










-
Yeasts are ubiquitously present[1]. There are hundreds of yeast species, yet merely a handful of yeasts are commercially utilized, such as Saccharomyces cerevisiae. Saccharomyces cerevisiae is used predominantly as a feed additive. Annual production of feed yeast has increased significantly from 80.8 thousand tons in 2013 to 208.6 thousand tons in 2022 in China. Globally, the feed yeast market was valued at 1.3 billion (USD) in 2022 with a projected annual growth rate of 4.83% to 2029 (
www.imarcgroup.com/yeast-market ).A vast array of forms of yeast and substances derived from yeast are used as feed additives which include viable yeasts, nutritional yeasts, yeast cell culture, and hydrolyzed yeast[2]. Yeast-derived postbiotic are produced by anaerobic solid-state fermentation of yeast cells under certain process conditions. The resulting yeast biomass contains multiple components, such as yeast cell metabolites, short-chain fatty acids[3], yeast cell walls (polysaccharides and proteins), proteases, nucleases, nucleotides, amino acids, peptides, minerals, and vitamins[4].
Interest in yeast-derived products for feeding pigs has increased in the last 20 years[5]. Viable yeast or autolyzed yeast lysate consistently improves growth performance, feed intake, feed efficiency, immunity, and gut health in weanling piglets[6−10]. In growing-finishing pigs, yeast-derived postbiotic, hydrolyzed yeast, or mannose supplementation positively impacted growth performance (based on 36 comparisons) and feed efficiency (based on 33 comparisons) in a meta-analysis of studies conducted from 1971 to 2021[5]. However, the functional mechanisms through which yeast-derived products influence the performance of growing-finishing pigs are not clear. Many factors such as feedstuff, dietary protein levels, temperatures, nutrient restriction, yeast form, stages of pig, and antibiotics were included in the analysis to elucidate potential mechanisms of action. Interestingly, improved feed efficiency occurred in high-fiber when yeast-derived products were included in diets[11−14]. These characteristics of yeast supplementation are useful as pig feed in China contains relatively low energy and uses fiber as part of energy feed instead due to the low cost of these diets. In addition, yeast-derived postbiotics usually impart antioxidant and anti-inflammatory effects that are beneficial to the gut health of pigs.
Therefore, in this study, yeast-derived postbiotics were evaluated in growing pigs fed diets with normal or reduced net energy for their effects on growth performance, feed conversion rate, antioxidant status, anti-inflammatory indices, and gut microbiota composition, and its mechanism was explored in vitro.
-
Crossbred growing pigs, 192 healthy Duroc × Landrace × Yorkshire (initial weights 30 ± 0.6 kg) were used in an 11-week feeding trial. A 2 × 2 factorial design was employed. The four diets were basal diet (CON); CON supplemented with 0.5 g/kg XPC Ultra diet (YC); CON with net energy (NE) reduced by 75 kcal/kg supplemented (RED75–); and RED75– with 0.5 g/kg XPC Ultra diet (RED75+; Table 1). XPC Ultra, yeast-derived postbiotic product, was kindly gifted by Diamond V (Cedar Rapids, IA, USA). It contained oligosaccharides, proteins (15.21% crude protein), peptides, amino acids, yeast-derived enzymes, nucleic acids, and other nutrients. All diets were provided in pellet form and formulated to meet nutrient requirement of Chinese pigs as described by China Agricultural University[15] on growing pigs.
Table 1. Ingredient composition and calculated nutrient composition of experimental diets (as-fed basis).
Item 30–50 kg BW 50–75 kg BW CON YC RED75− RED75+ CON YC RED75− RED75+ Ingredients (%) Corn 58.50 58.50 52.00 52.00 62.00 62.00 56.00 56.00 Soybean meal 13.00 13.00 10.95 10.95 9.30 9.30 7.55 7.55 Wheat bran 17.00 17.00 20.00 20.00 5.50 5.45 13.20 13.15 Corn DDGS 5.00 5.00 5.00 5.00 8.00 8.00 8.00 8.00 Rice bran meal 1.00 0.95 7.20 7.15 10.00 10.00 10.00 10.00 Wheat flour 2.00 2.00 2.00 2.00 2.00 2.00 2.00 2.00 Limestone 1.00 1.00 0.83 0.83 1.16 1.16 1.20 1.20 Dicalcium phosphate 0.90 0.90 0.82 0.82 0.60 0.60 0.55 0.55 Salt 0.35 0.35 0.35 0.35 0.35 0.35 0.35 0.35 L-lysine·HCl 0.440 0.440 0.468 0.468 0.376 0.376 0.414 0.414 DL-Methionine 0.093 0.093 0.097 0.097 0.032 0.032 0.038 0.038 Threonine 0.122 0.122 0.134 0.134 0.092 0.092 0.105 0.105 Tryptophan 0.030 0.030 0.036 0.036 0.025 0.025 0.028 0.028 Premix1 0.40 0.40 0.40 0.40 0.40 0.40 0.40 0.40 Choline chloride 0.08 0.08 0.08 0.08 0.08 0.08 0.08 0.08 Phytase 0.02 0.02 0.02 0.02 0.02 0.02 0.02 0.02 XPC Ultra 0.00 0.05 0.00 0.05 0.00 0.05 0.00 0.05 Mold inhibitor 0.05 0.05 0.05 0.05 0.05 0.05 0.05 0.05 Antioxidants 0.015 0.015 0.015 0.015 0.015 0.015 0.015 0.015 Total 100.00 100.00 100.00 100.00 100.00 100.00 100.00 100.00 Calculated nutrient levels (%)2 NE (kcal/kg) 2,325.00 2,325.00 2,250.00 2,250.00 2,325.00 2,325.00 2,250.00 2,250.00 Calcium 0.65 0.65 0.65 0.65 0.62 0.62 0.62 0.62 Total phosphorous 0.63 0.63 0.74 0.74 0.60 0.60 0.65 0.65 Available phosphorus 0.28 0.28 0.28 0.28 0.23 0.23 0.23 0.23 Crude protein 15.50 15.50 15.51 15.51 14.48 14.48 14.52 14.52 NDF 17.85 17.85 18.25 18.25 19.60 19.60 20.37 20.37 SID AA Isoleucine 0.51 0.51 0.49 0.49 0.46 0.46 0.45 0.45 Leucine 1.12 1.12 1.08 1.08 1.16 1.16 1.11 1.11 Lysine 0.92 0.92 0.92 0.92 0.80 0.80 0.80 0.80 Methionine 0.30 0.30 0.30 0.30 0.25 0.25 0.25 0.25 Methionine + cysteine 0.52 0.52 0.52 0.52 0.45 0.45 0.45 0.45 Threonine 0.55 0.55 0.55 0.55 0.49 0.49 0.49 0.49 Tryptophan 0.17 0.17 0.17 0.17 0.14 0.14 0.14 0.14 Valine 0.60 0.60 0.60 0.60 0.57 0.57 0.56 0.56 Experimental diets were a corn-soybean meal-based diet: (1) CON: basal diet with normal net energy meet with China National Feeding Standard of Swine (GB/T 39235-2020); (2) YC: CON supplemented with 0.5 g/kg yeast derived postbiotic XPC Ultra diet; (3) RED75–: diet with the net energy reduced by 75 kcal/kg (comparing with CON); (4) RED75+: RED75– supplemented with 0.5 g/kg XPC Ultra diet. BW: Body weight; NE: Net energy; ME: Metabolizable energy; NDF: Neutral detergent fiber; SID: Standardized ileal digestibility. 1 The premix provided the following per kilogram of the diet: vitamin A, 9,000 IU; vitamin D3, 3,000 IU; vitamin E, 24 IU; vitamin K3, 1.8 mg; vitamin B1, 2 mg; vitamin B1, 2.0 mg; vitamin B6, 3.0 mg; vitamin B2, 5.0 mg; vitamin B12, 0.1 mg; niacin, 40 mg; vitamin B5, 15 mg; folic acid, 1.0 mg; biotin, 0.05 mg; choline chloride, 500 mg; Fe, 80 mg; Cu, 20 mg; Zn, 90 mg; Mn, 80 mg; I, 0.35 mg; Se, 0.3 mg. 2 Gross energy, Crude protein, Calcium, Phosphorous NDF contents are measured values, while all others are calculated values. During Phase I and Phase II (on d 34 and d 69, respectively), 32 pigs from each dietary treatment were selected for blood collection via vena puncture in the morning after 12 h of fasting (Supplementary File 1). During Phase I and Phase II (d 35 and d 70), 16 pigs from each treatment were selected randomly for feces collection for the determination of microbiota. The fecal samples were collected by anal stimulation to capture a pea-sized subsample of feces. Fecal samples were transferred aseptically to a sterile storage tube and placed on ice immediately. Within minutes, storage tubes were placed in liquid nitrogen to snap-freeze them before transferring them to a freezer (−80 °C) for storage until processed for microbiota sequencing.
Nutrient digestibility
-
The fecal and feed samples were analyzed for dry matter (DM), crude protein (CP), neutral detergent fiber (NDF), Ash, Ca, and P[16−18]. The apparent total tract digestibility of DM, N, and energy was calculated according to the method of Stein et al.[19] (Supplementary File 1).
Serum biochemistry
-
Sample pretreatment: Blood was collected from the porcine anterior vena cava via a vacuum tube. After gentle inversion (5–8 times) to mix anticoagulant, samples were centrifuged at 4 °C, 3,000 × g for 15 min. Serum supernatants were transferred to endotoxin-free EP tubes and stored at −80 °C until analysis. Assays were performed according to the kit instructions. Serum was analyzed for concentrations of interleukin-1β (IL-1β), interleukin-6 (IL-6), interleukin-10 (IL-10), interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), immunoglobulin A (IgA), immunoglobulin G (IgG), immunoglobulin M (IgM), total superoxide dismutase (T-SOD), malondialdehyde (MDA), glutathione peroxidase (GSH-Px), total antioxidant capacity (T-AOC), and catalase (CAT), and using commercial kits according to the instructions of the manufacturer (Nanjing Jiancheng, China). The ELISA kit was validated with intra-assay CV < 5%, inter-assay CV < 8%, and a standard curve r² of 0.9999. Testing instrument: Multiskan MK3 enzyme label instrument (Thermo Fisher Scientific, China).
Microbiome analysis
-
The processing of sequencing data was conducted as previously described. Sequenced raw reads were deposited in the China National GeneBank DataBase (CNGBdb) under CNP0006729 (Supplementary File 1).
Determination of bacterial growth curve in vitro
-
Lactobacillus strains were obtained from colonic chyme of weaned piglets in this study and the identities of L. amylovorus, L. johnsonii, and L. reuteris and gut microbes (ASV7, ASV8, ASV27, ASV30) in the 16S rRNA gene sequence matched completely. Lactobacillus strains were inoculated on MRS broth (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) at a 1% inoculation rate to then be placed in an incubator at 37 °C. L. amylovorus, L. johnsonii, and L. reuteris were grown in XPC-Ultra contained MRS (0.1% v/v) at the pH of 3.0, 7.5, and 8.4 respectively. OD600 absorbance was measured every 2–3 h. Each experiment was performed in triplicate.
Statistical analysis
-
Data analyses of growth performance, serum indices, and nutrient digestibility were performed using Univariate analysis with SPSS (SPSS 27.0, IBM). Each pen served as an experimental unit and individual pig was considered the experimental unit. Duncan's multiple comparisons analysis of variance (ANOVA) was used for analyzing differences among groups (Supplementary File 1).
-
In Phase I, ADG, ADFI, and F:G were not statistically different among the four dietary treatments (Fig. 1a−c). Across the four dietary treatment groups, no statistically significant variation of ADFI was observed in Phase II or Phase I + II (Fig. 1d, g). Yeast postbiotic improved ADG in the net energy reduced dietary treatments (RED75− vs RED75+) in Phase II and Phase I + II (p = 0.01) (Fig. 1e, h), tended to increase ADG in normal energy dietary treatments (CON vs YC) in Phase II (p = 0.06) (Fig. 1e). Compared with RED75–, F:G of RED75+ was decreased significantly by yeast postbiotic in Phase II (p = 0.04) (Fig. 1f), and tended to be decreased over the entire experiment period (p = 0.08) (Fig. 1i).
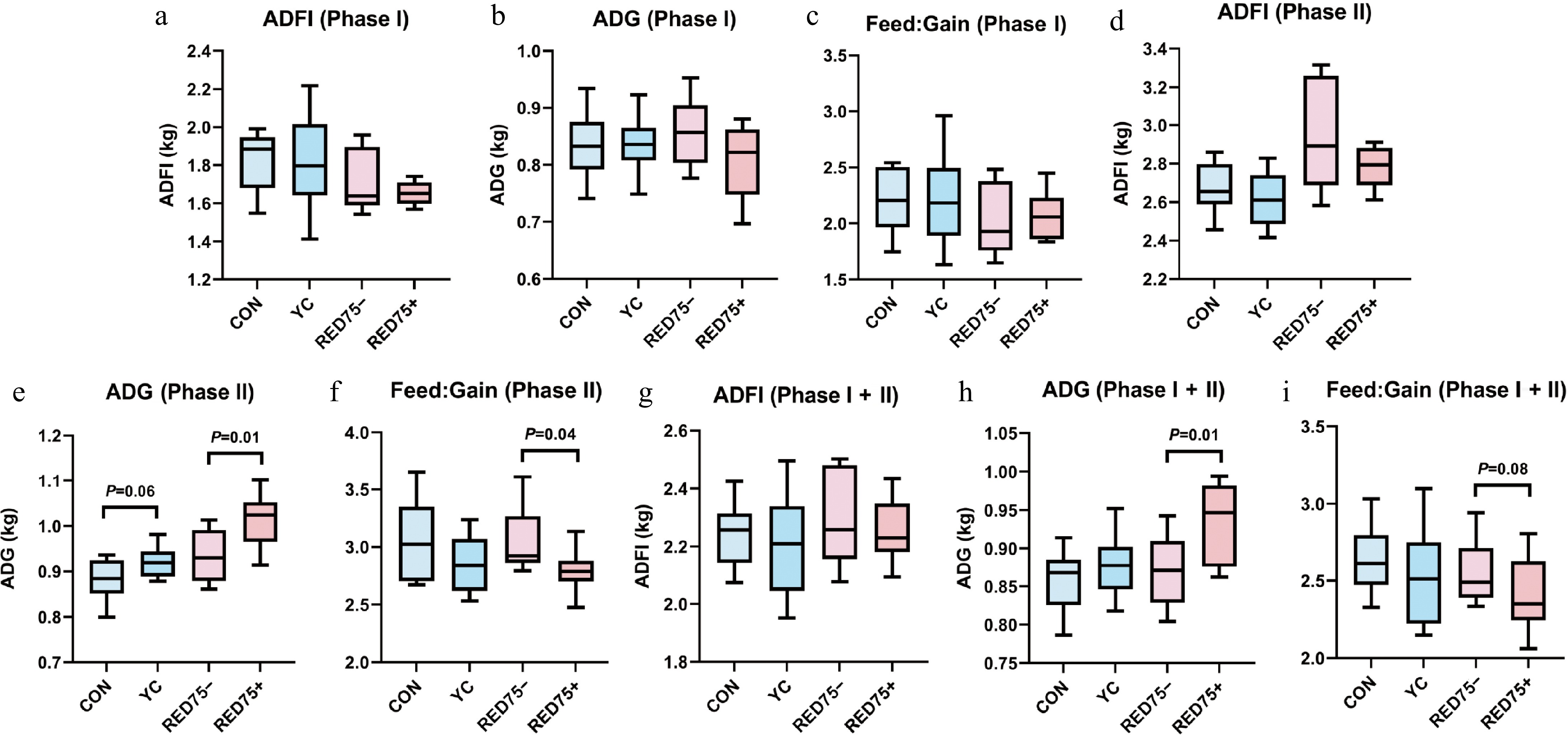
Figure 1.
Effects of supplementation of yeast-derived postbiotic on growth performance of growing pigs. Dietary treatments were: (1) CON: basal diet with normal net energy meet with China National Feeding Standard of Swine (GB/T 39235-2020); (2) YC: CON supplemented with 0.5 g/kg yeast derived postbiotic XPC Ultra diet; (3) RED75–: diet with the net energy reduced by 75 kcal/kg (comparing with CON); (4) RED75+: RED75– supplemented with 0.5 g/kg XPC Ultra diet. Pigs were fed for Phase I (30–50 kg) and Phase II (50–75 kg). ADG, average daily gain; ADFI, average daily feed intake. Bars represented standard error of mean (SEM). Student's t-test was performed when comparing CON and YC, or RED75– and RED75+. The significant level was set as p < 0.05. A trend was set as 0.05 ≤ p < 0.10.
Nutrient digestibility
-
The effects of energy levels and supplementation of yeast-derived postbiotics on nutrient digestibility were presented in Table 2. In Phase I, XPC and energy level variations had no significant effects on nutrient digestibility (p > 0.05). XPC and energy levels exhibited a significant interaction effect on NDF digestibility (p = 0.01). In Phase II, energy levels had no significant effects on nutrient digestibility (p > 0.05). XPC had no significant effects on the digestibility of DM, Ash, and Ca (p > 0.05), but significantly improved the digestibility of CP, NDF, and P, and the GE utilization rate. No significant interaction was observed between XPC and energy levels for nutrient digestibility (p > 0.05). These results indicated that yeast-derived postbiotics had greater improvement in the digestibility of calcium, phosphorus, and gross energy in the low net energy contained diets than the normal energy diets.
Table 2. Effects of yeast-derived postbiotic supplementation and dietary energy concentration on total tract apparent nutrient digestibility in growing pigs (%).
Item CON YC RED75– RED75+ SEM p-value XPC Eng XPC × Eng Indicate number of observations for means Phase I (30–50 kg) Dry matter 87.63 87.61 87.76 87.51 0.06 0.95 0.99 0.95 Crude protein 86.37 88.48 87.95 87.07 0.41 0.75 0.97 0.45 Ash 44.25 45.06 45.11 43.91 0.25 0.84 0.88 0.32 NDF 56.37b 58.25a 57.59ab 56.31b 0.34 0.39 0.31 0.01 Calcium 61.39a 61.55a 60.27a 62.01b 0.36 0.02 0.38 0.44 Phosphorus 57.17 58.09 56.43 58.27 0.41 0.29 0.83 0.72 Gross energy 86.69 87.76 89.08 87.97 0.64 0.99 0.51 0.58 Phase II (50–75 kg) Dry matter 88.10 87.89 87.60 87.70 0.07 0.98 0.86 0.94 Crude protein 83.32a 85.47b 84.65a 86.67b 0.23 < 0.01 0.02 0.91 Ash 40.45 41.27 40.51 41.29 0.21 0.38 0.97 0.98 NDF 53.69a 55.89b 53.01a 55.01b 0.31 < 0.01 0.27 0.76 Calcium 65.07 65.75 64.33 65.34 0.40 0.56 0.69 0.91 Phosphorus 63.11a 64.08a 63.58a 65.45b 0.36 0.01 0.27 0.26 Gross energy 83.25a 84.53ab 82.78a 85.45b 0.61 0.01 0.66 0.18 Experimental diets were a corn-soybean meal-based diet: (1) CON: basal diet with normal net energy meet with China National Feeding Standard of Swine (GB/T 39235-2020); (2) YC: CON supplemented with 0.5 g/kg yeast derived postbiotic XPC Ultra diet; (3) RED75–: diet with the net energy reduced by 75 kcal/kg (comparing with CON); (4) RED75+: RED75– supplemented with 0.5 g/kg XPC Ultra diet. BW: Body weight; NE: Net energy; ME: Metabolizable energy; NDF: Neutral detergent fiber; SID: Standardized ileal digestibility; Eng: energy. Statistical significance was determined using one-way ANOVA among all dietary treatments (n = 16). a, b Values with different superscripts within a row differ (p < 0.05). Serum indices of inflammatory factors, immunoglobulins, and antioxidants
-
For serum indices of inflammatory factors (Table 3), in Phase I, energy level variations had no significant effects on inflammatory cytokine levels (p > 0.05), whereas yeast-derived postbiotics significantly influenced serum inflammatory cytokine levels in growing pigs (p < 0.01). XPC significantly reduced IL-1β, TNF-α, and IFN-γ concentrations at normal energy levels and reduced IFN-γ concentrations at low energy levels (p < 0.01). A significant interaction effect was observed between XPC and energy levels on inflammatory cytokine concentrations. In Phase II, XPC significantly influenced IL-1β and IFN-γ concentrations (p < 0.01, p = 0.01), while energy level variations significantly affected IL-6 and IL-10 concentrations (p = 0.01, p = 0.04). Only a significant interaction effect was observed between XPC and energy levels for the pro-inflammatory factor IL-1β concentrations (p = 0.03).
Table 3. Effects of yeast-derived postbiotic supplementation and dietary energy concentration on serum indices of inflammatory, immunoglobulins, and antioxidant factors in growing pigs.
Item CON YC RED75– RED75+ SEM p-value XPC Eng XPC × Eng Indicate number of observations for each mean Phase I (30–50 kg) IL-1β, pg/mL 29.04a 21.96b 24.84ab 27.02b 0.65 < 0.01 0.25 0.01 IL-6, pg/mL 43.11 52.15 49.23 49.92 2.03 < 0.01 0.12 0.02 IL-10, pg/mL 22.18a 17.09b 18.90ab 20.13ab 0.49 < 0.01 0.81 < 0.01 TNF-α, pg/mL 97.56a 78.28b 93.07a 95.86a 1.92 < 0.01 0.08 0.01 IFN-γ, pg/mL 86.77a 54.14c 65.07c 73.57b 1.94 < 0.01 0.22 < 0.01 IgA, g/L 0.96a 1.06a 1.14b 1.00ab 0.02 0.05 0.28 < 0.01 IgG, g/L 7.05 6.87 6.89 6.78 0.07 0.47 0.37 0.73 IgM, g/L 0.98a 1.08ab 1.15b 1.19b 0.02 0.03 < 0.01 0.15 T-SOD, U/mL 137.32 134.47 138.81 134.13 0.68 < 0.01 < 0.01 < 0.01 MDA, nmol/mL 3.84a 3.68a 3.54a 2.28b 0.12 < 0.01 < 0.01 < 0.01 GSH-Px, U/mL 375.22a 386.69a 395.07a 406.10b 3.86 0.05 0.52 0.68 T-AOC, mmol/L 0.26 0.26 0.25 0.25 0.01 0.94 0.06 0.94 CAT, U/mL 5.78 6.28 5.96 6.64 0.12 < 0.01 0.08 0.54 Phase II (50–75 kg) IL-1β, pg/mL 27.04 28.00 24.09 29.43 0.67 < 0.01 0.26 0.03 IL-6, pg/mL 51.43 52.15 44.08 49.92 1.51 0.13 0.01 0.45 IL-10, pg/mL 18.32 18.15 17.71 16.01 1.27 0.38 0.04 0.84 TNF-α, pg/mL 72.60a 63.99a 71.06b 71.89b 2.33 0.22 0.09 0.07 IFN-γ, pg/mL 60.65 69.64 59.18 71.94 1.83 0.01 0.79 0.25 IgA, g/L 0.96 1.06 1.14 1.02 0.02 0.59 0.01 < 0.01 IgG, g/L 7.07 6.69 6.75 7.05 0.06 0.80 0.90 0.47 IgM, g/L 0.92 1.08 1.24 0.96 0.06 0.05 < 0.01 < 0.01 T-SOD, U/mL 136.34 142.46 143.19 139.71 0.82 0.7 0.56 0.18 MDA, nmol/mL 3.61a 2.99a 3.02a 2.75b 0.11 < 0.01 < 0.01 0.02 GSH-Px, U/mL 392.87 384.26 400.34 400.37 3.53 0.66 0.23 0.66 T-AOC, mmol/L 0.23a 0.24ab 0.28b 0.25ab 0.01 0.12 < 0.01 0.01 CAT, U/mL 5.22 5.80 5.54 6.00 0.14 < 0.01 < 0.01 < 0.01 Experimental diets were a corn-soybean meal-based diet: (1) CON: basal diet with normal net energy meet with China National Feeding Standard of Swine (GB/T 39235-2020); (2) YC: CON supplemented with 0.5 g/kg yeast derived postbiotic XPC Ultra diet; (3) RED75–: diet with the net energy reduced by 75 kcal/kg (comparing with CON); (4) RED75+: RED75– supplemented with 0.5 g/kg XPC Ultra diet. BW: Body weight; NE: Net energy; ME: Metabolizable energy; NDF: Neutral detergent fiber; SID: Standardized ileal digestibility; Eng: energy. Values listed for means and pooled standard error of mean (SEM) of all data are actual data. Statistical significance was determined using one-way ANOVA among all dietary treatments (n = 32).a, b, c Values with different superscripts within a row differ (p < 0.05). For serum indices of immunoglobulins (Table 3), both yeast-derived postbiotic and energy levels influenced the serum levels of the immune factor IgM concentrations. In Phase I, no significant interaction effect was observed (p = 0.15), and low energy levels significantly increased IgM concentrations (p < 0.01). In Phase II, a significant interaction effect was observed between XPC and energy levels (p < 0.01).
For serum indices of antioxidants (Table 3), in Phase I, yeast-derived postbiotic and dietary energy levels significantly influenced the activity of the antioxidant enzyme T-SOD and MDA content, with a significant interaction (p < 0.01). At low energy levels, XPC significantly reduced MDA content and increased GSH-PX activity, enhancing antioxidant capacity. In Phase II, XPC and dietary energy levels significantly influenced MDA content and CAT activity, with significant interactions. XPC significantly reduced MDA content at low energy levels, and alleviated the oxidative stress response.
Microbiota composition
-
Feces collected at the termination points of Phase I and Phase II contained 25,869 (± 3,023) and 27,394 (± 10,825) high-quality sequences respectively. These sequences mapped to 513 (± 84.58) and 549 (± 96.76) unique amplicon sequence variants (ASV) per feces sample of Phase I and Phase II, respectively.
Yeast supplementation reduced α diversity (Chao and Shannon) (CON vs YC or RED75– vs RED75+, p < 0.05) (Fig. 2a). This effect was more significant when yeast was supplemented in reduced energy diets that the alteration occurred in Phase I and Phase II. Considering the similar effect of yeast supplementation in normal energy or reduced energy diets, the following comparison and analysis were conducted using the treatment of RED75– and RED75+. Yeast supplemented diet treated pigs contained the microbial communities that were phylogenetically distinct from those of RED75– treated pigs in Phase I (p < 0.01) and Phase II (p < 0.01; Fig. 2b). A total of 210 and 205 genera were found in RED75– and RED75+ in Phase I, and a total of 235 and 222 genera were found in RED75– and RDE75+ in Phase II (Fig. 2c). The distribution of genera mainly overlapped between RED75– and RDE75+, except that 22 specific genera were unique to RED75– while 17 genera were unique to RED75+ in Phase I, 27 and 14 genera were unique for RED75– and RDE75+, respectively in Phase II (Fig. 2c).
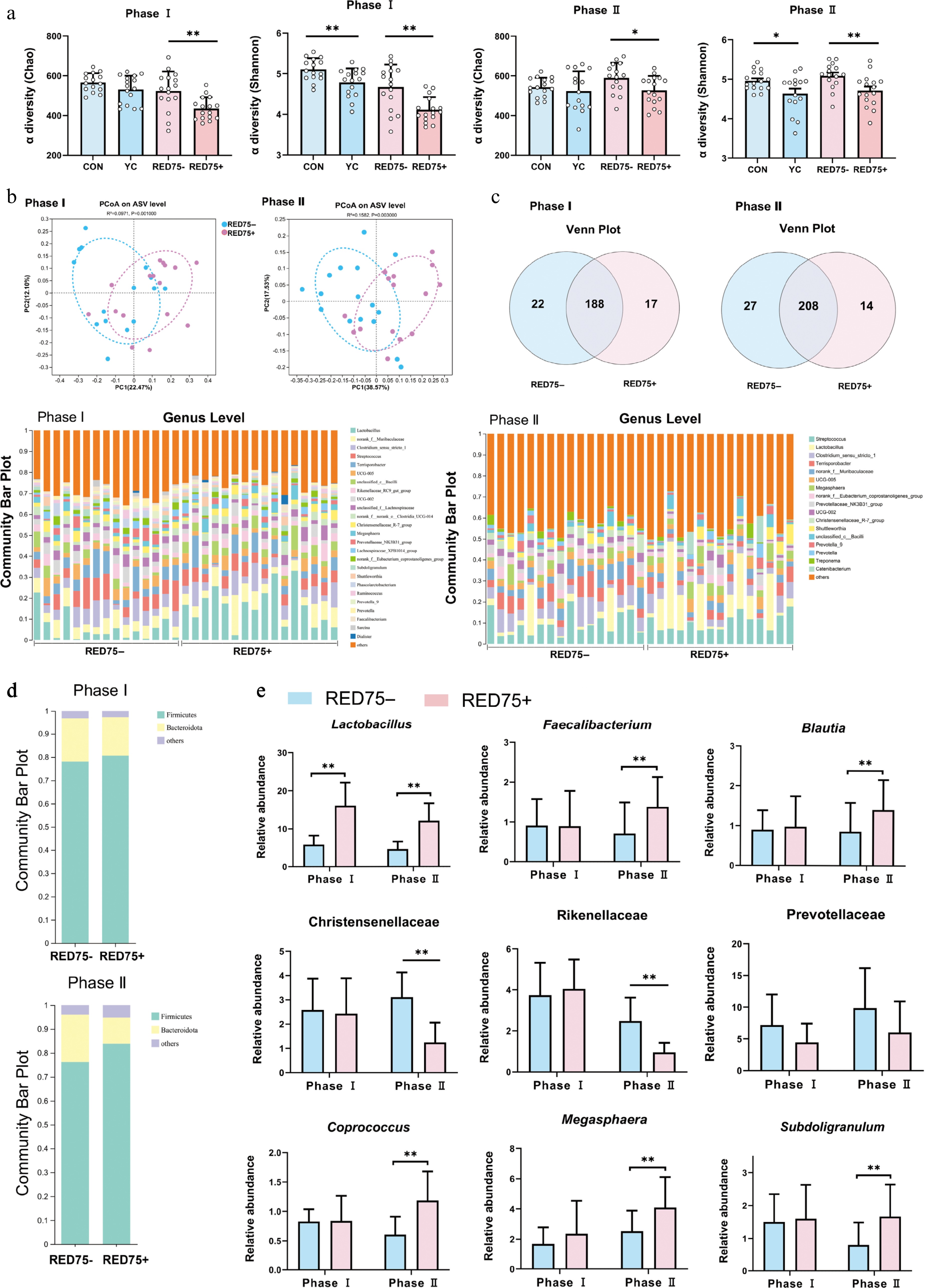
Figure 2.
Effects of supplementation of yeast-derived postbiotic and dietary energy concentration on fecal bacterial composition in Phase I and Phase II. (a) Effect on alpha-diversity represented by Chao and Shannon. (b) Effect on principal coordinates analysis (PCoA). (c) Venn diagram of genera composition of CON and YC. (d) Genus composition of CON and YC represented by the composition of individual fecal sample (n = 16). (e) Abundance of significantly altered genera. Dietary treatments were CON (basal diet); YC (CON with 0.5 g XPC Ultra/kg); RED75– (CON reduced net energy by 75 kcal); RED75+ (CON reduced net energy by 75 kcal with 0.5 g XPC Ultra/kg). Pigs were fed for Phase I (30–50 kg) and Phase II (50–75 kg). p-values were calculated using Adonis test and were adjusted by false discovery rate (FDR) correction according to Benjamini-Hochberg procedure. The significant level was set as FDR < 0.10; * presented FDR < 0.10.
Firmicutes and Bacteroidota were prevailing phyla in both phases and accounted for about 96% of the total phyla composition (Fig. 2e). RED75+ treated pigs had higher Firmicutes and lower Bacteroidota than RED75– in feces (p < 0.05). At the genus level, Lactobacillus was dominant in RED75+ (16% of the total bacteria), comparing to 5% of Lactobacillus in RED75– in Phase I (p < 0.05) (Fig. 2d). In Phase II, beneficial bacteria such as Lactobacillus, Faecalibacterium, and Blautia were more prevalent in RED75+ than RED75– (p < 0.05); presence of harmful bacteria at the family level such as Christensenellaceae, Rikenellaceae, and inflammatory-related Prevotellaceae were lower in RED75+ than RED75– (p < 0.05); the presence of fiber-digesting bacteria such as Megaphaera, Subdoligranulum, and Coprococcus were higher in RED75+ than RED75– (p < 0.05) (Fig. 2f).
Correlation between microbiota and metabolic pathway
-
Correlation analysis was conducted between the screened bacterial genera abundance and metabolic pathways (Fig. 3). By comparing the bacterial compositions of RED75– and RED75+ feces at the genus level, six genera (Lactobacillus, unclassified_c_Bacilli, Rikenellaceae_RC9_gut_group, UCG-002, Prevotellaceae_NK3B31_group, and Ruminococcus) in Phase I, and 12 genera (Clostridium_sensus_stricto_1, Terrisporobacter, Norank_f_Muribaculaceae, Megasphaera, Prevotellaceae_NK3B31_group, UCG 002, Christensenellaceae_R7_group, Rikenellaceae_RCg_qut_group, Lachnospiraceae_XPB1014_group, Subdoligranulum, Blautia, Faecalibacterium) in Phase II, were screened which showed statistically difference (p < 0.05). The abundances of these genera were among the top abundances genera among all analysed microbes. Similarly, there were eight significantly altered metabolic pathways in Phase I, and eight in Phase II when comparing RED75– and RED75+. Correlation analysis revealed that comparing Phase I with Phase II, fewer metabolic pathways were altered. The most abundant Lactobacillus and unclassified_c_Bacilli decreased the paths of biosynthesis of amino acids but increased the paths of energy/glucose metabolism. Oscillospiraceae UCG-002 showed an active role in multiple paths and its alteration trends were negative to those of Lactobacillus. In Phase II, compared to RED75–, more bacteria in RED75+ contributed to altering the pathways of energy/glucose metabolism like ATP-binding cassette transporters (ABC transporters), fructose/mannose/galactose/sucrose metabolism, and carbon fixation. These pathways involved beneficial bacteria such as Lactobacillus, Feacelibacterium, Blautia, and the fiber-digesting Megasphaera, Subdoligranulum, and the harmful Christensenellaceae_R-7_group, Rikenellaceae_RC9_gut_group.
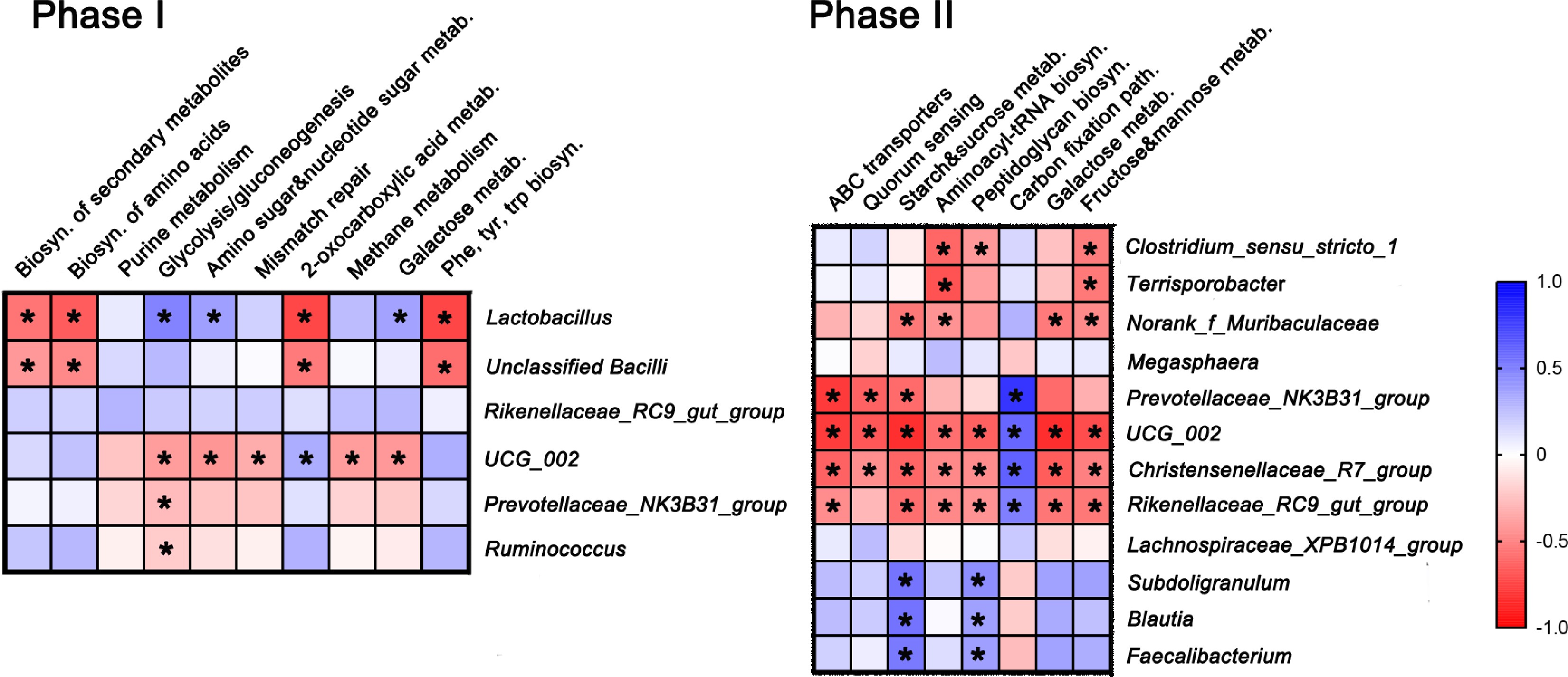
Figure 3.
Correlation analysis between the screened bacterial genera abundance and metabolic pathways. The statistically altered bacterial genera (the abundance was within the top 15 bacteria of the total bacterial composition) and the significant metabolic pathways between CON and YC were screened. Spearman correlation coefficient between bacterial taxa and the numbers of unique sequences mapped to various metabolic pathway were calculated, and p < 0.05 was considered as significant and labeled with *. Dietary treatments were CON (basal diet); YC (CON with 0.5 g XPC Ultra/kg) for growing pigs during Phase I (30–50 kg) and Phase II (50–75 kg).
Effect of growth promotion on Lactobacillus strains in vitro
-
From the above results, yeast-derived postbiotics could enhance the abundance of Lactobacillus strains, L. amylovorus (ASV8), L. johnsonii (ASV7), and L. reuteris (ASV27, ASV30). Therefore an in vitro investigation was carried out to assess the effect of yeast derivatives on the promotion of the growth on these strains (Fig. 4). The intestinal pH values of the stomach, jejunum, and colon were measured in preliminary experiments (Table 4). The results showed that at pH 3.0, mimicking the pH in the stomach, yeast derivatives significantly enhanced the growth of three Lactobacillus strains, especially for L. reuteris the promoted effect occurred instantly. At pH 7.5, mimicking the pH in the jejunum, the promoting effects on three Lactobacillus strains were similar and at pH 8.4 yeast derived postbiotics showed an inhibition effect on the growth of L. amylovorus and L. johnsonii, but no significant difference for L. reuteris. The results indicated that yeast-derived postbiotics helped the growth of Lactobacillus strains in the proximal intestines which is critical for their inhabitation in the gut.
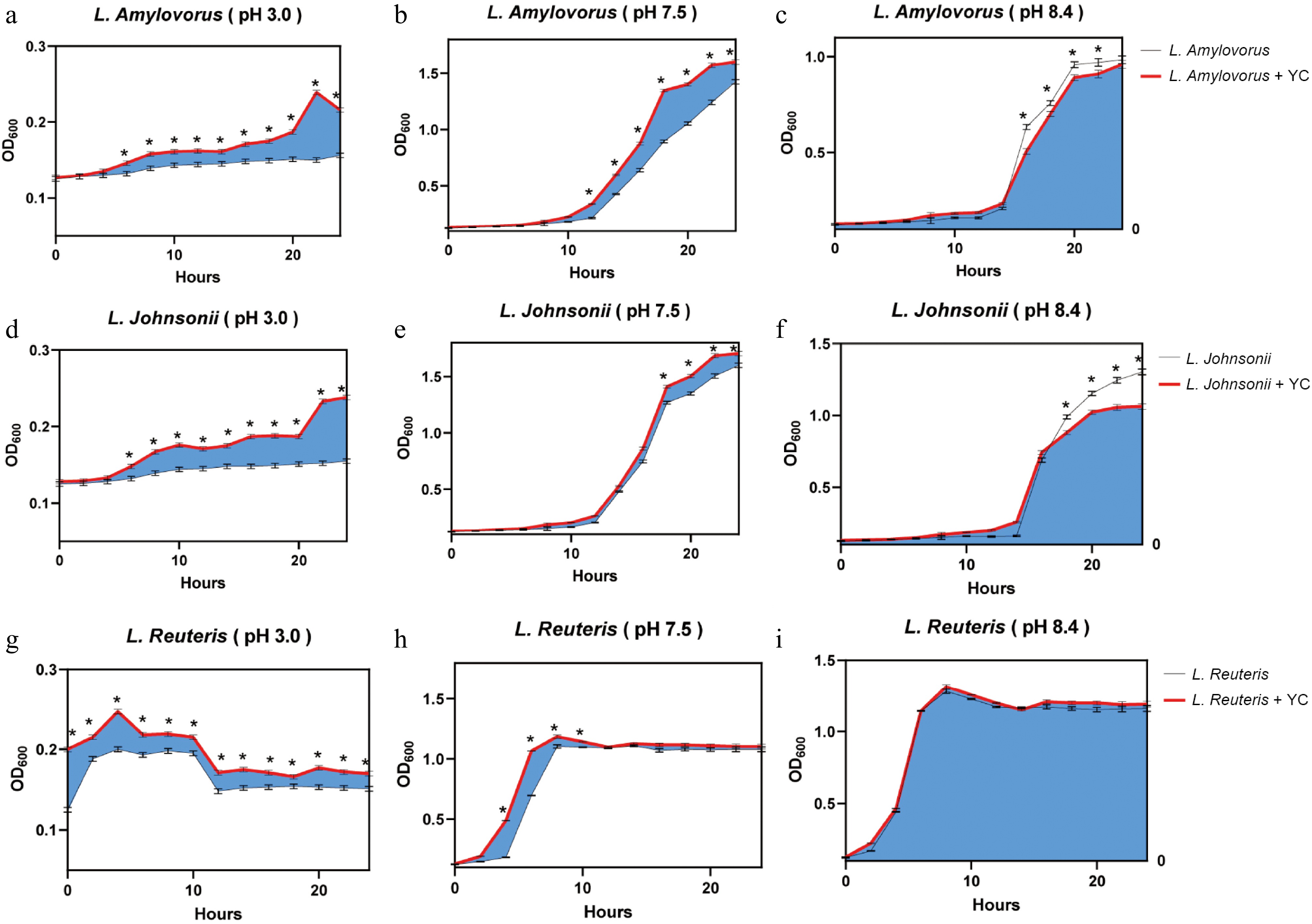
Figure 4.
Growth curves of Lactobacillus strains with yeast-derived postbiotic XPC Ultra at different pH in vitro. Different pH values were mimic the regional pH in stomach, jejunum, and colon. The growth curves (OD absorbance at 600 nm) of L. amylovorus, L. johnsonii, and L. reuteris at pH 3.0, 7.5, and 8.4 (n = 3) were shaded with either blue (MRS broth with XPC Ultra) or white (MRS broth without XPC Ultra). The overlap areas of the growth curves were colored according to the shade color of the growth curve of lower-growing bacteria. Student t-test was performed to statistically differ treatments. The significant level was set as p < 0.05 and labelled as *.
Table 4. The intestinal pH values of stomach, jejunum, and colon in growing pigs.
Item pH SEM Stomach 3.0 0.09 Jejunum 7.5 0.12 Colon 8.4 0.43 Values listed for means and pooled standard error of mean (SEM) of all data are actual data. Statistical significance was determined using one-way ANOVA among all dietary treatments (n = 3). -
Energy in feed constitutes a significant proportion of total feed costs, prompting the implementation of diverse strategies to enhance feed conversion efficiency in the pig industry. One approach is to elevate the utilization of cereal constituents which contain various types of fibers. Against this backdrop, the present study aims to investigate the application of reduced-energy diets in swine nutrition. Living yeast cells or yeast lysate often improved the performance of growing pigs with diets containing high-fiber substances like wheat bran, distillers dried grains with soluble (DDGS), and cassava[11−13]. Yeast-derived postbiotic or yeast lysate improved growth performance via multiple aspects. The in vitro results in this study might explain why yeast-derived postbiotics facilitated the inhabitation of Lactobacillus strains in the gut that yeast derives promoted the growth of Lactobacillus at pH 3.0 and pH 7.5 but not pH 8.4. Early promotion effect on the growth is essentially a benefit for increased abundance of Lactobacillus strains in the intestines.
To enhance the health status and growth performance of finishing pigs, XPC Ultra yeast postbiotics culture was incorporated into the dietary regimen. Postbiotics refer to cell structures and metabolites secreted or released after microbe lysis in complex microbial cultures. XPC Ultra yeast postbiotics are yeast propagated by culture and followed by solid fermentation. It contains yeast cell walls, intracellular metabolites (enzymes and short-chain fatty acids), and secreted metabolites (functional proteins and enzymes)[20]. Compared to probiotics, postbiotics have several advantages such as higher stability, no risk of passing antibiotic-resistant genes, and a wider range of action targets[21]. The cell wall of yeast contains about 75%–85% polysaccharides, which consist of water-soluble mannans, and water-soluble and water-insoluble glucans[22]. In S. cerevisiae, the content of mannans and glucans is nearly equivalent[23]. Mannans can stimulate macrophages in vivo to enhance pathogen clearance by activation of mannosylated compound-recognizing receptors[24] and in vitro to facilitate endosomal degradation of bacteria[25]. The binding of mannans to the macrophage mannose receptor induces the liberation of anti-inflammatory factors IL-1 and TNF for further antigen clearance[26]. During the experimental period, it was observed that dietary supplementation with yeast postbiotics resulted in a reduction in serum inflammatory factor concentrations in finishing pigs. β-glucans can be delivered to small intestines and transmitted to Payer's patches of the gut-associated lymphoid tissue. β-glucans can activate monocytes, macrophages, natural killer cells, and T-helper cells, and stimulate the release of IL-1, IL-2, IFN-γ, and TNF-α[27,28]. Both α-D-mannans and β-D-glucans can inhibit the deleterious effects of mycotoxins by binding to specific receptors or direct absorption of the toxins[29,30].
Second, bioactive components in yeast-derived postbiotic metabolites, including sugars from fermented medium, organic acids, esters, peptides, vitamins, phospholipids, polyphenols, amino acids[31] mannan oligosaccharides, β-glucan[32−34], and nucleotides[35,36], can improve antioxidant status, intestinal morphology, and provide additional elements for body growth. This study demonstrates that yeast extract supplementation significantly elevates serum antioxidant capacity and body growth in finishing pigs. These active components are either substrates for anabolism or active factors for gene expression involved in the body physiological processes. For weanling pigs, particularly when they are under pathogen infection, Saccharomyces cerevisiae fermentation postbiotic enhances the growth performance and intestinal barrier function and lessens the occurrence of diarrhea among piglets[37]. During late gestation and lactation stages, yeast-derived postbiotic supplementation decreases inflammatory response in sows and increases weaning weight and immune function in piglets[38]. In fattening calves whose food is relatively rich in concentrate, yeast-derived postbiotics could notably increase ruminal pH, promote microbial colonization and progression, and improve production performance[39]. In aged laying hens, yeast-derived postbiotic promotes the egg production by improving the intestinal health status[40].
Third, yeast-derived postbiotics play a pivotal role in the modulation of gut health by altering gut microbes. This aspect of function may largely contribute to the benefits of yeast-derived postbiotics on improved feed conversion rate and fiber degradation. Gut microbes can produce numerous enzymes to aid animal digestion, especially in the digestion of cereal cell walls that contain xylan, fructan, and lignin because animals can not directly degrade these compounds due to a lack of endogenous enzymes. Yeast S. cerevisiae can enhance the growth of relevant bacteria that degrade fiber. In the current study, compared to un-supplemented control, yeast-derived postbiotics numerically improved the apparent digestibility of NDF in growing pigs and increased the abundance of fiber-degrading bacteria Megasphaera[41,42], Subdoligranulum[43,44], and Coprococcus[45]. These results suggest that yeast-derived postbiotics promote the colonization of fiber-degrading bacteria and improve the ability to degrade fiber in growing pigs. Yeast-derived postbiotics contain a large proportion of β-glucan, mannan, and mannan oligosaccharides which can be broken down by fiber-degrading bacteria. The small molecules of oligosaccharides, disaccharides (such as sucrose, fructose, lactose, maltose, rhamnose, and raffinose) from gut fermentation are substrates for the growth of Lactobacillus[46−52], Faecalibacterium[53,54] and Blautia[55]. In this study, the abundance of Lactobacillus was significantly enriched after feeding yeast-derived postbiotics. Cross-feeding interactions occur among the bacteria. For example, Lactobacillus produce acetate and lactic acid[49] which are substrates for butyrate producing Faecalibrium, Blautia, and Coprococcus. Faecalibacterium receive beneficial feeding from Lactobacillus by upregulating its genes for cell wall formation and inhibition of cell wall degradation[56]. Acetate producing Blautia enhanced the growth of butyrate producing Faecalibacterium to provide energy for colonocytes[57].
In this study, Lactobacillus was profoundly increased by yeast-derived postbiotic supplemented in RED75+ as the abundance of Lactobacillus represented 16% of the total bacterial abundance compared to un-supplemented RED75– (in Phase I). This indicated yeast-derived postbiotic were favorable for growth of Lactobacillus. Previous researchers reported some evidence of yeast S. cerevisiae benefiting growth of Lactobacillus. For example, surface mannans of Saccharomyces cerevisiae interacted with surface proteins of Lactobacillus forming a mixed-species biofilm[58] that protected Lactobacillus from pathogenic bacteria[59,60] and antibiotics[61]. β-glucan could increase the adhesion ability of Lactobacillus to intestinal epithelia[48] by inducing synthesis of exopolysaccharides, surface proteins, and transporters[62]. Arabinoxylan (AX) is the predominant fibrous polymer in wheat bran and rice bran[63]. Arabinoxylan fractions from wheat stimulated Lactobacillus growth[46] with shorter AX being more effective[64]. Therefore, both yeast-derived postbiotic and AX from wheat bran and rice bran benefit the growth of Lactobacillus.
Lactobacillus strains can produce a series of enzymes including protease, peptidase, ureases, amylases, and esterases that aid nutrient digestion[65]. These secreted enzymes may explain the increased digestibility of protein associated with an increased abundance of Lactobacillus in this study. In addition, some strains of Lactobacillus can produce antimicrobial peptides[66−68]. These antimicrobial substances could contribute to a lower abundance of pathogenic biomarker bacteria such as Eubacterium-scoprostanoligenes-group[69], infectious Prevotella[70,71], and Terrisporobacter[72,73]. Terrisporobacter induces gut oxidative stress and inflammation in pigs after weaning[74]. Like other probiotics, Lactobacillus itself is an immune modulator. Lactobacillus can induce recognition of Toll-like receptors and interact with host intestinal epithelial cells to recruit immune cells to infection sites by activating mucosa-associated lymphoid tissue which recruits intestinal immune cells[75]. Lactobacillus can induce specific immune markers by producing pro- and anti-inflammatory cytokines[76,77]. Lactobacillus can enhance the production of IgA that does not directly participate in inflammatory responses but prevents antigen binding to or penetration of epithelial cells[78,79]. Overall, immune modulation by Lactobacillus is systemic and involves physical (mucus), molecular (antimicrobial proteins), and cellular components to synergistically protect the body from pathogenic microbial invasions.
High-fiber ingredients are extensively used in developing countries. Great efforts are being made to save protein and energy from animal feed for direct human consumption. A huge number of alternative feedstuffs are considered agricultural waste by-products such as wheat bran, and rice bran. DDGS are used in feed production. However, these by-products are characterized as high-fiber ingredients. The presence of non-digestible polysaccharides such as β-glucan, xylan, mannan, and fiber-bound protein limits the digestibility and utilization of dietary nutrients. Supplementation of yeast-derived postbiotics, which subsequently alters the gut microbial composition, provides exogenous enzymes to digest these polysaccharides and produce energy substrates for body metabolism. Acetate, propionate, and butyrate are the predominant microbiota-derived short-chain fatty acids (SCFA). Butyrate is the key energy source for colonocytes[57]. Butyrate directly activates intestinal gluconeogenesis via a cAMP-dependent pathway[80]. Propionate is transferred to the liver to participate gluconeogenesis[80,81]. SCFA can interact with gut G protein-coupled receptors (GPR), GPR 41 and GPR43, to activate intestinal gluconeogenesis[82,83]. Synthesized glucose through gluconeogenesis using SCFA as substrates provides extra energy for metabolism.
In the current study, we designed a dietary treatment with reduced net energy compared to the required net energy for Chinese pigs[15]. The treatment with reduced energy was supplemented with yeast-derived postbiotics, un-supplemented treatment as a counterpart. Diets were formulated by reducing proportions of corn and soybean meal with replacement by wheat bran and rice bran meal. Body weight results revealed a compensatory effect of yeast-derived postbiotics in reduced energy diets by improving the digestibility of NDF and crude protein. In high-fiber diets, digestibility of NDF often increased as NDF content increased as long as the fermentation capacity of the hindgut microbe increased proportionately. But if NDF content exceeded the fermentation capacity of gut microbes, the digestibility of NDF decreased[84]. Increased NDF in diets decreased ADG, and digestibility of CP and gross energy in growing pigs[85,86]. In the current study, supplemented yeast-derived postbiotics maintained similar digestibility of NDF in the diets with normal energy (YC) and reduced energy (RED75+), which indicated that yeast-derived postbiotic supplementation extracted energy from NDF to maintain a similar growth of pigs as pigs fed a normal energy diet.
Improved antioxidant status of pigs fed yeast-derived postbiotics might be attributed to MDA, GSH-Px, T-AOC, and CAT, were contributed by both yeast-derived postbiotics and wheat/rice bran. Wheat bran and rice bran are antioxidants due to their content of phenolic acid, flavonoids, tocophenols, tocotrienols, and c-oryzanol[87−89]. Even though wheat and rice bran likely inferred some antioxidant capacity, we cannot rule out the direct antioxidant effects of yeast-derived postbiotics.
-
Low-energy diets may reduce feed costs and alleviate inflammatory responses. However, supplementing yeast-derived postbiotics is necessary to synergistically alleviate oxidative stress under low-energy conditions. Supplementation of yeast-derived postbiotics to the diets of growing pigs was beneficial to the pigs' growth performance and feed conversion rate. Yeast-derived postbiotics improved digestibility of NDF, protein, calcium, and phosphorus, decreased serum inflammatory factors, and improved anti-oxidant status. Yeast-derived postbiotic improved gut microbial composition by increasing the abundance of Lactobacillus, Faecalibacterium, and Blautia, increasing fiber-digestible bacteria, Megaphaera, Subdoligranulum, and Coprococcus, and decreasing the Christensenellaceae, Rikenellaceae. Yeast-derived postbiotics promoted the growth of Lactobacillus strains in vitro at pH 3.0 and pH 7.5. These results demonstrated that yeast-derived postbiotic supplementation was beneficial to growing pigs by promoting the growth of Lactobacillus.
-
All procedures were reviewed and preapproved by the Ethics Management Committee of China Agricultural University, identification number: 202205410479536, approval date: 2022.05.11. The research followed the 'Replacement, Reduction, and Refinement' principles to minimize harm to animals. This article provides details on the housing conditions, care, and pain management for the animals, ensuring that the impact on the animals was minimized during the experiment.
-
The authors confirm their contributions to the paper as follows: study conception and design: Li Q, Zhang Y, Zhang S, Lu P, Shi Q; data collection: Li Q, Zhang Y, Guo X, Guo S, Ma C; analysis and interpretation of results: Li Q, Zhang Y; draft manuscript preparation: Li Q; language edit: Johnston LJ; Dong B: study design, financial support, and manuscript revision. All authors reviewed the results and approved the final version of the manuscript.
-
16S rRNA sequencing data of gut microbiota that support the findings of this study are available in the the China National Genebank Database repository (CNGBdb CNP0006729).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Qianxi Li, Ying Zhang
- Supplementary File 1 Supplementary materials and methods to this study.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Q, Zhang Y, Guo X, Guo S, Ma C, et al. 2025. Effects of yeast-derived postbiotic in different energy diets on growth performance, immunity, antioxidant status, and intestinal microbiota in growing pigs. Animal Advances 2: e028 doi: 10.48130/animadv-0025-0024 

Effects of yeast-derived postbiotic in different energy diets on growth performance, immunity, antioxidant status, and intestinal microbiota in growing pigs
- Received: 09 March 2025
- Revised: 09 June 2025
- Accepted: 11 June 2025
- Published online: 11 October 2025
Abstract: To explore the impacts of yeast-derived postbiotic on growth performance and intestinal microbiota of growing pigs, 192 healthy Duroc × Landrace × Yorkshire (initial weights 30 ± 0.6 kg) were subjected to four dietary treatments, with eight replicates per treatment and six pigs per replicate. The diets were: CON: basal diets; YC: CON with 0.5 g/kg yeast-derived postbiotic; RED75–: CON with net energy reduced by 75 kcal/kg; RED75+: RED75– with 0.5 g/kg yeast-derived postbiotic. The results showed that yeast supplementation improved average daily gain in the low energy dietary treatments, tended to improve nutrient digestibility and feed conversion rate. Yeast supplementation in normal energy diets decreased the levels of serum inflammatory factors including IL-1β, IL-10, TNF-α, IFN-γ, and IgA. Yeast in low-energy diets improved antioxidant status by decreasing serum MDA and increasing GSH-Px. Yeast significantly reduced α diversity of the pig feces. Compared to RED75–, Lactobacillus, Faecalibacterium, and Blautia were more prevalent in RED75+, at the family level Christensenellaceae, Rikenellaceae, and inflammatory-related Prevotellaceae were lower in RED75+, the abundance of fiber-digesting bacteria Megaphaera, Subdoligranulum, and Coprococcus were higher in RED75+. Correlation analysis revealed that altered bacterial composition involved glucose/mannose/galactose/fructose metabolism. In vitro, yeast-derived postbiotics promoted the growth of L. amylovorus, L. johnsonii, and L. reuteris at different pH. Our findings provide evidence that supplementation of yeast-derived postbiotics is beneficial to growth performance, anti-inflammatory, antioxidant status, and intestinal microbiota composition of growing pigs by promoting the growth of Lactobacillus.
-
Key words:
- Antioxidant /
- Digestibility /
- Inflammation /
- Nutrient digestibility /
- Fiber