









-
The plant cuticle is an extracellular lipid structure deposited on the surface of terrestrial plants that seals off aerial organs and protects them from biotic and abiotic stresses. It is composed of a cutin polymer matrix and wax, which are produced and secreted by epidermal cells[1]. GDSL genes are essential for wax biosynthesis and the formation of stomatal cutin protrusions. As a class of hydrolases with broad substrate specificity, GDSL genes are widely distributed across various organisms and exhibit both esterases and lipase activities. For the sake of consistency, they will be collectively referred to as GDSL lipases hereafter. Mutations in GDSL lipase genes leads to a significant reduction in leaf wax synthesis and impaired stomatal closure, thereby increasing epidermal permeability reducing the transpiration rate[2]. Classified as a distinct subfamily from classical lipolytic enzymes, GDSL lipases(GELPs) are characterized by a conserved G × S × G motif and contain four conserved amino acid residues—serine (S), glycine (G), asparagine (N), and histidine (H)—which are located within at least four conserved structural domains. Consequently, GELPs are also known as SGNH lipase[3,4]. Unlike other lipases, GELPs can undergo conformational changes in response to different substrates through the an induced-fit mechanism[5,6], enabling them to exhibit broad catalytic versatility and hydrolyze a wide range of substrates, including thioesters, aryl esters, phospholipids, and amino acids[7].
GELPs are widely distributed in microorganisms, animals, and plants, where they play crucial roles in growth, development, and stress responses. However, research on plant GELPs began relatively late[8,9]. In 1995, Upton & Buckley first reported a GELP in Aeromonas hydrophila, and named it based on its conserved structural domain characteristics(pfam PF00657)[7]. With the development of genomics and bioinformatics, a large number of GELPs have been identified in an increasing number of plants species. For instance, the model plant Arabidopsis thaliana harbors 105 members[10], of which Oryza sativa contains 114 members[11]. Pyrus spp. has 113 members[12]. In the genomes of other sequenced species, such as the grain crops sorghum and maize[13,14], there are 130 and 53 GELP members. Oilseed crops like soybean and sunflower have 194 and eight members, respectively[15]. Ricinus communis contains 96 GELPs members[16]. Through comparative analysis of the amino acid sequences of 604 GELP genes encoded by different terrestrial plants, it was found that members of the plant GELP family form three subfamilies (subfamily A, B, and C) in the phylogenetic tree, each containing GELP genes from different plant species[13].
In recent years, the biological functions of GELP family members have been gradually elucidated. Several GELPs have been shown to participate in plant growth, development, and organ morphogenesis[17]. For example, the Arabidopsis AtGELP gene AtEXL6 is involved in pollen development[18]. AtOSP1 (occlusion of stomatal pore 1), an endoplasmic reticulum and lipid-localized GELP, plays a role in early wax biosynthesis and stomatal epidermal wall formation[2]. AtGELP7 is a plasma membrane-localized protein that enhances saccharification efficiency[19]. The expression of many plant GELP genes can be induced by pathogens, hormones such as salicylic acid, ethylene, and jasmonic acid, as well as abiotic stress factors, suggesting their potential involvement in plant disease resistance and stress responses. For example, AmGDSL1 is most highly expressed in leaves, and is induced by drought stress, with its expression level showing a positive regulatory trend as stress duration increases[20]. Overexpression of longan DlGDSL57 activates the antioxidant system and contributes to the high-temperature stress response[21]. Cosuppression of AtGELP22 and AtGELP23 enhances drought resistance by regulating the ABA-mediated signaling pathway[22]. Furthermore, GELPs are widely involved in the synthesis of plant secondary metabolites and fatty acid metabolism in oilseeds, and can catalyze the formation of lutein esters[13]. When the cotton GELP gene GhGL1 was transferred into Brassica napus, seed oil content increased by 8.68%[23]. A rapid decline in GELP gene expression in Shatian grapefruit results in impaired intracellular ester metabolism[24].
With the exception of a 2024 study on the molecular mechanism by which the apple lipase MdGELP1 inhibits anthracnose infection[25], research on MdGELPs remains limited. As the most economically valuable cultivated fruit tree in the Rosaceae family, apple (Malus domestica Borkh.) is widely grown in temperate regions worldwide, and ranks first globally in both production and cultivation area. Apple wax is a critical component of the fruit cuticle, playing a essential role in apple development, storage, and adaptation to environmental stress. The wax covering the apple surface directly determines its appearance quality and market value[26]. In this study, we used bioinformatics approaches to characterize the genome-wide members of the MdGELPs family, including their gene and protein structures. Additionally, we employed fluorescence qRT-PCR to analyze the tissue-specific expression of representative MdGELPs gene and investigated changes in their expression patterns under low-temperature, drought, and salt stresses. These findings provide a foundation for further exploring the regulatory mechanisms of wax synthesis and stress response in apple.
-
The experiment was conducted from January 2025 to June 2025 at Shandong Agricultural University.
Plant materials and treatments
-
Two apple materials were selected for this study. Twenty-year-old'Royal Gala'apple trees, planted at the Horticultural Experimental Station of Shandong Agricultural University (Tai'an, Shandong, China), were used to collect roots, young stems, leaves, flowers, and pericarp, and pulp tissues separately (130 d after flowering). All tissue samples were immediately snap-frozen in liquid nitrogen after collection and stored at −80 °C in an ultra-low-temperature refrigerator for subsequent tissue expression analysis. Malus hupehensis seedlings that had been cultured under laboratory conditions for approximately 4 months and exhibited uniform growth were selected for abiotic stress analysis. Three experimental treatments were applied to the M. hupehensis seedlings, with three biological replicates per treatment. For ABA treatment, three seedlings were transferred to a hydroponic system containing 10 μM abscisic acid (ABA), and whole-plant samples were collected at 1, 2, 3, 6, 12, 24, and 48 h after treatment. For low-temperature stress treatment, three seedlings were incubated in a 4 °C light incubator, and whole plants were sampled at 0, 1, 2, 3, 6, 12, 24, and 48 h after treatment. For salt stress treatment, three seedlings were incubated in a hydroponic system containing 150 mM NaCl, and whole plants were sampled at 1, 2, 3, 6, 12, 24, and 48 h after treatment. All samples from the three treatments were quickly frozen in liquid nitrogen and stored at −80 °C for subsequent abiotic stress expression analysis.
Identification of the MdGELPs genes in apple
-
To identify MdGELP family members across the entire Malus domestica genome, we first downloaded 105 AtGELP protein sequences from the TAIR database (
https://arabidopsis.org )[10]. Sequences shorter than 300 amino acids (aa), or longer than 350 aa, were filtered out, leaving 16 AtGELP protein sequences that were used as query sequences. A local BLASTP search was performed against the complete apple protein sequence library with an E-value threshold of 1e-5. The M. domestica whole-genome sequence and GFF3 genome annotation data were retrieved from the Genome Database for Rosaceae (GDR,https://rosaceae.org ). Additionally, the GDSL conserved domain (PF00657) file was downloaded from the Pfam database (https://pfam.xfam.org ), and an HMMSEARCH was conducted with an E-value cutoff of 1e-5. The candidate MdGELP proteins obtained through these two approaches were deduplicated and integrated using TBtools[27]. Subsequent domain confirmation via NCBI Batch CD-Search led to the identification of 38 MdGELP family members, which were designated as MdGELP1 to MdGELP38.Analysis of MdGELP proteins
Physicochemical properties of proteins
-
The molecular weight (MW), isoelectric point (pI), amino acid (aa) length, and other physicochemical properties of the 38 MdGELP proteins were calculated using the Phylogenetics plugin in TBtools.
Multiple sequence alignment and conserved domain analysis
-
Multiple sequence alignment of the 38 MdGELP protein sequences was performed using DNAMAN (
https://dnaman.software.informer.com ) with default parameters, and the alignment results were visualized via the online tool WebLogo (https://weblogo.berkeley.edu/logo.cgi ). Conserved domains of MdGELP proteins were analyzed using the online Batch CD-Search tool provided by the National Center for Biotechnology Information (NCBI), and the results were visualized with TBtools.Phylogenetic analysis
-
Using the previously obtained 16 AtGELP and 38 MdGELP protein sequences, a phylogenetic tree was constructed with MEGA7 using the neighbor-joining (NJ) method, and 1,000 bootstrap replications for validation. The phylogenetic tree was optimized and visualized using the online software Evolview (
https://evolgenius.info/evolview-v2/#login ).Conserved motif analysis
-
Conserved motif analysis was performed using the 'Simple MEME Wrapper' plugin in TBtools, with the number of motifs set to 8.
Secondary and tertiary structure analysis
-
The secondary structures of the 38 MdGELP proteins were predicted using SOPMA (
https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html ). Homology modeling of the protein three-dimensional (3D) structures was conducted via the online tool SWISS-MODEL (https://swissmodel.expasy.org ).Prediction of transmembrane domains, signal peptides, and phosphorylation sites
-
Transmembrane domains of the 38 MdGELP proteins were analyzed using TMHMM v.2.0 (
https://services.healthtech.dtu.dk/services/TMHMM-2.0 ), while the presence of signal peptide structures in these protein sequences was annotated with SignalP 4.1 (https://services.healthtech.dtu.dk/services/SignalP-4.1 ). Phosphorylation sites of MdGELP proteins were predicted using the online software NetPhos 3.1 (https://services.healthtech.dtu.dk/services/NetPhos-3.1 ).Subcellular localization prediction
-
Subcellular localization was predicted using the online tool CELLO (
https://cello.life.nctu.edu.tw ), with the organism set to 'Eukaryotes'.Protein interaction network and GO functional annotation (MdGELP to AtGELP homology mapping)
-
The protein interaction network of AtGELPs was predicted using the STRING database (
https://string-db.org ), with the organism specified as Arabidopsis thaliana. To further explore the functional homology of MdGELP proteins, we retrieved Gene Ontology (GO) term information for the 16 AtGELP proteins from the STRING database, followed by detailed bioinformatics processing and visual presentation of the data.Analysis of MdGELP genes
Chromosomal localization
-
The chromosomal location information of the 38 MdGELP genes was extracted from the genome annotation file (gene_models_20170612.gff3), and submitted to TBtools for visualization of gene chromosomal distribution.
Gene structure (exon-intron) analysis
-
Using the gene GFF annotation data, the exon-intron structures of the 38 MdGELP genes were visualized with TBtools.
Analysis of MdGELP gene promoter cis-acting elements
-
The 2,000 bp upstream fragment of the translation start site (ATG) in the apple genome was defined as the promoter region. Promoter sequences of the 38 MdGELP genes were submitted to PlantCARE (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ), and cis-acting elements were identified using default parameters. The results were visualized via TBtools.Intraspecific and interspecific collinearity analysis
-
To explore genetic homology across plant species, genome, and annotation files of Arabidopsis thaliana and Oryza sativa were downloaded from the NCBI database. Collinearity analysis was performed using the MCScanX plugin in TBtools, and the non-synonymous substitution (Ka)/synonymous substitution (Ks) ratios of gene pairs were calculated using the Simple Ka/Ks Calculator (NG) plugin in TBtools.
Analysis of MdGELP expression patterns
-
Total RNA was extracted from different tissues (roots, stems, leaves, flowers, pericarp, and pulp) of 'Royal Gala' apple trees, as well as from Malus hupehensis seedlings sampled at various time points under ABA, low-temperature, and salt stress treatments, using an RNA extraction kit (R2060; Beijing Solarbio Science & Technology Co., Ltd, Beijing, China). RNA concentration and purity were determined using a Thermo NanoDrop 2000 spectrophotometer. Complementary DNA (cDNA) was synthesized via reverse transcription using a commercial kit (Beijing Solarbio Science & Technology Co., Ltd, Beijing, China), followed by quantitative real-time PCR (qRT-PCR) analysis performed on a QuantStudio 7 Flex system. Gene-specific primers were designed using Primer3Plus (
https://primer3plus.com ), and their specificity was verified via NCBI Primer-BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi ). All primers used in this study are listed in Table 1, with 18S rRNA serving as the internal reference gene for apple. Relative gene expression levels were calculated using the 2−ΔΔCᴛ method.Table 1. Real-time fluorescence quantitative PCR primers for relative expression of MdGELPs.
Gene Forward primer (5′-3′) Reverse primer (5′-3′) MdGELP2 GACTTTGATGAAGCGTTGAGGAG GAATGAACGACCCGAGTCTTTTG MdGELP3 AAATAGGGATTGCTCCGAGACTG TAACGAGTACACAGGACGCTAAC MdGELP6 TGCTAGATCTCGCCAATTCTCTC CGACTAGTTGTGGGTTGAACAAG MdGELP8 CTTTGAAGGGGGAGTGAATTTCG CTCCCATGAGAATTAGAGAGCCC MdGELP14 GTTACAACAATGTGGCCTTGGAG GTAGCCCATTTCAAACATCCCAG MdGELP18 CCAAAACACAGTGTCTCAAGTGG GTTGACCGTAATCGTGAATGAGC MdGELP26 ATCTATATGCACTGGGAGCAAGG AGCGGTTGGTAGATATCAAGGAC MdGELP28 GCAGCAGGTATTCGTGATGAATC GGCGGCTAGTTGAGTATAACCTT MdGELP31 GGTGATATTGTCCAGATGCATGC GCCGGTATCAACAATCGAATCTC -
A total of 38 MdGELP genes were identified in the apple (Malus domestica) genome and designated as MdGELP1 to MdGELP38. The amino acid lengths of the encoded proteins ranged from 337 (MdGELP36) to 419 (MdGELP21), with molecular weights (MW) of 36.76–46.37 kDa, and isoelectric points (pI) of 4.59–9.12. Most proteins (34 out of 38, 89.5%) exhibited a stability index < 40, indicating good in vitro stability. Subcellular localization prediction revealed that 21 members (55.3%) were localized to the plasma membrane, 14 (36.8%) to the extracellular space, and the remaining three (MdGELP10, MdGELP15, MdGELP29) to the cytoplasm or vacuoles. These results suggest that MdGELPs may exert diverse functional roles in membrane transport, secretion, and metabolic regulation (Table 2).
Table 2. Physical and chemical properties of MdGELPs.
Gene name Gene ID Number of amino acid Molecular weight (Da) Theoretical pI Instability index Aliphatic index Grand average of hydropathicity Subcellular localization prediction MdGELP1 MD00G1093600 365 40,038.95 6.72 26.17 78.6 −0.168 Extracellular MdGELP2 MD01G1156400 361 39,941.92 9.12 40.29 88.31 −0.106 Extracellular MdGELP3 MD02G1126900 366 40,088.96 8.42 37.79 96.69 0.029 PlasmaMembrane MdGELP4 MD02G1127000 359 39,242 8.95 34 92.56 0.009 PlasmaMembrane MdGELP5 MD02G1127100 349 38,230.71 8.67 33.06 96.07 0.02 PlasmaMembrane MdGELP6 MD04G1234300 362 39,890.29 5.26 31.61 92.18 0.002 PlasmaMembrane MdGELP7 MD05G1293600 330 36,765.6 7.54 38.02 84.52 −0.125 PlasmaMembrane MdGELP8 MD05G1304000 376 41,660.12 4.59 44.12 86.14 −0.002 PlasmaMembrane MdGELP9 MD07G1225100 361 39,543.43 8.13 37.94 91.27 0.025 Extracellular MdGELP10 MD08G1009700 381 41,885.18 6.82 31.83 93.1 −0.02 Cytoplasmic MdGELP11 MD08G1009800 380 41,341.35 5.03 40.88 95.34 0.099 PlasmaMembrane MdGELP12 MD08G1215700 377 41,683.24 9.03 33.69 81.78 −0.102 PlasmaMembrane MdGELP13 MD09G1116400 350 39,094.76 8.34 34.53 86.11 −0.052 PlasmaMembrane MdGELP14 MD09G1116500 349 38,651.25 8.34 35.09 83.87 −0.045 PlasmaMembrane MdGELP15 MD09G1116600 350 39,579.22 5.54 28.05 84.69 −0.159 Vacuole MdGELP16 MD09G1148500 349 39,053.98 6.35 30.2 88.88 −0.038 PlasmaMembrane MdGELP17 MD09G1199700 388 41,943.68 8.88 25.24 85.31 −0.036 Extracellular MdGELP18 MD10G1270600 366 40,134.91 4.95 24.81 82.1 −0.151 Extracellular MdGELP19 MD10G1270700 368 40,247.69 5.65 22.19 84.84 −0.149 Extracellular MdGELP20 MD10G1270800 362 39,373.54 5.59 34.47 84.09 −0.047 Extracellular MdGELP21 MD10G1270900 419 46,368.76 8.68 30.98 76.09 −0.252 Extracellular MdGELP22 MD10G1283000 367 40,296.18 8.52 36.87 85.64 0.028 Extracellular MdGELP23 MD10G1283400 371 41,088.41 4.65 44.34 87.82 −0.019 PlasmaMembrane MdGELP24 MD10G1286100 356 38,944.27 8.5 25.47 97.47 0.092 PlasmaMembrane MdGELP25 MD11G1058600 377 41,838.96 6.6 37.12 89.71 −0.105 Extracellular MdGELP26 MD11G1153800 354 38,492.65 5.41 28.38 92.37 0.056 Extracellular MdGELP27 MD11G1210200 349 39,157.06 7.51 26.79 89.66 −0.003 PlasmaMembrane MdGELP28 MD12G1049900 369 40,378.84 6.97 32.11 90.38 0.038 Extracellular MdGELP29 MD15G1008700 368 41,202.58 8.89 28.53 89.73 −0.111 Cytoplasmic MdGELP30 MD15G1008800 379 42,085.47 6.88 30.74 94.59 −0.005 PlasmaMembrane MdGELP31 MD15G1008900 400 44,117.95 7.49 30.63 100.15 0.075 PlasmaMembrane MdGELP32 MD15G1009100 379 42,003.4 6.02 31.44 97.18 0 PlasmaMembrane MdGELP33 MD15G1009500 370 40,696.89 6.87 37.02 93.73 0.076 PlasmaMembrane MdGELP34 MD15G1009600 362 39,653.51 4.83 22.11 95.86 0.071 PlasmaMembrane MdGELP35 MD15G1241700 373 41,369.63 9.1 32.72 88.53 −0.014 PlasmaMembrane MdGELP36 MD15G1241800 337 37,835.66 5.63 29.19 101.72 0.249 PlasmaMembrane MdGELP37 MD15G1400200 391 42,794.36 5.29 33.22 84.35 −0.137 Extracellular MdGELP38 MD17G1180300 357 38,251.51 6.5 22.84 91.06 0.099 Extracellular Phylogenetic tree and conserved region analysis
-
Phylogenetic analysis of 38 MdGELP and 16 AtGELP proteins was performed using the neighbor-joining (NJ) method with 1,000 bootstrap replicates. The resulting tree divided the family into three major clades (Clade A, B, and C), containing 25, 28, and one member, respectively. Clade B was the largest, comprising 13 MdGELPs and 15 AtGELPs, while Clade C contained only AtGELP15. Both Clade A and Clade B could be further subdivided into two subclades. Several orthologous relationships were identified, including between MdGELP27 and AtGELP9 (Fig. 1a). Multiple sequence alignment confirmed high conservation of key motifs among these proteins (Fig. 1b). Motif analysis using MEME identified eight conserved motifs, with Motif 1, Motif 3, and Motif 4 present in all members (Fig. 1c). Notably, all MdGELP proteins in Clade B contained Motif 8, suggesting that this motif may contribute to their shared functional characteristics. Conserved domain analysis revealed the presence of both SGNH and GDSL domains in all MdGELP proteins (Fig. 1d). The vast majority of Clade B proteins contained only the SGNH domain. To further explore similarities and differences among MdGELPs at the nucleic acid level, we visualized their exon–intron structures (Fig. 1e). The results showed that most MdGELPs (28/38) contained either 3 or 5 exons, with MdGELP36 having the highest (7 exons). The relatively conserved gene structures suggest similar regulatory mechanisms, while variations in intron number (2–6)[16] may reflect evolutionary divergence through intron gain/loss events[28].
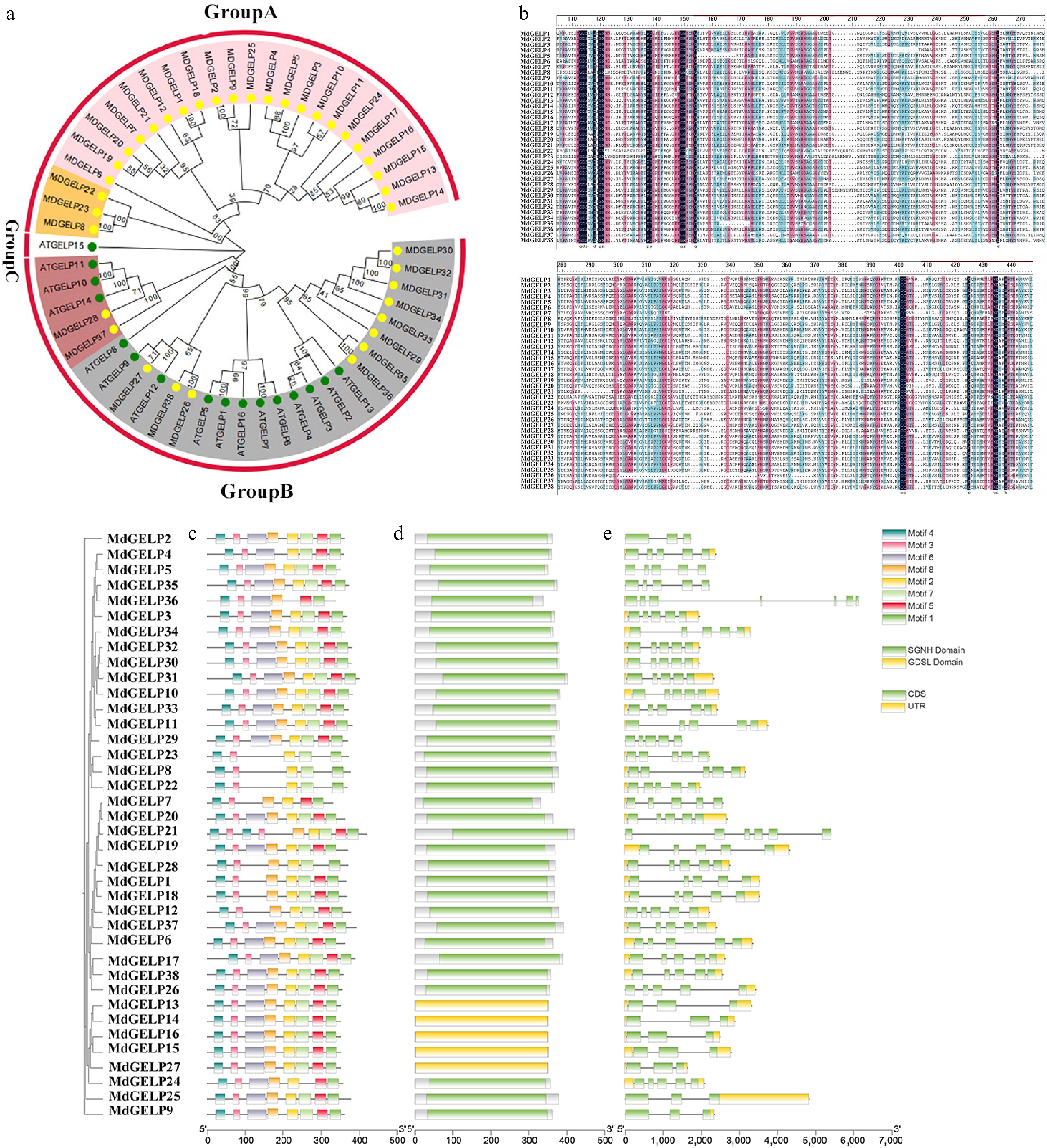
Figure 1.
Phylogenetic tree and conserved region analysis. (a) Phylogenetic tree of GELP proteins from Malus domestica and Arabidopsis thaliana. (b) Multiple sequence alignment of MdGELP family protein. (c) Distribution of conserved motifs MdGELP family proteins. (d) Distribution of conserved domains in MdGELP family proteins. (e) Gene structure analysis of MdGELPs.
Characteristics and structural predictions of MdGELP proteins
-
Secondary structure prediction revealed that all MdGELP proteins are primarily composed of α-helices and random coils, with minimal β-turns (Fig. 2a). Tertiary structure modeling showed highly similar three-dimensional (3D) conformations among family members, further supporting functional conservation (Fig. 2b). TMHMM analysis identified 13 MdGELPs (MdGELP2/8/9/12/17/18/20/22/26/28/33/36/38) containing a single transmembrane domain, suggesting potential roles in membrane-associated signaling (Fig. 2c). Signal peptide prediction indicated that 23 proteins harbor secretory signals, with the cleavage site of MdGELP37 localized between amino acids 30 and 31 (Fig. 2d). Protein phosphorylation plays a core regulatory role in plant growth and development, environmental adaptability, and signal transduction[29]. Phosphorylation site analysis detected 1,283 total sites, predominantly on serine (49.26%), threonine (27.83%), and tyrosine (22.91%) residues, with MdGELP35 protein exhibiting the highest phosphorylation site density (Fig. 2e). Collectively, these results indicate that MdGELP proteins are functionally conserved, with some members having transmembrane properties, and may be regulated through phosphorylation modifications.
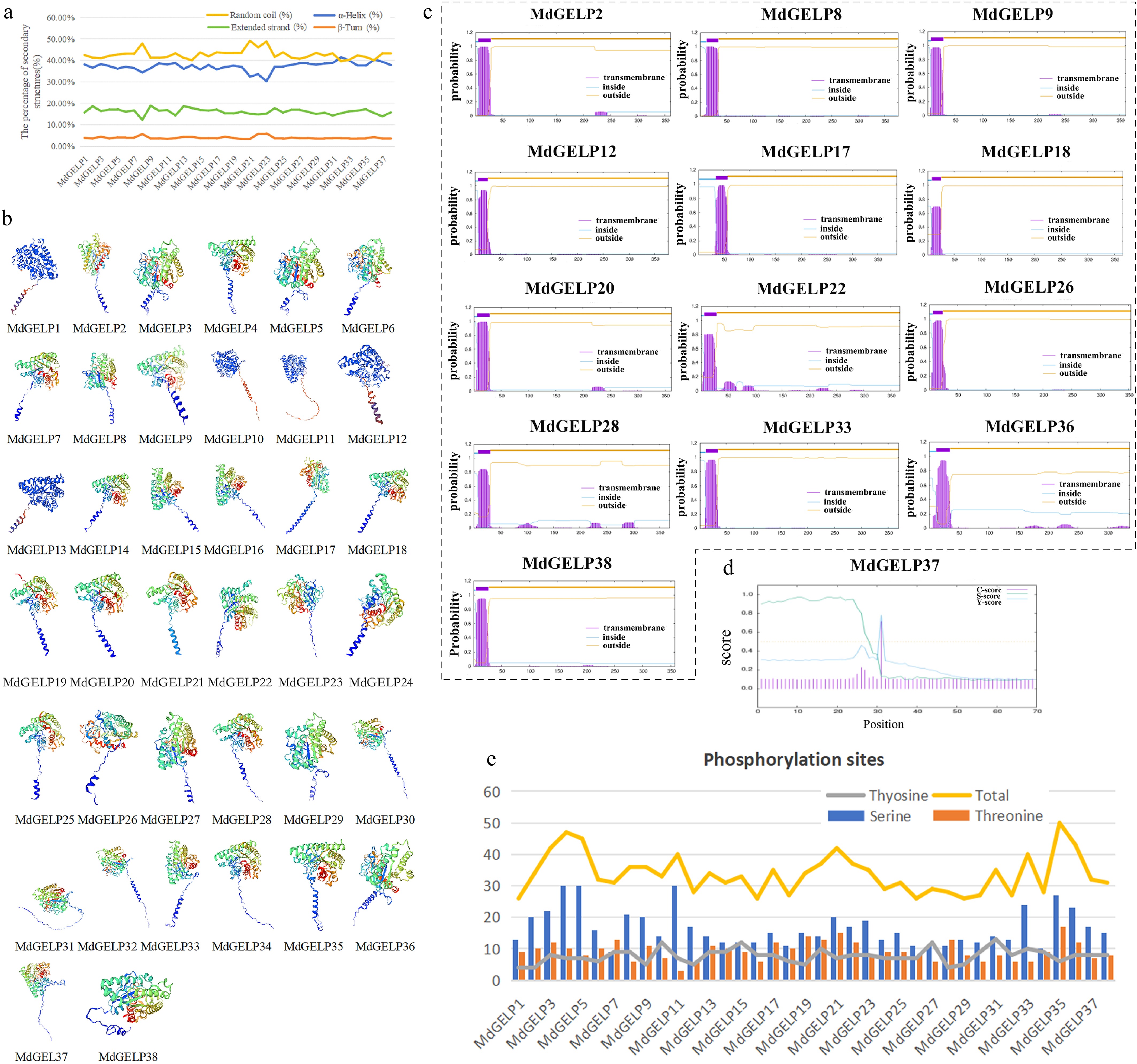
Figure 2.
Characteristics and structural prediction of MdGELP protein. (a) Secondary structure prediction of MdGELP proteins; the four colors represent different secondary structure elements, respectively. (b) Tertiary structure prediction of MdGELP proteins; the four colors represent different subunits, respectively. (c) Transmembrane domain prediction of MdGELP proteins. Gene names are listed above the plot. In the plot, the x-axis represents protein sequence length and the y-axis represents the probability of transmembrane regions. (d) Signal peptide prediction for MdGELP37. (e) Prediction of phosphorylation site distribution in MdGELP amino acid sequences.
Chromosomal localization and collinearity analysis
-
We conducted a detailed analysis of the chromosomal localization and gene structural features of the 38 MdGELP genes. These genes were mapped to 14 apple chromosomes. Notably, chromosome 15 exhibited the highest gene density, harboring 10 MdGELP genes (26.3% of the total) (Fig. 3a). MdGELP1 was located on an unanchored scaffold, indicating that it has not yet been assigned to a specific chromosome in the current genome assembly, which reflects limitations in the current annotation. Genes such as MdGELP18–21 were closely clustered on the same chromosome, suggesting potential tandem duplication events during evolution. Tandem duplication, a key mechanism of gene amplification, increases gene copy numbers and may enhance adaptive potential.To clarify the phylogenetic relationships among MdGELP family genes, we performed an intraspecific collinearity analysis (Fig. 3a). The analysis identified seven tandem duplication events, and 11 segmental duplication events within the family. Segmental duplication, which may arise from whole-genome duplication or large-scale chromosomal recombination, occurred 1.57 times more frequently than tandem duplication. Synteny analysis is a key strategy in comparative genomics for evaluating molecular evolutionary relationships between species[30]. We performed interspecific collinearity analysis among Arabidopsis thaliana, apple (Malus domestica), and rice (Oryza sativa). The results showed more collinear gene pairs between apple and Arabidopsis than between apple and rice, indicating a closer evolutionary relationship and higher functional conservation between apple and Arabidopsis (Fig. 3b). This finding enhances our understanding of plant genome evolution and facilitates the prediction of MdGELP biological functions. We calculated the Ka/Ks ratios of all paralogous gene pairs. The results showed that the vast majority of values were significantly less than 1, indicating that the MdGELP family has been primarily influenced by purifying selection during evolution (Supplementary Table S1). This analysis provides key evolutionary evidence supporting the functional conservation of the family.
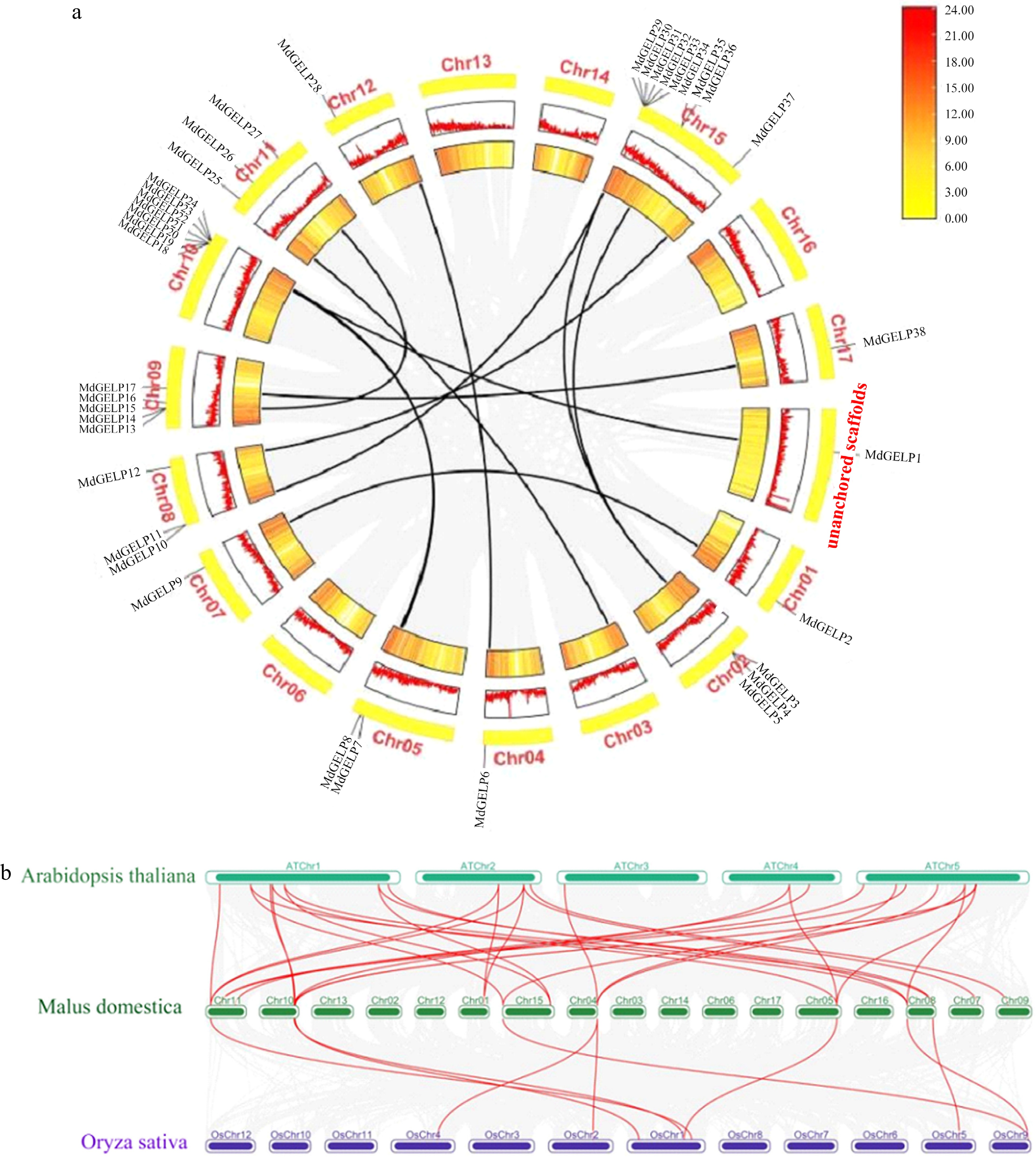
Figure 3.
Chromosomal localization and collinearity analysis. (a) Chromosomal distribution of MdGELPs and tandem/fragment repetition, with MdGELP1 located in unanchored scaffolds. (b) Collinearity analysis among apple, Arabidopsis thaliana, and rice. Dark green boxes represent the 17 apple chromosomes, green boxes represent the five Arabidopsis thaliana chromosomes, and purple boxes represent the 12 rice chromosomes. Red lines indicate homologous gene pairs.
Promoter cis-acting elements, tissue expression patterns, and responses to abiotic stress
-
Using PlantCARE, we predicted and analyzed cis-acting elements in the 2,000 bp promoter regions of the 38 MdGELP genes (Fig. 4a). These elements were associated with stress responses (e.g., anaerobic conditions, low temperature, and drought, etc.), phytohormone regulation (e.g., abscisic acid and auxin), and growth-related processes, and were classified into 18 functional categories. Among all MdGELP promoters, that of MdGELP37 contained the highest number of elements (18). To investigate the tissue-specific expression patterns of MdGELP family genes, we performed qRT-PCR analysis on nine representative MdGELPs. The results showed that these genes are broadly expressed across different tissues but exhibit distinct expression profiles in roots, stems, leaves, flowers, pericarp, and flesh. Most of the nine genes were more highly expressed in leaves and flowers (Fig. 4b, Supplementary Table S2), suggesting potential roles in the development or function of these organs. Further studies are needed to determine whether other family members share similar expression patterns. The observed tissue specificity is consistent with findings in soybean[15].
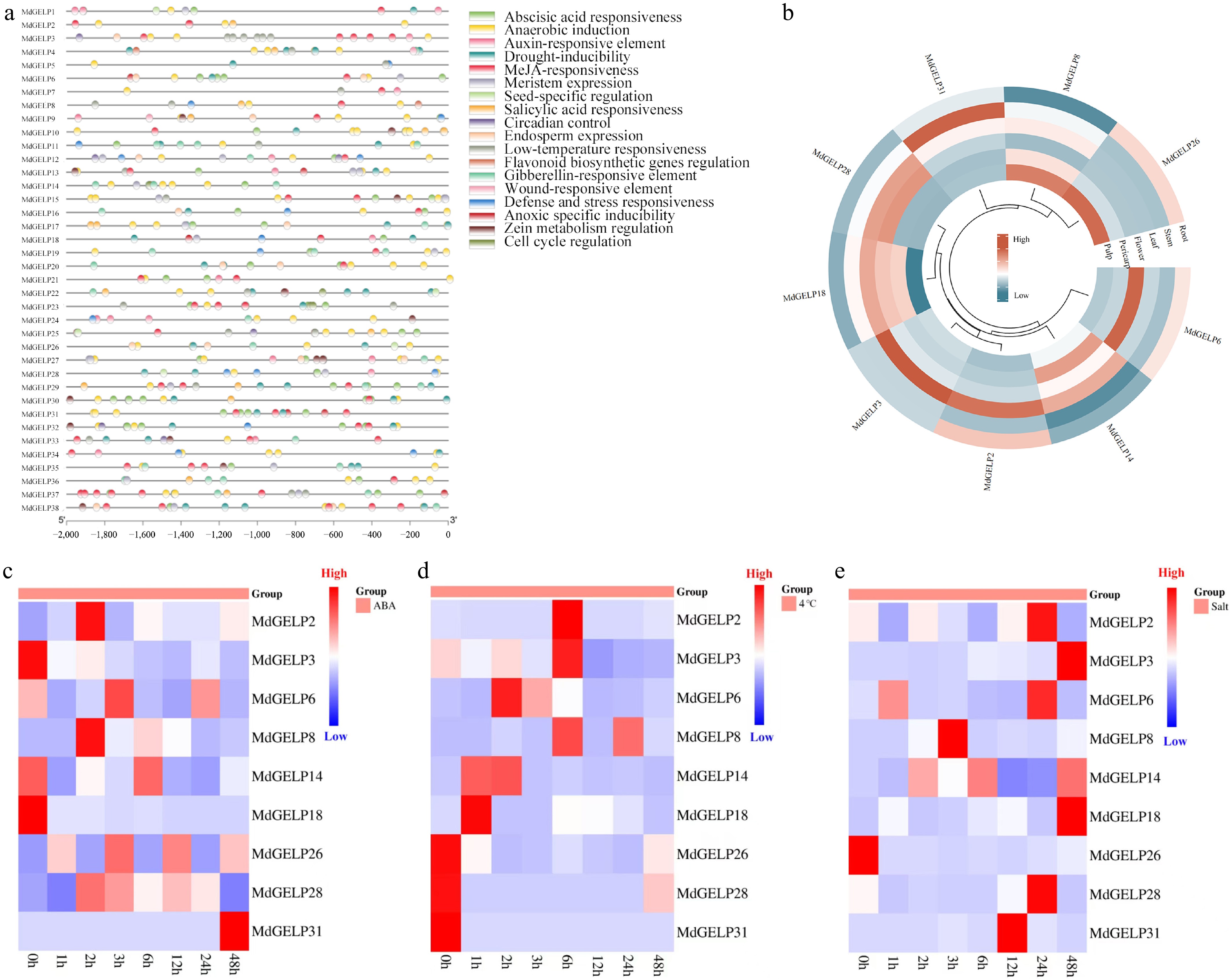
Figure 4.
Promoter cis-acting element analysis and expression profiling. (a) Analysis of cis-acting elements in the promoter regions of MdGELPs. (b) Expression levels of MdGELPs in roots, stems, leaves, flowers, pericarp, and flesh as determined by qRT-PCR. (c) Relative expression under ABA treatment. (d) Relative expression under low-temperature stress. (e) Relative expression under salt stress.
Since abscisic acid (ABA) plays a crucial role in plant tolerance to abiotic stresses such as heavy metals, drought, and extreme temperatures[31], we treated Malus hupehensis seedlings with ABA to examine MdGELP responses (Fig. 4c, Supplementary Table S3). Some genes, including MdGELP2 and MdGELP6, were initially upregulated and then downregulated, with peak expression at 2-6 h post-treatment, while MdGELP31 showed significant upregulation throughout the treatment period. Given that GELPs may be involved in plant tolerance to low-temperature stress, we analyzed MdGELP expression under low-temperature conditions (Fig. 4d, Supplementary Table S3). Except for MdGELP26, MdGELP28, and MdGELP31, the other six genes showed a pattern of initial upregulation followed by downregulation as the duration of low-temperature stress increased. Previous research has suggested that GELPs may mediate plant responses to salt stress through osmotic regulation and phytohormone signaling[32]. We therefore analyzed MdGELP expression patterns under salt treatment (Fig. 4e, Supplementary Table S3). Most MdGELPs, including MdGELP2 and MdGELP3, were upregulated. MdGELP8 showed an initial increase, followed by a decrease, with peak expression at 3 h.
Protein interaction network and GO annotation
-
The STRING database was used to predict the protein interaction network of AtGELPs, revealing several key nodes such as GRP17, which is involved in pollen wall formation[33](Fig. 5a, b). GO enrichment analysis categorized AtGELPs into three main functional groups: biological processes (e.g., lipid degradation, GO:0016042), molecular functions (e.g., lipase/hydrolase activity, GO:0016289/GO:0016788), and cellular components (e.g., extracellular region/extracellular space, GO:0005576/GO:0070505). These results support the conserved roles of GELP proteins in lipid metabolism and stress responses (Fig. 5c, Supplementary Table S4).
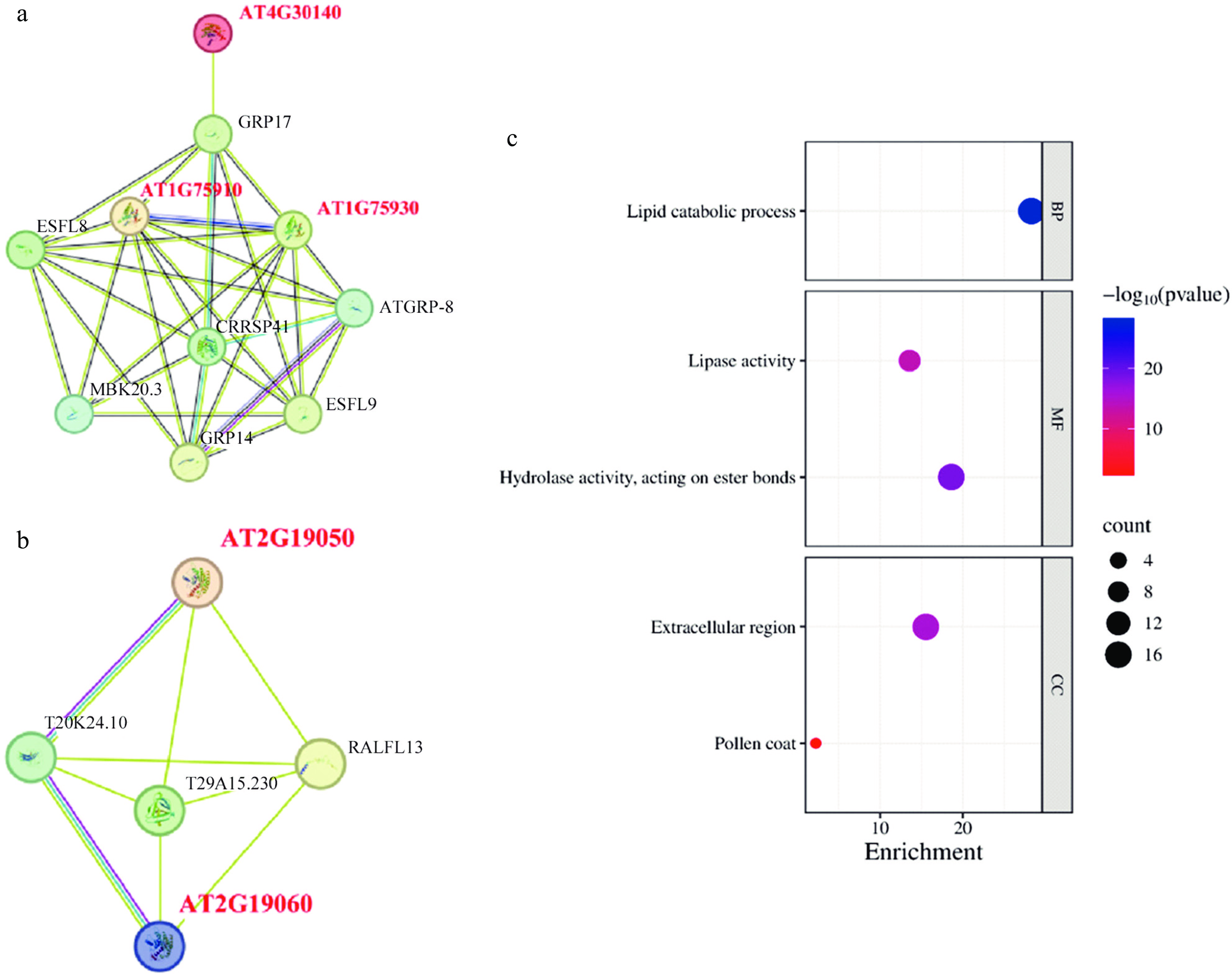
Figure 5.
Protein interaction network and GO annotation. (a), (b) Prediction of the AtGELP protein interaction network. (c) GO enrichment analysis of AtGELP proteins. Bubble size indicates the number of genes, and color indicates the p-value.
-
GELPs are a recently identified subfamily of lipolytic enzymes[10]. Although they play key roles in plant wax synthesis and stress responses[34], their functions in apple (Malus domestica Borkh.) remain largely unclear. In this study, we identified 38 MdGELPs containing both GDSL and SGNH domains. Bioinformatics analyses were used to characterize their gene and protein features, while expression patterns across different tissues and in response to ABA, low-temperature, and salt stress were investigated. These findings provide a foundation for understanding GELP function in apple and may be valuable for breeding stress-resilient crop varieties. Subcellular localization predictions revealed that MdGELPs predominantly accumulate in the extracellular matrix and cell membrane, consistent with previous reports[35], suggesting that GELP genes may mediate signal transduction from the extracellular environment to the inside of the cell[16]. Additionally, some proteins were predicted to localize to the cytoplasm and vacuoles. Such differential subcellular distributions imply that MdGELPs may possess diverse biological functions.
As an evolutionary concept, homology is most reliably inferred through phylogenetic analysis[36]. Our phylogenetic results indicate that this gene family is widely distributed across species ranging from lower to higher plants. In this study, MdGELP proteins were classified into three groups (Fig. 1a), consistent with previous findings that terrestrial plant GELPs form three subfamilies[37]. Most Arabidopsis thaliana GELP genes contain four introns, which aligns with our results[37]. The intron number in the GELP family varies from 2 to 5 (Fig. 1e), likely due to intron gain events. Intron acquisition and loss are crucial processes in eukaryotic evolution[38,39], contributing to differences in gene structure and functional divergence. Notably, MdGELPs exhibit highly similar gene structures and domain compositions, suggesting a conserved evolutionary trajectory. Collectively, this study provides important references for future functional research on the MdGELP family.
The expansion of gene families provides the foundation for functional diversification. Because of functional redundancy, multiple genes can be rapidly silenced or lost from duplicated genomes[40]. During plant evolution, increases in gene family size primarily occur through tandem duplication and segmental duplication events[41,42]. In Arabidopsis, approximately 44% (46/105) of GELPs form 16 tandem clusters on chromosomes, with 2–9 genes per cluster. In rice, about 47% (54/114) of OsGELPs are tightly clustered[15]. In apple, 68% (26/38) of MdGELPs exist as tandem repeats (Fig. 3a), highlighting tandem duplication as a major driver of family expansion. In addition, 11 segmental duplication events were detected in the apple genome. These findings are consistent with previous studies on GELP family evolution in terrestrial plants[37]. Collinearity analysis revealed that the number of collinear gene pairs between MdGELPs and AtGELPs is more than three times that between MdGELPs and OsGELPs (Fig. 3b). Thus, functional knowledge of AtGELPs can serve as a valuable reference for inferring the potential functions of MdGELPs.
After family expansion, members may undergo functional differentiation, and diversity in transcriptional regulation is an important manifestation of this process. To explore this, we analyzed cis-acting elements in the promoter regions of MdGELPs. To elucidate the roles of MdGELPs in apple development, we identified diverse cis-acting elements in their promoters (Fig. 4a). These results indicate that MdGELP expression may be modulated by phytohormones and environmental stresses, consistent with previous studies in Carya illinoensis[43]. Hormone-related elements, including those responsive to abscisic acid (ABA), methyl jasmonate (MeJA), and gibberellin (GA), were detected in nearly all MdGELP promoter regions, suggesting that MdGELPs are involved in apple tissue development and growth. In our study, cis-acting elements related to seed-specific regulation, circadian rhythm, and endosperm expression were identified in the promoters of MdGELP1, MdGELP3, and MdGELP6.Analogously, GER1 in rice is regulated by MeJA and light[44]. Furthermore, numerous anaerobic stress-associated elements were found, consistent with previous observations in Carya illinoensis[43]. Overall, the diverse cis-acting elements in MdGELP promoters imply that their expression is fine-tuned by multiple environmental and endogenous signals, enabling MdGELPs to perform varied functions throughout apple development.
The presence of the aforementioned regulatory elements suggests that MdGELPs may exhibit tissue-specific expression patterns. We tested this hypothesis by analyzing the tissue expression profiles of selected MdGELP genes. Studies of GELP expression in different plant tissues have shown that these genes are widely distributed across nearly all examined organs. In soybean, GmGELP22, GmGELP149, and GmGELP186 are highly transcribed in most tissues throughout the growth cycle[15]. In Brassica napus, the GELP family member BnaLIP1 is expressed in roots, stems, leaves, flowers, pods, and flower buds[45]. Consistent with these findings, the nine MdGELPs examined in this study are expressed in roots, stems, leaves, flowers, pericarp, and flesh, but their expression levels vary significantly among tissues (Fig. 4b). From an evolutionary perspective, gene duplication often increases genetic diversity and can lead to gene loss, whereas duplicated genes that undergo functional differentiation tend to be retained by natural selection[46]. The results of this study are consistent with these concepts.
Finally, to investigate the potential role of MdGELPs in stress responses, we analyzed their expression under several abiotic stress conditions. Analysis of MdGELP expression profiles under ABA, low-temperature, and salt stress (Fig. 4c–e) showed that MdGELP14 and MdGELP18 were significantly downregulated during ABA and low-temperature treatments. In contrast, MdGELP2, MdGELP3, MdGELP6, MdGELP14, MdGELP18, MdGELP28, and MdGELP31 were upregulated under salt stress. Notably, MdGELP3 was significantly induced under both low-temperature and salt stress, indicating that it may act as a core regulatory factor in cross-stress responses. In particular, it exhibited a typical 'induction followed by inhibition' expression pattern under low-temperature stress. The promoter region of MdGELP3 contains multiple elements related to low-temperature stress, which is consistent with its rapid induction during the early phase of low-temperature response (Fig. 4a). In addition, previous research has demonstrated that MeJA participates in plant defense responses against various abiotic stresses[47]. In tomato, for example, MeJA promotes abscisic acid (ABA) biosynthesis and plays a crucial role in low-temperature stress resistance[48]. The MdGELP3 promoter region was predicted to contain an abundance of MeJA-responsive elements, suggesting that MdGELP3 may integrate hormone signals to coordinate defense responses (Fig. 4a).
-
Despite the importance of the MdGELP gene family, the functions of its members remain poorly understood in apple, with only a few genes having been characterized in any detail. In this study, we identified 38 MdGELP genes from the apple genome, and classified them into three distinct groups. Promoter analysis revealed numerous cis-acting elements associated with growth, abiotic stress responses, and phytohormone regulation. Tissue-specific expression profiling showed that MdGELPs preferentially accumulate in leaves and flowers. Under abiotic stresses—including ABA treatment, low temperature, and salt exposure—MdGELPs exhibited dynamic expression changes, highlighting their potential roles in stress adaptation. Comparative analysis revealed both conserved and divergent features in the protein structures and expression patterns of MdGELPs. While sharing several similarities, these genes also exhibit subtle differences, suggesting that they have both overlapping and distinct functions in mediating responses to abiotic stress.
-
Not applicable
-
The authors confirm their contributions to the paper as follows: writing of original draft: Sha RX, Feng ZQ; formal analysis, methodology, and validation: Zhai JXY, Li T, Tian YS, Lyu HM, Jiang H, Wu S; supervision, resources, funding acquisition, writing − review and editing: Zhang YL, Li YY; made equal contributions to the article: Feng ZQ, Sha RX, Zhai JXY. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
This work was supported by the National Key Research and Development Program (2023YFD2301000), the National Natural Science Foundation of China (32472705 and 32402536), Natural Science Foundation of Shandong Province (ZR2022JQ14), and Taishan Scholar Project.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/frures-0026-0004.
- Supplementary Table S1 Paralogous gene pairs and Ka/Ks ratios among MdGELP genes.
- Supplementary Table S2 Expression of MdGELP genes in different tissues.
- Supplementary Table S3 Expression of 9 MdGELP genes following abiotic stress treatments.
- Supplementary Table S4 Gene Ontology annotation of AtGELP genes.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sha RX, Feng ZQ, Zhai JXY, Li T, Tian YS, et al. 2026. Genome-wide identification and expression analysis of GDSL-type esterases/lipases (GELP) gene family in apple (Malus × domestica Borkh.). Fruit Research 6: e014 doi: 10.48130/frures-0026-0004 

Genome-wide identification and expression analysis of GDSL-type esterases/lipases (GELP) gene family in apple (Malus × domestica Borkh.)
- Received: 05 November 2025
- Revised: 08 January 2026
- Accepted: 13 January 2026
- Published online: 14 April 2026
Abstract: The GDSL-type esterase/lipase family plays a crucial role in regulating plant growth and development, modulating responses to abiotic stresses, activating pathogen defense mechanisms, and governing lipid metabolism pathways. In this study, a comprehensive genome-wide analysis using integrated BLASTP and HMMSEARCH strategies identified 38 MdGELP genes in apple, which were mapped to 14 chromosomes. Phylogenetic analysis classified these genes into three major clades (A, B, and C). Multiple sequence alignment, combined with analyses of conserved domains and motifs, revealed a high degree of sequence conservation among MdGELP proteins. Promoter cis-acting element analysis uncovered diverse regulatory motifs, while gene duplication analysis indicated that segmental and tandem duplications were the primary evolutionary forces driving the expansion of the GELP family in apple. Collinearity analysis further highlighted significant homology between MdGELPs and AtGELPs. Quantitative expression assays revealed tissue-specific expression patterns, with MdGELPs showing preferential expression in leaves and flowers. Expression profiling under abiotic stresses (ABA, low-temperature, and salt) demonstrated their positive responsiveness to environmental challenges. This study not only establishes a theoretical foundation for exploring the biological functions of MdGELPs but also provides critical insights for future research in this field.