








-
Despite the continuous development of synthetic drugs, plants remain among the most important sources of drugs. Even today, more than half of the world's population still relies on medicinal plants to treat different human diseases[1,2]. However, in the past 100 years, global climate change has posed an extreme and irreversible threat to the cultivation, growth, and harvest of medicinal plants, with a decreasing number of medicinal plant species and active ingredients in traditional herbal medicine[3−5]. These adverse effects seriously constrain the in-depth development and utilization of traditional herbal medicines. Numerous investigations have focused on the effects of abiotic stresses such as drought, extreme precipitation, warming, UV radiation, salt stress, or atmospheric nitrogen deposition on the secondary metabolites of medicinal plants[6,7], but studies on the effects of serious ground-level ozone stress on medicinal plants are insufficient.
Ozone (O3) is a highly reactive colorless gas in the atmosphere that is composed of three oxygen atoms. The production of ozone involves the splitting of molecular oxygen, O2, into two atoms by strong UV radiation for photodissociation and then combining with another atom. An oxygen molecule recombines with a pungent odor and is mostly concentrated in the stratosphere[8,9]. Through absorbing UV radiation, it regulates the temperature of the stratosphere and has a protective effect on the Earth[10]. In the troposphere 15 km from sea level, O3 accounts for only 10% of the total volume of ozone, which is mainly secondary pollutants generated by photochemical reactions such as atmospheric nitrogen oxides (NOx), volatile organic compounds (VOCs), and carbon monoxide (CO). A highly toxic global air pollutant that has a major impact on the survival of humans, animals, plants, and microorganisms. The annual average concentration of ground-level ozone has increased from 10–15 ppb at the end of the 19th century to 40–50 ppb at present. Several regions are generally facing the threat of ozone, including Europe, North America, the Middle East, China, and India, where it is projected to increase by 60% from current levels by 2100[11−13]. Currently, hundreds of terrestrial plant species are threatened by elevated ozone concentrations[14−16]. Ozone stress has caused yield decreases of 5%–48% in several major crops in East Asia, and economic losses will reach USD$2.21–29.1 billion[17−19]. Generally, the ozone level peaks in summer, which overlaps with the growing season of most plants. For example, Ziziphi Spinnsae Semen (ZSS) is a traditional Chinese herbal medicine widely cultivated in the Taihang Mountains. It has the effects of nourishing the liver, calming the mind, soothing the spirit, and reducing sweating. However, the daily maximum 8 h ozone mean concentration (MDA8) exceeds 90 ppb in some ZSS production areas, such as Shijiazhuang and Xingtai, Hebei, China[20]. When the hourly ozone concentration is greater than 40 ppb, plants experience oxidative stress[21].
The effects of ozone on plants have been reviewed by scholars at different scales and perspectives. Feng et al. reviewed the macroscale effects of terrestrial ecosystems on ozone stress[22]. Zhang et al. focused on the impacts of elevated atmospheric ozone on plant-related carbon cycle processes, including carbon dioxide (CO2) assimilation, soil carbon sequestration, and litter decomposition[23]. Singh et al. conducted a detailed summary of plant secondary metabolites under ozone exposure[24]. Nowroz et al. reviewed the potential negative effects of increased tropospheric ozone on crops, the antioxidant defense mechanisms of plants, and the measures taken to mitigate ozone stress[25]. Nevertheless, the above studies focused primarily on the impact of ozone stress on crops, horticultural species, and a limited number of fruit trees and vegetables. The impact on medicinal plants has been explored in only a small number of studies. Notably, research on medicinal active ingredients remains limited.
In comparison to other abiotic stresses, the effects of ozone stress on plants have been the subject of relatively few studies, and therefore, the existing literature was relatively easy to collect and organise. However, it must be acknowledged that there are instances of ambiguity in the delineation of medicinal plants, particularly in the context of medicine-food homology and ornamental plants. Consequently, the following principles were followed in the literature screening: (1) the selected papers were obtained from the Web of Science Core Collection, Scopus Preview, and the China Knowledge Network; (2) priority should be given to the retention of species that have explicit medicinal value, as stated in the paper; (3) studies on conventional crops, vegetables and fruits were excluded; (4) the indicators determined contain plants with recognized medicinal active ingredients. Here, this paper elaborates on the effects of ozone stress on phenotypes, antioxidant defenses, secondary metabolites, gene regulation, and symbiotic microorganisms and proposes the understanding of future research directions. This review is important for understanding the effects of ground-level ozone stress on medicinal plants, guiding the production of medicinal plants, and protecting human health.
-
Ozone stress affects the photosynthesis, respiration, and other physiological and biochemical processes of medicinal plants, which eventually leads to alterations in the phenotypic characteristics of different parts used in medicine, which often manifests as slow plant growth, weak seedlings, and reduced leaf color and fruit setting rates. Ozone first enters plants through their stomata and bears the brunt of its effects on leaves, especially whole-herbal medicinal plants. Compared with herbaceous plants, trees and shrubs grow slowly, have a long life cycle, thicken the cuticle after leaf maturation, and have a strong ability to regulate stomatal opening. Therefore, ozone stress often causes delayed damage to trees and shrubs, which requires continuous observation for a longer time. Studies have shown that high concentrations of ozone can reduce the photosynthetic rate and stomatal conductance, reduce chlorophyll and carotenoid contents, destroy leaf cell walls and cell membranes, accelerate leaf senescence, and eventually lead to decreased plant height and carotenoids[26]. Table 1 shows the phenotypic changes in medicinal plant leaves after ozone stress, and slight differences were detected depending on the glandular trichomes, thickness of the mesophyll, and degree of waxing. However, unlike those of diseases, the effects of ozone stress on plant leaves are often uniform rather than localized. According to the criteria of the 'Forest Health Expert Advisory System', the characteristics of ozone damage to plant leaves are first manifested as the appearance of uniform fine spots, most of which are yellow to brown[27]. In particular, damage occurs between the leaf veins. The symptoms of injury on old leaves are more severe than those on young leaves without obvious insects or mildew spots[28]. This may lead farmers to inappropriately consider other diseases as the causative agent[29]. For the cultivation of medicinal plants, some farmers often mistake the damage induced by ozone stress on leaves for disease or insect damage and thus misuse pesticides, causing environmental stress and irreparable economic losses.
Table 1. Phenotypic changes in medicinal plants under elevated levels of ozone.
Country Plants Medicinal parts Fumigation method Ozone treatment Leaves symptom Ref. Brazil Tibouchina pulchra (Cham.) Cogn. Leaves OTCs Ambient, no-filtered air + 40 ppb for 8 h, 25 d Interveinal red stippling da Rosa Santos et al.[95] China Ginkgo biloba L. Leaves OTCs 80 ppb for 9 h, 110 d Leaf blades turning yellow with yellow spots, leaf margins curling He et al.[58] China Monarda didyma L. Leaves OTCs 160 ppb for 10 h, 14 d Localized yellow‒brown spots Wang et al.[26] China Ziziphus jujuba Mill. Var. spinosa (Bunge) Hu ex H. F. Chou Leaves OTCs 95 ppb for 8 h, 60 d Yellow spots Fu et al.[46] Egypt Mesembryanthemum crystallinum L. Leaves, root OTCs Ambient + 20 ppb for 8 h, 120 d Red pigmented leaves appeared; nonsignificant reductions Mohamed et al.[33] Germany Melissa officinalis L. Leaves Growth chambers 80 ppb for 5 h Chlorotic spots Döring et al.[96] Germany Melissa officinalis L. Leaves OTCs 80 ppb for 5 h Small chlorotic spots on the adaxial leaf surfaces Döring et al.[97] Italy Melissa officinalis L. Leaves Growth chambers 200 ppb for 5 h Roundish and dark-blackish necrosis in the interveinal adaxial areas Pellegrini et al.[43] Italy Hypericum perforatum D. Don Apical shoots OTCs 110 ppb for 5 h No visible symptoms Pellegrini et al.[39] Italy Salvia officinalis L. Leaves OTCs 200 ppb for 5 h No visible symptoms Marchica et al.[36] Italy Salvia officinalis L. Leaves OTCs 200 ppb for 5 h, 36 d Interveinal stippling and chlorotic spots Marchica et al.[38] India Sida cordifolia L. Leaves, floral OTCs Ambient + 20 ppb, 90 d Chlorosis and necrotic spots; the lengths of calyx, corolla, and stamen decreased Ansari et al.[30] India Costus pictus D.Don Leaves OTCs 57 ppb for 5 h, 120 d Interveinal stippling and chlorotic spots Ansari et al.[54] India Sida cordifolia L. Leaves OTCs 63 ppb for 5 h, 90 d Trichome density increased Ansari et al.[4] India Cymbopogon flexuosus (Steud.) (Wats.) Leaves OTCs 76.3 ppb for 5 h, 90 d Shoot and root length, leaf area decreased Madheshiya et al.[37] India Andrographis paniculata (Burm. f.) Nees Leaves OTCs Ambient + 20 ppb for
8 h, 90 dChlorosis and necrotic spots Ansari et al.[98] Ozone stress further affects the flowering, pollination, and fruiting of medicinal plants. For instance, when Sida cordifolia L. (Mallow) was fumigated at an ozone concentration of 70 ppb for 90 d, the start of flowering was 3 d earlier; the lengths of the calyx, corolla, and stamens were reduced by approximately 10%, and pollen viability was significantly decreased by 41.6%[30]. The number of viable pollens plays a very important role in the success of sexual reproduction and may affect the fertilization process. Ozone stress can also cause changes in the color of petals and the scent of flowers, thus affecting the interaction between plants and pollinating insects and even producing toxicity for some herbivorous insects, reducing the potential foragers for bees, butterflies, flies and moths[31], all of which may lead to a decrease in the reproductive ability of medicinal plants and, further, a low seed setting rate, poor growth or even death of fruit and seed plants, thus affecting the phenotypes of flower and fruit seed plant characteristics and active pharmaceutical ingredients[32].
Herbal medicines derived from roots and tubers are significantly more prevalent than those sourced from whole plants, flowers, seeds, fruits, or other plant parts. Ozone has the potential to modify the underground biomass accumulation and rhizosphere microenvironment of medicinal plants by impacting the aboveground parts of these plants (Fig. 1). The adverse effects of roots and rhizomes, when utilized as medicinal components, are variable under ozone stress, yet these impacts have been largely neglected. As ozone exposure increases, carbon fixation by plants decreases. To maintain their survival under ozone stress, plants attempt to 'trade off' resources among different organs. This strategy reduces carbon allocation to the root system, resulting in a decrease in the root-to-shoot ratio and the biomass of fine roots in Mesembryanthemum crystallinum L. (ice plant)[33]. Similarly, a meta-analysis revealed that ozone stress exacerbated the loss of plant root biomass[34]. When roots are under the influence of ozone, their ability to absorb water and nutrients decreases, which results in the feedback of shoot growth and photosynthesis. Optimal shape and high quality are important manifestations of genuine medicinal materials[35]. Phenotypic alterations in medicinal plants due to ozone stress may significantly impact the collection, processing, and clinical application of these materials.
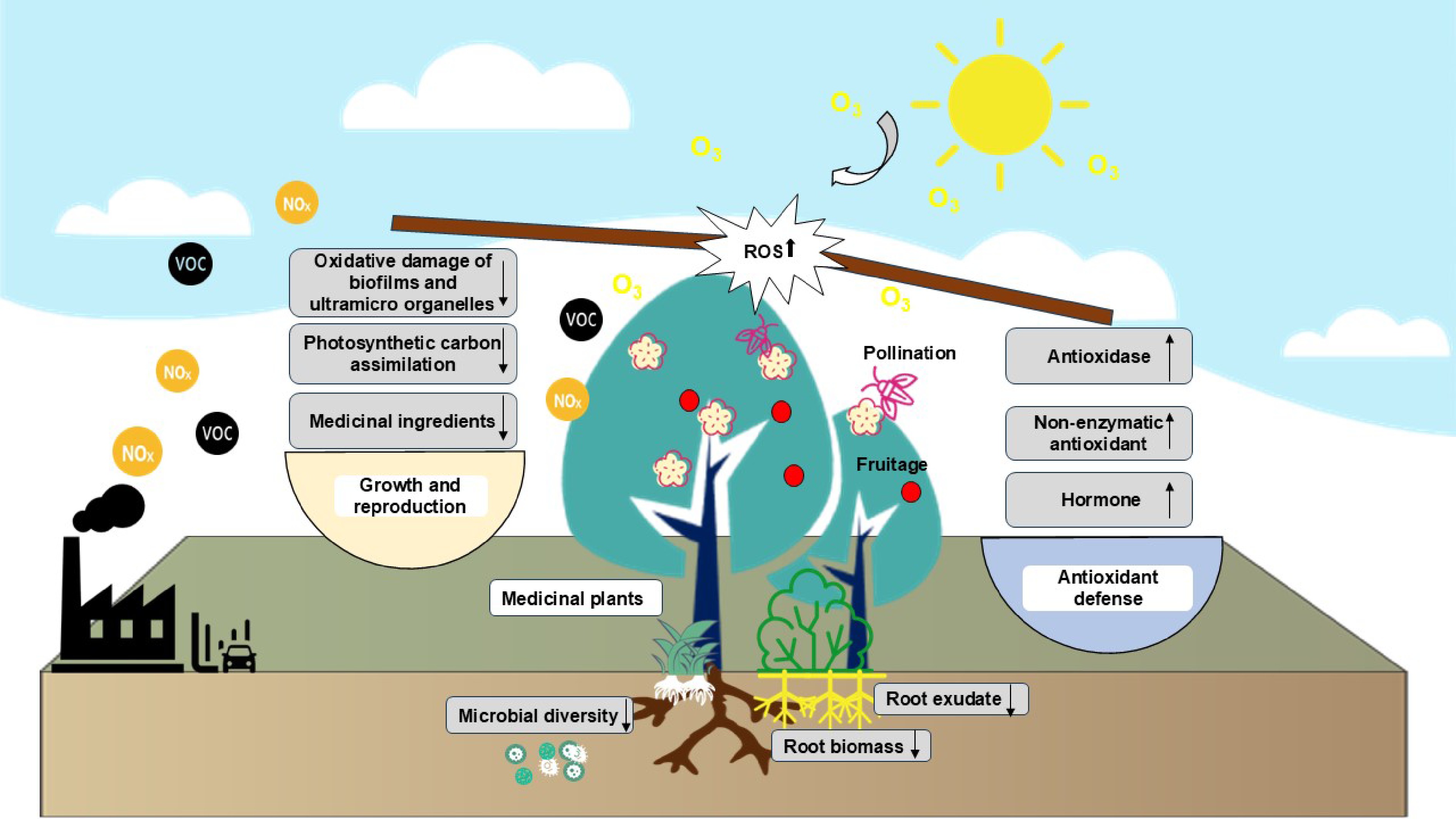
Figure 1.
Effects of ozone stress on the aboveground-underground ecosystem of medicinal plants.
-
When medicinal plants are under environmental stress, the signaling, production, and removal of reactive oxygen species (ROS) are regulated inside plants to control their toxic effects[36]. As the impact of ozone stress on medicinal plants is primarily initiated at leaves, this study employed a leaf-based model to delineate the antioxidant defence mechanisms (Fig. 2). O3 stress induces the antioxidant system in medicinal plants to initiate the defense mechanism. Antioxidant enzymes such as peroxidase (POD), catalase (CAT), superoxide dismutase (SOD), and glutathione reducing enzyme (GR) and nonenzyme substances such as ascorbic acid (AsA) and glutathione (GSH) are the most important antioxidant substances in plants and play important roles in preventing cell damage caused by biological free radicals and maintaining cell redox homeostasis[37−39]. The ASC‒GSH (ascorbate‒glutathione) cycle has received much attention because it can directly remove H2O2 produced in plants; additionally, the antioxidants produced in this cycle can also remove other types of H2O2 through other pathways[39]. When exposed to high concentrations of ozone, ROS are formed in plant cells, alter gene expression, and ultimately cause cell death, which also plays dual roles, acting as signaling molecules to regulate different physiological processes and responses to stress and inducing oxidative damage[40]. However, plants possess a finite capacity to withstand ozone stress. When the intracellular accumulation of ROS surpasses the threshold of cellular antioxidant defenses, it can impair the functionality of the cellular protective mechanisms. This leads to an exacerbation of membrane lipid peroxidation, a decrease in enzyme activity, and ultimately, damage to the physiological integrity of the cell[41].
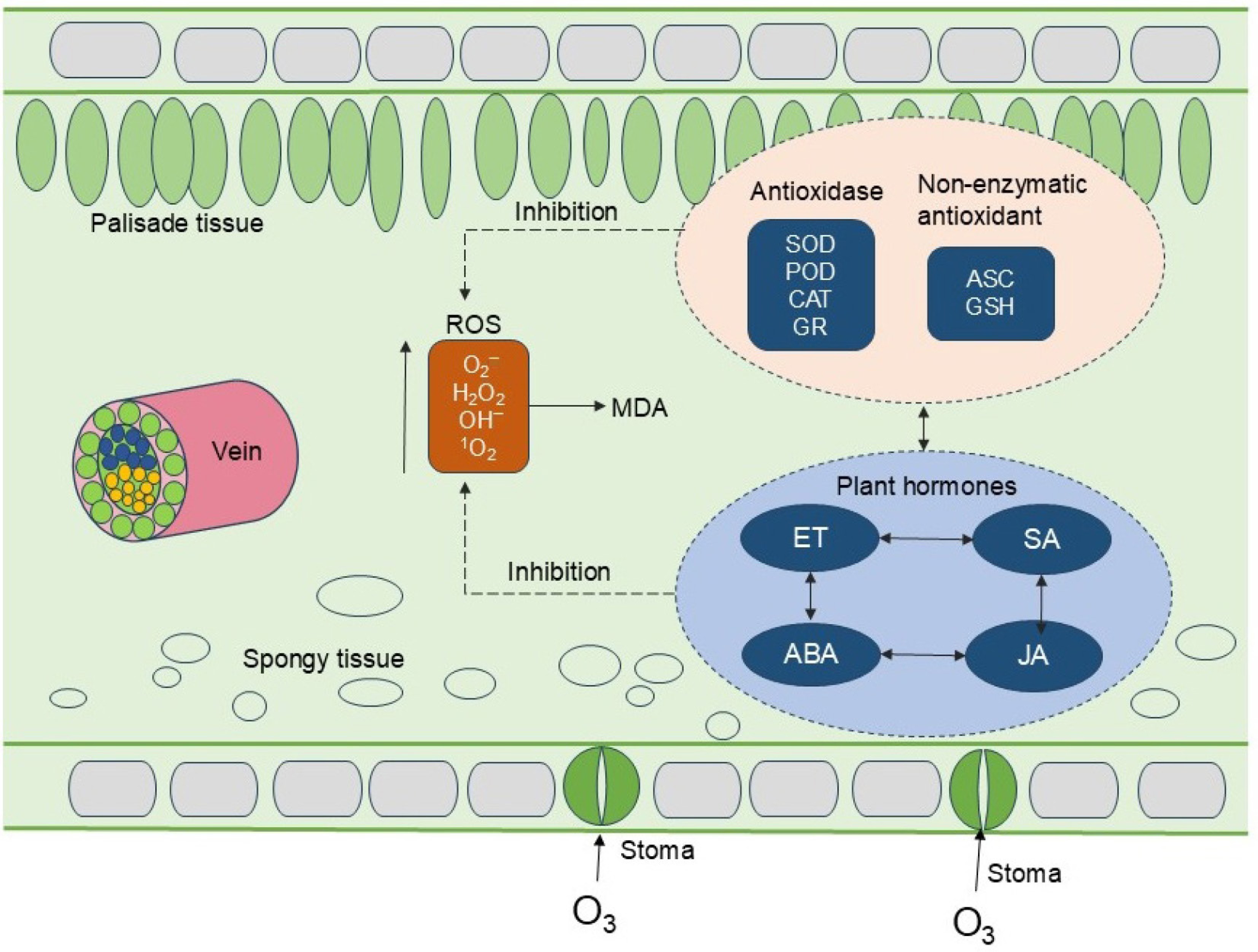
Figure 2.
Antioxidant defense mechanism of medicinal plant leaves under ozone stress. ROS, reactive oxygen species; O2−, superoxide anion; OH−, hydroxide ion; 1O2, singlet oxygen; H2O2, hydrogen peroxide content; MDA, malondialdehyde content; SOD, superoxide dismutase activity; POD, peroxidase activity; CAT, catalase activity; GR, glutathione reductase activity; ASC, apoptosis-associated speck-like protein containing a CARD; GSH, glutathione reduced; ET, ethylene; SA, salicylic acid; ABA, abscisic acid; JA, jasmonic acid.
ROS signaling and phytohormone defense responses are interrelated[42]. As key signaling molecules involved in the regulation of plant growth and development, plant hormones play important roles in the plant response to ozone stress[43]. As stress signals, ROS also promote the defense responses of hormones such as abscisic acid (ABA), salicylic acid (SA), and jasmonic acid (JA) to induce stomatal closure and reduce ozone uptake and water evaporation, thereby helping plants cope with the oxidative stress caused by ozone[25,44]. After exposure to an ozone concentration of 200 ppb for 5 h, the ABA content in Salvia officinalis L. (Common sage) increased by 25%, and ethylene release was twice that of the control[38]. ABA can promote H2O2 production through NADPH oxidase[44]. Ozone exposure may affect JA signaling, an important hormone involved in plant defense responses. Under 200 ppb ozone stress, the JA content in the leaves of Melissa officinalis L. (Lemon balm) was much greater than that under the nonfumigation treatment and remained high for a long period[45]. However, the interactions and synergistic effects of different plant hormones under ozone stress have not been clearly revealed and should be further investigated.
-
The allocation of resources by medicinal plants often involves a trade-off between growth and defence in response to ozone stress. This enables the medicinal plants to cope with the stress and attain homeostasis. The limitation of ozone on photosynthesis affects the accumulation of primary metabolites (e.g., sugars, soluble protein, amino acids, organic acids, and lipid metabolites), which in turn directly alter carbon skeletons and energy for secondary metabolite biosynthesis. When the air was treated with a high level of ozone (160 ppb), the amount of soluble protein in the leaves of Monarda didyma L. decreased by 20.9%[26]. A recent study in China found that after 60 d of treatment with a higher level of ozone in the air (40 ppb), the levels of lipids in sour jujube leaves decreased a lot, while the levels of gamma-aminobutyric acid (GABA) and L-glutamine significantly increased[46]. Indian scholars found that the proline of lemongrass (Cymbopogon flexuosus [Steud.] [Wats.]) was elevated after it was exposed to ozone for 90 d[37]. This shows that amino acids in medicinal plants do more than just build proteins. They also have many other important roles, such as helping with metabolism, sending signals, and protecting against stress[47]. This helps plants adapt easily to changes in their environment. Therefore, changes in the main metabolic products of traditional herbal medicines under ozone stress may alter their medicinal targets. This can occur through mechanisms such as covalent modification, transport competition, and signalling pathway regulation. It is important to pay more attention to this in the future.
Compared with the primary products of horticultural plants and crops, the secondary metabolites of medicinal plants have gained more attention as key active medicinal components. Ground-level ozone is an abiotic stressor, and plants can mitigate cell and tissue damage by synthesizing various secondary metabolites[7], thereby establishing defense responses to counteract this stress. Some secondary metabolites exhibit robust antioxidant properties, particularly in the scavenging of ROS. This efficacy is attributed to the varying ROS sensitivities and metabolic profiles across different cellular compartments. Ozone exposure significantly accelerated carbon flux toward secondary metabolic pathways, resulting in a shift in resources that changed the synthesis of secondary products[28]. On the basis of their biosynthetic pathways and chemical structures, plant secondary metabolites (PSMs) can be broadly classified into three categories: (1) terpenoids (plant volatiles, sterols, carotenoids, saponins, and glycosides), (2) phenolic compounds (flavonoids, phenolic acids, lignin, lignans, coumarins, stilbenes and tannins), and (3) nitrogen-containing compounds (alkaloids, glucosinolates and cyanogenic glycosides)[7]. Moderate environmental stress enhances the release of secondary metabolites such as polyphenols, flavonoids, and terpenes from medicinal and aromatic plants[6,28]. The effects of ground-level ozone stress on 12 typical medicinal species were reviewed (Table 2): Ginkgo biloba L., Melissa officinalis L., Tibouchina pulchra, Salvia officinalis L., Capsicum baccatum L., Hypericum perforatum L., Monarda didyma L., Sida cordifolia L., Costus pictus D. Don, Ziziphus jujuba Mill. Var. spinosa (Bunge) Hu ex H. F. Chou), Andrographis paniculata (Burm. f.) Nees, and Cymbopogon flexuosus (Steud.) (Wats.), the study area is located in India and China in Asia, Italy and Germany in Europe, and Brazil in South America. As shown in Fig. 3, this section aims to summarize the effects of ozone stress on key medicinal constituents, such as polyphenols, terpenoids, and alkaloids, and to explore their molecular response mechanisms and the effect of ozone dosage.
Table 2. Changes in secondary metabolites of medicinal plants under elevated levels of ozone.
Country Plants Medicinal parts Fumigation method Ozone treatment Secondary metabolite change Ref. Brazil Tibouchina pulchra (Cham.) Cogn. Leaves OTCs Ambient no-filtered air + 40 ppb for 8 h, 25 d, Anthocyanins (↑), total tannins (↓), total phenolics (↓), flavones/flavonols (↑) da Rosa Santos et al.[95] Brazil Capsicum baccatum Pericarps and seeds OTCs 80 ppb for 5 h, 62 d Pericarp: capsaicinoid (↓); dihydrocapsaicin (↓);
lutein (↓); β-Carotene (↑); total phenolics (↑);
Seed: capsaicinoid (↓); dihydrocapsaicin (↓); total phenolics (↑)Bortolin et al.[57] China Ginkgo biloba L. Leaves OTCs 80 ppb for 9 h, 110 d Condensed tannins (↓), isorhamnetin (↓), quercetin (↑), keampferol (↑), bilobalide (↑), ginkgolides C (↑), ginkgolides A (↑), ginkgolides B (↑) He et al.[58] China Monarda didyma L. Leaves OTCs 80, 120, and 160 ppb for 10 h, 14 d Abscisic acid (↑) Wang et al.[26] China Ziziphus jujuba Mill. Var. spinosa (Bunge) Hu ex H. F. Chou Leaves OTCs 95 ppb for 8 h, 60 d Organic acids (↑), nucleoside acids (↑), glycerophosphocholines (↓), lipids (↓), tiliroside (↓), kaempferitrin (↓), spinosin (↓), swertisin (↓) Fu et al.[46] Germany Melissa officinalis L. Leaves Growth chambers 80 ppb for 5 h Rosmarinic acid (↓) Döring et al.[97] Italy Melissa officinalis L. Leaves Growth chambers 200 ppb for 5 h Salicylic acid (↑), jasmonic acid (↑),
dehydroascorbate (↓), β-Carotene (↓), lutein (↓), violaxanthin (↑), total carotenoids (↓)Pellegrini et al.[45] Italy Salvia officinalis L. Leaves OTCs 120 ppb for 5 h, 90 d Lutein (↓), β-Carotene (↓), gallic acid (↑),
catechinic acid (↑), rosmarinic acid (↑), caffeic acid (↑)Marchica et al.[38] Italy Hypericum perforatum D. Don Apical shoots OTCs 110 ppb for 5 h Total phenols (↓), flavonoids (↑), anthocyanins (↑), condensed tannins (↓), quercetin (↑), hyperoside (↓), rutin(↑) Pellegrini et al.[39] Italy Salvia officinalis L. Leaves OTCs 200 ppb for 5 h Ethylene (↑), aalicylic (↓), jasmonic (↓), abscisic acid (↑) Marchica et al.[36] India Sida cordifolia L. Leaves, roots OTCs Ambient + 20 ppb, 90 d Leaves: α-thujene (↑), ar-turmerone (↓),
neophytadiene (↑), palmitic acid (↑), cis-linoleic acid (↑), e-phytol (↑), monopalmitin (↑), squalene (↑), tetracosane (↑), vitamin E (↑), phytyl palmitate (↑);
Roots: 1-decyne (↓), pelargonic acid (↓), calcium pelargonate (↓), tetradecane (↑), clonitazene (↑), n-hexadecanoic acid (↓), cholesterol (↑), cholesterol (↑)…Ansari et al.[30] Italy Salvia officinalis L. Leaves OTCs 200 ppb for 5 h, 36 d Coumaric acid (↓), caffeic acid (↓), danshensu (↑), rosmarinic acid (↑), salvianolic acid (↓), gallic acid (↑), vanillic (↑), syringic acids (↑), apigenin (↑), luteolin (↑), quercetin (↑) Marchica et al.[38] India Costus pictus D. Don Leaves, rhizomes OTCs 57 ppb for 5 h,120 d Leaves: thiols (↓), anthocyanins (↑), flavonoids (↓), ascorbic acid (↑), total phenolics (↑), phytosterol (↓), tannin (↑), lignin (↑), saponins (↑), alkaloids (↑);
Rhizomes: phytosterol (↑), tannin (↑), lignin (↑), saponins (↑), alkaloids (↑)Ansari et al.[54] India Sida cordifolia L. Leaves, roots OTCs 63 ppb for 5 h, 90 d Leaves: total phenolics (↑), thiols (↑), ascorbic acid (↓), phytosterol (↑), tannin (↓), alkaloids (↑), lignin (↑), saponins (↑);
Roots: phytosterol (↑), tannin (↑), alkaloids (↑), lignin (↑), saponins (↑)Ansari et al.[4] India Cymbopogon flexuosus (Steud.) (Wats.) Leaves OTCs 76.3 ppb for 5 h, 90 d Total phenolics (↑), anthocyanins (↑), flavonoids (↑), alkaloids (↑), ascorbic acid (↑) Madheshiya et al.[37] India Andrographis paniculata (Burm. f.) Nees Leaves OTCs Ambient + 20 ppb for
8 h, 135 dLignin (↓), saponins (↓), tannin (↑), phytosterol (↑), alkaloids (↑), andrographolide (↑) Ansari et al.[98] Note: '↑' indicates that the secondary metabolite content increased, and '↓' indicates that the secondary metabolite content decreased. 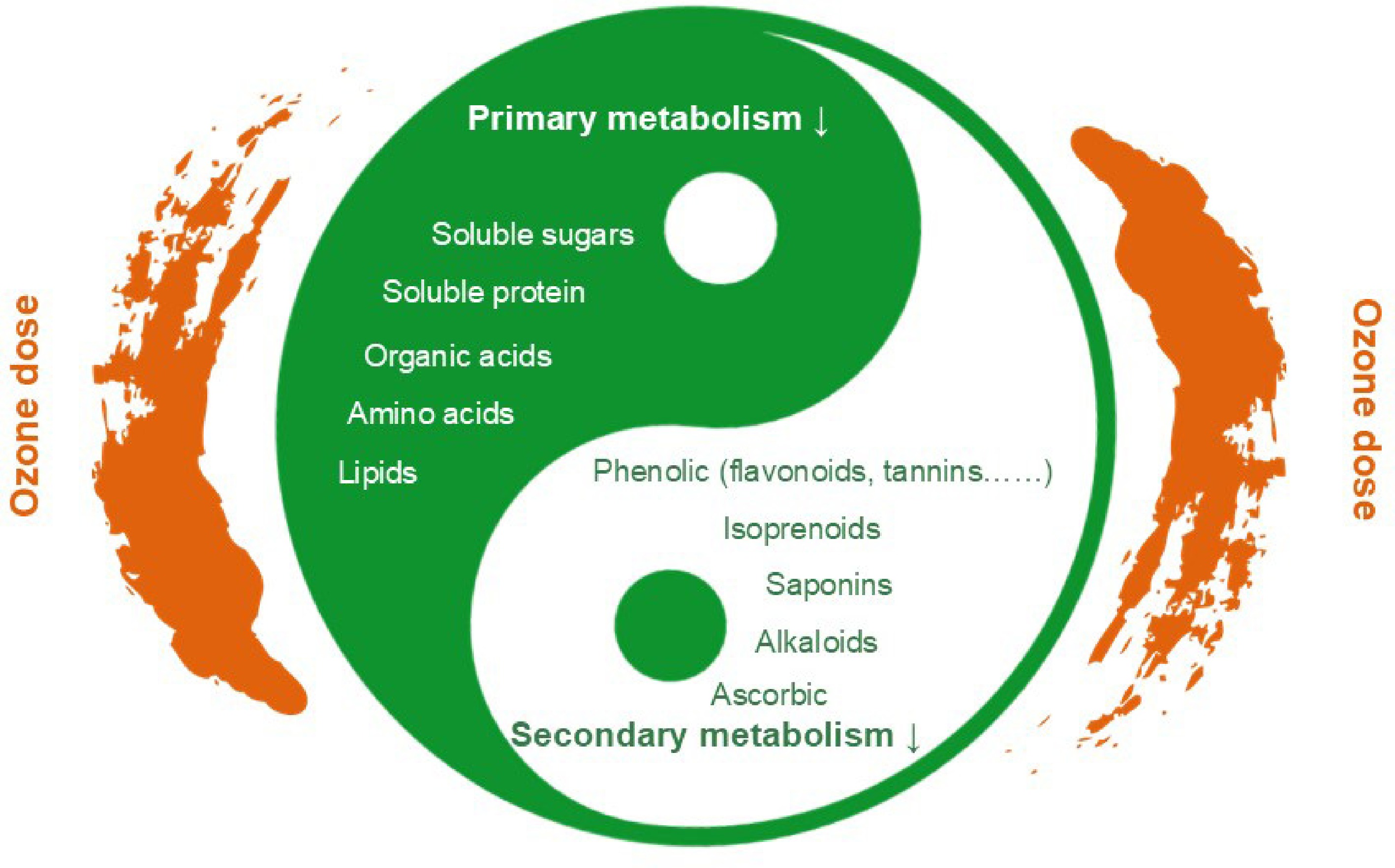
Figure 3.
Effects of ozone stress on medicinal plant metabolites. Note: ozone dose, the cumulative value of hourly O3 concentrations greater than 40 ppb.
Polyphenols are natural antioxidants found in many plants. They help prevent diseases like heart disease and diabetes, slow aging, and support skin and nerve health by getting rid of free radicals and stopping inflammation[48]. The phenolic compounds play a more significant role than do antioxidant enzymes in protecting plants against lipid and protein oxidation induced by ozone stress[49]. The total phenolic content of H. perforatum (St. John's wort) was increased under ozone stress at 110 ppb for 5 h[39]. The antioxidant activity of flavonoids arises from the catechol group on the B ring, where the hydroxyl group can directly bind to and neutralize ROS[50], forming stable products that help defend against lipid damage caused by ozone. This mechanism is more effective than the antioxidant enzyme system in addressing substance and protein oxidation. Additionally, O3 exposure stimulates phenylpropanoid metabolism and leads to the accumulation of anthocyanin-related compounds[51].
Terpenes are a class of biogenic volatile organic compounds (BVOCs) released by plants in response to abiotic stress[52]. It also has anti-inflammatory, antibacterial, anthelmintic, analgesic, and anti-tumor properties[53]. Together with other terpenes, they form one of the largest families of metabolites, collectively known as isoprenoids[24]. Isoprene can react with and neutralize ROS, resulting in antioxidant properties that help plants resist oxidative stress. Notably, various plant organs (such as leaves, flowers, and fruits) also produce different types of low-boiling, highly volatile hydrocarbons through secondary metabolic pathways. While isoprenes and monoterpenes constitute a significant portion of these compounds, the extensive release of BVOCs in many aromatic medicinal plants can lead to alterations in active medicinal components or a reduction in their therapeutic potency.
Alkaloids are a crucial class of plant secondary metabolites that play significant roles in the response of plants to adverse stress. Exposure to 60 ppb ozone for 90 d resulted in notable increases in lignin, saponin, and alkaloid levels in the leaves and roots of S. cordifolia[4]. After fumigation for 120 d under ozone at 60 ppb, the alkaloid contents in the leaves and rhizomes of C. pictus (Insulin plant). were still greater than those in the non-fumigation treatment[54]. It should be noted that some alkaloids are toxic and can adversely affect the cardiovascular, central nervous, and digestive systems[55], and ozone stress may also alter their pharmacological activity.
However, the impact of ground-level ozone stress on physiological processes is complex. Oxidative stress induced by ozone continuously consumes the secondary metabolites accumulated in medicinal plants at the early stage, which ultimately leads to a decline in the content of secondary metabolites. Agathokleous et al. reported that plants exhibit a broad range of hormetic dose-responses (HDRs) in response to ozone stress, which was defined as AOT40, the cumulative value of hourly O3 concentrations greater than 40 ppb[56]; specifically, low concentrations of ozone promote plant growth, which is considered a hormesis effect in plants, whereas high concentrations cause adverse effects on plants. It was similarly found that the response of secondary metabolites of medicinal plants to ozone dose effects was more pronounced. For example, after an elevated level of ozone (ambient + 20 ppb) fumigation for 90 d in open-top chambers (OTCs), the tannin content in the leaves of S. cordifolia decreases, whereas that in the rhizomes increases[4]. After fumigation with 120 ppb ozone for 30 d, the phenolic and monoterpene contents of S. officinalis initially increased but then decreased[38]. Bortolin et al. observed that continuous exposure to an ozone concentration of 170 ppb for 62 d led to a 20%–30% reduction in polyphenol content in pepper seeds[57]. The changes in secondary metabolite levels varied significantly with plant species, ozone fumigation duration, and ozone concentration. Furthermore, variations in the contents of specific compounds across different organs or tissues of the same plant are also inconsistent.
The secondary metabolites of medicinal plants result from long-term adaptation to their environment and serve as the active ingredients for treating diseases. Although more and more research has been done in recent years on how ozone affects the secondary metabolism of its medicinal plants, there are still very few studies focusing on the quality-indicating components specified in the pharmacopoeia. For example, the andrographolide in A. paniculata (king of bitter) was increased under elevated 20 ppb O3 after 45 d[39]. The andrographolide is an important active ingredient that helps to clear heat and remove toxins. It is also antibacterial and anti-inflammatory, and is an index ingredient stipulated in the Chinese Pharmacopoeia. A further study found that after 60 d of elevated 40 ppb O3, the spinosin content in the sour jujube (Ziziphus jujuba Mill. Var. spinosa (Bunge) Hu ex H. F. Chou) significantly decreased[46]. Spinosin is precisely the main sedative and tranquilising substance of ZSS. This provides a novel method for addressing the issue of declining spinosin levels in sour jujube plants in relation to their growth environment. Ginkgo has been designated the 'living fossil' of plants, and the content of ginkgolides can be elevated even after 110 d of fumigation at an ozone concentration of 80 ppb, further indicating that ancient plants possess a greater degree of environmental adaptability[58]. Excessive abiotic stress compels plants to utilize secondary metabolites, hindering normal growth and development; conversely, moderate stress enhances the accumulation of secondary metabolites. Given the predicted rising levels of ozone in the future, the active components of medicinal plants may change, potentially enhancing or inhibiting the medicinal properties of certain species. Future research should focus on how to utilize the beneficial effects of ozone and mitigate the detrimental effects of ozone on medicinal plants.
Plants regulate their responses by activating or repressing the expression of specific genes. For example, they modulate the biosynthesis of secondary metabolites by adjusting the activities of key enzymes and the expression levels of genes involved in secondary metabolic pathways[36]. Ozone stress stimulated the expression of antioxidant enzyme-related genes in Ginkgo biloba L. leaves, resulting in the upregulation of SOD, CAT, APX, glutathione thioltransferase (GST), and class III POD to varying degrees[59]. Under ozone stress, genes involved in hormone-mediated signaling, general defense mechanisms, redox regulation, signal transduction, transcription, and trafficking, along with those shaping genome-wide expression profiles, were upregulated[60]. Additionally, genes involved in alternative splicing may also play a potential role in linking to newly synthesized or metabolically active genes, such as chalcone isomerase, shikimate O-hydroxycinnamoyltransferase, and trans-cinnamate 4-monooxygenase[61]. The transcription factor families involved in the response to ozone stress include AP2-ERF, WRKY, JAZ, and MYB, which may be involved in cell regulation. The WRKY family may be crucial in the signaling mechanisms associated with plant responses to ozone. WRKY transcription factors are distinct plant transcription factors extensively involved in various physiological activities, including biotic and abiotic responses as well as signal transduction[44]. Marchica et al. identified six WRKY gene sequences in S. officinalis and reported that WRKY4, WRKY5, WRKY11, and WRKY46 were significantly upregulated after 2 and 5 h of ozone exposure[36]. McWRKY75 was identified as a key transcription factor affected by ozone stress, and its expression was positively correlated with that of flavonoid-related structural genes. Moreover, the ethylene response factors McERF019 and McERF109-like are also upregulated under ozone stress[62].
In summary, within the outer cell body, there is no single regulatory pathway governing gene expression in response to ozone. Instead, a combination of multiple pathways regulates gene expression and cell death induced by ozone stress. Yadav & Mina established the ozone stress-responsive gene database (OSRGD ver. 1.1)[63]. The database utilizes ozone stress-specific transcriptome and microarray datasets to investigate various plant species, primarily crops and horticultural plants (
www.osrgd.com ). Therefore, elucidating the molecular mechanisms underlying the effects of ozone stress on medicinal plants is imperative. -
Under the influence of ozone stress, the medicinal plants may promote allelopathic effects by releasing various secondary metabolites, thus influencing interactions among plants, animals, and microorganisms, which warrants further investigation. Although no studies addressing the effects of ozone stress on endophytes of typical medicinal plants in the published literature have been identified, it is believed to be essential to investigate this impact further, given the significant role of endophytes in the expression of active ingredients in medicinal plants. The diversity of endophytic bacteria with antioxidant activity in medicinal plants is notably high, particularly in actinomycetes (Methylobacterium radiotolerans)[64]. Huang et al. isolated 292 species of endophytic bacteria from 29 plant species, all of which mainly produce active antioxidant substances and phenolic substances[65]. Under environmental stresses, plants can influence their endophytes by synthesizing or secreting various metabolites. Conversely, endophyte communities may also impact the secondary metabolism of the host plant. The diversity and community characteristics of endophytes are influenced by environmental factors such as the physiological state of medicinal plants, season, temperature, and humidity, which serve as indicators of the host's physiological state[66]. Oscillations in ozone intensity may alter the strength and direction of species interactions. Plant adaptation to ozone stress can be enhanced by symbiotic microbes. An increasing body of research has demonstrated that endophytes can improve host adaptation to stresses such as high temperature, drought, ozone, and pests by increasing the activity of antioxidant enzymes, scavenging free radicals, and producing active antioxidant substances[67−69].
Under high ozone exposure, the endophytic fungi of Euonymus japonicus L. fail to maintain community richness and diversity, leading to the dominance of pathogenic fungi and a reduction in bacterial abundance, which poses a threat to plant health[70]. Another study found that ozone stress did not significantly change the taxonomic composition of fungal mycobiomes, such as Leotiomycetes and Phaeomoniella. These results suggest that fungal communities play a role in mitigating the oxidative burst caused by ozone[71]. Endophytic microbes can also colonize various plant organs, including the stem, leaf, curd, and carpel ring, thereby colonizing seeds[72,73]. As important reproductive organs of plants, seeds serve as carriers of various beneficial bacteria that can be transmitted from generation to generation through vertical transmission, promoting the adaptation of plants to local biotic and abiotic conditions. Seed endophytes play crucial ecological roles as the first microbiota to colonize new seedlings[74]. Studies indicate that elevated ozone concentrations reduce the survival time of endophytes in plant seeds and that maternal endophytes are vertically transmitted to offspring, increasing seedling survival rates[75,76]. Furthermore, endophytes not only produce secondary metabolites akin to those of host plants but also metabolize secondary compounds produced by these hosts, potentially influencing the accumulation of unique active components in certain medicinal plants[77].
In addition to the ability of endophytes to colonize various tissues of medicinal plants, alterations in rhizosphere microbes warrant attention. Rhizosphere microbes play key roles in information exchange and environmental adaptation and are often referred to as the 'second genome' of plants[78]. Rhizosphere microorganisms regulate the expression of genes related to secondary metabolite synthesis and increase the activity of key enzymes involved in the production of specific chemical components. Moreover, ground-level ozone stress adversely impacts both foliar chemistry and soil processes in plants. These chemical processes govern plant–microbe–soil interactions, including litter mass, turnover of plant root exudates, soil enzyme activity, and nutrient cycling, all of which are detrimental to soil ecosystem health[79]. Overall, research on the effects of ozone on plant–microbiota interactions and biodiversity is largely restricted to rhizosphere processes, with few studies addressing both above- and below-ground components as an integrated system[80]. Rhizosphere microbes are interconnected within the 'microbe–root–shoot' loop, facilitating compensatory regulation of plant growth, albeit at the cost of some defensive capabilities[6].
Consequently, investigating how plant versatility is influenced by the structure and function of the rhizosphere microbial community is crucial, as it may provide new insights into the induction and quality response mechanisms of the antioxidant system in medicinal plants. Importantly, the host plant is a critical factor in shaping the richness and composition of the symbiotic microbial flora. Plants can selectively favor beneficial colonizing endophytes, influencing the roles of various endophytes in conferring resistance to ozone stress. Despite these variations, future efforts should focus on isolating and identifying additional symbiotic microorganisms that exhibit resistance to ozone stress, as well as applying synthetic microbial communities (SynComs) to the cultivation of medicinal plants[81].
-
Most traditional studies have focused primarily on mesoscopic scales, such as photosynthesis, growth indicators, and physiological and biochemical parameters. In the open-top chamber (OTC) method, the top of the chamber is open to the atmosphere, creating a small environment that closely mimics natural conditions. This method has proven effective and has been widely used in recent decades because of its high precision, consistent internal conditions, and reliable results[82]. Currently, most studies on ozone stress in medicinal plants are conducted as pot experiments in open-top chambers (OTCs), and the research methods are relatively limited. Free-air controlled exposure (FACE) experimental technology effectively simulates variations in ozone levels and other atmospheric components within natural ecosystems. A high-concentration gas field is created by introducing elevated levels of ozone gas directly into the experimental area to investigate its effects on medicinal plant growth, physiological and ecological responses, and overall yield. Compared with traditional greenhouse or OTC experiments, FACE experiments more closely mimic natural conditions and provide a more accurate simulation of crop growth under anticipated climate change scenarios[83]. Regardless of the experimental type, the study durations were relatively brief, ranging from 30 d to two to three years. Since herbs are often harvested on a perennial basis, the results of short-term experiments do not reflect long-term changes. Therefore, long-term ozone fumigation experiments are essential for field studies, as they are the most effective way to collect the critical empirical data necessary for modelling the effects of ozone stress on primary and secondary metabolic pathways in plants.
However, plants typically respond to ozone stress by integrating various signaling pathways and molecular regulatory networks, allowing them to adapt positively to stress. Owing to advancements in sequencing technologies and methodologies, statistical methods characterized by high throughput, large scale, and high sensitivity have increasingly been applied in the comprehensive analysis of adverse stress[3]. These analytical objectives can be categorized into five main areas: genomics, transcriptomics, proteomics, epigenetics, and metabolomics. To further investigate this systemic response mechanism, integrated multiomics approaches are essential[84]. The metabolome directly reflects the functional interactions between the microbiota and the host, with both components complementing each other[77,85]. In the distant future, gene editing (e.g., CRISPR/Cas9) and molecular biology will be combined to systematically study the expression, regulation, modifications, and interactions of key genes[86], and the changes in metabolites in the primary metabolite synthesis and catabolism pathways of medicinal plants under ozone stress. These findings help us to understand exactly how these genes respond to ozone stress.
Data derived from single-component experiments are unreliable for predictions, as ecosystem responses to interacting global changes can vary significantly depending on individual factors and their combinations. Salt stress, elevated temperatures, and the interaction between increased atmospheric CO2 concentrations and ozone exert significant effects on plants[87,88]. Combined stress typically results in three types of effects on plants: synergistic, antagonistic, or independent effects. However, the antagonistic or synergistic effects of compound stress are not absolute. The rapid increase in ozone concentration is often driven by ample light and high temperatures, frequently coinciding with drought conditions, which expose plants to the dual stresses of elevated ozone concentrations and drought simultaneously[89,90], making this a prominent area of current research. Although drought can mitigate ozone damage at the leaf level by promoting stomatal closure, it reduces carbon fixation. When considering the plant as a whole, drought does not alleviate ozone stress but rather exacerbates biomass loss[91]. Several studies have indicated that elevated CO2 mitigates the O3-induced reduction in photosynthetic activity[87], likely due to an increase in carbon assimilation products. In a study of M. crystallinum, salt stress alleviated the oxidative stress caused by elevated ozone concentrations. This effect benefits from the increase in SOD activity, increased antioxidant compounds, and reduced production of O2−·[33]. There are definite interspecies differences in the interactions between ozone and other environmental factors, which are determined by the amount of ozone entering the cells or the types and activities of plant antioxidant enzymes stimulated by adverse stress[92]. It is widely accepted that plant secondary metabolites increase to varying degrees in response to environmental stresses. However, the synergistic effects of multiple stresses may be influenced by the plant's physiological state, as well as the duration and intensity of the stress. Consequently, future research should aim to elucidate how various climatic factors impact net primary productivity and carbon sequestration and their subsequent effects on the synthesis of secondary metabolites in medicinal plants under fluctuating climatic conditions.
As demonstrated in Fig. 4, in response to increasing ozone stress, scholars have implemented measures to mitigate ozone damage through various means, such as cultivated varieties, water and fertilizer management, and exogenous spraying of antioxidants[25]. Numerous studies highlight ethylenediurea (EDU) as the most promising compound for alleviating the adverse effects of ozone on crops. Feng et al. demonstrated that EDU significantly reduced visible ozone damage by 76%[93]. At a concentration of 200–400 mg·L−1, EDU as a soil drench solution had the most pronounced beneficial effect on field crops. Under ozone stress, the yield of crops treated with EDU was on average 15% greater than that of those without EDU treatment. However, the use of synthetic protectants pollutes the environment and may cause dose-dependent toxicity to vegetation. Therefore, there is an urgent need to develop alternative and eco-friendly antiozonants, including natural plant antioxidant-based compounds such as ascorbic acid and its derivatives, phytohormones, flavonoids, and polyamines[94]. In addition to the established functions of NO, polyamines, and defense hormones in combating environmental stressors such as ozone, as multiomics technologies have advanced, a growing array of antistress signaling molecules have been characterized for mitigating ozone impacts. For example, stress molecules such as γ-aminobutyric acid (GABA) can reduce wheat grain yield by 10% under elevated ozone conditions[61]. In addition to merely maximizing agronomic practices to increase ozone resistance, including irrigation, fertilization, and tillage, there is a growing acceptance of ozone-tolerant medicinal varieties. Notably, the implementation of these measures should consider the specific growth conditions and environmental factors of medicinal plants to ensure their efficacy and sustainability. Concurrently, the effects of ozone stress on medicinal plants require rigorous monitoring and research. It is also very important to keep improving and perfecting how the fields are managed, especially by using antioxidants in the right way. This will help to stop negative effects on the environment. In the future, it will be necessary to combine pharmacological and toxicological methods to systematically assess the effects of changes in the content and proportion of active ingredients in medicinal plants under ozone stress on their overall efficacy and safety. This will provide a basis for the development of scientific standards for the quality of traditional Chinese medicinal herbs and cultivation practices.
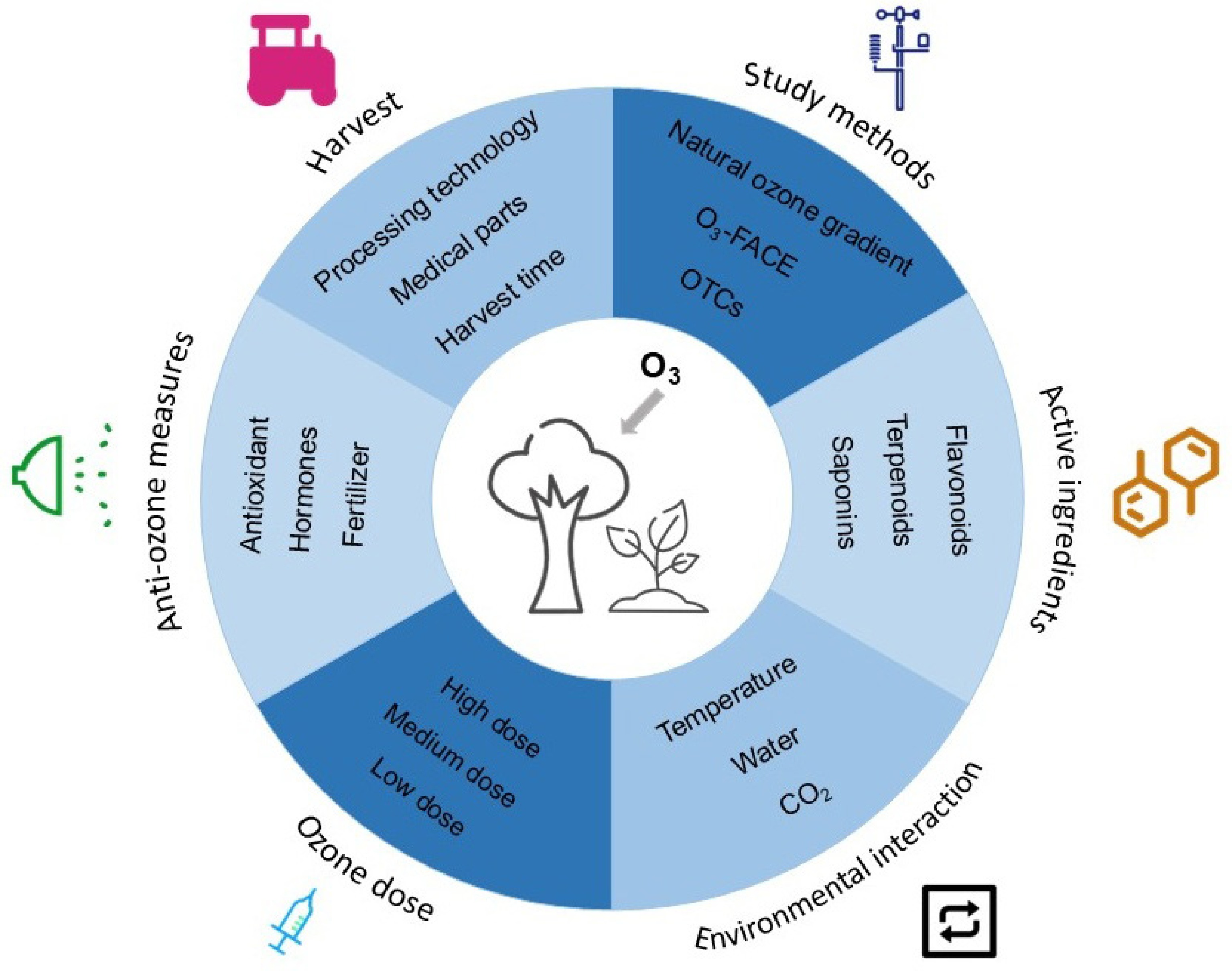
Figure 4.
Future work on medicinal plants under ozone stress.
-
Ozone has multiple effects on the physiology and ecology of medicinal plants, including both direct and indirect damage to plant leaves, impacts on photosynthesis and respiratory metabolism, and the induction of plant defense mechanisms. Although some studies have investigated the effects of ozone on several medicinal plants, more investigations are essential to provide a comprehensive understanding of the long-term effects of ozone stress on medicinal plants. Furthermore, different medicinal plant species exhibit varying sensitivities to ozone, with these sensitivities frequently interacting with other environmental factors. Future research directions should encompass: (1) enhancing the examination of the impacts of ozone stress on plant physiology at both the individual and community levels; (2) investigating the mechanisms of gene regulation and associated signaling networks in plants exposed to ozone; and (3) developing new medicinal plant varieties to improve resilience against ozone stress. Additionally, future research must comprehensively assess the ecological impacts of ozone stress to establish a scientific basis for environmental protection policies. In conclusion, managing ozone stress is crucial for preserving the stability of traditional herbal medicines.
This research was supported financially by the National Key Research and Development Program of China (Grant No. 2023YFC3503801 and 2023YFC3503502); the Scientific Research Plan of Hebei Provincial Administration of Traditional Chinese Medicine (Grant No. 2025416); the Science and Technology Research Program of Higher Education Institutions in Hebei Province (Grant No. QN2023026); the Innovation Team of Hebei Province Modern Agricultural Industry Technology System (Grant No. HBCT2024110201 and HBCT2024110205).
-
The authors confirm contributions to the paper as follows: study conception and design: Gu X, Li C; data collection: Fu X, Ma D; analysis and interpretation of results: Gu X, Yan Y, Zheng Y; draft manuscript preparation: Gu X, Fu X, Ma D; supervision: Guo L; writing, review and editing: Yan Y, Zheng Y, Guo L, Li C. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gu X, Fu X, Ma D, Yan Y, Zheng Y, et al. 2025. Ground-level ozone stress in medicinal plants: response mechanisms and adaptive strategies. Medicinal Plant Biology 4: e039 doi: 10.48130/mpb-0025-0036 

Ground-level ozone stress in medicinal plants: response mechanisms and adaptive strategies
- Received: 16 May 2025
- Revised: 05 September 2025
- Accepted: 16 September 2025
- Published online: 11 December 2025
Abstract: Ground-level ozone (O3) has strong oxidative properties and is a significant threat to the health of humans, animals, and plants. Currently, in comparison with studies on crops and horticultural plants, studies evaluating the impact of ozone stress on medicinal plants are relatively rare, and the methodologies employed are somewhat limited. Ozone stress can impact medicinal plant growth and reproduction at multiple levels, including the community, whole plant, and cellular levels, potentially leading to a decline in the quality of medicinal materials. This study systematically reviewed the effects of ground-level ozone on the phenotypic changes in the leaves, flowers, fruits, and seeds of medicinal plants, as well as its impact on harvest timing. Furthermore, a comprehensive analysis was carried out at the level of antioxidant defense, secondary metabolite production, gene regulatory networks, and symbiotic microbial interaction mechanisms. This review advocates the use of diverse research methods to enhance the effects of ozone stress on the active constituents of medicinal plants, considering the combined effects of ozone and other abiotic factors, such as drought, salinity, and high temperatures. It is imperative to elucidate the molecular regulatory mechanisms underlying the tolerance of various types of medicinal plants, including trees, shrubs, and herbaceous species, to develop targeted production management strategies against ozone stress. It also aims to clarify the impacts of ozone stress on medicinal plants and to offer insights for their cultivation and conservation efforts.