








-
Antibiotics play a crucial role in treating infectious diseases in humans, animals, and plants. With the misuse of antibiotics, an increasing amount of these substances are entering the environment, driving the emergence and spread of antibiotic resistance genes (ARGs)[1]. The global proliferation of ARGs has become a significant challenge in public health[2,3]. Aquatic environments serve as critical reservoirs and transmission pathways for ARGs, playing a pivotal role in their spread[4]. Over the past decade, numerous researchers have focused on the detection, screening, source identification, and risk assessment of ARGs in river ecosystems[5−7]. These macro-scale studies have significantly enhanced public awareness of AMR, and deepened the research on the environmental behavior of ARGs. However, the spread of ARGs discharged into rivers is influenced at the micro-scale by various biological mediators, such as microalgae[8], and abiotic mediators, such as microplastics[9].
Microalgae and bacteria are two dominant microbial groups in the photic zone of natural water bodies, having coexisted in aquatic ecosystems for billions of years. Currently, research on microalgae-bacteria symbiotic systems primarily focus on two aspects. One involves the microalgae-bacteria granular sludge (MBGS) for the removal of conventional and emerging contaminants, while the propagation and removal efficiency of ARGs within these systems are also investigated. However, conclusions from these studies and review articles regarding the impact of microalgae on ARGs in MBGS systems are inconsistent. The mechanisms by which microalgae inhibit the propagation of ARGs in MBGS systems generally involves roles as follows: (1) the removal of antibiotics, thereby reducing the selective pressure on ARGs[10]; (2) the reduction in the abundance of potential hosts[11]; and (3) the adsorption and encapsulation of microalgae on the surface of granular sludge, acting as a barrier to hinder horizontal gene transfer (HGT) between bacteria[12,13]. However, other studies have confirmed that microalgae can increase the abundance of mobile genetic elements (MGEs) and potential hosts, thereby promoting the propagation of ARGs[14]. As such, a consistent conclusion regarding the effect of microalgae on ARGs in MBGS systems has not yet been established in the existing research. The other main area focuses on microalgae and bacteria outside of granular sludge, which more closely mirrors the relationship between microalgae and bacteria in actual watersheds. Microalgae can act as carriers for bacteria and interact with them over certain distances, forming a unique microecological environment known as the phycosphere[15]. The phycosphere creates favorable conditions for the propagation of ARGs, making them a potential environmental hotspot[8,16,17]. As shown in Fig. 1, the two research approaches involve radically different spatial structures of microalgae-bacteria symbiosis, which results in various pathways and potential mechanisms for ARG spread.
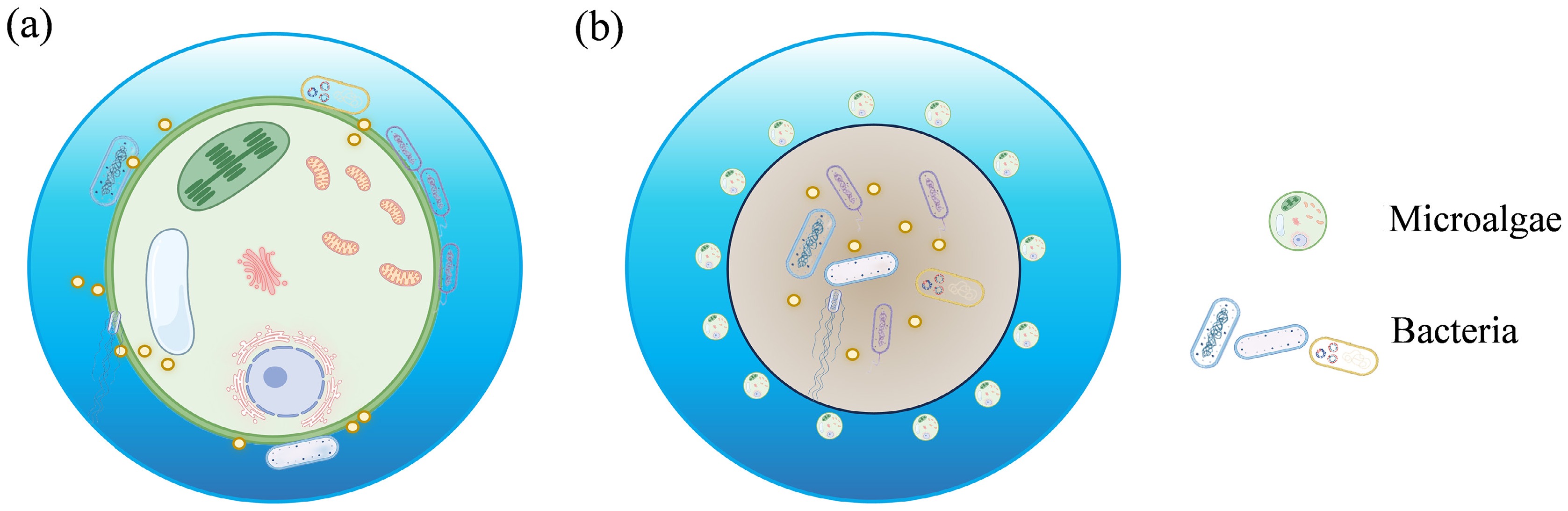
Figure 1.
Schematic diagram of the spatial structure of the (a) phycosphere, and (b) MBGS.
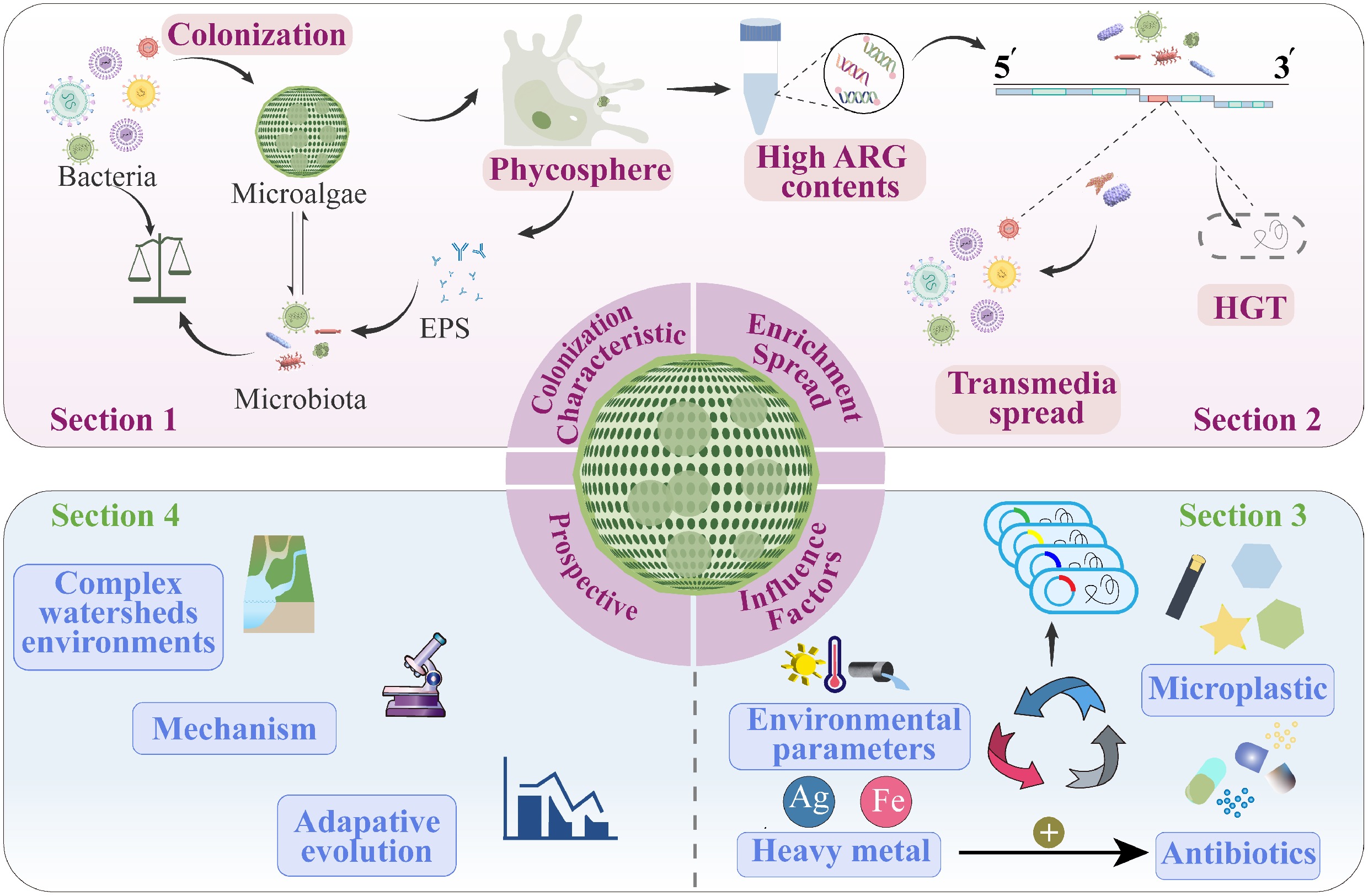
The phycosphere has confirmed the significant potential of microalgae to harbor and disseminate ARGs, positioning them as critical nodes linking different domains of the One Health framework. Research on AMR in the phycosphere remains in its infancy, and the role of microalgae in the dissemination of ARGs within watersheds has not yet been systematically reviewed, representing a significant knowledge gap. This review focuses on the phycosphere within watersheds, highlighting its unique characteristics, the enrichment and spread of ARGs, and the influence of contaminants. This shows that microalgae can exacerbate the proliferation of ARGs in watersheds, posing a significant threat to ecosystems and humans. Therefore, in the final section of this review, prospects are proposed for the mechanism and adaptive evolution of ARG spread induced by microalgae. The results shed light on the role of microalgae in driving the proliferation of ARGs, which has crucial scientific significance for preventing and controlling the global spread of AMR.
-
The Web of Science (
http://webofscience.clarivate.cn/ ) and Science Direct (www.sciencedirect.com ) databases were used to retrieve publications that evaluated the impacts of microalgae on ARGs in watersheds. The topics used were [microalgae AND antibiotic resistance genes; algae AND antibiotic resistance genes]. This study searched the published literature database as of September 18, 2025, and reviewed all publications individually to exclude duplicate and irrelevant literature. These studies focused on natural water bodies and pilot-scale experiments. However, the MBGS were not included in this study. The research information on microalgae and ARGs in the phycosphere are summarized in Table 1. Field studies primarily target algae such as Microcystis or green tide during bloom events, whereas Chlorella has mainly been investigated in laboratory studies. The characteristics of microalgae and the features of the phycosphere microenvironment are shown in Fig. 2.Table 1. The summary of the microalgae and ARGs related to the phycosphere in the previous studies
Order Microalgae species ARGs types Exogenous pollutants Sampling point/bacterial source Article type Topic Ref. 1 Cyanobacteria intI1, 16S rRNA, sulfonamide resistance genes (sul1, sul2, sul3, sulA), tetracycline resistance genes (tetA, tetB, tetD, tetE, tetJ, tetK, tetM, tetO, tetQ, tetS, tetW), macrolide resistance genes (ermB, ermC, ereA, ereB, mphA, mphB), quinolone resistance genes (qnrB, qnrS, aac(6′)-Ib), β-lactam resistance genes (blaTEM, blaOXA, blaOXY, blaSHV, blaCTX-M), and aminoglycoside resistance genes (strA, strB) Cyanobacteria bloom Taihu Lake, China Sampling from
surface waterAntimicrobial resistance in the phycopshere [52] 2 / Sulfonamide resistance genes (sul1, sul2, sul3), tetracycline resistance genes (tetA, tetB, tetC, tetG, tetM, tetO, tetQ, tetW, tetZ), quinolone resistance genes (gyrA, qnrA, qnrB, qnrS), macrolide resistance genes (ermB, ermC, ermF), chloramphenicol resistance genes (cat1, floR, cmlA), β-lactams resistance genes (blaNDM1, blaIMP4, blaCTX-M, blaSHV), integron integrase genes (intI1, intI2), transposases (tnpA-01, tnpA-02, tnpA-03, tnpA-04, tnpA-05, tnpA-07, IS613), 16S r RNA River contamination Ba River, China Sampling from
surface waterAntimicrobial resistance in the phycopshere [51] 3 Cyanobacteria 296 primer pairs that targeting one 16S gene, 285 major ARGs and 10 MGEs (1 clinical class 1 integron, 1 class 1 integron and eight transposases) Cyanobacterial bloom Taihu Lake, China Sampling from
surface waterAntimicrobial resistance in the phycopshere [36] 4 Green tide Metagenomic sequencing Green tide bloom Yellow Sea, China Sampling from
surface waterAntimicrobial resistance in the phycopshere [37] 5 / / / Lake Mochou,
ChinaSampling from
surface waterBacterial colonization and assembly in microalgae [63] 6 / / / North Sea island Helgoland Sampling from
surface waterBacterial colonization and assembly in microalgae [23] 7 Ulva sp., Saccharina sp., Grateloupia sp., and Gelidium sp. / / Weihai, China Sampling from
surface waterBacterial colonization and assembly in microalgae [54] 8 Chlorella pyrenoidosa Two sulfonamide resistance genes (sul1 and sul2), 15 tetracycline resistance genes (tet(36)-01, tet(36)-02, tetA-02, tetG-01, tetG-02, tetL-02, tetM-01, tetM-02, tetQ, tetR-01, tetR-02, tetS, tetT, tetW-01 and tetX), and five MEGs (intI1, tnpA-03, tnpA-04, tnpA-05, tnpA-07) Silver nanoparticles (AgNPs)
Hematite nanoparticles (HemNPs)
Tetracycline
SulfadiazineChicken farm wastewater Laboratory experiment Antimicrobial resistance in the phycopshere [75] 9 Chlorella pyrenoidosa Sulfonamide resistance genes (sul2, sul1, dfrA1), tetracycline resistance genes (tetX, tetG-02, tetG-01, tetM-01, tetM-02, tetA-01, tet(36)-02, tetL-02, tetA-02), aminoglycoside resistance gene (aadA2–03, aadA1, aadA2–01, aadA2–02, aadA-02, aadA-01, strB, aadA5–01, aac(6′)-Ib-3, aadA5–02, aac(6')-Ib-1, aac(6')-Ib-2, aadE, aac (6')-II, aphA1, aadA9–02), MGEs (tnpA-04, intI1, tnpA-03, tnpA-07) Sulfadiazine Soil leachate near poultry farms Laboratory experiment Antimicrobial resistance in the phycopshere [70] 10 Chlorella vulgaris sul1, sul2, intI1 Sulfadiazine Wastewater from fish ponds Laboratory experiment Antimicrobial resistance in the phycopshere [71] 11 Chlorella pyrenoidosa Sulfonamides resistance genes (sul1 and sul2), phenicol resistance genes (floR, fexA, fexB, optrA, cfr, and pexA), integrase and transposase (intI1, IS613, tnpA01, tnpA02, and tnpA03) Polylactic acid MP
FlorfenicolDonghu lake and fish ponds Laboratory experiment Antimicrobial resistance in the phycopshere [72] 12 Auxenochlorella pyrenoidosa Metagenomic sequencing Contents of N and P Donghu lake Laboratory experiment Antimicrobial resistance in the phycopshere [61] 13 Chlorella vulgaris tetQ, tetE, sul1, sul2, strA, strB, blaOXA, blaPSE / Effluent from the sewage treatment plant Laboratory experiment Antimicrobial resistance in the phycopshere [10] 14 Chlorella vulgaris sul1, sul2, sul3, intI1, 16S rRNA gene Sulfamethoxazole Jinggong Lake Laboratory experiment Antimicrobial resistance in the phycopshere [16] 15 Chlorella vulgaris, Scenedesmus acuminatus, Chlamydomonasnoctigama, Chlorolobion braunii, Raphidocelis subcapitata Tetracycline resistance genes (tetA, tetB, tetC, tetZ), quinolone resistance genes (gyrA, qnrA, qnrB, qnrS), macrolide resistance genes (ermB, ermC, ermF), chloramphenicol resistance genes (catA1, floR, cmlA), aminoglycoside resistance genes (aadA, aacC, aph(2’)-Id), and β-lactam resistance genes (blaCTX-M, blaSHV, blaTEM), intI1, intI2, 16S rRNA gene Rivers with varying
degrees of pollutionQishui River Laboratory experiment Antimicrobial resistance in the phycopshere [17] 16 Chlorella vulgaris 16S rRNA, tetracycline resistance genes (tetA, tetB, tetC, tetG, tetM, tetO, tetQ, tetW, tetZ), quinolone resistance genes (aaC(6')-Ib, gyrA, oqxAB, qepA, qnrA, qnrB, qnrC, qnrD, qnrS), aminoglycoside resistance genes (aph(2')-Id), β-lactam resistance genes (blaSHV), macrolide resistance genes (ermC), chloramphenicol resistance genes (floR), sulfonamide resistance genes (sul2), mobile genetic elements (intI1, intI2, IS613, tnpA-02, tnpA-03) Tetracycline
CiprofloxacinWei River Laboratory experiment Antimicrobial resistance in the phycopshere [50] 17 Chlorella pyrenoidosa pEASY-T1 plasmid Contents of N / Laboratory experiment Antimicrobial resistance in the phycopshere [35] 18 Cyanobacteria Metagenomic sequencing Temperature and phosphorus concentration The water and sediments of the Three Gorges Reservoir Laboratory experiment Bacterial colonization and assembly in microalgae [84] 19 / / / / Laboratory experiment Bacterial colonization and assembly in microalgae [15] 20 Diatom / / / Laboratory experiment Bacterial colonization and assembly in microalgae [21] 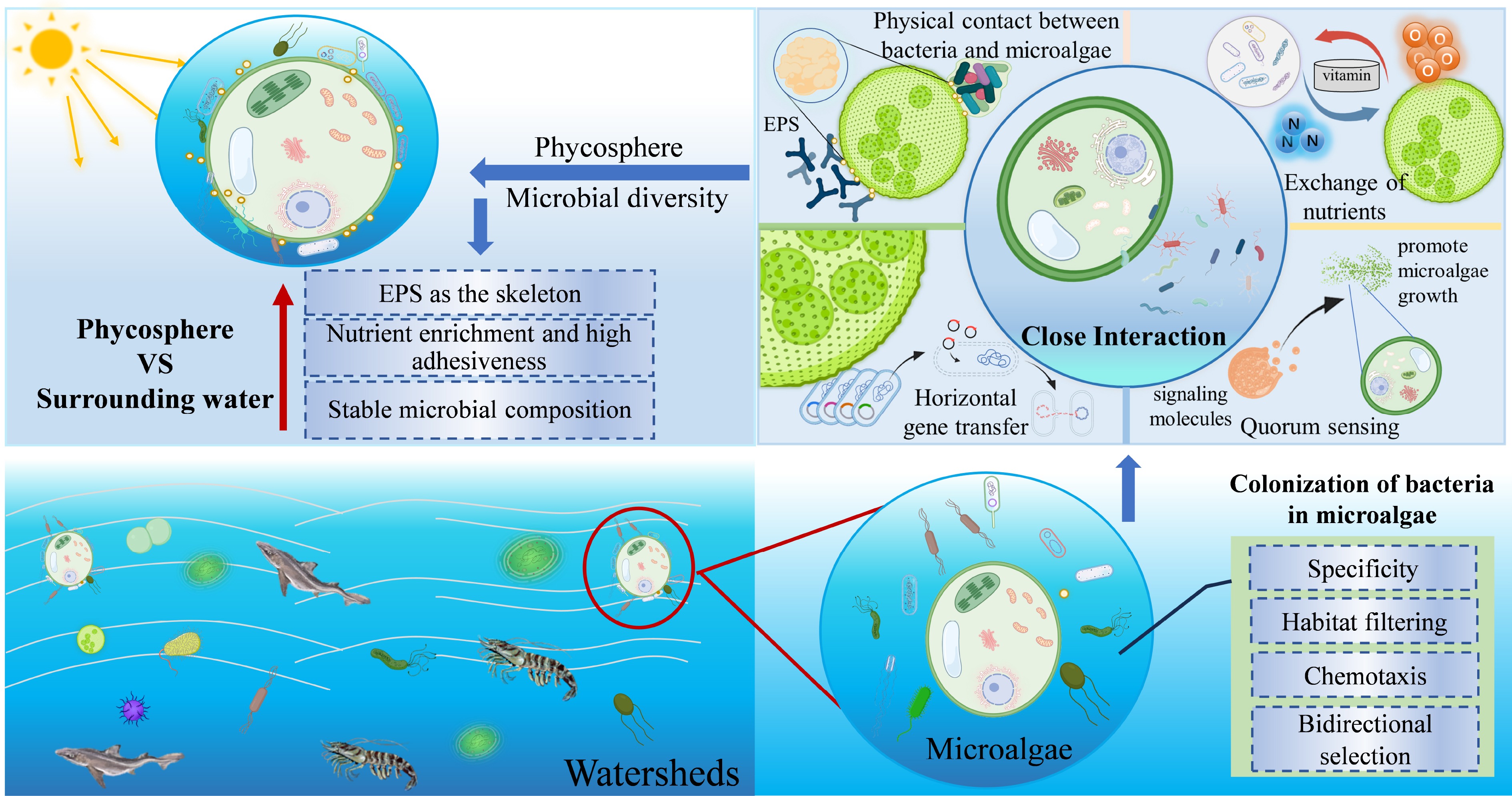
Figure 2.
Bacterial colonization and characteristics of the phycosphere environment.
Bacterial colonization on microalgae
-
In aquatic ecosystems, microalgae, with their substantial biomass and wide distribution, serve as the primary producers, and account for over 40% of global organic carbon fixation. Microalgae and bacteria have long coexisted in these ecosystems, where bacteria can colonize microalgal surfaces through random encounters, chemotaxis, and vertical transmission[8]. Furthermore, the reciprocal release of extracellular polymeric substances (EPS), charge disparities, and mutualistic interactions via carbon and oxygen exchange facilitate substrate interactions, and create a specific microenvironment. This in turn attracts bacterial colonization of microalgal surfaces, and promotes the formation of the phycosphere microbiota[18,19]. Both microalgae and bacteria can secrete secondary metabolites that influence the biological responses within the microalgae-bacteria system. For instance, bacteria associated with macroalgae can release quorum-sensing signal molecules that alter algal physiology and metabolism, thereby enhancing bacterial adhesion to microalgal surfaces[20]. These intricate and tight interactions make the phycosphere microbiota regarded as a 'second genome' of the microalgal host[21].
The colonization of algae by bacteria is a dynamic and challenging research topic. However, recent studies have demonstrated a degree of universality in the outcomes of bacterial colonization on microalgae. The shift from the lottery hypothesis to the habitat filtering hypothesis underscores that microalgae, such as diatoms, may drive the initial screening or selection process, thereby facilitating the subsequent enrichment of specific microorganisms[21]. The species-specific interactions between individual diatom and bacterial taxa have been extensively explored with the development of omics technology and electron microscopy[22]. Certain bacteria, such as Flavobacteriia, Gammaproteobacteria, and Roseobacter were the dominant bacterial groups during diatom blooms or in diatom-dominated phytoplankton communities due to their preference for specific polysaccharide exudates from diatoms[23,24]. Sugars and amino acids released by microalgae serve as key chemoattractants, enabling the enrichment of flagellated and chemotactic bacterial populations within the phycosphere[21]. Marine macroalgae harbor epiphytic bacterial biofilms that were species-specific and temporally adaptive[25]. The diffusion boundary layer around large seaweeds attracted unique bacterial communities to colonize on the surface of the seaweed through released substances[26], which was likely to dominate the deterministic process of phycosphere microbial assembly through competition, reciprocity, and trade-offs[8]. These findings collectively illustrate that the three-step sequential process of bacterial colonization on microalgal surfaces (i.e., contact, transient attachment, and irreversible adhesion) represents a specific outcome of bidirectional selection between microalgae and bacteria.
Characteristics of the phycosphere environment
Nutrient enrichment and high adhesiveness
-
Although the phycosphere exhibits significant divergence across algal groups, such as dinoflagellates, green algae, and diatoms[27], it is unified by a rigid structural framework primarily formed by EPS. This framework dictates the key characteristics of the phycosphere: EPS-mediated nutrient enrichment and high adhesiveness. EPS extended outward from the microalgal cells, creating a spatially structured gradient of tightly bound, loosely bound, and dissolved EPS[27]. Xiao & Zheng reviewed the composition and function of EPS secreted by microalgae[28]. The composition of these microalgal EPS primarily comprises polysaccharides, proteins, nucleic acids, and lipids, providing diverse nutritional substrates for the metabolic processes of microalgae and bacteria. In addition, within EPS, proteins and polysaccharides stabilize the structure via non-covalent interactions (hydrogen bonding, hydrophobic interactions), and covalent reactions (Maillard reaction), while lipids link other molecules via hydrophobic interactions, thereby enhancing the structural integrity of the network[29,30]. At the nanoscale, the three-dimensional polymer network is anchored by specific protein cross-linking nodes and ordered polysaccharide backbones. For example, microalgal EPS proteins (e.g., tryptophan-rich fractions) form β-sheet-rich domains or α-helical segments that act as 'bridges' to cross-link polysaccharide chains, while bacterial biofilm studies reveal conserved assembly mechanisms, such as donor-strand exchange in TasA fibers and cross-β stacking in curli amyloid fibers, which are analogous to microalgal EPS protein assembly. Molecular docking and density functional theory (DFT) calculations further demonstrate that polysaccharide hydroxyl groups (-OH) form hydrogen bonds with protein amide groups (-NH-) or aromatic rings (e.g., tryptophan benzene rings), while hydrophobic interactions between lipid chains and protein nonpolar residues consolidate the network[31,32].
Of particular significance is the stable matrix structure of EPS, which forms a three-dimensional polymer network that promotes cell-to-cell interactions and mediates surface adhesion. EPS forms a hierarchical assembly with proteins as the core and polysaccharides distributed in the middle-periphery, protein α-helices (rich in polar groups) have stronger hydrogen bonding with polysaccharides than β-sheets[29]. Consistent with this, Wang & Tao[32] observed that the ratio of α-helix/(β-sheet + random coil) in microalgal EPS proteins decrease from 72.4% to 61.6% during cell proliferation, loosening the spatial conformation to expose more polysaccharide binding sites and facilitate π-π stacking between tryptophan benzene rings, and aromatic moieties. This structural dynamism is critical for network flexibility and cross-linking efficiency[32]. Exopolysaccharides are involved in the adherence to abiotic and biotic surfaces, e.g., phycosphere-bacteria[27]. Humic acid within EPS can influence its adsorption properties[33]. Carboxyl groups in humic acid induce electrostatic repulsion between biopolymers, this repulsion can be mitigated by metal ions, enabling humic acid to form hydrophobic pockets with lipids and regulate adsorption[29,34]. The higher PS content in the phycosphere may contribute to adhesion and cell aggregation due to polyanionic and polycationic compounds, thereby mediating the deterministic processes of bacterial assembly[28].
Stable microbial composition
-
The biofilm-like structure of the phycosphere, analogous to that of other microbial aggregates, confers greater robustness to its microbiota. Attached microorganisms benefit from a relatively stable environment provided by their biofilm structures and the attachment surface[35]. During outbreaks of Enteromorpha and cyanobacteria, the phycosphere environment was also observed to be more stable than the surrounding water. This enhanced stability helps protect bacteria from external environmental disturbances[36,37]. Mutualistic interactions between microalgae and bacteria within symbiotic systems can promote enhanced polysaccharide secretion, helping to stabilize mutualism and improve adaptability to environmental fluctuations[28]. Polysaccharides (such as alginate, hyaluronic acid, and sulfated polysaccharides) form the skeleton of EPS, providing adhesion and a physical barrier to the mucus layer, which is essential for algae to resist environmental stress and further confirms the stable microenvironments in the phycosphere[38]. In previous study, it was also indicated that microalgae can enhance bacterial adaptive capacity to antibiotic stress, thereby contributing to the maintenance of the structural integrity and composition of the phycosphere microbiota[16].
Close interaction between microalgae and bacteria
-
The interactions between microalgae and bacteria have been extensively studied and reviewed. Overall, beyond serving as carriers for bacteria, microalgae engage in diverse interactions with them. These microalgae-bacteria interactions primarily include physical contact at the cellular level, and substrate exchange, signal transduction, and HGT at the molecular level[39]. Through EPS release, charge disparities, and mutualistic exchanges of carbon and oxygen, microalgae and bacteria facilitate substrate interactions, create a specific microenvironment, thereby attracting bacterial colonization on microalgal surfaces, promoting the formation of the phycosphere microbiota[18,19], and mediating their initial physical contact. The exchange of nutrients between microalgae and microorganisms is a fundamental relationship, essential not only for enhanced growth but also for their survival in polluted environments. During photosynthesis, algae provide oxygen to bacteria, while bacteria produce CO2, inorganic nitrogen, and inorganic phosphate that support algal growth[40]. The exchange of nutritional metabolites is considered the foundation of mutualistic interactions between algae and bacteria, providing both partners with enhanced growth and greater resilience to environmental fluctuations.
Signal transduction via signaling molecules is another form of interaction between bacteria and algae. Bacterial and algal signaling molecules regulate each other's behaviors by activating or inhibiting gene expression and/or physiological activities[25]. For instance, bacteria associated with macroalgae can release acyl-homoserine lactones (AHLs), quorum-sensing signal molecules that alter algal physiology and metabolism[20]. Bacteria-produced vitamins B12, B1, and B7 can promote diatom biomass growth[41]. Certain bacterial taxa (such as Vibrio) regulate photoactivity through quorum sensing, providing diatom cells with stable soluble iron in exchange for organic compounds released by diatoms[42]. Interkingdom signaling between microalgae and bacteria has been demonstrated over the past decade and has only recently gained broad scientific attention. As reviewed by Zhou et al.[43], the signal molecules/mechanisms that mediate communication between microalgae and bacteria can be divided into three categories: (1) lipid molecules, which can freely cross the cell membranes in an energy-independent manner; (2) some structurally conserved molecules have three-dimensional structures related to the functional domains of regulatory proteins; and (3) bacterial signal molecules (such as AHLs) and microalgal allelochemicals (such as flavonoids and ectocarpene).
However, due to the substantial evolutionary divergence between bacteria and microalgae, the frequency of HGT between them is extremely low, exerting minimal impact on the structure of the phycosphere microbiota[44]. Although studies have found that HGT can occur between microalgae and bacteria; there is currently no report of the HGT of ARGs between them. Microbial communities primarily determine the fate of ARGs. Consequently, microalgae-bacteria interactions (physical contact, substrate exchange, and signal transduction) influence the diversity of the phycosphere microbiota, alter the phycosphere environment, and ultimately mediate the formation and dissemination of ARGs among phycosphere bacteria.
-
The relationship between microalgae and bacteria has long been a subject of interest and research. In recent years, the transmission characteristics and impacts of ARGs at the microscale within the phycosphere has garnered increasing attention from researchers. Studies have confirmed the significant potential of microalgae to carry and disseminate ARGs, positioning them as critical nodes linking different domains under the 'One Health' framework. This poses a serious threat to the integrated health of humans, animals, plants, and the environment[45−47], while also offering new perspectives on the transmission, control, and risk assessment of ARGs in watersheds. Therefore, investigating the cross-media occurrence and dissemination patterns of ARGs in aquatic systems involving microalgae is of great scientific significance for curbing the global spread of AMR (Fig. 3).
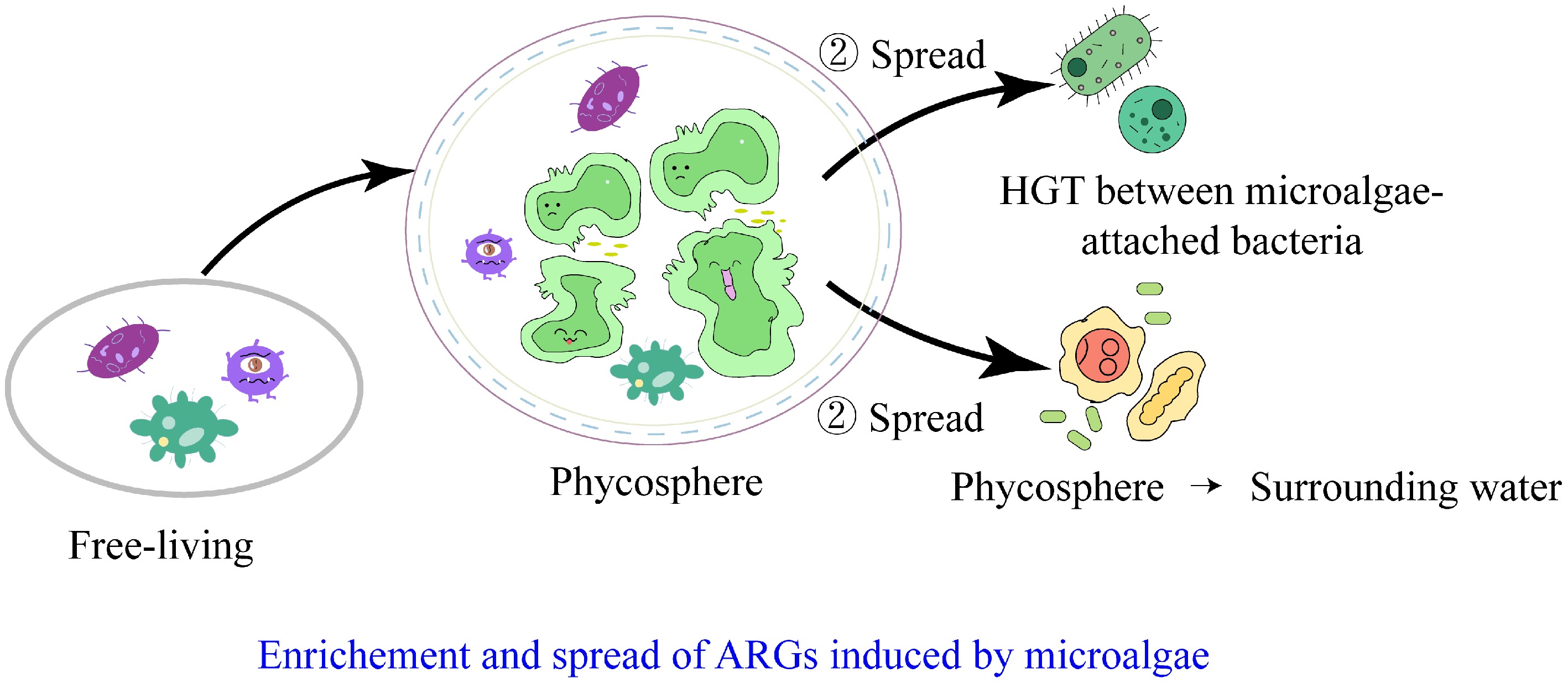
Figure 3.
Pathways of ARGs enrichment and spread induced by microalgae in watersheds.
Enrichment of ARGs in the phycosphere
-
Microalgae enrich and transfer ARGs, making the phycosphere a significant reservoir for these genes. Gao et al.[48] collected 123 water and sediment samples from 35 sites across the Pearl River Basin, and Zhang et al.[49] obtained 76 samples from 18 urban rivers in eastern China. Their combined results identified over 200 ARG subtypes in total (95% confidence interval [CI]: 185–215), with a detection rate of ≥ 89% across all samples. Xue et al.[50] focused on Chlorella vulgaris, a common freshwater microalga, and quantified 26 ARG subtypes using qPCR analysis. Among them, the abundance of ARGs in the phycosphere exceeded that of the surrounding water by up to 47 times (95% CI: 38.2–55.8; p < 0.001), highlighting the phycosphere as a significant reservoir of ARGs. A total of 30 samples, from five sites along the Ba River (China) over two seasons, were collected to quantify sulfonamide, tetracycline, and quinolone ARGs (amplification efficiency: 90%–110%; coefficient of determination, R2 ≥ 0.99). In different freshwater ecosystems (e.g., urban rivers, reservoirs, lakes), the total absolute abundance of ARGs enriched in microalgal niches (phycosphere and attached biofilms) was 1–2 orders of magnitude higher than in the surrounding water, when normalized to 16S rRNA gene abundance, the relative ARG abundance in microalgal niches was 2.67–3.54 times greater than in bulk water (95% CI: 2.13–4.01; p < 0.01), demonstrating that microalgal niches serve as larger ARG reservoirs in freshwater systems. This significant enrichment is likely driven by the combined effects of nutrient concentration, high cell density, which promotes HGT, and the protective nature of the EPS matrix, which collectively create a favorable microhabitat for ARG hosts[51].
Furthermore, microalgae facilitate the proliferation and dissemination of ARGs. Zhang et al.[36] investigated cyanobacterial blooms in Lake Taihu (China) by collecting 16 water samples (covering bloom and non-bloom periods), and performing metagenomic analysis. They found that the total relative abundance of ARGs increased from 0.32 ± 0.08 copies per 16S rRNA copy (non-bloom period) to 1.87 ± 0.23 copies per 16S rRNA copy (bloom period; 95% CI: 1.52–2.22; p < 0.001). A trend attributed to the bloom, which provides a nutrient-rich microenvironment for bacterial colonization and growth. In addition, Wang et al.[52] analyzed 12 samples from Lake Taihu and identified tetracycline resistance genes (tetA/C/Z) and sulfonamide resistance genes (sul1/2) as the dominant ARG types during blooms, accounting for 62% ± 7.3% of total ARGs (95% CI: 58.1%–65.9%). Zhao et al.[37] studied Ulva prolifera (a marine macroalga) during green tide outbreaks in the South Yellow Sea, collecting samples from the phycosphere of U. prolifera and bulk seawater at nine bloom sites. They performed high-throughput sequencing using the Illumina NovaSeq 6000 platform, and standardized ARG abundance to 16S rRNA gene copies. The results showed that ARG abundance in the U. prolifera phycosphere (3.25 × 10−4–8.26 × 10−4 copies/cell) was significantly higher than in bulk seawater (2.30 × 10−4–4.02 × 10−4 copies/cell; p < 0.05). Thus, microalgae also facilitate the proliferation and dissemination of ARGs, particularly during algal bloom events. Ji et al.[53] conducted laboratory experiments using Microcystis (a bloom-forming cyanobacterium). They set up six treatment groups (three replicates each), and exposed the system to microcystin-leucine-arginine (MC-LR, 10 μg/L), a toxin produced by Microcystis, then quantified horizontal gene transfer (HGT) of ARGs via qPCR targeting mobile genetic elements (MGEs: intI1 and tnpA). The results showed that MC-LR exposure increased the conjugative transfer frequency of ARGs by 25-fold compared to the control group (95% CI: 21.3–28.7; p < 0.001). During Microcystis blooms, the production of microcystins inhibits protein phosphatases, or increases reactive oxygen species (ROS), inducing oxidative stress and enhancing membrane permeability, leading to DNA damage. This, in turn, increased the selective pressure for ARGs and opportunities for HGT. Additionally, Lu et al.[54] examined the genomic characteristics of epiphytic bacteria in the phycosphere, which may support the storage of ARGs. They isolated 5,527 bacterial strains (4,426 from macroalgal phycospheres), and reconstructed 1,619 metagenome-assembled genomes (MAGs) from coastal reef ecosystems in Weihai (China). Genome sizes were calculated using QUAST v5.0.2, revealing that phycosphere bacteria had a larger average genome size (4.2 ± 0.31 Mb; 95% CI: 3.89–4.51 Mb) than bacteria from sediments (3.5 ± 0.27 Mb; 95% CI: 3.23–3.77 Mb), or seawater (3.3 ± 0.22 Mb; 95% CI: 3.08–3.52 Mb; p < 0.01). The higher bacterial abundance and diverse community structure in the phycosphere may increase the number of potential ARG hosts, and the elevated ARG abundance creates favorable conditions for the dissemination of antibiotic resistance genes. Therefore, the phycosphere is a critical niche for the proliferation and transmission of ARGs.
Spread of ARGs induced by microalgae
-
The dissemination of ARGs in the phycosphere occurs in two directions: first, the horizontal transfer of ARGs among epiphytic bacteria within the phycosphere, and second, the transmission of phycosphere ARGs to the surrounding water. HGT plays a critical role in the development of microbial antibiotic resistance in the phycosphere. Microorganisms in the phycosphere exchange genetic material via MGEs, such as plasmids and transposons, facilitate the spread of resistance genes among different microbial species. This HGT not only accelerates the dissemination of resistance but may also lead to the emergence of new resistant strains. Conjugation is likely the primary mechanism of HGT in microalgae-bacteria systems, with extracellular polysaccharides and ABC transporters potentially involved in the dissemination of ARGs[55]. Conjugation is recognized as the dominant pathway of ARG transmission among environmental bacteria as well[56]. The EPS matrix in the phycosphere may actively facilitate conjugative transfer of ARGs by enhancing cell-to-cell adhesion and stabilizing plasmid DNA in the extracellular environment. Additionally, the presence of divalent cations (e.g., Ca2+ and Mg2+) within the EPS can bridge negatively charged bacterial membranes, thereby promoting the formation of conjugation pilus and genetic exchange. Therefore, it remains unclear whether the horizontal transfer of ARGs among epiphytic bacteria in the phycosphere differs from that among isolated bacteria. While the mechanisms of ARG transfer among phycosphere epiphytic bacteria are the same, the frequency is hypothesized to be higher than in the surrounding water, based on the reasonable assumption of high bacterial density and ARG abundance.
The dissemination of ARGs between the phycosphere and the surrounding water is a bidirectional dynamic process. Throughout algal growth, the fate of ARGs carried by bacteria gradually shifted from a 'two-way exchange' between the phycosphere and the surrounding medium to a 'one-way transfer' from the phycosphere to the ambient water. In the context of the study on phycosphere microbes and ARGs, 'two-way exchange' refers to the back-and-forth movement of ARG-carrying bacteria or ARGs between the phycosphere (the microenvironment around algal cells), and the surrounding water during algae’s early growth stages, where those from the water, enter the phycosphere, and those in the phycosphere spread to the water. 'One-way transfer' describes the shift that occurs as algae mature, such as when they reach the stationary phase: the phycosphere becomes a key source of ARGs, sending its resistant bacteria and ARGs outward to the surrounding water with little to no reverse movement of ARGs or resistant microbes from the water into the phycosphere. This key shift is likely driven by physiological changes in algae during their later stable or aging stages; as algal cell vitality deteriorates and the phycosphere microenvironment disintegrates, ARG-rich bacteria and free DNA are released in large quantities into the surrounding water, thereby transforming the phycosphere from a relatively self-contained mutualistic niche into a significant source and active disseminator of ARGs in the broader aquatic environment. This transformation also highlights the dynamic role of the algal community in shaping environmental resistance groups, especially under stress conditions[17]. Additionally, studies have found that, under pollution stress, the phycosphere can transfer ARGs to the surrounding water through passive processes, such as microbial cell lysis or detachment, as well as active mechanisms, such as quorum sensing and biofilm dispersal[37]. Co-culturing bacteria and microalgae significantly increased the release of bacterial plasmid DNA into the aquatic environment, facilitating HGT among bacteria[57]. The process by which microalgae first enrich and subsequently disseminate ARGs into surrounding water makes them a critical link in the spread of ARGs within watersheds, thereby increasing AMR risk in these ecosystems.
-
In natural environments, the planktonic nature of microalgae exposes them to various environmental challenges. The development of antimicrobial resistance in the phycosphere is a complex process that involves the interplay of multiple factors[58]. In addition to the misuse of antibiotics, environmental factors, and human activities can all contribute to the development and spread of ARGs (Fig. 4).
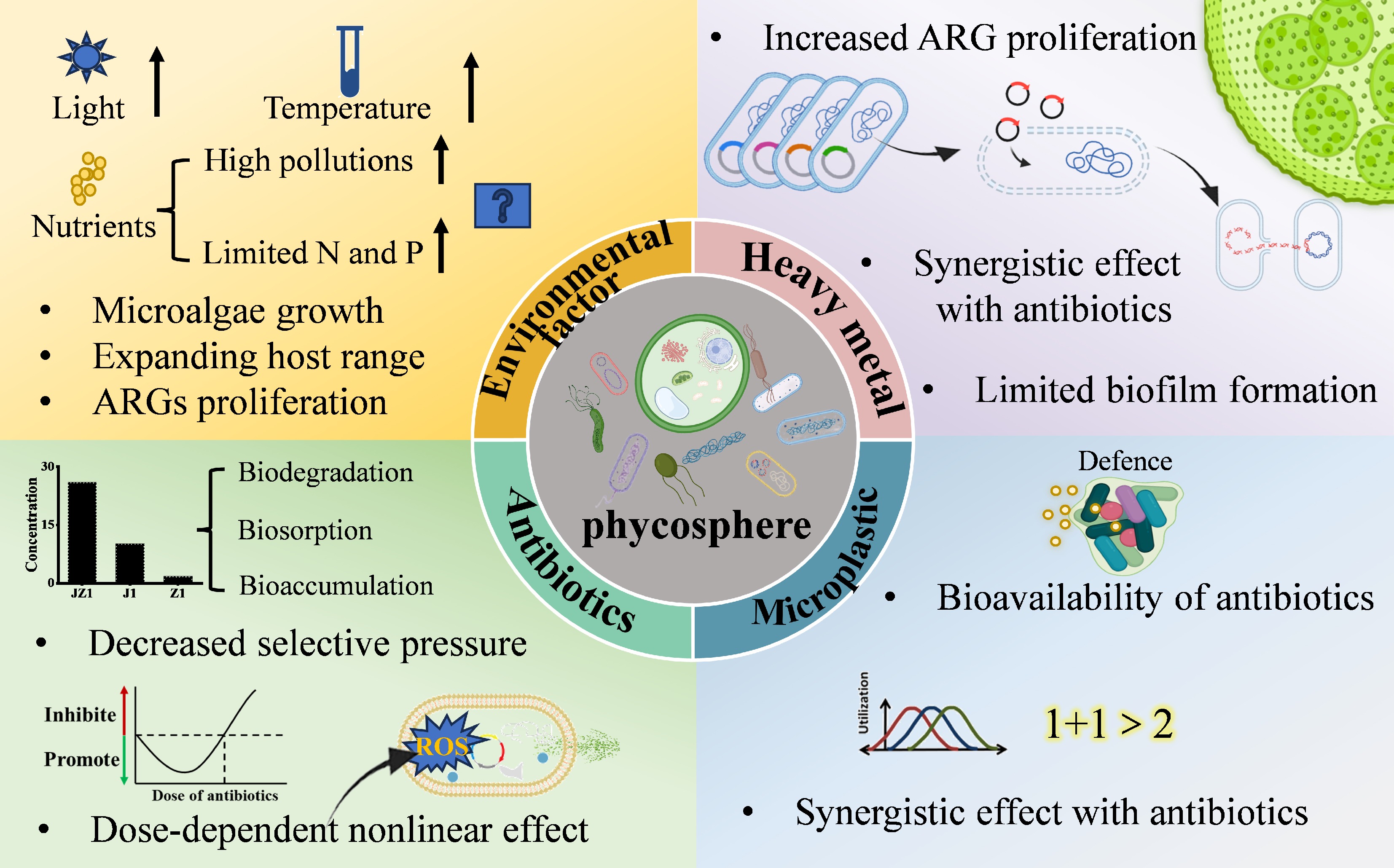
Figure 4.
The effects of contaminants on antibiotic resistance genes in the phycosphere.
Light, temperature, and nutrients
-
Light is crucial for microalgae growth, and optimal light conditions promote their proliferation. Climate change is warming aquatic environments and intensifying extreme weather events, expanding the geographical and seasonal distribution of harmful microorganisms, and increasing risks to human health, food security, and the economy[59]. Elevated temperatures and higher cyanobacterial concentrations favor the conjugative efficiency of bacterial assimilation of ARGs[52]. The five classic aquatic environments, namely seas, lakes, rivers, bays, and estuaries, are affected by human activities to varying degrees. Due to the input of land-based pollutants and human influence, freshwater environments are usually less stable than marine environments. Therefore, the interactions between freshwater microalgae and their associated bacteria are more susceptible to interference and remodeling. In marine environments, human activities influenced the expression of ARGs through impacts on nitrogen, phosphorus, and temperature, with high-risk ARGs tending to transfer towards lower latitudes and areas of intensive human activity[60]. Increases in nutrients such as nitrogen and phosphorus may promote microalgal growth, thereby providing more abundant niches for the colonization of bacteria/resistant bacteria. In polluted waters, the abundance, richness, and diversity of phycosphere epiphytic bacteria increased significantly, strengthening connections between algae and bacteria and inducing a more complex and stable ecological network in the phycosphere. Water pollution expanded the host range of phycosphere ARGs, significantly stimulated the directional selection of bacteria towards microalgae, and enhanced the proliferation of ARGs[17]. Therefore, adequate availability of nitrogen, phosphorus, and carbon facilitates and drives the proliferation of ARGs within microalgae-bacteria systems. However, limiting nitrogen or phosphorus concentrations in water bodies increased the richness and diversity of phycosphere epiphytic bacterial communities and promoted the proliferation of ARGs in the phycosphere as well[61]. Significant environmental differences exist between the phycosphere and the surrounding water, particularly in terms of core bacterial communities and physicochemical properties. The formation of microbial communities is influenced by both stochastic and deterministic processes. Deterministic processes are typically driven by niche selection, influenced by environmental factors and species interactions, while stochastic processes involve random events such as dispersal and drift[62]. Jiao et al.[63] found that in eutrophic water environments, the assembly processes of phycosphere communities are more dominated by stochastic processes.
Antibiotics
-
Antibiotics, as the primary drivers of AMR development, have received the most extensive attention in research on epiphytic bacterial resistance in the phycosphere. They are widely used globally for the prevention and treatment of diseases in humans and animals, which inevitably leads to the discharge of residual antibiotics into rivers[64]. Although antibiotic concentrations in rivers are relatively low, ranging from ng/L to μg/L[65], the continuous release of these pollutants exerts persistent selective pressure on epiphytic bacteria in the phycosphere. Antibiotics generally do not cause acute toxicity to the growth and survival of larger aquatic species, such as fish and invertebrates. However, they can directly affect particular phytoplankton species[66]. These effects are typically sublethal, manifesting as changes in physiological and morphological characteristics, such as the inhibition of photosynthetic activity and protein synthesis[67,68]. Cyanobacteria are often the most sensitive algal species to antibiotic pollution because their cellular structure closely resembles that of bacteria[69]. When antibiotics enter the phycosphere environment, they exert disruptive and toxic effects on the resident microorganisms. While both bacteria and microalgae contribute to removing antibiotics from water, they also mount specific responses to antibiotic stress. The development of AMR in phycosphere microorganisms represents an adaptive mechanism and survival strategy to cope with antibiotic contamination.
Antibiotic stress on the phycosphere system exhibits a typical 'dose-dependent non-linear effect'[70]. This likely describes a phenomenon in which low, sub-inhibitory concentrations of antibiotics can stimulate stress responses and significantly promote Horizontal Gene Transfer (HGT). In contrast, high bactericidal concentrations may kill bacteria, thereby impeding HGT opportunities and resulting in a non-linear correlation between antibiotic dosage and ARG abundance. Antibiotic-induced dose-dependent non-linear effects on the phycosphere are regulated by exposure time scales (acute vs chronic). Acute exposure (short-term, high-dose; e.g., 1 mg/L Sulfadiazine (SDZ) for hours to 3 d) rapidly restructures the microbiome, with nonlinearity shown as 'high-dose inhibition (reducing ARG hosts) vs low-dose slight stimulation (negligible ARG enrichment)', due to insufficient time for MGE-mediated HGT. Chronic exposure (long-term, low-dose; e.g., 0.001 mg/L SDZ for weeks to months) exerts sustained selection pressure, leading to non-linear 'sustained ARG enrichment even at low doses' via recruiting beneficial symbionts, facilitating ARG host colonization, and enhancing HGT, which ultimately increases ARG abundance by one order of magnitude vs the control. This timescale dependence reflects the dynamic nature of the effect, critical for explaining ARG dissemination risk in rivers under continuous low-concentration antibiotic pressure[70]. The addition of sulfadiazine to a microalgae-bacteria system resulted in ARG abundance in the treatment group being approximately one order of magnitude higher than in the control group[71]. Antibiotic stress prompts the phycosphere to recruit more symbiotic bacteria that support the growth and reproduction of Chlorella. At the same time, antibiotic-induced specific assembly of bacterial communities and MGEs drives the formation of a distinct ARG profile in the phycosphere[50]. Furthermore, antibiotic exposure promotes the colonization of potential ARG hosts in the Chlorella phycosphere, leading to the specific enrichment of ARGs around the phycosphere and even enhancing the potential for bacteria carrying ARGs to spread from the phycosphere to broader environmental niches[71,72]. Exposure to antibiotics such as tetracycline, hygromycin, and enrofloxacin induces closer contact between microalgae and bacteria[73,74]. In a previous study, a microcosm of microalgae-bacteria co-culture was established to investigate the transmission of ARGs in the phycosphere under sulfamethoxazole pressure. Microalgae enhance the adaptability of epiphytic bacteria to antibiotic stress, increase the diversity of the phycosphere epiphytic bacterial community, and promote the proliferation of ARGs[16]. Moreover, even low concentrations of antibiotics can induce the proliferation of ARGs in the phycosphere[75]. Therefore, the continuous exposure to low concentrations of antibiotics in rivers may play a significant role in the dissemination of ARGs within the phycosphere.
Beyond their facilitative effects, antibiotic stress can trigger a stress response in microalgae-bacteria consortia, wherein specific ROS can act synergistically with sunlight to degrade ARGs[76]. Additionally, the high metabolic activity of phycosphere epiphytic bacteria contributes to antibiotic removal[77]. Under antibiotic stress, EPS content in the microalgae-bacteria system increased significantly, with a notable rise in the proportion of proteins (PN) within the EPS, thereby limiting antibiotic penetration into cells[33]. These findings demonstrate that microalgae and bacteria can collaborate to reduce antibiotic exposure, thereby mitigating the potential environmental selection pressure on phycosphere epiphytic bacteria. However, the distribution and enrichment patterns of antibiotics within this natural microecosystem, the phycosphere, remain poorly understood. One study revealed that both concentrated pollution sources from wastewater treatment plants and moderately dispersed pollution upstream of treatment plants leads to detectable 'resistance selection pressure' from antibiotics in rivers, existing both upstream and far downstream of WWTP discharges. This indicates that the impact of antibiotic pollution from WWTPs on phycosphere ARGs can extend to areas distant from the pollution source[78], suggesting a potential underestimation of the risks associated with antibiotic-mediated dissemination of AMR.
Microplastics
-
Microplastics have recently gained significant attention, and biofilms on plastic particles (plastispheres) are also recognized as hotspots for the transmission of ARGs. Microplastics can likewise impact microalgae and their phycosphere. Microalgae can adapt to microplastic stress through homo- and hetero-aggregation, while microplastics can influence the bioavailability of antibiotics[72]. Combined exposure to the exogenous contaminants, microplastics, and florfenicol increased the abundances of ARGs, MGEs, and potential ARG hosts within the phycosphere, thereby promoting the dissemination of ARGs between the surrounding environment and the phycosphere[72].
Heavy metals
-
Heavy metals are widely distributed in river systems. Under copper stress, antibiotic-resistant bacteria (ARB) form biofilms more readily than antibiotic-sensitive bacteria (ASB), as reflected in higher bacterial abundance and elevated EPS content in ARB biofilms[79]. Under pollutant stress, the relative abundance of ARGs increases. Moreover, at environmentally relevant concentrations, antibiotics have a greater impact on ARGs than silver nanoparticles (AgNPs) and hematite particles. The combined effect follows this order: antibiotics + metallic nanoparticles > antibiotics > metallic nanoparticles[75]. Both heavy metals and nanoparticles can reduce the expression of genes related to bacterial motility and/or chemotaxis, thereby inhibiting cell adhesion and biofilm formation[80], which may impede the enrichment of ARGs in the phycosphere.
-
This study reviews the formation and characteristics of the phycosphere, and the enrichment and spread of ARGs within it, demonstrating that the phycosphere serves as a crucial ecological niche for the proliferation and transmission of ARGs. The widespread distribution of microalgae in aquatic environments, coupled with their exposure to various contaminants, highlights their significant role in the dissemination of ARGs within watersheds. Such microscopic-scale research provides a theoretical foundation for curbing the global spread of AMR. However, the study of microalgae-mediated abundance and spread of ARGs in watersheds are still in their early stages, and many key questions remain theoretical. The main challenges and future directions are outlined below.
Microalgae-mediated proliferation and transmission of ARGs in complex watershed environments
-
Research on the transmission of ARGs mediated by microalgae has predominantly focused on laboratory studies under single-variable controls, and specific microalgal bloom events. In actual aquatic ecosystems, particularly during non-bloom periods, the cross-media occurrence patterns and dissemination mechanisms of ARGs within the water-microalgae system under complex multi-environmental factors remain unclear. Furthermore, the current understanding of the directionality of ARG transmission between microalgae and their surrounding environment is based on source-sink speculation, with no direct evidence to substantiate this process. Some studies have shown that bacteria can horizontally transfer specific genes to microalgae[81,82]. Finally, under pollution stress, the tolerance genes in bacteria may be horizontally transferred to algae, becoming an essential mechanism for algae to cope with environmental stress[80]. However, the transfer of ARGs from bacteria to microalgae (particularly eukaryotic microalgae) has not yet been conclusively demonstrated. Fluorescently labeled resistant bacteria or more in-depth genomic data analysis may resolve this issue. However, key challenges were as follows: fluorescence tracers suffer from poor stability in complex water matrices due to interference from dissolved organic matter (DOM), indigenous microbes, or marker dilution via bacterial reproduction, while genomic and transcriptomic analyses struggle with detecting low-abundance ARGs, exogenous ARG integration in complex microalgal genomes, and the transient expression of conjugation-related genes.
To address these bottlenecks, optimized technical strategies are proposed: fluorescence tracking adopts dual-tracer systems (fluorescent dyes combined with stable isotopes) validated in simulated complex environments; genomic studies integrate metagenomic binning with digital droplet PCR (ddPCR) or targeted gene capture sequencing; transcriptomic analyses use targeted RNA-seq focusing on conjugation genes (e.g., tra/trb) complemented by proteomic validation, such as Western blot. To further clarify the impacts of complex multi-environmental factors on phycosphere ARG dynamics, and make future research directions more concrete, a tiered experimental framework is proposed. First, a Plackett-Burman design (PBD) will be employed to screen the key environmental variables with significant effects on ARG transmission efficiency from a candidate pool, including temperature, nitrogen (N)/phosphorus (P) nutrient levels, antibiotic concentrations, pH, and light intensity. On this basis, a full-factorial design coupled with response surface methodology (RSM) will be applied to quantify the individual, synergistic, and antagonistic effects of core factors (e.g., temperature gradients of 15/25/35 °C, TN levels of 0.5/2/5 mg/L, and tetracycline concentrations of 0.1/1/10 μg/L) on ARG horizontal transfer frequency and microbial community structure in the phycosphere. Moreover, the optimized tracing and omics techniques mentioned above will be integrated into this multi-factor experimental system to identify the molecular regulatory mechanisms underlying the environmental factor-driven ARG dynamics. Finally, laboratory-scale results will be validated using in-situ mesocosm systems constructed with natural water samples to ensure ecological relevance. These approaches aim to obtain reliable visual and molecular evidence and are expected to reveal the mechanisms and directionality of ARG transmission from the phycosphere to the surrounding water, while providing a standardized methodological paradigm for disentangling the interactive effects of multi-environmental factors on aquatic ARG behavior in future studies.
Adaptive evolution of ARGs in the phycosphere
-
The four mechanisms of AMR evolution include spontaneous resistance mutations, HGT of ARGs, selection of pre-existing resistance, and immigration of resistant lineages[83]. Among these, spontaneous mutations and the HGT of ARGs represent processes that create resistance de novo. In contrast, the selection of pre-existing resistance and the immigration of resistant lineages are ecological evolutionary mechanisms manifested through microbial self-regulation to adapt to the environment. However, research on how environmental heterogeneity influences the evolution of pre-existing ARGs has predominantly been conducted in clinical settings. Studies in natural ecological media have focused mainly on mutation and horizontal transfer. Significant environmental gradients exist between the phycosphere and the surrounding water environment, enabling bacteria to adapt to changing conditions by regulating metabolic pathways and gene expression[84]. Cascade effects among microalgae, bacteria, and viruses within microalgae-bacteria granular sludge highlight the interconnected co-evolutionary dynamics of these microorganisms[85]. Similarly, biofilm-based studies have revealed that the evolutionary pathways of antibiotic resistance depend on environmental structure and bacterial lifestyle (sessile/planktonic)[86]. Nevertheless, after cross-media transmission of antibiotic resistance genes from water to microalgae, the mechanisms by which they adapt to the microecological environment of the phycosphere and subsequently drive the evolution of these genes remain to be elucidated. Thus, it is critical to investigate whether the phycosphere microenvironment selectively enriches for ARG variants located on mobilizable plasmids rather than chromosomes, or favors mutations that confer fitness advantages in this unique niche.
-
Not applicable.
-
The author confirm contributions to the paper as follows: Shaojing Sun: conceptualization, writing − original draft; Chao Chen: methodology; Jie Wang: investigation; Yan Sun: data curation; Qing Wang: funding acquisition, supervision. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
This work was supported by the National Natural Science Foundation of China (Grant Nos 42477469, 42507546); Hebei Natural Science Foundation (Grant No. B2024402004); Science and Technology Project of Hebei Education Department (Grant No. QN2024101); State Key Laboratory of Water Pollution Control and Green Resource Recycling Foundation (Grant No. PCRRF25032); Post-graduate's Innovation Fund Project of Hebei Education Department (Grant No. CXZZSS2025085).
-
The authors declare that they have no conflict of interest.
-
Full list of author information is available at the end of the article.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun S, Chen C, Wang J, Sun Y, Wang Q. 2026. Microalgae in the enrichment and spread of antibiotic resistance genes in watersheds: a review. Biocontaminant 2: e003 doi: 10.48130/biocontam-0025-0028 

Microalgae in the enrichment and spread of antibiotic resistance genes in watersheds: a review
- Received: 07 November 2025
- Revised: 16 December 2025
- Accepted: 19 December 2025
- Published online: 04 February 2026
Abstract: The widespread use of antibiotics has led to antimicrobial resistance (AMR), which has become one of the most serious threats to global health. The coexistence of microalgae and bacteria enhances the abundance and transmission of antibiotic resistance genes (ARGs), thereby increasing their environmental hazard. This study summarizes the roles of microalgae in the enrichment and spread of ARGs. First, the characteristics of microalgae and the features of the phycosphere microenvironment are systematically reviewed. Second, the role and mechanisms of ARG enrichment and spread mediated by microalgae are discussed. As the core innovation in this article, scattered research evidence is integrated to comprehensively elucidate the environmental drivers and underlying mechanisms that render the phycosphere a critical niche for the proliferation and transmission of ARGs. Furthermore, the contaminants influencing the transfer of ARGs within the phycosphere further confirm the elevated ARG risks induced by microalgae in watersheds. However, research on ARGs in the phycosphere remains in its infancy. Research challenges and prospects related to mechanism, transmedia spread, and adaptive evolution are also highlighted. This timely synthesis consolidates fragmented studies, clarifies the pathways of microalgae-mediated ARG transmission, and provides the scientific community with a clearer understanding of this hidden environmental process. It also offers a scientific basis for assessing and intervening in the diffusion of ARGs in watersheds.
-
Key words:
- Microalgae /
- Antibiotic resistance genes /
- Bacteria /
- Spread /
- Contaminants