









-
Domesticated pigs are among the most important livestock breeds; they provide most of the meat consumed by humans worldwide and are important in the daily life of humans. Breeding high-quality boars is an effective way to improve population genetics and increase the production efficiency of the porcine industry, as it directly reflects the ability to produce the offspring necessary to offset costs in modern pig production systems. Determining production performance is a goal of boar breeding because it can not only be used to screen excellent breeders and provide strong support for the improvement and development of boar breeds but also provide excellent offspring to the market and promote pork production. Back fat thickness, a common metric for subcutaneous fat, is inversely related to meat's leanness[1]. Excessive subcutaneous fat leads to a reduced lean meat rate, resulting in less edible meat and consequently diminishing the profitability of pig farming[2]. Hence, reducing fat content and increasing the lean meat percentage are central objectives in commercial pig breeding. Additionally, previous studies have shown that testicular volume impacts the age of puberty and the postpartum interval in genetically linked females[3,4]. Testicular weight has also been found to be positively correlated with the sperm count in the cauda epididymis of Nigerian Sahel goats[5], suggesting a connection between testicular size and sufficient sperm reserves for reproductive efficiency. Therefore, selecting for testicular size could be a crucial strategy to accelerate breeding processes and ensure the reproductive capacity of sires. In regions where advanced semen assessment technology and laboratory support are unavailable, farmers may need to rely on testicular size as a proxy for assessing male fertility. Overall, it is crucial to select boars with larger testicular sizes and lower back fat thicknesses as breeding candidates. However, in practical breeding situations, selecting individuals with larger testicles as backup pigs can be performed through visual inspection alone. However, for other production performance indicators, such as fat and lean meat percentages, precise selection based solely on visual assessment poses a significant challenge.
The gut of mammals is home to trillions of microbiota that play crucial roles in reproduction, metabolism, immunity, and behavior[6−9]. Recent studies have revealed correlations between gut microbial profiles and various parameters in pigs, including reproduction and productivity[10−12]. For example, certain gut microbes are capable of affecting testosterone synthesis and metabolism. Testosterone, the main male sex hormone produced by Leydig cells in the testes, is crucial for testicular development and maintenance, as well as for fostering reproductive organ growth and sperm production[9]. Insufficient testosterone production may result in testicular dysplasia, which is characterized by reduced testicular volume[13]. Imbalances in the gut microbiota can lead to abnormal testosterone levels, thereby impacting testicular function[13]. Moreover, a healthy gut microbiota aids the host in nutrient absorption and utilization, providing the necessary nutritional support for the host's growth[14]. Conversely, an imbalanced gut microbiota may cause nutrient malabsorption, indirectly affecting production performance and testicular size. Hence, analysis of the gut microbiota is a key research field for enhancing the production efficiency of the porcine industry. Junmu No. 1 white pigs are hybrids of Chinese Sanjiang hybrid sows and Belgian Seghers hybrid boars and have a fast growth rate, a high percentage of carcass lean meat, plump and prominent legs and buttocks, obvious double-muscle buttocks and strong adaptability. However, the relationships among testicular development, production performance, and the gut microbiota in Junmu No. 1 white boars (JN1WBs) remain poorly understood. In the present study, 41 170-day-old JN1WBs were divided into two groups according to their testicular index to (1) determine the primary differences in testicular development indices and production parameters between JN1WBs with high and low testicular indices, (2) identify and compare the fecal microbiota composition between JN1WBs with high and low testicular indices, and (3) elucidate the relationship of the fecal microbiota with testis development and production performance in JN1WBs. Elucidating these characteristics not only provides a reference for investigating the relationships among the fecal microbiota, testicular development, and production performance but also offers theoretical support for the preliminary selection of sires for breeding on the basis of the fecal microbiota from sexually mature JN1WBs.
-
Forty-one 80-day-old healthy JN1WBs with similar initial body weights (IBW) (mean ± standard error of the mean [SEM]: 35.33 ± 1.70 kg) were randomly divided into five pens with eight or nine boars per pen. The animals were raised at the Agricultural Experiment Base of Jilin University, China, for 90 days under uniform conditions. A commercial formula diet (Table 1) and water were provided ad libitum throughout the trial. On the final day of the experiment, at an age of 170 days, which generally encompasses the late stage of puberty, the boars were transported from the farm to a commercial slaughterhouse, where they were fasted for 12 hours prior to slaughter.
Table 1. Ingredients and nutrient composition of the commercial diet provided to the boars.
Item Body weight (kg) 30–60 > 60 Ingredients Corn (%) 61.00 65.00 Soybean meal (%) 15.30 7.50 Wheat bran (%) 15.00 15.00 Corn gluten meal (%) 5.00 9.00 Limestone powder (%) 1.50 1.50 Calcium hydrogen phosphate (%) 0.90 0.70 Sodium chloride (%) 0.30 0.30 Premix compound (%)a 1.00 1.00 Total (%) 100 100 Nutrient
compositionDigestible energy (MJ/kg)b 13.17 13.33 Crude protein (%) 16.15 14.16 Calcium (%) 0.81 0.79 Total phosphorus (%) 0.56 0.49 Total lysine (%) 0.65 0.49 Total methionine + cystine (%) 0.42 0.35 a The premix provided the following per kilogram of diet: Vitamin A, 8,000 IU; Vitamin B1, 4 mg; Vitamin B2, 3.6 mg; Vitamin B5, 40 mg; Vitamin B6, 4 mg; Vitamin B12, 0.02 mg; Vitamin D3, 3,000 IU; Vitamin E, 20 IU; Vitamin K3, 2 mg; biotin, 0.15 mg; folic acid, 1 mg; nicotinic acid, 10 mg; D-pantothenic acid, 11 mg; antioxidant, 100 mg; Cu, 10 mg (as copper sulfate); Fe, 80 mg (as ferrous sulfate); Mn, 30 mg (as manganese sulfate); Zn, 75 mg (as zinc sulfate); I, 0.4 mg (as potassium iodide); and Se, 0.3 mg (as sodium selenite). b The digestible energy was calculated, while the others were analyzed. Sample collection
-
The final body weight (FBW, in kg) and total feed intake (TFI, in kg) of the boars were recorded. These data were used to calculate body weight gain (BWG, in kg), average daily gain (ADG, in kg/day), and the feed conversion ratio (FCR, in kg/kg). Fresh fecal samples from the boars were collected in sterile 5-mL polypropylene tubes, with care taken to avoid contact with urine, soil, or other contaminants to prevent cross-contamination. Immediately after collection, the samples were stored at –80 °C for subsequent DNA extraction. Following slaughter, measurements were taken for testicular weight (TW, kg), testicular length (cm), testicular width (cm), carcass weight (CW, in kg), fat mass weight (FMW, in kg), lean meat weight (LMW, in kg), eye muscle length (cm), and eye muscle width (cm). These measurements were used to determine the testicular index (TI, in %), testicular volume (cm3), fat mass percentage (FMP, in %), lean meat percentage (LMP, in %), and eye muscle area (EMA, in cm2). The commercial slaughtering process included electroshock, bloodletting, scalding, hair removal, head and foot removal, gutting, washing, and cooling. Furthermore, the testes and serum were also obtained from each boar and stored at –80 °C for analysis of testosterone levels.
Testicular development and production performance analysis
-
Testosterone levels were determined via a commercially available ELISA kit (ENZO Life Sciences Inc., NY, USA) following the protocol provided by the manufacturer. In the case of testicular tissue, the samples were initially homogenized in a 1.15% KCl solution at a ratio of three parts solvent to one part tissue (w/v). This homogenate underwent centrifugation at 9,000 × g for 20 min at 4 °C, followed by an additional centrifugation step at 105,000 × g for 60 min at the same temperature. The resulting supernatant was then subjected to two rounds of extraction via diethyl ether to isolate testosterone for quantification. The results for testosterone in the tissue samples are presented as the quantity per 1 g of testicular tissue. In contrast, the serum samples were analyzed directly without any preliminary processing steps. The ELISA kit for testosterone testing spans a range from 7.81 to 22,000 pg/mL and has a sensitivity level of 5.67 pg/mL. The intra-assay variability coefficients for the serum and testicular testosterone measurements were 6.80% and 6.75%, respectively, as determined by analyzing a single boar sample on 20 occasions throughout a standard testing procedure.
The BWG, ADG, FCR, TI, testicular volume, FMP, LMP, and EMA were calculated via the following formulas: BWG (kg) = FBW (kg) – IBW (kg); ADG (kg/day) = BWG (kg)/90 (day); FCR (kg/kg) = TFI (kg)/BWG (kg); TI (%) = TW (kg)/FBW (kg) × 100; testicular volume (cm3) = 4 × πab2/3 (here, a represents half of the testis length, whereas b represents half of the testis width)[15]; FMP (%) = FMW (kg)/CW (kg) × 100; LMP (%) = LMW (kg)/CW (kg) × 100; and EMA (cm2) = eye muscle height (cm) × eye muscle width (cm) × 0.7. Finally, the JN1WBs were divided into two groups according to the TI: the low testicular index (LT, n = 21) group and the high testicular index (HT, n = 20) group.
DNA extraction, microbiota analysis, and functional prediction
-
The total genomic DNA in the fecal samples was extracted via a Magnetic Soil and Stool DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer's protocol. The DNA concentration and purity were assessed via 1% agarose gel electrophoresis. The DNA was diluted to a concentration of 1 ng/µL with sterile water. The 16S rRNA hypervariable V3–V4 regions were amplified with the primers 341F (CCTAYGGGRBGCASCAG) and 806R (GGACTACNNGGGTATCTAAT)[16]. An equal volume of 1× loading buffer containing SYBR Green was mixed with the polymerase chain reaction (PCR) products, and the DNA was assessed via electrophoresis on a 2% agarose gel. The PCR products were mixed in equal proportions and then purified via the QIAquick Gel Extraction Kit (Shanghai Canspec Scientific Instruments Co., Ltd., Shanghai, China). Finally, the amplicon libraries were sequenced on the Illumina NovaSeq platform, and 250-bp paired-end reads were generated. The generated reads were evaluated via QIIME2 (version 2-202006). Next, the DADA2 plugin was used to denoise low-quality reads and to merge high-quality paired-end clean reads into tags with FLASH
[17], followed by chimera removal[18] to obtain the initial amplicon sequence variants (ASVs); after that, ASVs with abundances less than 5 were filtered out[19]. Species annotation was performed via QIIME2 software according to the Silva database (release 138.1, www.arb-silva.de ). The alpha diversity in the sample was calculated on the basis of the rarefied ASVs from four indices in QIIME2, namely, the Chao1, Shannon, Pielou's evenness (Pielou_e), and Simpson indices. Beta diversity was calculated on the basis of unweighted UniFrac distances in QIIME2 and displayed via principal coordinate analysis (PCoA) in R software (version 2.15.3). Community structure similarities between groups were analyzed via the anosim function, which is based on unweighted UniFrac distances with 999 permutations in QIIME2 software. Species differences at each taxonomic level (top 10 phyla, genera, and species) were analyzed using R software (version 3.5.3) via MetaStat. This methodology involves performing hypothesis tests on taxa's abundance data across groups to derive p-values, with a significance threshold set at p < 0.05. Thereafter, taxa demonstrating statistically significant differences between the groups are pinpointed on the basis of these p-values. The predicted functional features of the communities in the samples were analyzed with PICRUSt2 software (version 2.1.2-b) on the basis of the 16S rRNA sequencing data[20].Relationships of the fecal microbiota with testicular development indices and production parameters
-
To investigate the associations between the fecal microbiota and both testicular development indices and production parameters, we utilized R (version 2.15.3) to calculate the Spearman's correlation coefficients and assess their significance.
Statistical analysis
-
The normality of the data distribution was assessed via the Shapiro‒Wilk and Kolmogorov‒Smirnov tests. Significant differences between groups were determined via either a two-tailed unpaired t-test or the Mann‒Whitney U-test, depending on the data distribution, with analyses conducted in GraphPad Prism software (version 8). The data are presented as the means ± SEMs. Significance levels are denoted by asterisks as follows: *p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
-
Compared with the LT group, the HT group presented significantly greater values for several key metrics. Specifically, the TI and TW were significantly greater in the HT group (p < 0.0001 and p < 0.0001, respectively) (Fig. 1a,b). Similarly, the left testis volume (p = 0.003), right testis volume (p = 0.001), and total testicular volume (p < 0.0001) were also significantly greater in the HT group (p < 0.0001) (Fig. 1c–e). Additionally, the serum testosterone (p = 0.049) and testicular testosterone (p = 0.039) levels were markedly elevated in the HT group compared with those in the LT group (Fig. 2a,b). Moreover, compared with the LT group, the HT group presented significantly greater LMP but lower FMP (p = 0.014 and p = 0.014, respectively) (Fig. 3e,f). However, no significant differences were detected between the HT and LT groups in terms of IBW (p = 0.338), FBW (p = 0.640), EMA (p = 0.458), ADG (p = 0.454), or FCR (p = 0.614) (Fig. 3a–d,g).
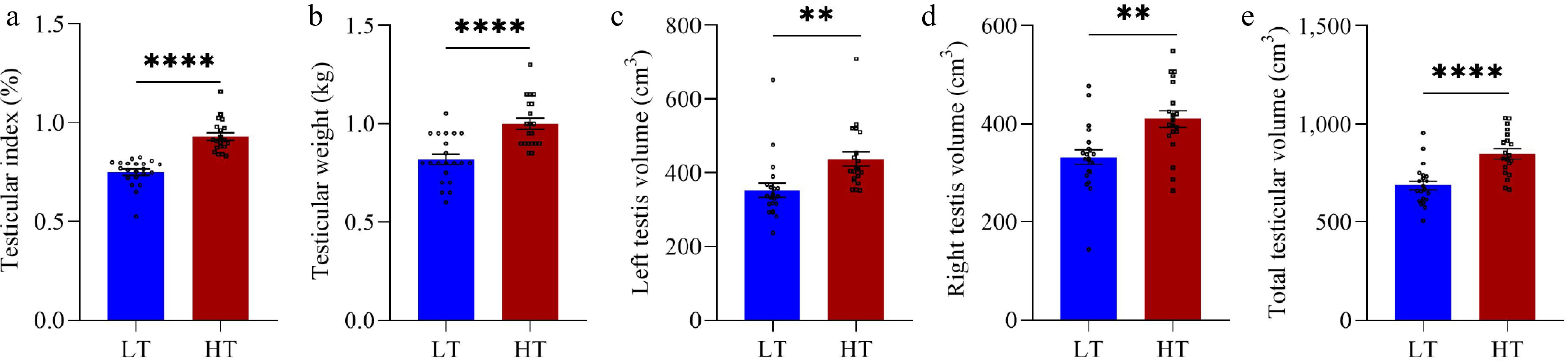
Figure 1.
Testicular development indices between boars in the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups. Testicular index (a), testicular weight (b), left testicular volume (c), right testicular volume (d), and total testicular volume (e) were measured on the final day of the experiment. The vertical bars represent the means ± SEMs. ** p < 0.01 and **** p < 0.0001 indicate significant differences between the LT and HT groups.
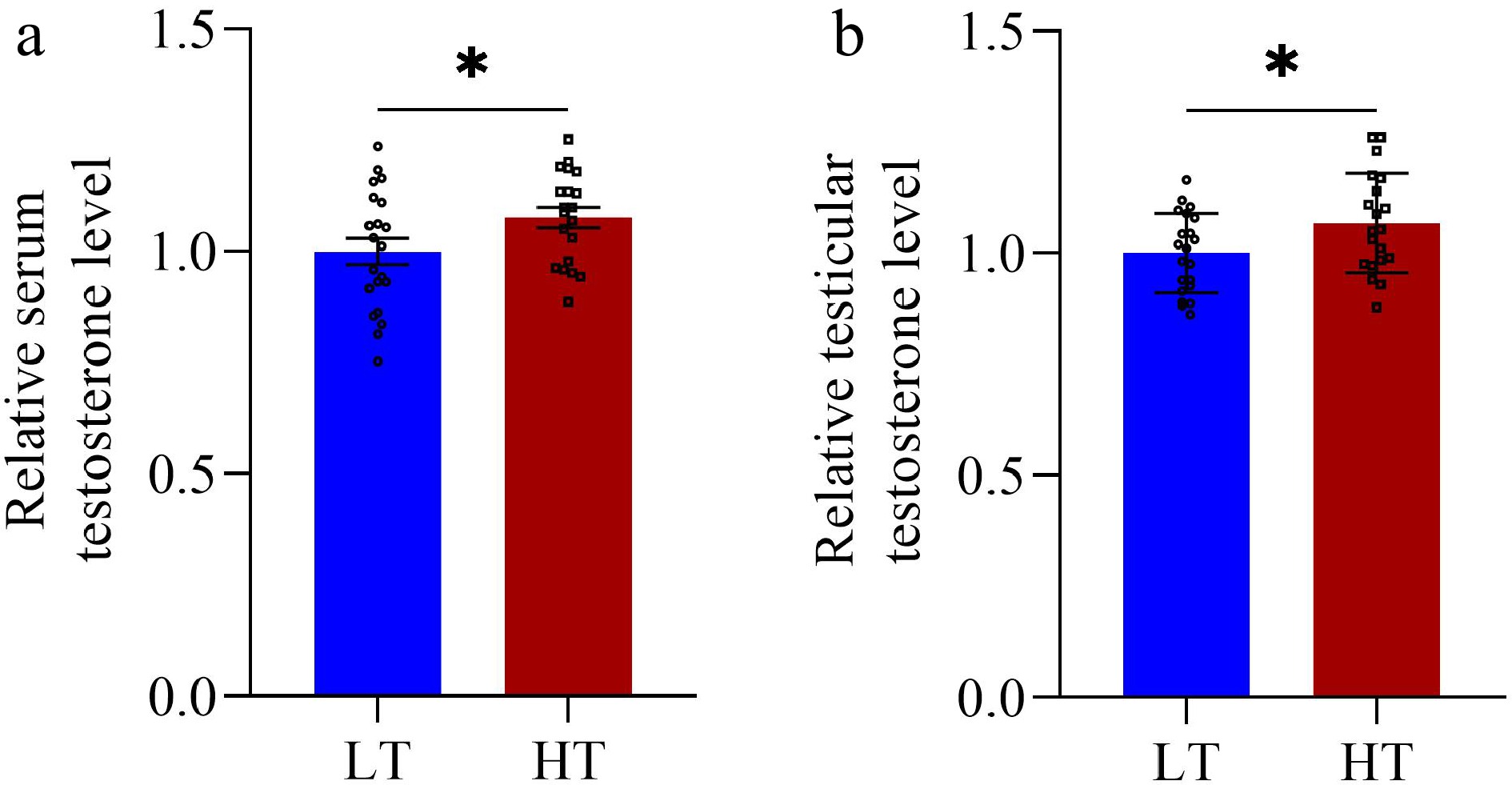
Figure 2.
Testosterone levels between boars in the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups. The relative levels of serum testosterone (a) and testicular testosterone (b) were measured on the final day of the experiment. The vertical bars represent the means ± SEMs. * p < 0.05 indicates a significant difference between the LT and HT groups.
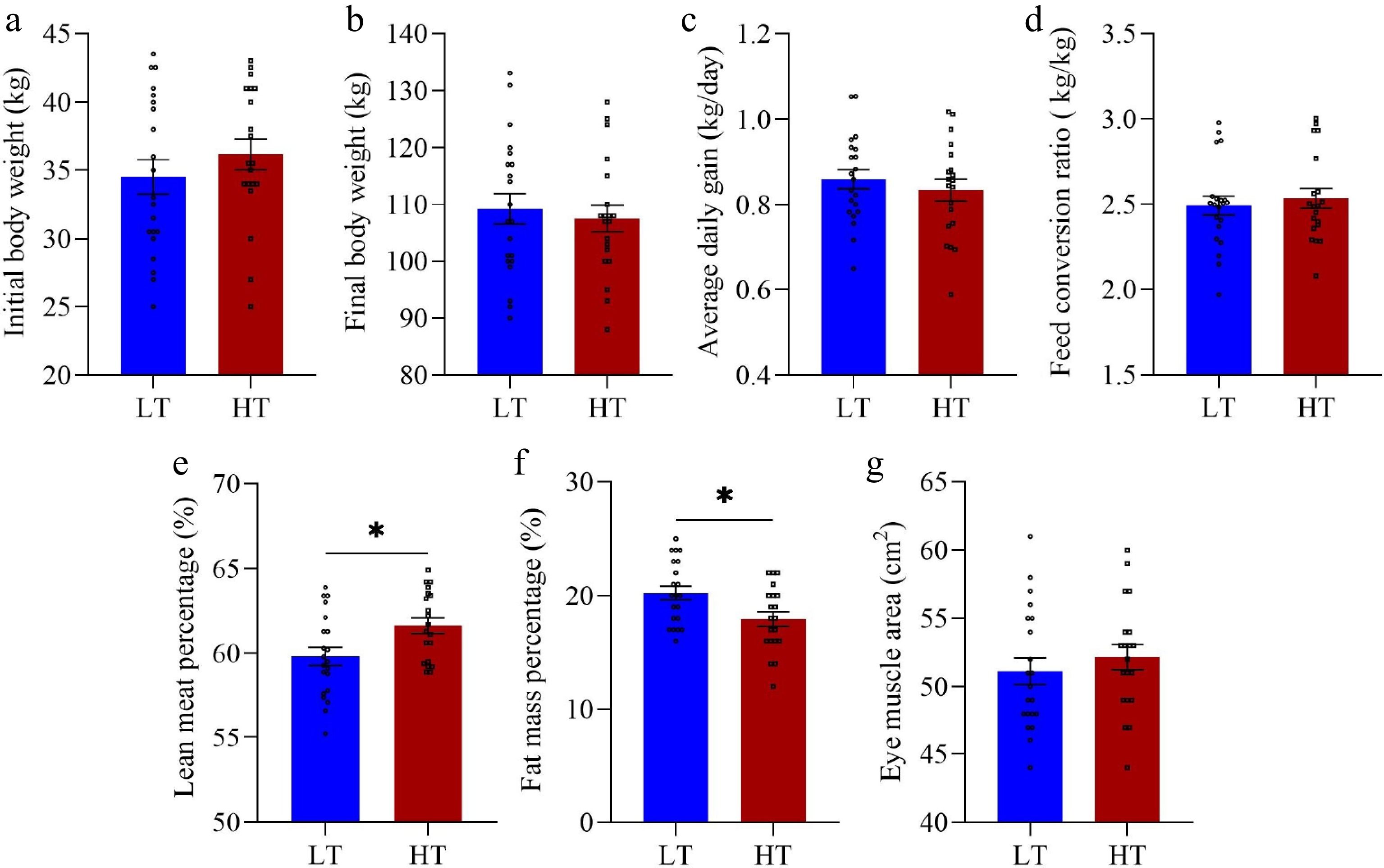
Figure 3.
Differences in production performance between the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups of boars. Initial body weight (a), final body weight (b), average daily gain (c), feed conversion ratio (d), lean meat percentage (e), fat mass percentage (f), and eye muscle area (g) were measured on the final day of the experiment. The vertical bars represent the means ± SEMs. * p < 0.05 indicates a significant difference between the LT and HT groups.
Differences in fecal microbiota composition and functional prediction between LT and HT boars
-
The microbiota of the fecal samples was analyzed via 16S rRNA gene sequencing. In total, 3,143,903 high-quality reads were obtained from 41 fecal samples (with an average of 76,680 reads per sample). The increase in total ASVs tended to stabilize as the sequence number increased (Fig. 4a). The observed ASVs (p = 0.008) (Fig. 4b) and Chao1 index (p = 0.008) (Fig. 4c) in the fecal samples of LT boars were significantly greater than those in the fecal samples of HT boars, but there were no significant differences in the Pielou_e (p = 0.315), Shannon (p = 0.197), or Simpson (p = 0.334) indices between the LT and HT groups (Fig. 4d–f). Furthermore, PCoA and unweighted UniFrac dissimilarity distance analysis (Fig. 4g) revealed significant differences in the microbial composition of the feces between the LT group and the HT group (p = 0.005). Specifically, the LT group presented significantly greater abundances of Campilobacterota (p = 0.046) and Streptococcus (p = 0.003) but lower abundances of Lactobacillus (p = 0.044) and Anaerovibrio sp. (p = 0.028) than did the HT group (Fig. 5a–g).
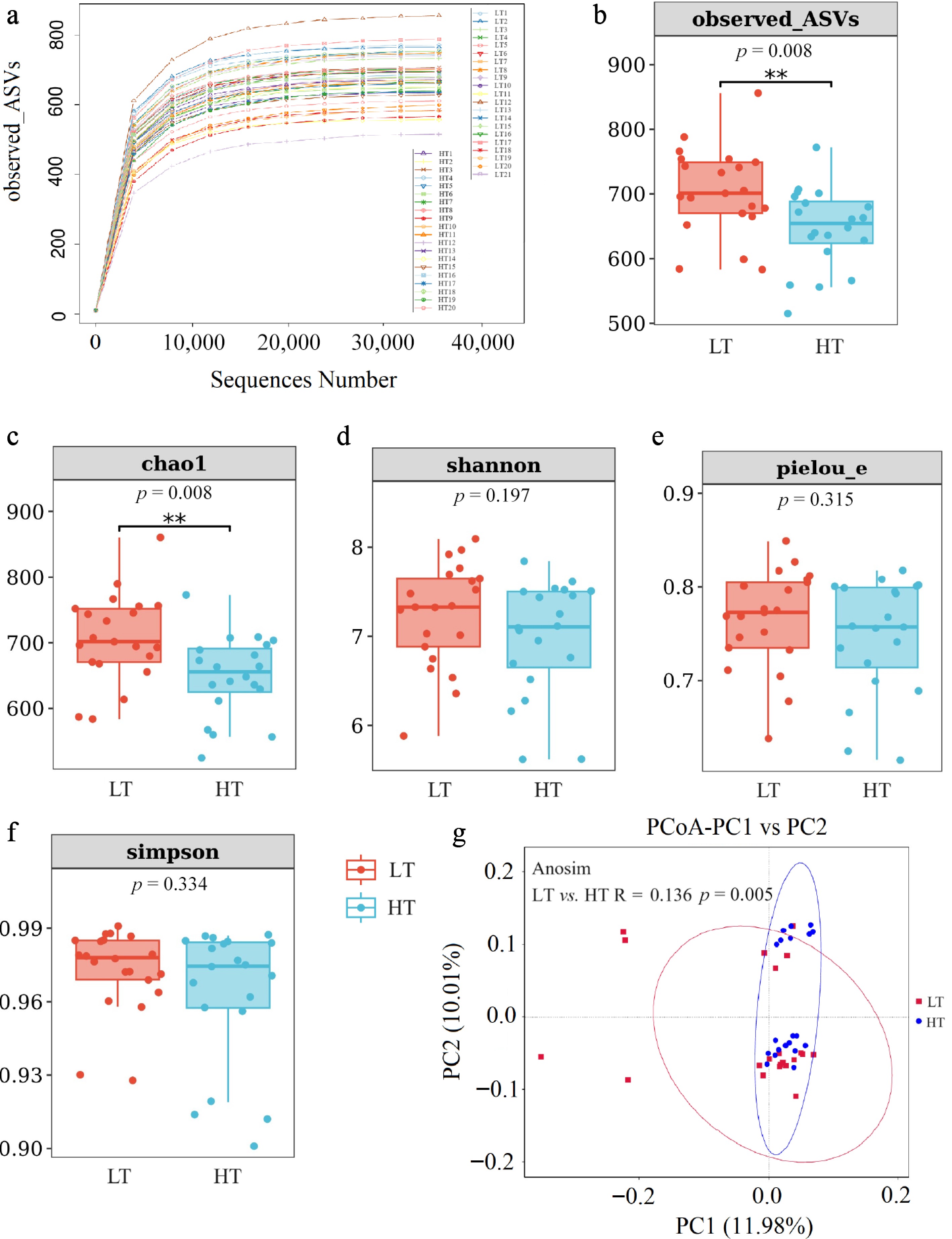
Figure 4.
Comparison of microbial diversity in feces between boars in the low testicular index (LT) (n = 21) and high testicular index (HT) (n = 20) groups. The curve shows the relationship between the total number of ASVs and the sequence number (a). Bar chart of the number of ASVs (b) and alpha diversity measured by the Chao1 (c), Shannon (d), Pielou_e (e) and Simpson (f) indices. Beta diversity, as depicted by the principal coordinate analysis (PCoA) plot based on unweighted UniFrac distance dissimilarities, revealed distinct fecal microbial compositional differences between the LT and HT groups (g). ** p < 0.01 indicates a significant difference between the LT and HT groups.
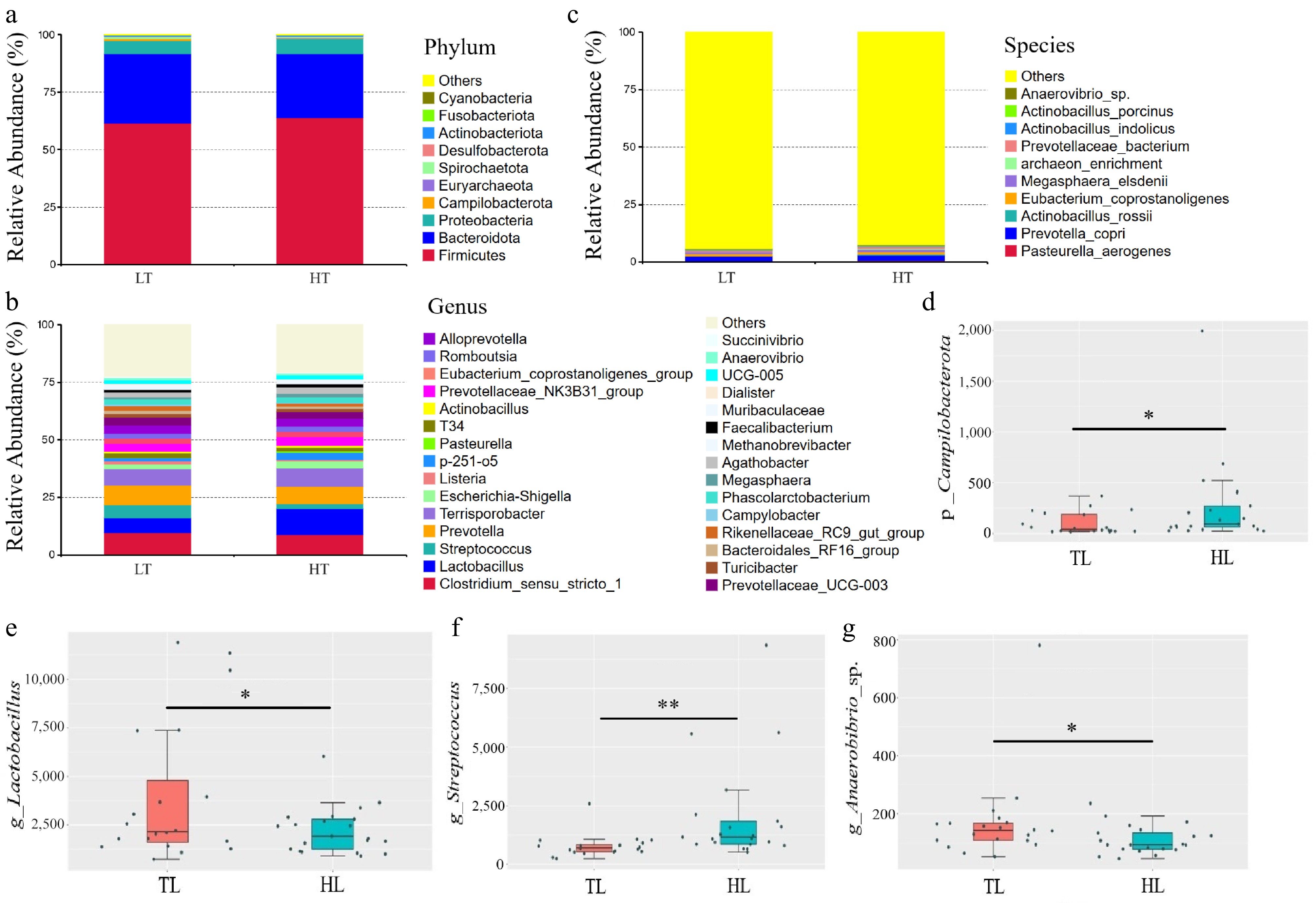
Figure 5.
The difference in the composition of the fecal microbiota between the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups was assessed. The relative abundances of major (a) phyla, (b) genera, and (c) species are shown. The absolute abundances of the top 10 significantly different taxa at the phylum (d), genus (e, f), and species (g) levels are presented. * p < 0.05 and ** p < 0.01 indicate significant differences between the LT and HT groups.
Alterations in the fecal microbiota, particularly those involving microbes with specialized functions, may signal a shift in the overall functional profiles of the fecal microbiota. To investigate this, we utilized PICRUSt software to predict the functional profiles on the basis of ASV-assigned taxa (Fig. 6). Our findings revealed that the LT group presented greater abundances of several key pathways, including K00059 (3-oxoacyl-[acyl-carrier protein] reductase, p = 0.014), K02078 (acyl carrier protein, p = 0.004), K03111 (single-strand DNA-binding protein, p = 0.027), K00790 (UDP-N-acetylglucosamine 1-carboxyvinyltransferase, p = 0.019), K03574 (8-oxo-dGTP diphosphatase, p = 0.013), and K00058 (D-3-phosphoglycerate dehydrogenase/2-oxoglutarate reductase, p = 0.018), than did the HT group. These biochemical processes are associated with fatty acid biosynthesis, energy metabolism, carbohydrate metabolism, amino acid metabolism, and DNA replication and repair. Conversely, the HT group presented significantly greater abundances of genes related to chromosomes and associated proteins, adenosine triphosphate (ATP) hydrolysis, ATP-binding cassette (ABC) transporters, peptidoglycan biosynthesis, and nucleotide metabolism. Specifically, pathways such as K03497 (chromosome partitioning protein, p = 0.034), K03569 (rod shape-determining protein MreB and related proteins, p = 0.014), K01537 (Ca2+-transporting ATPase, p = 0.042), K02035 (peptide/nickel transport system substrate-binding protein, p = 0.017), K07258 (serine-type D-Ala-D-Ala carboxypeptidase, p = 0.022), and K00760 (hypoxanthine phosphoribosyl transferase, p = 0.034) were more prevalent in the HT group than in the LT group. These processes are linked to chromosomes and their associated proteins, ATP hydrolysis, ABC transporters, peptidoglycan biosynthesis, and nucleotide metabolism.
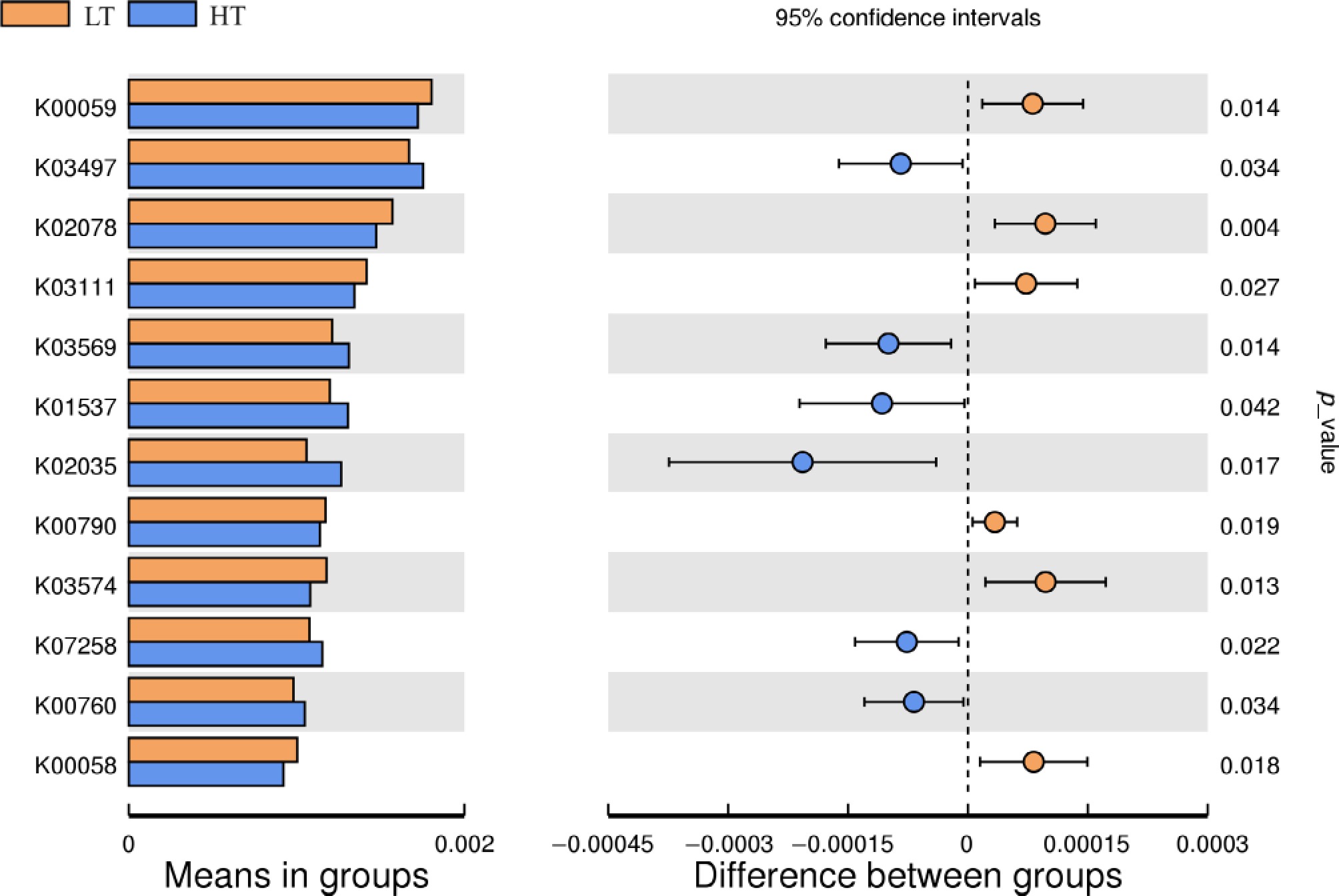
Figure 6.
Differences in predicted fecal microbial functional profiles between boars in the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups.
Relationships among the fecal microbiota composition, testicular development indices, and production parameters
-
To further elucidate the interrelationships among the fecal microbiota composition, testicular development, and production performance indices, we conducted Spearman's correlation analyses (Fig. 7). The results indicated that the TI was positively correlated with serum testosterone levels, testicular testosterone levels, total testicular volume, TW, and LMP but was negatively correlated with FMP. Additionally, LMP was negatively correlated with FMP. Testicular testosterone levels were positively correlated with total testicular volume and serum testosterone levels. The abundance of Streptococcus was negatively correlated with TI, serum testosterone levels, testicular testosterone levels, total testicular volume, and Anaerovibrio sp. abundance but positively correlated with Campilobacterota abundance. Anaerovibrio sp. abundance was positively associated with TW, TI, and Lactobacillus abundance but negatively correlated with Campilobacterota abundance. However, no statistically significant associations were found between Lactobacillus abundance and testicular development or production performance indices.
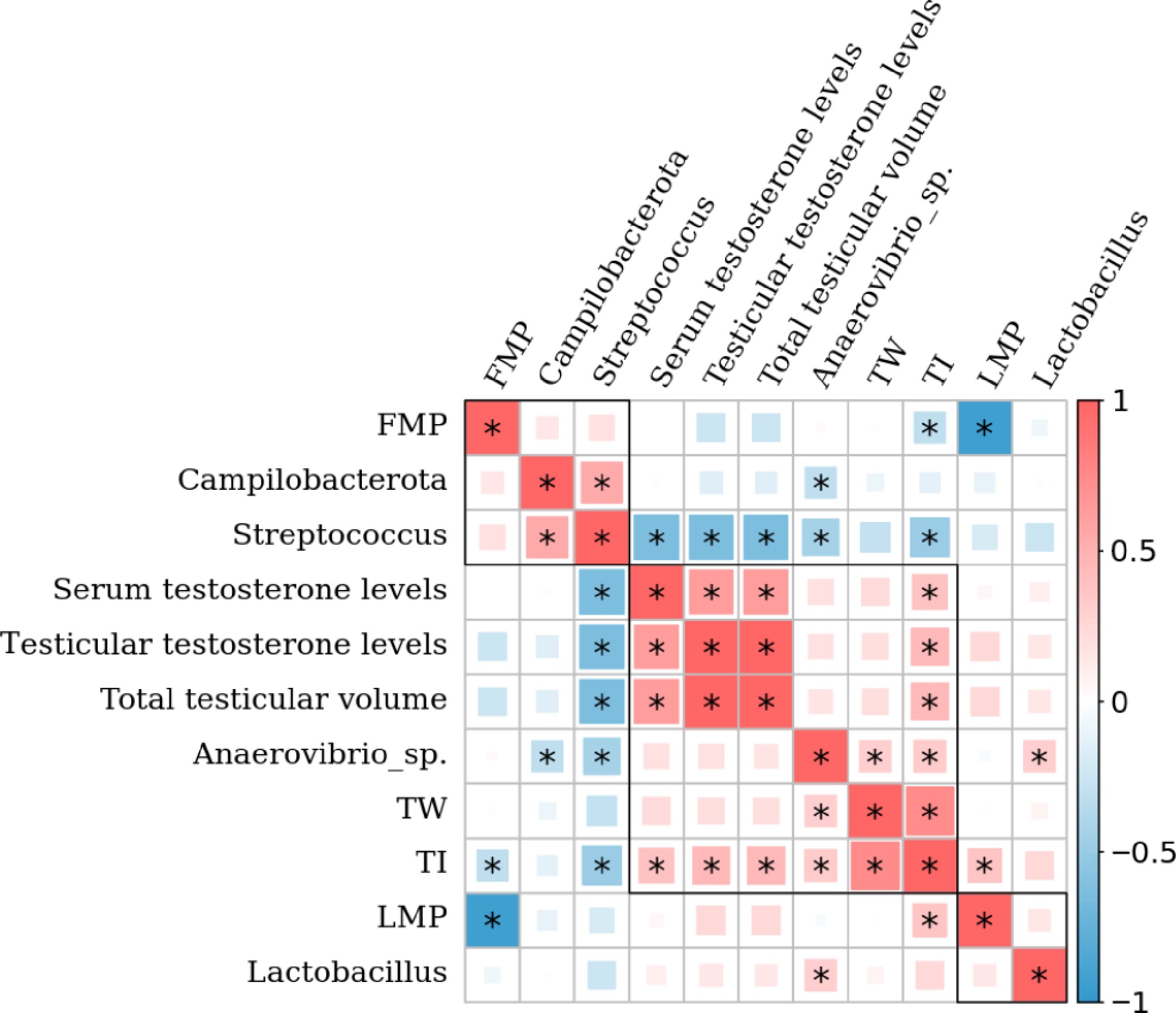
Figure 7.
Heatmap of Spearman's correlation analysis between the fecal microbial microbiota, testicular development indices, and production performance indices of boars in the low testicular index (LT, n = 21) and high testicular index (HT, n = 20) groups. Red represents a positive correlation, whereas blue represents an inverse correlation. * p < 0.05.
-
With improvements in living standards, the demand for meat products has increased significantly. Domesticated pigs are among the most important livestock breeds that provide a substantial portion of meat for human consumption. Breeding high-quality boars is crucial in pig production because it directly impacts the efficiency of the porcine industry. However, from a breeding perspective, the process is constrained by a variety of factors, with testicular development being a core element. Production performance is also a key indicator determining the overall profitability of the pig industry. Moreover, a healthy gut microbiota plays an important role in assisting the host in nutrient absorption and utilization, thereby providing the necessary nutritional support for growth. However, the specific associations among the gut microbiota, testicular development indices, and production performance have not yet been clarified. Therefore, in the present study, the testicular development, production performance, and fecal microbiota diversity of JN1WBs in the LT and HT groups were evaluated. Understanding these relationships could provide valuable insights for optimizing breeding strategies and improving the efficiency of pig production.
Chen et al. reported that backfat thickness and LMP are important economic characteristics that affect the efficiency and profitability of modern pig production[2]. A large amount of fat is regarded as 'waste fat' with little commercial value during the maturation phase of farm animals. Therefore, developing strategies to decrease subcutaneous fat deposition in pigs has been one of the breeding goals for many years. Interestingly, in the present study, we detected no distinct differences in IBW, FBW, ADG, or FCR between the boars in the HT and LT groups; however, the boars in the HT group presented lower FMP and greater LMP than did those in the LT group. These results indicated that although boars in the LT and HT groups had similar growth performances, boars in the HT group had more efficient conversion of feed to meat yield than did boars in the LT group. Pig testicular volume is highly correlated with the production of mature sperm[21]. Testosterone plays a key role in testicular development and function. It supports testicular growth and sperm production by regulating hormone levels in the hypothalamic‒pituitary‒gonadal axis. Insufficient testosterone can lead to testicular dysplasia and reproductive dysfunction[13]. In line with these findings, boars in the HT group presented significantly greater TW, testicular volume, and testosterone levels but also presented a lower FMP than those in the LT group did. These results suggest that boars in the HT group may possess greater capacity for spermatogenesis than their counterparts in the LT group. However, further experiments are needed to confirm this hypothesis.
The gut harbors a complex ecosystem with a highly variable microbiome. Alpha diversity is frequently employed to assess the diversity of the microbial community within a single sample, whereas beta diversity is typically used to reflect differences in species complexity among samples at the group level. In this study, the HT group of boars presented significantly lower ASV and Chao1 values for their fecal microbiota than did the LT group. Moreover, the unweighted UniFrac distance revealed that the composition of the fecal microbiota in the LT group was distinct from that in the HT group, indicating differences in gut development and key microbial species between the two groups. Research suggests that there is a close connection between the male gut microbiome and testosterone levels. Harada et al.'s study indicated that testosterone has the capacity to significantly reshape the gut microbiome via intricate mechanisms[22]. Building on this, in 2024, Yan and coresearchers elaborated on how sex hormone-binding globulin (SHBG) exerts a causal effect on the gut microbiome[23]. In contrast, the gut microbiota may also affect testosterone levels[24]. In recent years, an increasing number of studies have suggested a close relationship between the male gut microbiota and testosterone levels. For example, Chen et al. proposed that the gut microbiota could regulate testosterone levels by impacting the hypothalamic‒pituitary‒gonadal (HPG) axis; specifically, it is believed that the gut microbiota might alter the secretion of gonadotropin-releasing hormone (GnRH), which, in turn, could influence the release of luteinizing hormone (LH), a hormone that directly promotes the production of testosterone[25]. Furthermore, it has been discovered that gut microbes with steroid-metabolizing enzymes can modulate testosterone levels by influencing hormone metabolism[26]. The results of this study revealed that, compared with the LT group, the HT group of boars presented significantly greater levels of serum and testicular testosterone. Testosterone, the principal male sex hormone secreted by Leydig cells in the testicles, is pivotal for testicular development and maintenance, as well as for the growth of reproductive organs and sperm production[9]. Inadequate testosterone secretion may result in suboptimal testicular development and reproductive dysfunction[13]. Hence, high levels of testosterone may play a significant role in promoting the testicular development of boars in the HT group. Furthermore, the HT group presented increased abundance of Lactobacillus and Anaerovibrio sp. but decreased abundance of Streptococcus and Campilobacterota. Analysis of the fecal microbial composition of Meishan pigs revealed a significant negative correlation between testosterone levels and the relative abundance of Streptococcus[27]. In a separate study, researchers explored the impact of Lactobacillus on sperm's kinematic parameters and discovered that Lactobacillus can significantly increase testosterone levels[28]. Conversely, few studies have explored the relationship between Anaerovibrio sp. and testosterone levels. Considering the multifaceted influence of the gut microbiota on the host's health and hormone levels, Anaerovibrio sp. may influence testosterone levels indirectly by modulating the gut environment. Nonetheless, definitive evidence confirming a direct relationship between the gut microbiome and testosterone levels is still lacking, and the microbiome hypothesis has yet to be thoroughly validated. Future studies may utilize functional interventions to modulate the microbiota and elucidate its direct impact on testosterone production. Potential approaches include the administration of antibiotics to reduce the microbial load, fecal microbiota transplantation (FMT) to alter the gut microbial community, and a comprehensive analysis integrating microbial metabolites with markers of androgenic steroidogenesis. Furthermore, experiments involving testosterone injections in boars could be conducted to investigate whether testosterone directly alters the composition of the gut microbiota, thereby further uncovering the underlying mechanisms. Moreover, Lactobacillus species are associated with changes in semen parameters, whereas Streptococcus species appear to have a negative effect on sperm quality parameters, such as the sperm concentration, total and progressive motility, viability, DNA integrity, and relative mRNA expression[29−31]. Campilobacterota has been linked to various inflammatory diseases[32]. Inflammation can lead to damage to the testis and epididymis, thereby affecting the production and maturation of sperm[33]. This may explain why higher TWs, testicular volume, and testosterone levels were observed in the HT group than in the LT group. In summary, while testosterone is likely a primary factor influencing testicular development, the composition of the gut microbiota may also contribute to this process. Further studies are needed to elucidate the interactions among these factors and their cumulative impact on testicular development.
We subsequently explored the relationships among the fecal microbiota composition, testicular development, and production performance indices in boars through Spearman's correlation analysis. Streptococcus abundance was negatively correlated with TI, serum testosterone levels, testicular testosterone levels, and total testicular volume. This finding is consistent with previous research results[27,29−31], indicating that the relative abundance of Streptococcus has a negative impact on testicular development. In addition, excessive fat accumulation has been suggested to reduce the efficiency of converting feed to meat yield[34]. High LMP and low FMP are usually associated with better feed conversion efficiency. An improved feed conversion efficiency may, in turn, support the overall health and nutritional status of boars. Both Lactobacillus and Anaerovibrio sp. are significantly positively associated with feed efficiency[35]. In this study, the feces of boars in the HT group presented increased abundance of Lactobacillus and Anaerovibrio sp., increased LMP, and decreased FMP. LMP was significantly negatively correlated with FMP and significantly positively correlated with TI, whereas FMP was significantly negatively correlated with TI. However, neither LMP nor FMP was significantly correlated with testosterone levels or the gut microbiota. TI was positively correlated with testosterone levels and testicular volume, and the abundances of Anaerovibrio sp. and Anaerovibrio sp. were also positively correlated with Lactobacillus. These results suggest that LMP and FMP may indirectly affect testosterone levels, and thereby testicular development, by modulating feed conversion efficiency. However, Lactobacillus abundance was not significantly associated with testicular development or production performance indices. Early studies have shown that Lactobacillus is associated with increased sperm motility[31,36], testosterone levels[28], and improved animal performance[37,38]. The lack of a significant association with Lactobacillus in this study was somewhat surprising. Notably, Lactobacillus was positively associated with Anaerovibrio sp., whereas Anaerovibrio sp. was negatively correlated with Streptococcus. These data suggest that Lactobacillus may influence the performance and testicular development of boars by modulating the abundance of related microbes.
In addition, the gut microbiota provides nutrients to the host by metabolizing various feed components to produce end products and other byproducts (vitamins, amino acids, and indole derivatives)[39]. The gut microbiota is vital for nutrient absorption, energy metabolism, and carbohydrate metabolism, especially in the processing of indigestible polysaccharides[35,40]. Differences in the fecal microbiota's composition may result in differences in the fecal microbiota's functional profiles. Therefore, we predicted the potential functions of fecal microbes with PICRUSt software on the basis of ASV-assigned taxa. In this study, there were significantly more biochemical processes related to fatty acid biosynthesis and nutrient metabolism in the LT group than in the HT group, which may be attributed to the greater community richness (observed ASVs and Chao1) and FMP in the LT group. A previous study revealed that adipose tissue can use a high proportion of carbohydrates (glucose and acetate) for fatty acid synthesis[41]. Furthermore, increased microbial community richness requires increased nutrient metabolism rates. Therefore, there were more nutrient metabolism pathways related to fatty acid biosynthesis, energy metabolism, and carbohydrate metabolism in the LT group than in the HT group. ATP hydrolysis is the most direct energy source in the host and can provide energy for sperm generation. The separation of chromosomes during male meiosis can lead to rapid production of sperm[42]. Nucleotide metabolism is crucial for purine and pyrimidine generation and is a vital substrate for DNA or RNA synthesis. ABC transporters are important for maintaining host health and boosting immunity. Here, we found that genes belonging to the functional categories associated with chromosomes and associated proteins, ATP hydrolysis, ABC transporters, peptidoglycan biosynthesis, and nucleotide metabolism were significantly more abundant in the HT group than in the LT group. These results suggested that, compared with LT boars, HT boars may have excellent sperm production capacity, which is consistent with the greater abundance of Streptococcus and Campilobacterota being associated with poor testicular development in the LT group compared with the HT group.
-
Our findings revealed that, compared with LT boars, HT boars had a reduced capacity for fat deposition in adipose tissue, a greater lean meat yield, and more advanced testicular development. Additionally, the HT boars presented significantly greater abundance of Lactobacillus and Anaerovibrio sp., whereas the LT boars presented greater abundance of Streptococcus and Campilobacterota. Collectively, these results indicate that HT boars not only demonstrate superior production performance and testicular development but also present a greater abundance of bacteria associated with increased nutrient absorption and utilization. Conversely, they present a lower abundance of bacteria linked to inflammation and health issues that can negatively impact production performance and testicular development. These findings offer valuable insights for the preliminary selection of breeding sires on the basis of the fecal microbiota among sexually mature JN1WBs. Notably, our study involved randomly assigning 41 pigs to five pens, with each pen containing eight or nine pigs. Despite all pigs being housed under identical conditions, the influence of cohabitation on the fecal microbiota is significant, potentially leading to variations among pigs within the same pen. These differences may result from interactions involving feed troughs or feces, which can transfer microbial communities and affect both the intestinal and fecal microbiota. Additional factors, including diet and social behaviors within the pen, also significantly contributed to these variations. These factors can influence microbial colonization and dietary habits, potentially impacting the gut microbiota independently of TI. To address this variability, future studies might consider increasing the sample size or using pigs from the same litter to minimize variability. Furthermore, to minimize dietary interference with the gut microbiota composition, all pigs were fed a standardized diet throughout the experiment. However, the composition of the diet could still indirectly affect the microbiota structure. Further research could explore the interactions between different dietary components and TIs, as well as the specific impact of these interactions on the gut microbiota. It is also important to record comprehensive field information to facilitate a thorough analysis of the potential impacts on the gut microbiota.
This research was funded by the Doctoral Start-up Fund from Jilin Agricultural Science and Technology College, grant number (2022)703.
-
All procedures were reviewed and preapproved by the Animal Welfare Ethics Committee of Jilin University (identification number SY202105020; approval date: 20 May 2021). The research followed the "replacement, reduction, and refinement" principles to minimize harm to the animals. This study provides details on the housing conditions, care, and pain management of the animals, ensuring that the impact on the animals was minimized during the experiment.
-
The authors confirm their contributions to the paper as follows: conceptualization, supervision, methodology, and investigation: Zhang Y, Choi SH; resources, data curation, and writing − original draft: Zhang Y, Qu HX; review and editing of the manuscript: Liang S, Yu J, Choi SH; funding acquisition: Zhang Y. All the authors have read and agreed to the published version of the manuscript.
-
The data that support the findings of this study are available in the NCBI Sequence Read Archive (SRA) repository: www.ncbi.nlm.nih.gov/sra/PRJNA1070801.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Qu HX, Liang S, , Choi SH. 2026. Comparative analysis of testicular development, production performance, and the fecal microbiota in boars with different testicular indices. Animal Advances 3: e004 doi: 10.48130/animadv-0025-0035 

Comparative analysis of testicular development, production performance, and the fecal microbiota in boars with different testicular indices
- Received: 27 April 2025
- Revised: 28 July 2025
- Accepted: 07 August 2025
- Published online: 28 January 2026
Abstract: A healthy gut microbiota is essential for the host's absorption and utilization of nutrients, providing necessary nutritional support for growth. Conversely, an imbalance in the gut microbiota can lead to poor nutrient absorption, which may indirectly affect production performance and testicular function. In this study, we observed that 170-day-old Junmu No. 1 white boars (JN1WBs) with a high testicular index (HT) presented superior testicular development and production performance indices compared with those with a low testicular index (LT). Fecal 16S rRNA gene sequencing revealed that Lactobacillus and Anaerovibrio sp. were significantly more abundant, whereas the abundances of Campilobacterota and Streptococcus were significantly lower in HT boars than in LT boars. Spearman's correlation coefficients revealed that the abundance of Streptococcus was negatively correlated with the testicular index (TI), lean meat percentage (LMP), testicular testosterone level, total testicular volume, and Anaerovibrio sp. abundance but was positively correlated with Campilobacterota abundance. Anaerovibrio sp. abundance was positively associated with testicular weight (TW), TI, and Lactobacillus abundance but negatively correlated with Campilobacterota abundance. Moreover, prediction of fecal microbiota function revealed that compared with those in the LT group, the expression of genes associated with nutrient metabolic pathways was significantly upregulated, but the expression of genes related to chromosomes and associated proteins, adenosine triphosphate (ATP) hydrolysis, ATP synthase-binding cassette (ABC) transporters, peptidoglycan biosynthesis, and nucleotide metabolism was downregulated in the HT boars. Taken together, the results of the present study indicate a significant correlation between the gut microbiota of boars and their testicular development, as well as production performance. These findings provide important fundamental information for improving the breeding efficiency of breeding boars.
-
Key words:
- Boar /
- Fecal microbiota /
- Testicular development /
- Production performance