









-
Pigs raised in intensive farming in the present pig industry are more productive, but they are also more susceptible to oxidative stress[1]. Post-weaning piglets are more susceptible to oxidative stress than pigs at earlier development stages due to a variety of reasons, including immunological stress, weaning stress, and dietary, environmental, and social difficulties[2]. Oxidative stress often results in poorer growth performance and a range of illnesses, leading to significant financial losses for the pig industry[3]. It has been reported that early weaning may damage intestinal barrier function, impair tight junctions, and reduce the activity of digestive enzymes of weaned piglets[4,5]. The use of antioxidant feed additives has been proposed as a nutritional strategy to enhance the development and overall health of piglets.
Sulforaphane (SFN) is one of the main glucosinolates found in cruciferous vegetables and is most abundant in broccoli and Brussels sprouts[6]. SFN is a small molecule whose molecular formula is C6H11NOS2, with a melting point between 58.6 to 91.2 °C and a molecular weight of 177.28 g/mol[7]. Due to its small molecular weight and relatively high lipophilicity, SFN can be rapidly absorbed by intestinal cells in the jejunum after oral administration[8,9]. SFN is crucial in promoting health, which is related to its ability to induce phase II detoxification enzymes, up to 14 times higher than other potent phytochemicals such as quercetin[10]. Antioxidant response element (ARE) is a promoter sequence that controls phase II enzyme production. Nf-E2-associated factor 2 (Nrf2), isolated by its cytoplasmic companion, Kelch-like ECH-associated protein 1 (Keap1), modulates the activity of the ARE promoter by promoting its ubiquitination and destruction by the proteasome[11]. In addition, SFN has the highest bioavailability among well-known phytochemical antioxidants, such as curcumin (80 times higher)[12], and quercetin (20-fold higher)[13]. Because of that, SFN has tremendous potential for use as a pharmaceutical to treat specific disease states or as a nutraceutical to improve health status[10]. Later, Bonnesen et al.[14] reported that SFN and other isothiocyanates have preventative effects on colon carcinogenesis by promoting apoptosis and strengthening cell defense against chemicals that cause gene toxicity. Since then, several in vitro and in vivo studies have been conducted on the role of SFN, including clinical trials, providing relevant data for disease prevention and treatment, such as pancreatic cancer[15], liver cancer[16], breast cancer[17], lymphomas[18], diffuse axonal injury[19], prostate cancer[20], and leukemia[21]. In addition, SFN can prevent aging and neurodegeneration[22] and protect from gastric ulcers[23]. Studies in vivo also observed the inhibitory effects of SFN on HDACs[24−26]. These findings suggest that SFN uptake reduces the number of lung, breast, and prostate cancers while increasing histone acetylation and decreasing HDAC activity[24,25]. In individuals receiving SFN-rich broccoli sprouts, histones H3 and H4 acetylation is primarily attributable to the inhibition of HDAC activity in circulating peripheral blood mononuclear cells (PBMCs)[26]. Glutathione peroxidase 4 (GPX4) is a crucial enzyme for inhibiting ferroptosis because it can convert harmful lipid peroxides into harmless associated alcohol[27]. Currently, the exploration of upstream regulators of GPX4 would facilitate new approaches for alleviating ileum resuscitation.
Histone modification may be impacted by oxidative stress in the ileum, which could affect gut health. According to studies, oxidative stress can modify histones in different ways, including changing the expression of histone methyltransferases, causing active DNA demethylation, and influencing the levels of histone deacetylases and histone acetyltransferases[28]. The location where methylation occurs may lead to inhibition or transcriptional activation. The histone H3 methyllysine 4 (H3K4me1/2/3) is enriched on the transcriptional start sites and gene promotors of activity and development regulatory genes[29]. The distal enhancers and promoters of transcriptionally active genes are linked to histone H3 acetyl-lysine 9 (H3K9ac) and acetyl-lysine 27 (H3K27ac)[30]. While histone H3 trimethylation at lysines 9 (H3K9me3) or 27 (H3K27me3) is a component of several gene-repressive pathways[31]. In response to oxidative stress, posttranslational modification of histone H3 is modified by epigenetic enzyme regulation, such as reduced HDAC activity[32]. ROS also regulates histone methylation, including inhibitory PTMs (H3K9me2, H3K9me3, H3K27me2, and H3K27me3) and activation PTMs (H3K4me2 and H3K4me3), which vary according to cell needs[33]. A study by Wang et al. on the porcine intestinal epithelial cells revealed that oxidative stress induced the acetylation of H4K16ac and H3K9ac and the loss of H3K4me3[34]. When VRK1 is depleted, these oxidative stress-induced modifications change, leading to increases in H3K9me3 and H3K4me3 levels and losses of H4K16ac and H3K9ac. As aforementioned, weaning piglets disrupt the oxidative balance in the ileum, leading to various health issues that can be alleviated through histone modification. We can restore oxidation balance and improve the intestinal health of weaning piglets by adding bioactive chemicals and using plant production to stimulate histone modification. Therefore, we hypothesized that SFN would activate anti-oxidative genes through histone modification to alleviate oxidative stress in the ileum of weaning piglets. This study aims to explore how SFN supplementation affects the expression of anti-oxidative genes in the ileum of weaning piglets and to clarify the underlying epigenetic mechanisms, with a special emphasis on histone modifications that may mediate these modifications.
-
Twelve piglets of (Duroc × [Landrace × Yorkshire]) crossbreed, with an average weight of 15 ± 0.5 kg, were selected on the 28th d post-weaning. The piglets were randomly allocated into two groups: one group was fed a basal corn-soybean meal diet (control group, CON), while the other group received the same diet supplemented with 1 g/kg of sulforaphane (SFN). The experiment lasted for one month. Throughout this period, all piglets had ad libitum access to feed and water, and the room temperature was maintained between 24 to 26 °C with relative humidity controlled at 60% to 70%.
Intestinal morphology
-
At the end of the experiment, piglets were sampled for slaughter per replicate, the small intestines of those were disemboweled and approximately 2 cm segments from the middle part of the ileum were excised. The collected segments were flushed with physiological saline, fixed in a 10% neutralized formalin solution, dehydrated, and embedded in paraffin. A section of 5-μm from each sample was cut, inserted on a glass slide, stained with hematoxylin and eosin (H&E), and used for some histomorphological measurements under a light microscope. The measured parameters included villus height (VH), villus width (VW), crypt depth (CD), and villus height to crypt depth ratio (VH/CD). The villus height was measured from the apex of the villus to the junction of the villus and crypt, while the crypt depth was measured from the base of the crypt upward to the region of transition between the crypt and villus.
RNA extraction and real-time PCR
-
The ileum tissues from piglets were utilized to the total RNA was isolated using Trizol (Invitrogen, Waltham, MA, USA) according to the manufacturer's instructions and stored at −80 °C. The quantity and purity of the extracted RNA were assessed via a protein-nucleic acid analysis instrument (ND-2000UV, Thermo Fisher, Waltham, USA) and confirmed through 1% agarose gel electrophoresis. Subsequently, the RNA was converted into cDNA using the transcript All-in-One First-Strand cDNA Synthesis Super MIX for qPCR (QIAGEN, Frankfurt, Germany). The reverse transcription mixture consisted of 0.5 μg of total RNA, 5 μL of 5 × TransScript All-in-one SuperMix for qPCR, 0.5 μL of gDNA Remover, and nuclease-free H2O was adjusted to a total volume of 10 μL. The reverse transcription was carried out at 42 °C for 15 min, followed by 85 °C for 5 s. Post-transcription, 90 μL of nuclease-free H2O was added to the mixture, then stored at −20 °C. Real-time PCR was performed using a LightCycler® 480 IIReal-time PCR Instrument (Roche, Basel, Switzerland) with a PCR efficiency ranging from 94% to 105%. The PCR reaction mixture (10 μL) included 1 μL of cDNA, 5 μL of 2 × PerfectStartTM Green qPCR SuperMix, 0.2 μL of forward primer, 0.2 μL of reverse primer, and 3.6 μL of nuclease-free water. The reactions were conducted in 384-well optical plates (Roche, Basel, Switzerland) with an initial denaturation at 94 °C for 30 s, followed by 45 cycles of 94 °C for 5 s and 60 °C for 30 s. A melting curve analysis was performed post-PCR to ensure the specificity of the PCR product. Each sample was analyzed in triplicate. Additionally, qRT-PCR was conducted using an ABI StepOne Plus Real-Time PCR System (Applied Biosystems, CA, USA) with AceQ® qPCR SYBR Green Master Mix (Vazyme, Nanjing, China). The mRNA expression levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and quantified using the 2−ΔΔCt method.
GSEA analysis of RNA-seq
-
To identify enriched GO terms among differentially expressed genes (DEGs), GSEA software (version 4.1.0) was used. Furthermore, statistically enriched biological processes or pathways of DEGs were ranked and classified according to GO and KEGG pathways using the Metascape database (
http://metascape.org/ ). KEGG pathway plots, volcano plots, and Venn diagrams were generated via an online data analysis and visualization platform (http://www.bioinformatics.com.cn ).ChIP-qPCR
-
The ileum tissues underwent crosslinking in a 1% formaldehyde solution for 5 min, followed by quenching with glycine for 5 min on ice. Subsequently, the samples were pelleted via centrifugation and resuspended in a lysis buffer containing 50 mM HEPES (pH 8.0), 140 mM NaCl, 1 mM EDTA, 10% glycerol, 0.5% NP40, and 0.25% Triton X100. The pellets were then resuspended in a 10 mM Tris (pH 8.0) washing buffer, 1 mM EDTA, 0.5 mM EGTA, and 200 mM NaCl. After washing, the samples were resuspended in a shearing buffer (0.1% SDS, 1 mM EDTA, pH 8, 10 mM Tris-HCl, pH 8) and sonicated according to the manufacturer's instructions. Chromatin fragments were precipitated using specific antibodies and protein G beads, then washed and treated with proteinase K and RNase A. The purified chromatin immunoprecipitation (ChIP) DNA was subsequently used for chromatin immunoprecipitation quantitative real-time PCR (ChIP-qPCR) analysis. The antibodies utilized in the ChIP-qPCR assay were as follows: PPARα (Millipore, MA, USA; MAB3890); H3K27ac (Abcam, Cambridge, UK; ab4729); H3K18ac (Abcam, Cambridge, UK; ab1191); H3K4me2 (Abcam, Cambridge, UK; ab32356); H3K18la (Abcam, Cambridge, UK; ab134047); H3K4me1 (Abcam, Cambridge, UK; ab8895). Each experimental point was performed in triplicate, and the entire experiment was repeated three times to ensure reliability.
Statistical analysis
-
All data analysis was processed with GraphPad Prism software 9.0. Data are presented as mean values ± SEM. Statistical analysis was performed using two-tailed Student's t-tests. A p < 0.05 was considered significant.
-
No discernible variations in body weight were detected across the different treatment groups (see Fig. 1a). A statistically significant elevation (p < 0.05) in the VH in the ileum was noted in piglets that received SFN supplementation, as opposed to those in the CON group (refer to Fig. 1c). No significant disparities were observed in VW, CD, or VH/CD in the ileum of piglets across the various treatments (as shown in Fig. 1d−f). These findings imply that the addition of SFN confers advantages to the ileum of piglets without any apparent adverse effects.
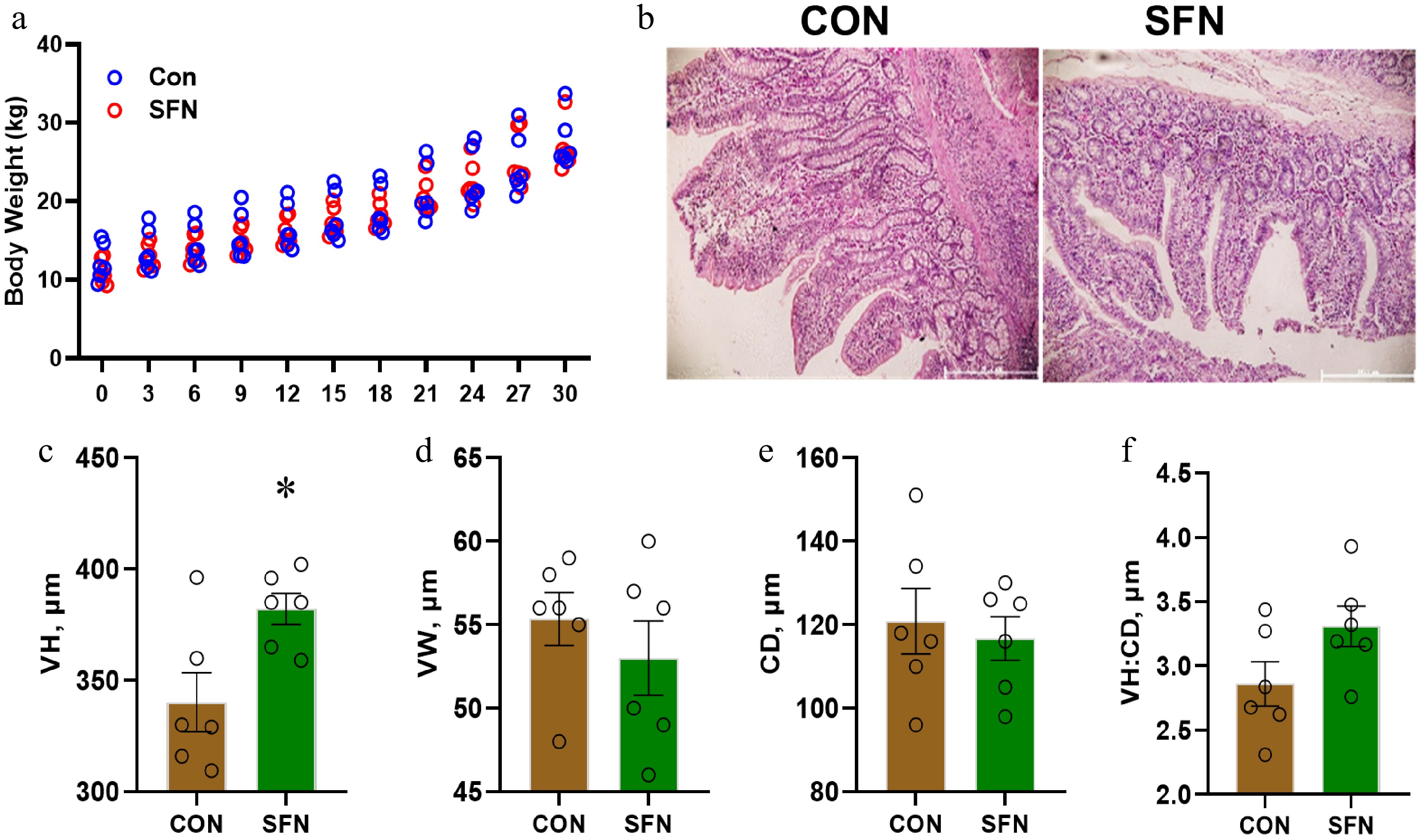
Figure 1.
This figure shows the effects of SFN supplementation on the ileum histomorphology of weaning piglets. (a) Body weight. (b) Representative histological image of the ileum. (c) Villus height (VH). (d) Villus width (VW). (e) Crypt depth (CD). (f) Villus height/Crypt depth (VH/CD). Data are presented as means ± SD. * p < 0.05.
Oxidative pathways in the ileum are sensitized to sulforaphane supplementation
-
To pinpoint the primary transcriptional pathways modulated by SFN, a transcriptome analysis was conducted on piglets' ileum tissues, comparing those supplemented with SFN to those without. Differentially expressed genes (DEGs) were identified in the SFN group compared to the CON group, with a log2 fold change greater than 1 and a p-value less than 0.05 (Fig. 2a). Further functional annotations of these transcripts are detailed in Fig. 2b and c. The Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of DEGs indicated that genes were predominantly enriched in peroxisome and lipid oxidation pathways. Gene Set Enrichment Analysis (GSEA) revealed that pathways related to oxidative phosphorylation, anti-oxidative stress, fatty acid β-oxidation, and cell-redox-homeostasis were significantly upregulated by SFN (Fig. 2d−g). These findings suggest that the oxidative pathways in the ileum are highly responsive to sulforaphane supplementation.
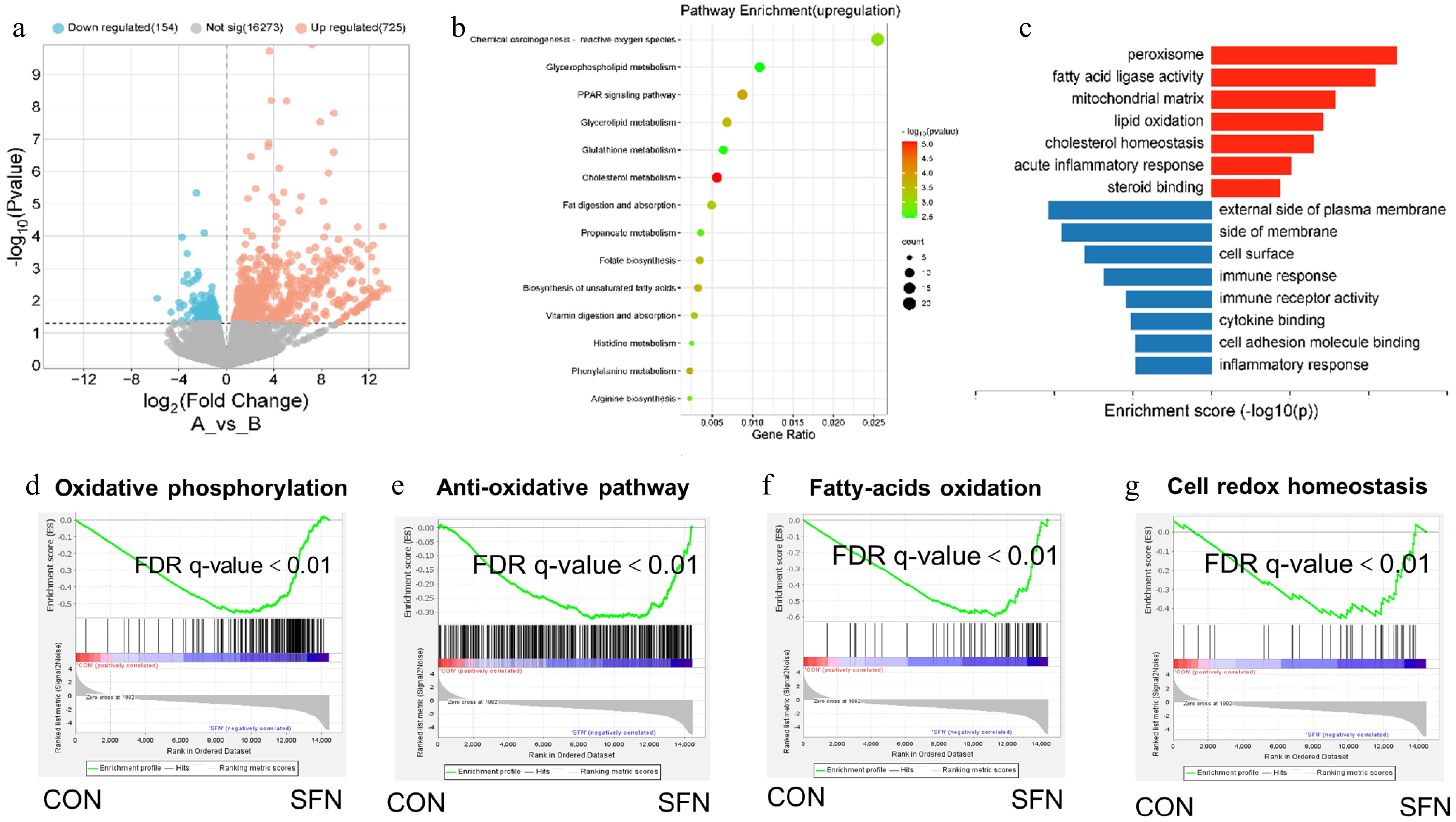
Figure 2.
(a) The differential gene expression profile between SFN and vector group was visualized by transcriptome analysis. (b) DEG involved in ROS and PPAR signal pathways is a rich enrichment of KEGG analysis. (c) Gene expression involved in the peroxisome and lipid oxidation pathways were among the most enriched pathways analyzed by gene ontology. (d) Oxidative phosphorylation. (e) Anti-oxidative pathway. (f) Fatty acid oxidation. (g) Cell-redox-homeostasis.
Sulforaphane enhances the anti-oxidative gene expressions in the ileum of piglets
-
In conjunction with GO and KEGG pathway enrichment analyses, a selection of pathway-specific genes revealed that the majority of pathways associated with oxidative phosphorylation, antioxidant stress, fatty acid β-oxidation, and cell-redox-homeostasis were notably upregulated in response to SFN (Fig. 3a−d). To further verify the expression pattern of DEGs, ileum-specific expression genes were measured by qRT-PCR (Fig. 3e). We investigated whether STRING-ELIXIR analysis supports the functions of PPARA, which is expected to be an important transcription factor interacting with essential proteins of lipid oxidation in SFN-fed pigs (Fig. 3f). Accordingly, 11 regulated DEGs, including NDUFB4, PRDX3, AIFM1, NDUFS8, GPX4, ETFDH, NDUFS2, IDH1, HTRA2, FXN, and LDHB in the anti-oxidative and oxidative phosphorylation pathways were identified by SFN supplementation in the Venn diagram (Fig. 3g). The preceding results indicate that SFN may influence oxidative pathways by regulating the transcription of particular genes associated with antioxidant functions.
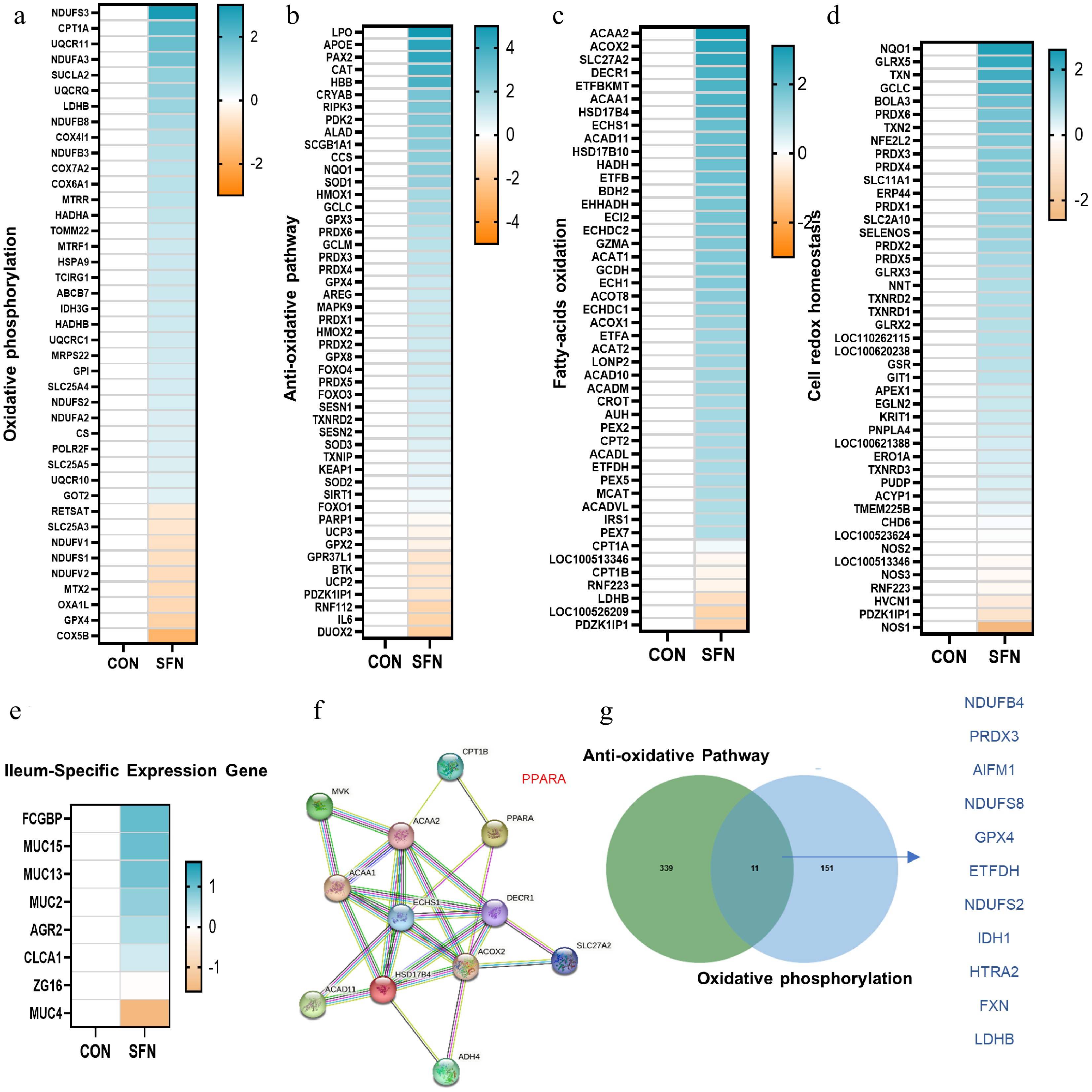
Figure 3.
(a)–(d) Heatmaps of mRNA expression (RNA-seq, log2 transformed) changes of oxidative phosphorylation, anti-oxidative stress, fatty acid β-oxidation, and cell-redox-homeostasis. (e) Heatmaps of mRNA expression (RNA-seq, log2 transformed) changes of the ileum-specific expression genes. (f) STRING-ELIXIR analysis predicts PPARA as a key transcript factor in interaction with lipid oxidation key proteins. (g) Venn diagram of the genes with significantly differential expression (log2 fold change < –0.5) shared by SFN vs control in anti-oxidative and oxidative phosphorylation pathways.
Histone modifications facilitate the transcriptional suppression of GPX4 and IDH1
-
In alignment with the heatmap depicting genes from the nuclear receptor superfamily (Fig. 4a) and their corresponding mRNA expression profiles (Fig. 4b), a marked increase in the mRNA expression of PPARα was evident in the SFN-supplemented group relative to the CON group (Fig. 4c). Furthermore, a significant upregulation in the mRNA expression levels of genes such as NDUFB4, PRDX3, AIFM1, NDUFS8, GPX4, ETFDH, NDUFS2, IDH1, HTRA2, and FXN was observed in piglets receiving SFN supplementation, contrasting with the CON group (Fig. 4d). Upon examining the ileum of piglets supplemented with SFN, a notable upregulation of PPARα activators was identified at the promoters of GPX4 and IDH1 (Fig. 4e). To further investigate, the presence of histone active markers H3K27ac, H3K18ac, H3K4me1, H3K4me2, and H3K18la at the GPX4 and IDH1 gene promoters was quantified using ChIP-qPCR analysis. The results indicated that SFN supplementation led to a significant enhancement in the binding of these histone markers at the GPX4 and IDH1 promoters compared to the CON group, as illustrated in Fig. 4f−j. These findings imply that SFN may exert its effects on oxidative pathways by influencing the transcriptional activity of genes central to antioxidant processes.
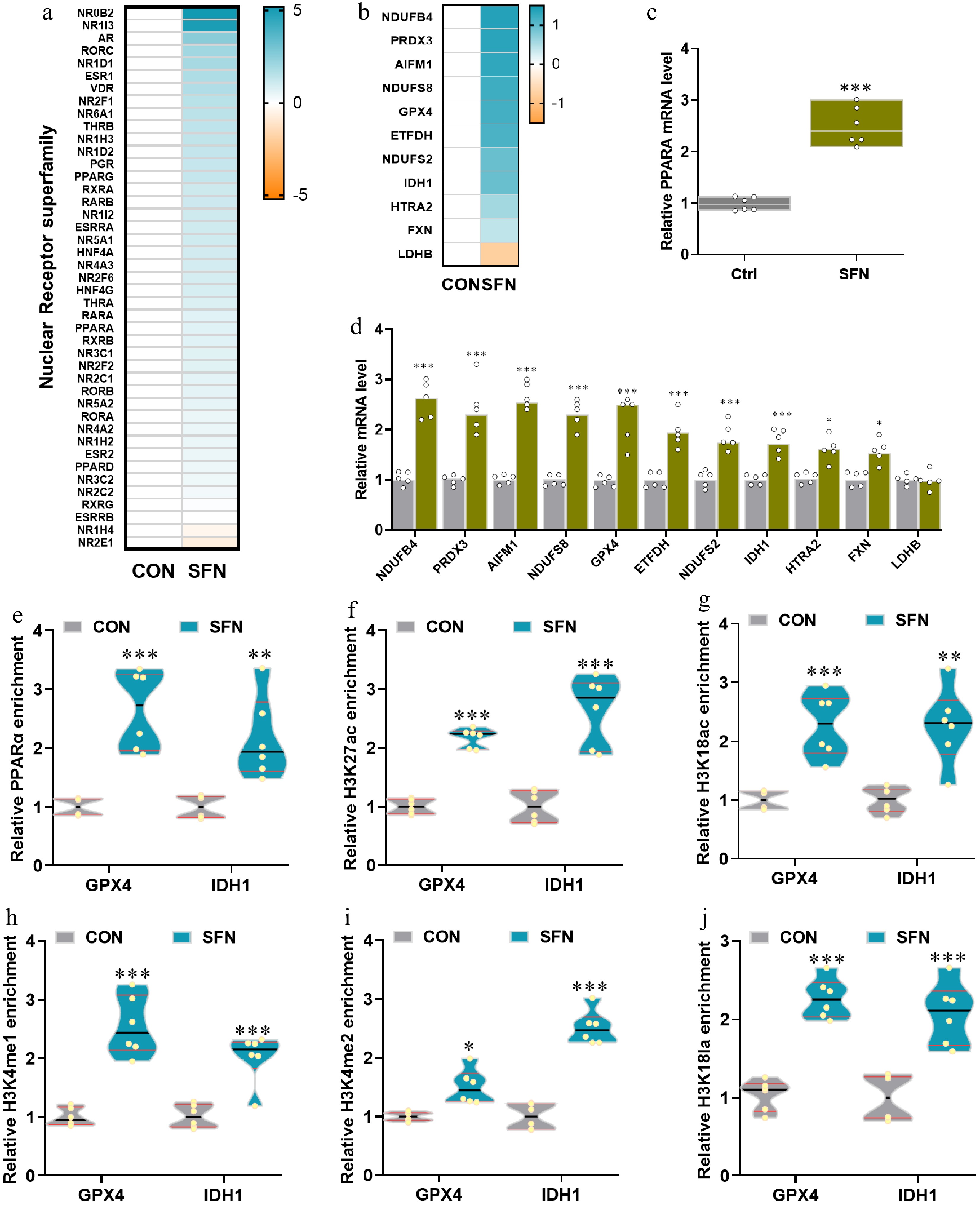
Figure 4.
(a), (b) Heatmaps of mRNA expression (RNA-seq, Log2 transformed) changes of the nuclear receptor superfamily and the genes with significantly differential expression shared by SFN vs control in anti-oxidative and oxidative phosphorylation pathways. (c), (d) qRT-PCR validations of the expression of PPARA and SFN-induced significance. (e)–(j) The relative enrichments of PPARA, H3K27ac, H3K18ac, H3K4me1, H3K4me2, and H3K18la at the key anti-oxidative genes GPX4 and IDH1 were analyzed by ChIP-qPCR. Data are presented as means ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.
-
According to recent research, SFN may have an impact on mitochondrial activity in addition to its well-established function of initiating the Nrf2 pathway[35]. Excessive SFN dosages have been shown to affect oxidative phosphorylation, bioenergetics, and mitochondrial dynamics, sometimes resulting in mitochondrial stress. There is evidence that SFN may change ATP synthesis, increase the formation of reactive oxygen species (ROS), or cause mitochondrial uncoupling without the involvement of Nrf2. In certain cell types or under particular circumstances, such effects may have therapeutic implications, but they may also result in cytotoxicity. SFN has been shown to affect the composition of the gut microbiota, favouring Lactobacillus and other helpful bacteria while perhaps suppressing pathogenic microorganisms. High SFN dosages, however, can disrupt microbial equilibrium and result in unexpected effects like dysbiosis or changes in the synthesis of short-chain fatty acids[36].
Piglets are particularly susceptible to oxidative stress, which leads to poor growth performance and intestinal damage. Sulforaphane can currently be sourced at a low cost from plants. This study investigated the effects of plant-derived SFN on growth performance and its potential beneficial roles against diquat-induced oxidative stress via histone modifications and ileum damage in piglets. We demonstrated that supplementation with plant-derived SFN improves growth performance under normal feeding conditions and effectively alleviates oxidative stress-related growth retardation and ileum injury by enhancing antioxidant capacity in piglets.
The intestine, as a site for nutrient digestion and absorption, is important for the development of intestinal morphology[37]. Intestinal villi extend from the tip to the base of the hilum, and their height is directly related to the rate of nutrient absorption in the small intestine[38]. As the height of the villi increases, the absorptive area of the small intestine becomes larger, thereby increasing digestion and absorption of nutrients[39]. The ability of pigs to absorb nutrients is based on the integrity of their intestinal morphology. The intestinal crypt refers to the vertical distance extending from the opening of the crypt to the base. A shallow CD is associated with a higher proliferation rate of intestinal cells[40]. The VH/CD ratio is a comprehensive index that reflects the nutrient absorption and digestion capacity of the small intestine in chickens. A low ratio of VH to CD indicates reduced digestive efficiency and intestinal mucosal damage in broilers, serving as a key indicator of decreased intestinal health. In this study, dietary SFN supplementation increased ileum villi height compared with CON, indicating improved intestinal function and maturation. Wang et al.[41] reported significant improvement in the villus height of piglets' ileum fed on a diet supplemented by phytogenic feed additives. Su et al.[42] reported an increase in the villus height of chickens fed on a diet supplemented by phytogenic feed additives.
The lack of selectively permeable cell membranes and environmental changes involving cell contents make it challenging to correlate data from intact cells with the homogeneity of these cells. Nevertheless, there is strong evidence that SFN penetrates normal tissues, although more readily in the latter, which accounts for the active transport of these substances[43]. It is, therefore, logical that the effects of SFN on oxidative phosphorylation, antioxidant stress, fatty acid β-oxidation, and cellular redox homeostasis reported in this study appear to be similar to the effects of SFN on these processes in intact ileum and liver tissues or cells prepared in vitro. If this hypothesis is correct, then it is clear that an increase in cellular oxidative phosphorylation in the presence of SFN is observed in porcine satellite cells[44], and rat ileum[45]. Excessive reactive oxygen species (ROS) during oxidative phosphorylation can inhibit the activity of histamin G, increase the expression of GAPDH, and cause damage to the microenvironment of glioblastoma[46], and significantly decrease the phosphorylation of the NF-κB/MAPK pathways in the ileum[47]. The high oxidative phosphorylation group also shows a more significant expression of immune checkpoint genes. These biological pathways may be responsible for these ileum tissues: elevated levels of oxidative phosphorylation may result in enhanced cellular metabolic activity, which may disrupt immune cell functions and cause immune dysfunction. In addition, this condition may help the ileum tissue evade immune system surveillance by influencing the expression of cell surface molecules. Lastly, increased oxidative phosphorylation activity can lead to the generation of reactive oxygen species, which can decrease DNA damage and result in genomic instability[48]. Oxidative stress and cellular redox homeostasis stem from an imbalance between oxidation and deoxidation in the body, resulting in elevated lipid peroxidation levels and reduced antioxidant enzyme activity[49]. Glutathione S-transferase is an important enzyme that reduces ROS and oxidative stress to detoxify the body[50]. According to reports, fatty acid oxidation plays a significant role in fatty acid catalysis when different fatty acid oxidase enzymes are present[51]. Fatty acid oxidation is often called fatty acid β-oxidation[51]. Fatty acids are divided into three types, namely medium chain, long chain, and short chain. Long-chain fatty acids produce fatty acyl-CoA through the action of long-chain acyl-CoA synthetase. Through CPT1, CPT2, and carnitine acylcarnitine translocase, fatty acyl CoA reaches the mitochondria, where it is subjected to fatty acid β-oxidation by fatty acid oxidase, the tricarboxylic acid cycle, and oxidative phosphorylation[52]. Notably, CPT1 is a significant enzyme in fatty acid β-oxidation, which controls the rate of fatty acid transfer to mitochondria during fatty acid β-oxidation. Fatty acid β-oxidation increases mitochondrial ROS production, and ROS accumulation activates the mitogen-activated protein kinase (MAPK) pathway[53]. The balance between the production and elimination of reactive oxygen species (ROS) and active nitrogen (RNS) in cells is known as cellular redox homeostasis. Aerobic metabolism produces reactive oxygen species (ROS) as by-products. Examples of ROS are superoxide anion (O2–), hydrogen peroxide (H2O2), and hydroxyl radicals (OH·). These compounds have inherent chemical properties that confer reactivity against various biological targets[54]. Both excess ROS/RNS and insufficient antioxidant defenses cause oxidative stress, which disrupts cellular components, including DNA, proteins, and lipids, the balance necessary for cell function and survival[55].
Peroxisome proliferator-activated receptors (PPARs) are ligand-induced transcription factors of the nuclear receptor (NR) superfamily. Three PPAR subtypes have been identified: PPARα (NR1C1), Pparβ/δ (NR1C2), and PPARγ (NR1C3)[56]. The synthesis and decomposition of triglycerides and lipid droplets, lipoprotein metabolism, gluconeogenesis, bile acid metabolism, and other metabolic pathways and genes are regulated by PPARα. Metabolic pathways and genes such as synthesis and decomposition of triglycerides and lipid droplets, lipoprotein metabolism, gluconeogenesis, and bile acid metabolism are regulated by PPARα[57]. In the present study, the mRNA expression of PPARα significantly increased in the SFN compared to the CON. NDUFB4, NDUFS2, and NDUFS8 (NADH: Ubiquinone Oxidoreductase Subunit B4, S2, and S8), expression in the ileum is necessary to preserve energy metabolism and mitochondrial function. The digestive system component ileum requires a lot of energy to perform its duties, such as nutrient absorption, peristalsis, and epithelial barrier maintenance[58]. NDUFB4 is a subunit of complex I that is critical for the production of ATP during oxidative phosphorylation of the mitochondrial electron transport chain[59]. Peroxorexin (PRDX3) protein reduces molecules that cause oxidative stress, such as ROS; for example, peroxides are essential metabolic intermediates and regulators of growth factor signaling while being a result of cellular stress[60]. PRDX-3 is associated with promoting the reduction of ROS, such as H2O2, generated during cellular metabolic processes such as ATP production. Mitochondria, the organelles that can regulate apoptosis, are a significant source of ROS generation[61]. Apoptosis-inducing factor mitochondrial Associated 1 (AIFM1) is a protein that regulates apoptosis or programmed cell death in addition to mitochondrial function[62]. It participates in energy synthesis and oxidative phosphorylation, which are essential for the high energy requirements of ileal cells for nutrient absorption and other metabolic functions[63]. When AIFM1 moves from the mitochondria to the nucleus in response to cellular stress or injury, it can cause apoptosis. This apoptotic process clears the ileum of unhealthy or dysfunctional cells, preserving tissue homeostasis[64]. Glutathione peroxidase 4 (GPX4) detoxifies lipid peroxides[65]. While hydrogen peroxide, alkyl hydrogen peroxide, and fatty acid hydroperoxides can all be broken down by glutathione peroxidase, only GPX4 can break down hydroperoxides present in lipoproteins and complex lipids, including cholesterol, cholesterol esters, and phospholipids[65]. The ileum of piglets is linked to many metabolic processes, including those of the weaning stage, in which isocitrate dehydrogenase 1 (IDH1) plays an important role. According to studies using single-cell mRNA sequencing, the ileum contains a variety of epithelial cell subtypes, including enterocytes, goblet cells, and enteroendocrine cells, each with unique gene expression patterns[66]. IDH1 is responsible for the oxidative decarboxylation of isocitrate to alpha-ketoglutarate, which results in the production of NADPH from NADP+; the citric acid cycle is one of the metabolic processes in cells that depends on this interaction[67]. In this study, dietary SFN supplementation has a significant positive effect on the relative expression levels of NDUFB4, PRDX3, AIFM1, NDUFS8, GPX4, ETFDH, NDUFS2, IDH1, HTRA2, and FXN mRNA in the ileum of weaned piglets. In conclusion, increased villus height and oxidative phosphorylation level of weaned piglets may be related to increased mRNA expression genes.
Further analysis showed that the enrichment degree of PPARα in the ileum of piglets was different among different treatments, consistent with the research results[56] in mice. We found that in the SFN-supplemented ileum, the activators of PPARα were significantly up-regulated at the GPX4 and IDH1 enhancers. Chemical changes in the histone proteins surrounding DNA are called histone modifications. These changes can influence gene expression through changes in chromatin structure that affect how accessible DNA is for transcription[65]. Posttranslational modifications of histones, including methylation, acetylation, phosphorylation, and ubiquitination, can alter DNA accessibility to transcription factors and other regulatory proteins and chromatin structure, which can affect gene expression[68]. The epigenetic report suggests that the ileum counteracts host epigenomic regulatory mechanisms by altering metabolic gene expression and promoting ileum replication and translocation[69]. Following the idea that H3K4me1 and H3K27ac, H3K9ac, and H3K18ac are primary modifiers of gene regulation during liver[70]. In this study, the dietary addition of SFN enhanced the concentration of H3K27ac, H3K18ac, H3K4me1, H3K4me2, and H3K18la on GPX4 and IDH1 in the ileum. An increase in p300 is responsible for these acetylation changes. Altered ileum expression of oxidative phosphorylation, anti-oxidative stress, fatty acid β-oxidation, and cell-redox-homeostasis genes may be explained by mechanisms other than PPARα-driven epigenetic control. However, epigenetic effector factors showed significant associations with most of the genes involved in these functions, which may be related to the small sample size of each group.
-
In summary, our study demonstrates that the administration of Sulforaphane significantly enhances the expression of genes associated with antioxidant functions in the ileum of weaned piglets, mediated through specific histone modifications. The upregulation of key genes related to metabolic pathways and the oxidative stress response, coupled with the observed increase in villus height, underscores the potential of Sulforaphane as a valuable dietary supplement for improving intestinal health and oxidative defense in young pigs. These results emphasize the importance of incorporating such phytochemicals into animal feeding strategies and expand our understanding of the molecular mechanisms by which plant-derived substances can modulate gene expression and support optimal growth and health in piglets.
-
All procedures were reviewed and preapproved by the Institutional Animal Care and Utilization Committee (IACUC) of the Animal Experimental Ethics Committee of Yangzhou University, identification number: No.202202160, approval date: 2022-02-24. The research followed the 'Replacement, Reduction, and Refinement' principles to minimize harm to animals. This article provides details on the housing conditions, care, and pain management for the animals, ensuring that the impact on the animals is minimized during the experiment.
This work was funded by the National Key R&D Program of China (2023YFD1301200), Jiangsu Provincial Double-Innovation Team Program (JSSCTD202147), Natural Science Foundation of Jiangsu Province (BK20220582, BK20210812), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
-
The authors confirm contribution to the paper as follows: writing – review and editing, project administration, and methodology: Wang S, Huang H; investigation: Zhu Q, Li K, Zhu C, Zhu CP, Ahmed AA, Yuan L, Liu HY, Kim IH; writing – original draft, methodology, investigation, data curation, and conceptualization: Muniyappan M; writing – review and editing, writing – original draft, methodology, investigation, and data curation: Cai D.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. The original transcriptome data proposed in this study has been preserved at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov), and the preservation number is PRJNA1187453.
-
The authors declare that they have no conflict of interests. Demin Cai is the Editorial Board member of Animal Advances who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
# Authors contributed equally: Shiqi Wang, Hao Huang
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang S, Huang H, Zhu Q, Li K, Zhu C, et al. 2025. Sulforaphane supplementation reprograms anti-oxidative genes via histone modifications in the ileum of weaning piglets. Animal Advances 2: e026 doi: 10.48130/animadv-0025-0023 

Sulforaphane supplementation reprograms anti-oxidative genes via histone modifications in the ileum of weaning piglets
- Received: 20 January 2025
- Revised: 24 April 2025
- Accepted: 20 May 2025
- Published online: 16 September 2025
Abstract: Currently, plant extracts are being extensively studied due to their anti-inflammatory and antioxidant properties. One is sulforaphane (SFN), an isothiocyanate found in cruciferous vegetables. This study aimed to determine the effects of SFN supplementation on anti-oxidation genes in the ileum of weaned piglets from the perspective of histone modification. A total of 12 piglets [Duroc × (Landrace × Yorkshire)] with an initial body weight of 15 ± 0.5 kg (weaned at d 28) were randomly divided into two groups (n = 6 of each): the control group (CON, basal diet) and the SFN treatment group (SFN 1 g/kg + basal diet). The experiment period lasted for 30 d. The results showed that compared to CON, SFN supplementation could significantly increase the villus height of piglets' ileum (p < 0.05) but had no significant effects on crypt depth, villus width, and VH/CD of piglets' ileum. Additionally, the analysis of GSEA revealed that SFN upregulated signatures involving oxidative phosphorylation, anti-oxidative stress, fatty acid β-oxidation, and cell redox homeostasis pathways compared to CON. In addition, an increase in the mRNA expression of PPARα and mRNA expression levels of anti-oxidative genes NDUFB4, PRDX3, AIFM1, NDUFS8, GPX4, ETFDH, NDUFS2, IDH1, HTRA2, and FXN were significantly up-regulated in piglets supplemented SFN compared to CON. Finally, compared with the CON group, the histone activity markers H3K27ac, H3K18ac, H3K4me1, H3K4me2, and H3K18ac, as well as the genes and GPX4 and IDH1 target site PPARα activators of the SFN supplement group, were significantly up-regulated. We conclude that SFN-supplemented reprograms anti-oxidative genes via histone modifications in the ileum of weaning piglets.
-
Key words:
- Sulforaphane /
- Histone modifications /
- Weaning piglets /
- Antioxidant /
- Epigenetics