and the inflorescence and flower development at different branches were observed.
QRT-PCR analysis
-
To explore the mechanism of dwarf in SlPYR1.1-OX lines, the mRNA levels of GID1, key gene related to GA signaling, in WT and SlPYR1.1-OX tomatoes were determined by qRT-PCR. The primers are listed in Supplementary Table S1.
Histological analysis
-
To examine the thickness of leaves in SlPYL8.1-OX lines, histological analysis was performed. Thirty-day-old leaves in WT and SlPYL8.1-OX plants were cut and immersed in Carnoy's Fluid for at least 24 h at 4°C. Then the samples were dehydrated in gradient ethanol, embedded in paraffin, sectioned, and stained with 0.05% toluidine blue. Finally, it was photographed by Leica DM3000 microscope (Leica, Wetzlar, Germany).
Determination of pigment content
-
The leaves and fruit of WT and SlPYL8.1-OX plants were sampled and subjected to pigment determination. A total of 0.2 g leaves was weighed, cut, and immersed in 95% (v/v) ethanol for 72 h at 4 °C in darkness, then the absorbance at wavelengths of 663, 645, and 470 nm were detected using an ultraviolet-visible spectrophotometer. The contents of chlorophyll a, b, and carotenoid were calculated using the formula by Beer-Lambert Law and Arnon. The method for lycopene extraction and determination was performed as follows. Briefly, approximately 0.5 g of the homogenized fruit sample was put into a 40 mL screw top amber glass vial containing 5 mL of 0.05% (w/v) butylated hydroxytoluene (BHT) in acetone, 5 mL of 95% ethanol, and 10 mL of hexane. Samples were extracted on a shaker at 180 rpm for 30 min on ice, and then 3 mL of ddH2O were added. The vials were then left at room temperature for phase separation and the upper (hexane) layer contained lycopene. The content of the total lycopene was obtained by measuring the absorbance of the solutions at 503 nm.
-
Based on the transcriptome data in our previous study[19], 15 PYR/PYL/RCAR family members were screened. The sequences of nucleotides and amino acids were obtained from version SL4.0 of the tomato genome (Supplementary Table S2). To avoid duplicate names, 15 SlPYR/PYLs were renamed according to the gene descriptions in the tomato genome. Based on the phylogenetic analysis, the SlPYR/PYL genes were divided into three subfamilies (Fig. 1). Subfamily I harbors the largest family members, including SlPYL4.1, SlPYL4.2, SlPYL4.3, SlPYL6.2, SlPYL6.3, and SlPYL6.4, which showed very close homology to Arabidopsis PYL4/PYL5/PYL6. Subfamily II harbors four SlPYR/PYLs, where SlPYR1.1 and SlPYR1.2 exhibited high amino acid homology with AtPYR1 and AtPYL1, while SlPYL2.1 and SlPYL2.2 displayed strong sequence homology with AtPYL2 and AtPYL3. In subfamily III, SlPYL8.1 shared close homology with AtPYL8 and AtPYL10, while three SlPYR/PYLs, including SlPYL8.2, SlPYL9.1, and SlPYL9.2, had close homology with AtPYL7 and AtPYL9. Additionally, SlPYL6.1 was found to be more closely related to AtPYL11/AtPYL12/AtPYL13 (Fig. 1).

Figure 1.
Phylogenetic analysis of the PYR/PYL proteins from tomato and Arabidopsis. A phylogenetic tree of 29 PYR/PYL proteins from tomato (15) and Arabidopsis (14) was constructed by using MEGAX based on the neighbor-joining method. These proteins were classified into three subfamilies as marked in the figure. The sequences used are listed in Supplementary Table S2.
Expression of SlPYR/PYL genes in response to hormones and abiotic stresses
-
To assess SlPYR/PYL expression responsiveness to ABA, tomato seedlings were treated with ABA for 72 h. Results showed that four genes, including SlPYR1.1, SlPYL4.3, SlPYL6.2, and SlPYL8.1, were remarkably down-regulated in response to ABA, while the transcription levels of SlPYL8.2 showed an upward and then a downward trend after ABA treatment (Fig. 2a). The SlPYR/PYL expressions respond to salinity are presented in Fig. 2b. The mRNA levels of SlPYR1.1 and SlPYL8.1 displayed significant declines in response to salt stress, whereas five SlPYR/PYLs, such as SlPYL4.1, SlPYL4.3, SlPYL6.2, SlPYL8.2, and SlPYL9.2, were obviously downregulated initially, but dramatically upregulated at 72 h after treatment. After 15 d drought stress, SlPYL4.3 exhibited an outstanding increase in its expression, while other SlPYR/PYL genes have no noticeable changes in their expressions (Fig. 2c). Compared with other genes in the SlPYL gene family (Fig. 2d), SlPYL9.1 and SlPYL9.2 were expressed at high levels throughout the process of fruit development and ripening, while two SlPYR/PYLs including SlPYL4.3 and SlPYL6.2 were consistently expressed at extremely low levels. During fruit maturation, the mRNA levels of four SlPYR/PYL genes (SlPYR1.1, SlPYL8.1, SlPYL8.2, and SlPYL9.1) initially decreased before increasing, reaching their minimum at the color-breaker stage (Br). In contrast, SlPYL9.2 expression progressively increased during fruit growth and ripening. Additionally, in response to ethylene treatment, the expressions of SlPYR1.1, SlPYR1.2, SlPYL6.2, and SlPYL8.1 were remarkably downregulated, while SlPYL4.1 and SlPYL8.2 showed significant upregulation (Fig. 2e). Based on the above-mentioned results, we hypothesized that SlPYR1.1 and SlPYL8.1 might play crucial roles during fruit maturation and responding to ABA, and ethylene, as well as salt stress.
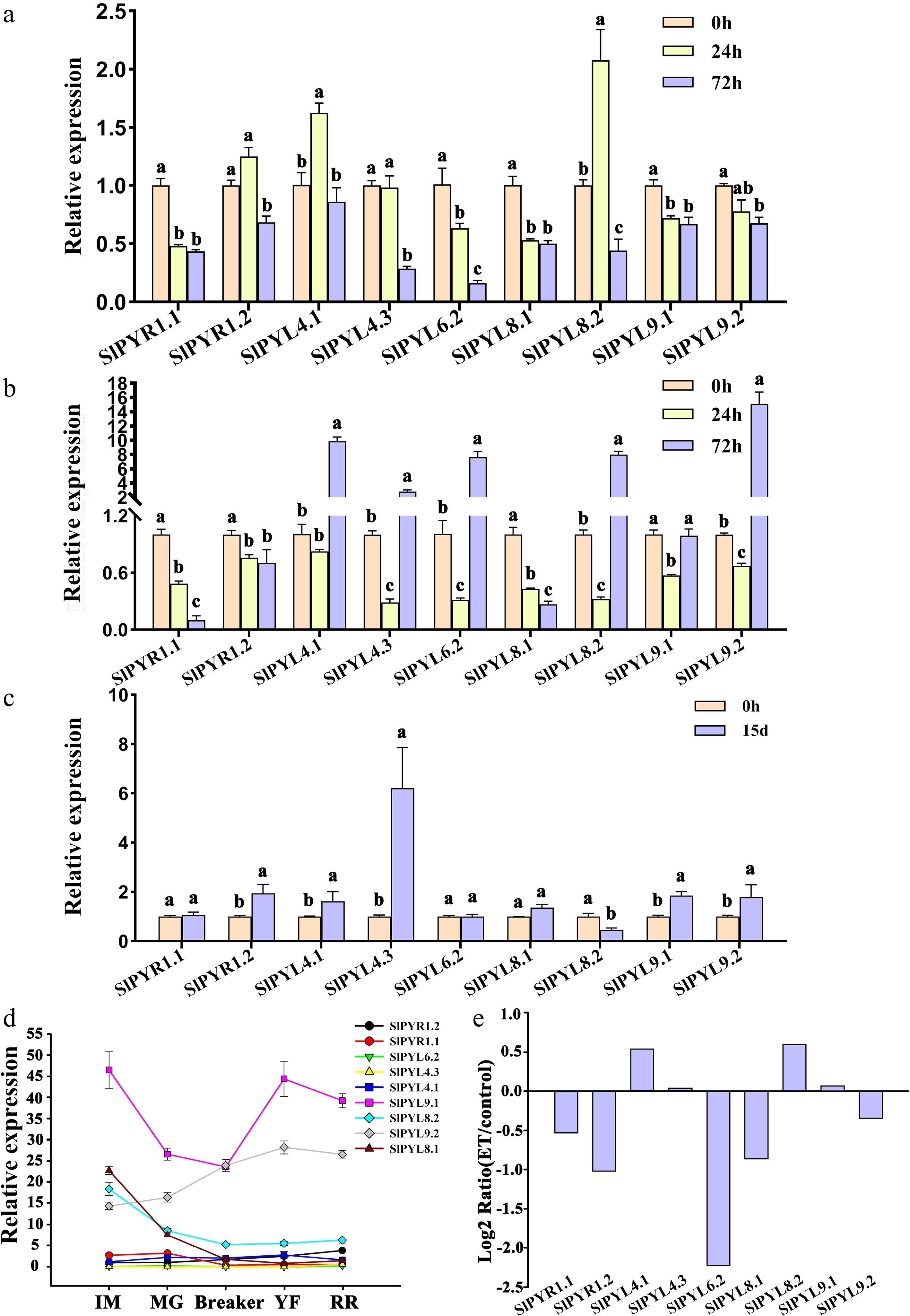
Figure 2.
Expression profiling of SlPYR/PYL genes responding to phytohormonal treatments and abiotic stresses in tomato determined by qRT-PCR. Expression patterns of SlPYR/PYLs at 0, 24, and 72 h after (a) ABA treatment, and (b) salt stress. (c) Expression of SlPYR/PYLs following 15-d progressive drought stress. (d) Expression of SlPYR/PYLs at different developmental stages during fruit maturation. IM, immature green (15 DPA). MG, mature green (35 DPA). BR, breaker (39 DPA). RR, red ripe (45 DPA). (e) Expression of SlPYR/PYLs in tomato fruit in response to exogenous ethylene (ET) application for 72 h. Data represents mean ± SEM of three biological replicates. Different lowercase letters indicate statistically significant differences (p < 0.05) between experimental groups.
Expression profiles and subcellular localization of SlPYR1.1 and SlPYL8.1 in tomato
-
To clarify their functions, the expressions of SlPYR1.1 and SlPYL8.1 in different tissues were analyzed by qRT-PCR. We found that SlPYR1.1 was expressed predominantly in roots, leaves, immature, and mature green fruits, while SlPYL8.1 displayed highest expression level in roots and leaves, and furthermore, their expression levels were dramatically downregulated following fruit ripening (Fig. 3a, e). In floral organs, SlPYR1.1 and SlPYL8.1 were dominantly expressed in sepals and stamens, respectively (Fig. 3b, f). During fruit set, SlPYR1.1 and SlPYL8.1 exhibited a remarkable upregulation 4 d after auxin treatment, but no obvious changes were observed in their expression after artificial pollination and gibberellin (GA3) treatment (Fig. 3c, g), indicating that these two genes might be involved in auxin-induced fruit setting in tomatoes. To determine their subcellular localization, GFP-fused SlPYR1.1 and SlPYL8.1 proteins were transiently expressed in Nicotiana benthamiana protoplasts. As shown in Fig. 3d and h, SlPYR1.1 and SlPYL8.1 localized to both the nucleus and cytoplasm, consistent with WoLF PSORT predictions.
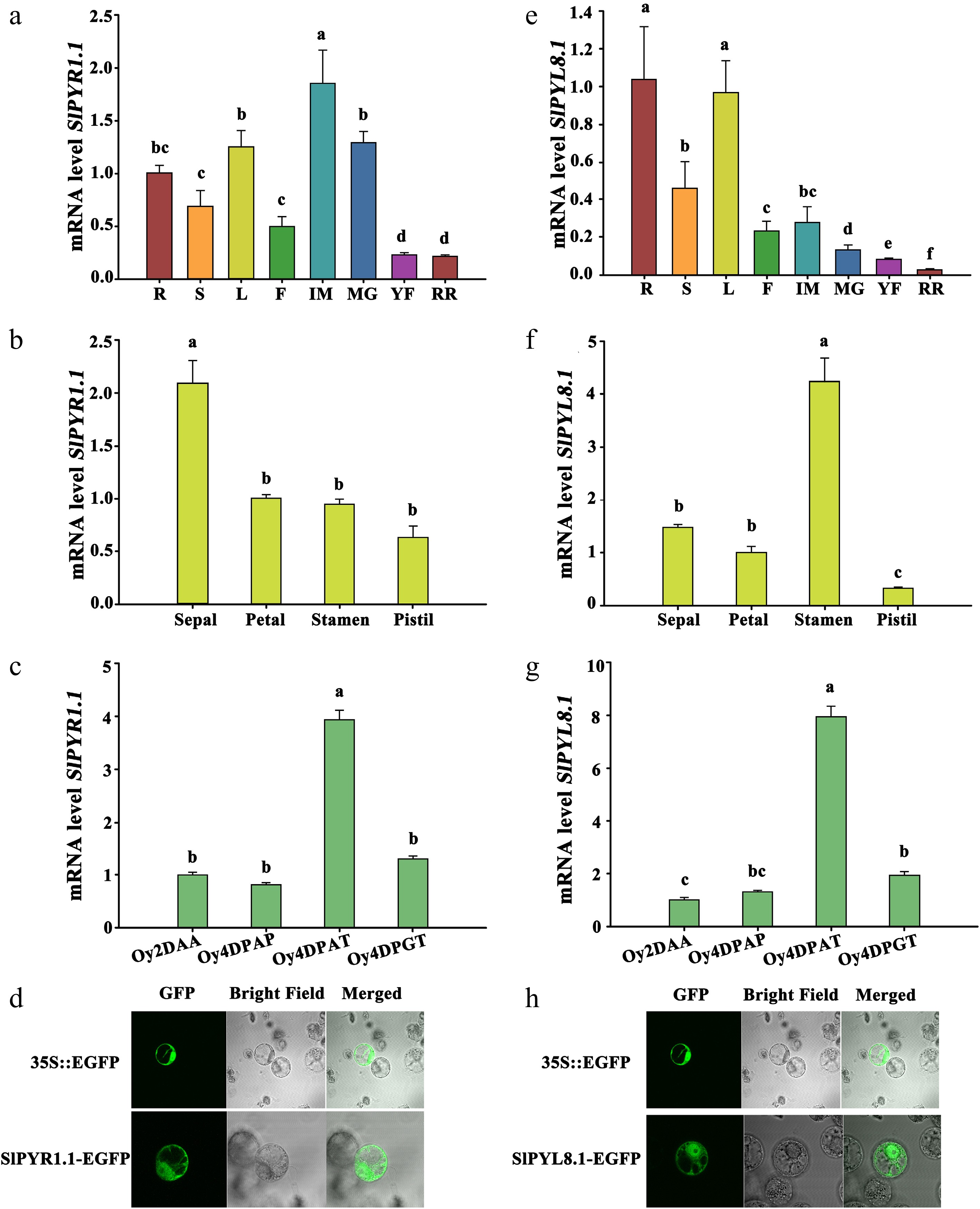
Figure 3.
Spatiotemporal expression and subcellular localization of SlPYR1.1 and SlPYL8.1. Expression patterns of (a) SlPYR1.1, and (e) SlPYL8.1 in various tissues/organs determined by qRT-PCR. R, S, L, F, IM, MG, YF, and RR indicate root, shoot, leaf, flower, and fruit at immature, mature green, yellow ripe, and red ripe stages, respectively. Expression profiles of (b) SlPYR1.1, and (f) SlPYL8.1 in four-whorled floral organs. Expressions of (c) SlPYR1.1, and (g) SlPYL8.1 in the ovaries at 2 d before anthesis (2 DAA), 4 d after pollination (4 DPAP), 2,4-dichlorophenoxyaceticacid (2,4-D) (4 DPAT), and GA3 application (4 DPGT). Data represents mean ± SEM of three biological replicates. Different lowercase letters indicate statistically significant differences (p < 0.05) between experimental groups. Subcellular localization of (d) SlPYR1.1, and (g) SlPYL8.1.
Overexpression of SlPYR1.1 leads to ABA hypersensitivity and plant dwarfism
-
To characterize the function of SlPYR1.1 in tomato, eight independent transgenic lines overexpressing SlPYR1.1 were generated. Among them, transcripts in three SlPYR1.1-OX lines were 12- to 20-fold higher than those in wild-type (WT) and were selected for phenotypic analysis (Fig. 4a). Here, the effect of ABA on seed germination and primary root growth in WT and transgenic plants was evaluated. Results showed that the seed germination rate (SGR) of SlPYR1.1-overexpressing plants was slightly lower than that of the WT on 1/2 MS medium. However, when 5 µM ABA was supplemented to MS medium, SGR of SlPYR1.1-OX plants was reduced to approximately 50% of the original rate, whereas SGR of WT was not affected. Additionally, compared with WT, ABA application dramatically inhibited the root growth and elongation of the transgenic plants (Fig. 4b−d; Supplementary Figs S1, S2). These findings indicate that SlPYR1.1 overexpression increased ABA sensitivity. We also found that overexpressing SlPYR1.1 in tomato led to a dwarf phenotype and delayed aging (Fig. 4e). The plant height, internode length, and rachis length were markedly declined in SlPYR1.1-OX lines, being approximately 70% of those in the WT (Fig. 4f−h). To elucidate the mechanism underlying the dwarfism of SlPYR1.1-OX plants, qRT-PCR analysis of GA signaling genes was conducted. The results demonstrated that SlGID1L mRNA levels were dramatically downregulated in three transgenic lines compared to those in WT (Fig. 4i), suggesting that GID1 might mediate SlPYR1.1-dependent regulation of plant height.
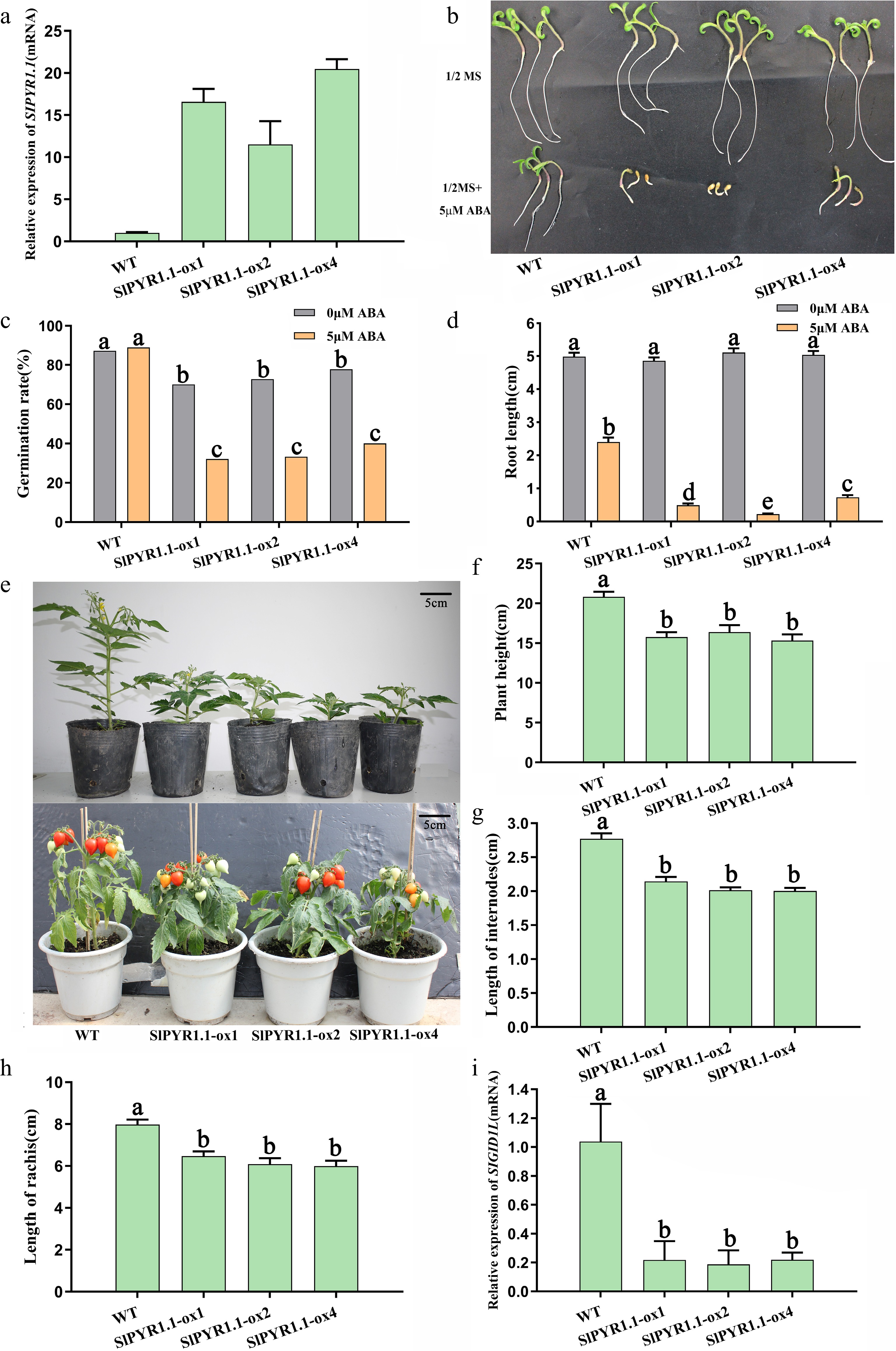
Figure 4.
Overexpression of SlPYR1.1 resulted in increased ABA sensitivity and plant dwarfism. (a) The relative mRNA level of SlPYR1.1 in three independent overexpression lines. (b) The seed germination and root growth of WT and transgenic plants on 1/2 MS medium supplemented with ABA. (c) Seed germination rate of WT and SlPYR1.1-OX plants under ABA treatment. (d) Root length of WT and SlPYR1.1-OX tomatoes under ABA treatment. (e) Phenotypic performance of transgenic plants across vegetative and reproductive phases. (f) Plant height, (g) length of internodes, and (h) rachis of WT and SlPYR1.1-OX tomatoes. (i) The relative mRNA level of SlGID1L in WT and SlPYR1.1-OX tomatoes. Data represents mean ± SEM of at least three biological replicates. Different lowercase letters indicate statistically significant differences (p < 0.05, one-way ANOVA with Tukey's HSD) among experimental groups.
Overexpression of SlPYL8.1 enhanced ABA sensitivity and delayed senescence
-
To address the functional significance of SlPYL8.1 in tomato, six independent transgenic lines overexpressing SlPYL8.1 were generated. Of them, three SlPYL8.1-OX lines exhibited 10- to 50-fold increases in their mRNA levels compared with that in WT (Fig. 5a). Consistent with SlPYR1.1-OX plants, overexpression of SlPYL8.1 led to a slight decline in seed germination rate (Fig. 5b). However, after ABA treatment, the seed germination rate and root elongation in SlPYL8.1-OX plants were strongly suppressed (Fig. 5c, d). It indicates that overexpression of SlPYL8.1 enhanced ABA sensitivity. Moreover, we observed that compared with the WT, the senescence of SlPYL8.1-OX tomato was delayed (Fig. 5e). During the fruit ripening stage, SlPYL8.1 overexpressing plants exhibited thickened, dark green, and upwardly curled leaves, whereas the leaves of the WT turned yellow (Fig. 5f). With respect to WT, the contents of chlorophyll a, chlorophyll b, and carotenoid were notably elevated in OX lines (Fig. 5g). Leaf size, including the length and width, was significantly increased in transgenic plants (Fig. 5h). Histological analysis demonstrated that in SlPYL8.1-overexpressing tomatoes, the number of mesophyll cells and cell layers increased significantly, while the size of epidermal and parenchyma cells in the veins was markedly larger compared to corresponding cells in WT leaves (Fig. 5i), which contributed to the increase in leaf size. Above all, we hypothesized that mesophyll cell proliferation in SlPYL8.1-OX tomatoes might lead to enhanced chlorophyll accumulation, thereby delaying leaf senescence.
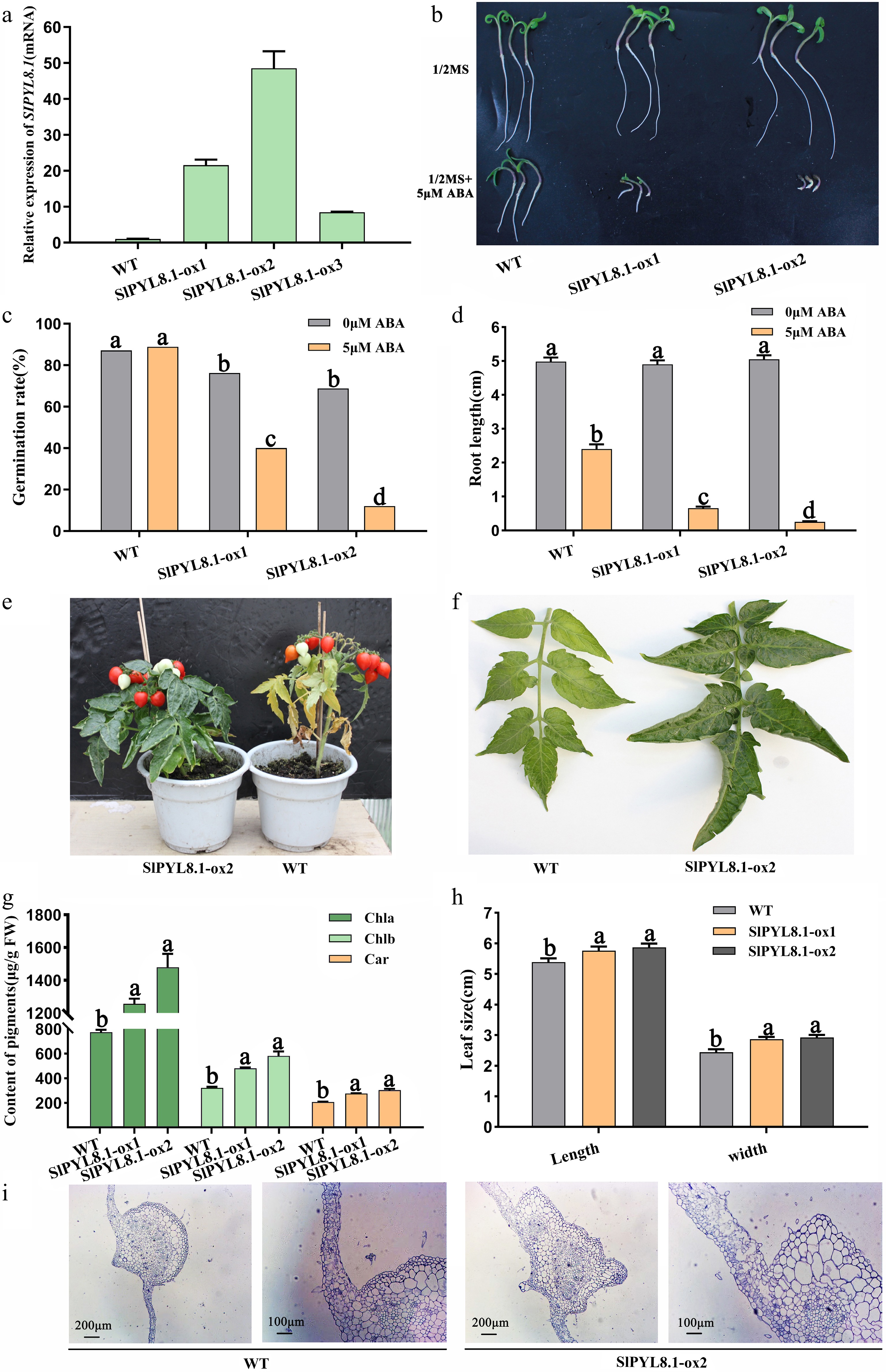
Figure 5.
Overexpression of SlPYL8.1 led to ABA hypersensitivity and delayed plant senescence. (a) The relative mRNA level of SlPYL8.1 in three independent overexpression lines. (b) The seed germination and root growth of WT and SlPYL8.1-OX tomatoes on medium supplemented with ABA. (c) Seed germination rate of WT and SlPYL8.1-OX tomatoes under ABA treatment. (d) Root length of WT and SlPYL8.1-OX lines under ABA treatment. (e) Overexpression of SlPYL8.1 led to delayed plant senescence. (f) Leaf architecture of WT and SlPYL8.1 overexpressing plants. (g) Pigment contents in WT and SlPYL8.1 overexpressing tomatoes. (h) Leaf size of WT and SlPYL8.1-OX plants. (i) Transverse leaf section showing the mesophyll cells and vascular bundle organization. Data represents mean ± SEM of at least three biological replicates. Different lowercase letters indicate statistically significant differences (p < 0.05, one-way ANOVA with Tukey's HSD) among experimental groups.
Overexpression of SlPYL8.1 alters inflorescence structure, fertility, and fruit pigment
-
Here, evidence showed that SlPYL8.1 participated in regulating reproductive growth in tomato. Generally, WT Micro-Tom plants exhibited normal flowering and fruit set in the first, second, and third inflorescences, whereas reduced fruit set occurred in higher-order inflorescences. However, in SlPYL8.1-OX plants, flowers in the first inflorescence frequently failed to open, as did the first flower in the first and second inflorescence, while flower buds aborted in higher-order inflorescences (Fig. 6a). Additionally, we observed that SlPYL8.1 overexpression caused abnormal inflorescence stem bending. In WT plants, inflorescence stems exhibited plagiotropic growth, forming a ~90° angle with the main stem, whereas inflorescence stems in transgenic lines displayed orthogravitropic growth and parallel alignment with the main stem (Fig. 6a & b). Moreover, it was noticed that WT fruit peel developed an orange-red color at full maturity stage, whereas SlPYL8.1-OX fruits exhibited a deep red coloration (Fig. 6c). Fruit flesh in tomato with SlPYL8.1 overexpressed exhibited significantly higher lycopene content relative to that in WT (Fig. 6d). It indicates that SlPYL8.1 can promote lycopene accumulation and thus modulate fruit color and maturation.
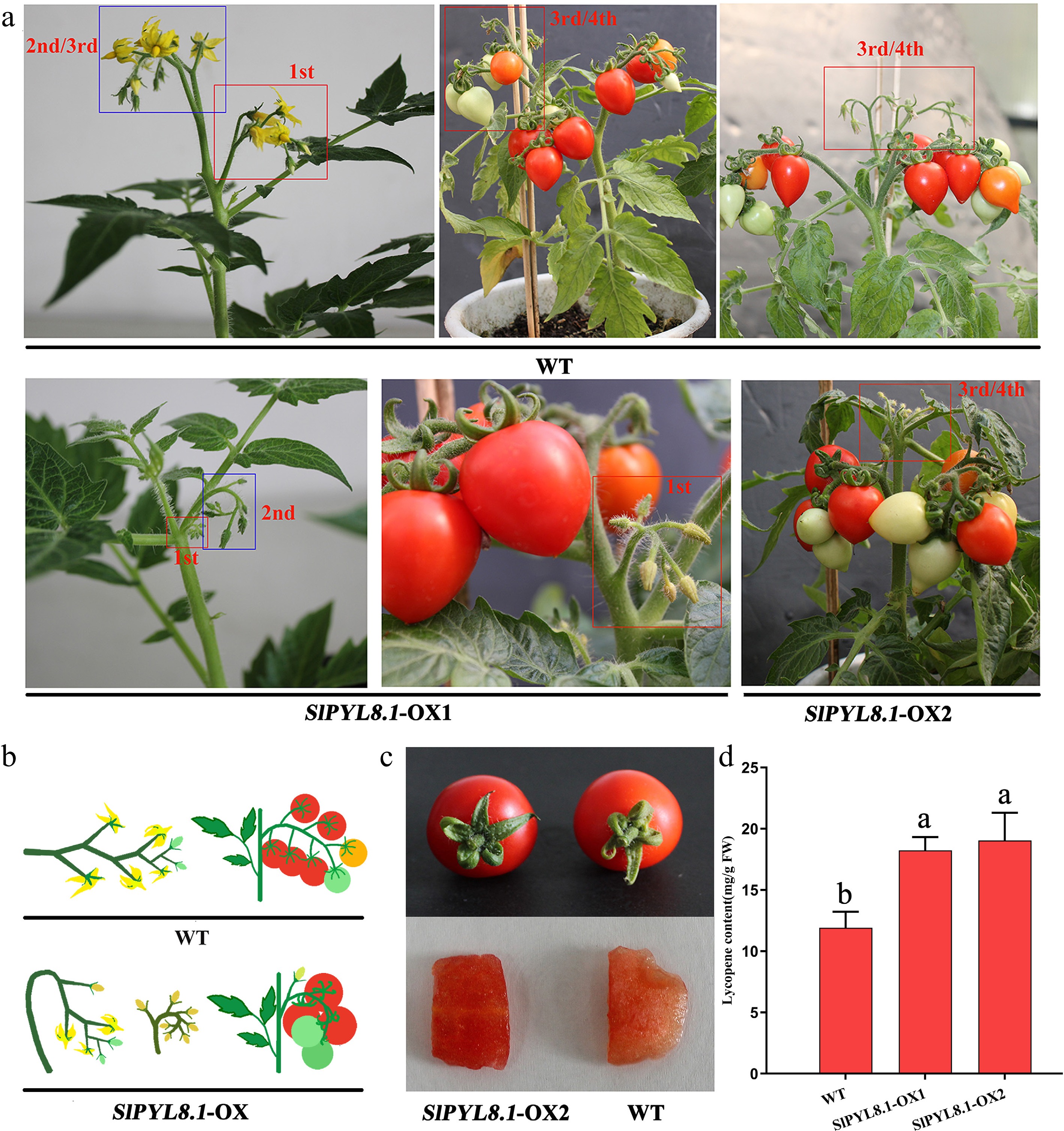
Figure 6.
SlPYL8.1 overexpression modified inflorescence architecture and altered fruit pigmentation. (a) SlPYL8.1 overexpression led to flower bud abortion. (b) Schematic diagram of inflorescence structures of WT and transgenic plants. (c) SlPYL8.1 overexpression resulted in alterations in fruit flesh color. (d) Lycopene content of WT and SlPYL8.1-OX plants. Data represents mean ± SEM of at least three biological replicates. Different lowercase letters indicate statistically significant differences (p < 0.05, one-way ANOVA with Tukey's HSD) among experimental groups.
-
The ABA receptor PYR/PYL family plays a pivotal role in mediating ABA-dependent plant stress responses and developmental regulation. To date, PYR/PYL members have been identified in many plant species, including 14 genes in Arabidopsis[20], 13 genes in rice[21], 14 genes in cucumber[22], nine genes in grape[23], seven genes in strawberry[24], and 14 genes in luffa[25]. In tomato, eight and 14 PYR/PYLs were identified according to Sun et al.[26] and Kai et al.[16], respectively. González-Guzmán et al.[27] discovered 15 PYR/PYLs based on the tomato genome. However, these genes were only recorded by numerical IDs. Previously, 15 receptors were designated as SlRCAR1 to SlRCAR15[15]. Herein, we have renamed these receptor genes according to their phylogenetic relationships with the orthologs in Arabidopsis (Fig. 1). These genes were divided into three subfamilies by using phylogenetic analysis, which is consistent with the study by Kai et al.[16]. In this study, to comprehensively uncover the significance, the responses of these ABA receptors to ABA, ethylene, salt and drought stress, developmental processes, as well as the roles of PYR1.1 and PYL8.1 in vegetative and reproductive growth were characterized.
SlPYR1 and SlPYL8 negatively regulate seed germination and plant height
-
ABA functions as the dominant phytohormone restraining seed germination initiation. Previous studies demonstrated that ABA-induced suppression of seed germination was mainly mediated by ABA receptors PYR1 and PYL1 in Arabidopsis[4,28−30]. Simultaneous knockout of PYR1, PYL1, PYL2 and PYL4 eliminated the inhibition of ABA on seed germination[31]. Zhao et al.[32] reported that PYL12 positively regulates ABA-mediated seed dormancy. In the presence of ABA, pyl12 mutant displayed less sensitivity to ABA and a higher seed germination rate than WT, while PYL12-overexpressing lines exhibited ABA hypersensitivity and a significant delay in seed germination. In accordance with previous studies, we observed that overexpressing SlPYR1.1 (the tomato homolog of AtPYR1 and AtPYL1) enhanced sensitivity to exogenous ABA while slightly reducing seed germination rates. When ABA was present, overexpression lines exhibited significantly reduced seed germination rates and shorter root lengths relative to WT tomato (Fig. 4b−d; Supplementary Fig. S2). Similarly, overexpression of AtPYL8/10 homolog SlPYL8.1 resulted in remarkably impaired germination capacity and stunted root elongation. In soybean, multiple knockouts of AtPYL8 homologs GmPYL17 and GmPYL19-1 by CRISPR/Cas9, also exhibited less susceptible to ABA and dramatically higher seed germination rate and root length than those in WT under 10 μM ABA treatment[33]. These findings indicate that functional conservation among angiosperm PYL orthologs and across tomato PYL subfamilies, evidenced by shared capacities to enhance ABA sensitivity, suppress seed germination, and inhibit root elongation. This provides critical insights into the evolutionary retention of core ABA signaling modules.
Evidence showed that ABA orchestrates plant dwarfing. Liu et al.[34] linked dwarfism in pear '601D' to endogenous ABA overaccumulation, which suppresses internode elongation via ABA signaling pathways and bHLH/WRKY transcription factors. In rice, CRISPR/Cas9-based editing of multiple PYLs, particularly pyl1/4/6, displayed significantly enhanced growth vigor compared to WT during seedling development, with increased shoot length and fresh weight, demonstrating that PYL1, PYL4, and PYL6 act as growth suppressors in rice[11]. In accordance with this, dwarf phenotype was observed in SlPYR1.1 overexpressing tomatoes, with declined plant height and stem internodes (Fig. 4f, g). Zhang et al.[33] demonstrated that in gmpyl17/19-1 mutants, plant height was higher than the WT. However, overexpression of their homolog SlPYL8.1 exhibited a slight decrease in plant height (Fig. 5e). Notably, heterologous overexpression of StPYL8-like, and AtPYL8/AtPYL9 homolog in potato, led to increased plant height, leaf number, and fresh weight in tobacco[35]. These results suggest functional diversification of PYL8 in plant growth across plant species. As is well known, GAs play a pivotal role in plant height control. ABA and GA antagonistically mediate plant development regulation. Overexpression of ABA-INSENSITIVE 4 (ABI4) exhibited a decline in plant height. Further analysis revealed that ABI4 regulates plant dwarfing via transcriptionally activating the ABA biosynthesis gene NCED6 and GA catabolism gene GA2ox7, thereby reducing GA/ABA ratio and inhibiting plant growth[36]. To date, many dwarfism-associated genes functionally linked to GA metabolism and signaling pathway have been molecularly characterized, including GA-INSENSITIVE DWARF1 (GID1), F-box GA-INSENSITIVE DWARF2 (GID2), slender rice1 (SLR1), dwarf1 (D1), spindly (SPY), and OsFBK4[37,38]. In this study, overexpression of SlPYR1.1 in tomato caused a strong downregulation of SlGID1 (Fig. 4i), indicating the regulation of SlPYR1.1 on dwarf might be mediated by GA receptor GID1. These findings also suggest that SlPYR1.1 functions as a central regulatory nexus mediating the antagonistic crosstalk between ABA and GA signals in tomato.
The involvement of SlPYR1 and SlPYL8 in pigment accumulation and senescence retardation
-
In an ABA-deficient tomato mutant high-pigment 3 (hp3), chlorophyll and carotenoid exhibited excessive accumulation[39], indicating the involvement of ABA in modulating pigment production. Jia et al.[40] demonstrated that ABA can activate MdABI5-MdMYBS1 transcriptional cascade to coordinate fruit carotenoid biosynthesis by directly upregulating carotenoid biosynthetic genes (MdPSY2-1, MdLCYb) in apple. Current studies also showed the pivotal role of PYLs in ABA mediated pigment accumulation. Overexpression of persimmon DkPYL3 (AtPYL7/8/9 homolog) in tomato resulted in a remarkable increase in chlorophyll levels in young fruit by elevating the transcripts of chlorophyll synthesis related genes. Additionally, DkPYL3-OX lines exhibited earlier ripening, and higher concentrations of total carotenoids, lycopene, and β-carotene in fully ripe fruits compared to those in WT[41]. Jia et al.[42] demonstrated that overexpressing apricot PaPYL9 (AtPYL7/9 homolog) led to a remarkable upregulation of GGPS and PSY, key enzymes involved in lycopene production. Consistent with these findings, SlPYL8.1-overexpressing plants accumulated elevated chlorophyll a/b, carotenoids in leaves during reproduction, and higher levels of fruit lycopene relative to those in WT (Figs 5 and 6). In our previous study, during postharvest storage of tomatoes under both ambient and moderate-low temperatures, PYL8.1 expression was positively correlated with lycopene content and PSY (Solyc02g081330.2) expression, but negatively correlated with LCY (Solyc04g040190.1) expression (Supplementary Fig. S3)[19]. These data establish SlPYL8.1 as a positive regulator of pigment biosynthesis, likely mediated through PSY/LCY modulation. Notably, contrasting with prior reports, our study revealed a metabolic uncoupling between carotenoid accumulation and fruit ripening phenotype, suggesting evolutionary subfunctionalization within these orthologs likely mediated by species-specific regulatory divergence.
It is well known that augmented ABA orchestrates plant senescence acceleration[43]. However, in this study, PYR1.1- and PYL8.1-overexpressing lines displayed delayed leaf senescence, as evidenced by transgenic foliage maintaining green compared to WT yellowing leaves during fruit maturation phases (Figs 4, 5). We hypothesized that SlPYL8.1 may counterbalance this by promoting cell division and chloroplast maintenance. Enlarged parenchyma cells and thickened leaves (increased palisade tissue thickness) in SlPYL8.1-OX plants (Fig. 5i) suggest enhanced photosynthetic capacity, which antagonizes senescence. This is in accordance with the results presented by Miret et al.[44] demonstrating that postharvest application of ABA significantly delayed leaf senescence during room-temperature storage by preserving chloroplast ultrastructure, enhancing cellular turgor pressure, and maintaining membrane integrity. In rice, SlPYL8 ortholog OsPYL3 overexpression resulted in significantly higher total chlorophyll, chl a, and chl b content compared to WT under drought and cold stress[45]. Similarly in Arabidopsis, after 20 d of drought treatment, the WT plants had withered and died, while the pRD29A::PYL9 transgenic lines remained vigorous[12]. These findings indicate that PYL8 and its homologs exhibit functional conservation in mitigating abiotic stresses and developmentally induced senescence processes.
SlPYL8 promotes floral bud dormancy and abortion during tomato reproductive growth
-
To date, the core role of ABA in regulating bud dormancy has been widely confirmed in numerous physiological, genetic, and molecular studies in higher plants. In pear, transcriptional upregulation of ABA biosynthetic genes (PpNCED2, PpNCED3), and higher ABA content were observed during flower bud endodormancy establishment[46]. Elevated ABA levels suppress the chromatin remodeler PKL (PICKLE), relieving its repression on dormancy master regulators DAM (dormancy-associated MADS-box) and SVP/SVL transcription factors. SVL directly activates callose Synthase 1 (CALS1) and represses FT, resulting in bud dormancy. Concurrently, ABA signaling via SnRK2-ABI5 phosphorylation cascades upregulates CALS1 expression, driving callose deposition at plasmodesmata. This callose barrier blocks intercellular communication, contributing to endodormancy establishment[47]. In this study, SlPYL8.1 overexpressing plants exhibited persistent dormant floral buds in both primary and higher-order inflorescences, characterized by loss of normal flowering ability and subsequent abortion (Fig. 6). Similar phenotypes were observed in the study by Huang et al.[48], demonstrating that heat stress (HS) induced tomato floral abortion on the primary inflorescence during transitional meristem and floral meristem developmental stages through HS-triggered ROS burst. In rice, HS-elevated ABA in developing anthers accelerated programmed cell death (PCD) initiation, subsequently leading to abnormal tapetum degradation and pollen abortion through ROS overaccumulation[49]. Therefore, we proposed that flower bud abortion in SlPYL8.1 overexpressing tomato might be attributed to PCD induction triggered by enhanced ABA signaling. Cao et al.[50] also demonstrated that PCD contributed to lotus flower bud abortion. However, ABA played an important role in alleviating flower bud abortion by suppressing PCD-related NnSnRK1, a positive regulator terminating the flowering processes in lotus.
These findings reveal an evolutionary trade-off, while ABA signaling enhances stress tolerance, its over-activation sacrifices reproductive success by hijacking developmental trajectories. Future research should elucidate the spatiotemporal dynamics of ABA receptor complexes coordinating the transition from dormancy to sterility. Engineering ABA signaling nodes could uncouple stress adaptation from yield loss in crops.
-
In summary, 15 tomato SlPYR/PYL genes were identified, renamed, and divided into three subfamilies. Spatial-temporal expression patterns revealed SlPYR1.1 and SlPYL8.1 displayed significant modulation by ABA, ethylene, and salt stress as well as fruit maturation phases, demonstrating its multifunctional integration of hormonal crosstalk, stress adaptation, and developmental transitions. Transgenic lines overexpressing these two genes exhibited hypersensitivity to ABA, dwarfism, and delayed senescence. SlPYR1.1 primarily governed GA-dependent vegetative growth and thereby inducing plant dwarfing via SlGID1L suppression. SlPYL8.1 delayed leaf senescence mediated by maintaining Chla level and chloroplast function. Additionally, SlPYL8.1 controlled reproductive plasticity, including inflorescence patterning, flower bud abortion, and fruit lycopene biosynthesis. These findings provide insights into how ABA receptors balance the priorities of growth and defense, offering genetic targets for breeding crops with optimized yield structures and enhanced stress resistance. The ability to enhance fruit pigmentation through manipulation of SlPYL8.1 was confirmed, highlighting its utility for targeted enhancement of horticultural traits.
This work was financially supported by Chongqing talent program (cstc2024ycjh-bgzxm0105), the Chongqing Natural Science Foundation Innovation and Development Joint Fund Project (CSTB2023NSCQ-LZX0146).
-
The authors confirm their contributions to the paper as follows: conceptualization: Tang N, Chen Z; methodology: Li Q, Wu P; software: Yang Z, Mashu H, Liu Z; validation: Tang N, Deng J; formal analysis: Wu P, Guo J; investigation: Li Q, Yang Z; resources: Mashu H; data curation: Li Q; writing − original draft: Li Q, Tang N; writing − review and editing: Wang N, Chen Z; visualization: Yang Z; supervision: Chen Z; funding acquisition: Tang N. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest. The funders have no role in study design, data collection, data analysis, data interpretation, or writing of the manuscript.
- Supplementary Table S1 The primers used in this study.
- Supplementary Table S2 The amino acid sequences of PYR/PYLs from Arabidopsis and tomato used in phylogenetic tree construction.
- Supplementary Fig. S1 Seed germination and primary root growth in wild-type tomato under ABA gradients range from 0 to 10 μm.
- Supplementary Fig. S2 Seed germination of SlPYR1.1-OX and SlPYL8.1-OX tomatoes under 5 μm ABA treatment.
- Supplementary Fig. S3 Expression patterns of SlPYL8.1, PSY and LCY during postharvest storage of tomatoes under both ambient and moderate-low temperatures (RT and MT).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Q, Wu P, Yang Z, Mashu H, Liu Z, et al. 2025. Overexpression of SlPYR1 and SlPYL8 confers ABA hypersensitivity, plant dwarfing, and delayed senescence in tomato. Plant Hormones 1: e013 doi: 10.48130/ph-0025-0013 

Overexpression of SlPYR1 and SlPYL8 confers ABA hypersensitivity, plant dwarfing, and delayed senescence in tomato
- Received: 07 May 2025
- Revised: 02 June 2025
- Accepted: 04 June 2025
- Published online: 08 July 2025
Abstract: Abscisic acid (ABA) functions as a master regulator in plant developmental processes and stress responses through signal transduction components. Extensive mechanistic dissection of PYR/PYL receptors has centered on stress adaption and fruit ripening. However, the roles of ABA receptors in regulating vegetative growth and floral organogenesis remain poorly characterized. Herein, 15 SlPYR/PYLs were identified and classified into three subfamilies in tomato. Expression profiling revealed that SlPYR1.1 and SlPYL8.1 were significantly downregulated under ABA, salt stress, and ethylene treatment while showing tissue-specific dominance in roots/leaves and dynamic reduction during fruit ripening. Subcellular localization confirmed their dual nuclear and cytoplasmic distribution. Functional characterization of overexpression lines demonstrated ABA hypersensitive phenotypes, including declined seed germination rate and root elongation in the presence of ABA. SlPYR1.1-overexpressing transgenics exhibited plant dwarfing, which might be attributed to SlGID1 downregulation, while SlPYL8.1-OX lines showed enhanced chlorophyll retention and mesophyll cell proliferation, thereby delaying leaf senescence. Notably, SlPYL8.1-OX tomatoes displayed reproductive abnormalities, including inflorescence stem orthogravitropism, and floral bud abortion in first- and higher-order inflorescences. In addition, SlPYL8.1 led to lycopene overaccumulation in fruit. These findings establish SlPYR1.1 and SlPYL8.1 as pivotal regulators of ABA-mediated growth-defense trade-offs, integrating phytohormone signaling with developmental plasticity in vegetative and reproductive organs.
-
Key words:
- Tomato /
- SlPYR1 /
- SlPYL8 /
- ABA hypersensitivity /
- Plant dwarfism /
- Leaf senescence