








-
Cannabis sativa L. (cannabis and hemp) is an important medicinal and industrial crop of the family Cannabaceae[1]. Originating from Central Asia, this annual herb has been widely cultivated for food, fiber, and medicinal and narcotic uses[2]. Cannabis or marijuana is grown primarily for psychoactive cannabinoids produced in glandular trichomes on female flowers[3], whereas hemp is a sustainable multipurpose crop valued for its efficient resource use and high-quality biomass for fiber and construction materials[4]. These two forms differ mainly in cannabinoid composition, particularly delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD), with THC being psychoactive and CBD non-psychoactive, but medically beneficial[5]. Cannabis and industrial hemp are cultivated in more than 40 countries under diverse legal frameworks, with industrial hemp defined by low THC content (generally ≤ 0.2%–0.3%)[6,7]. Despite its growing economic value, C. sativa production is constrained by pests and diseases, particularly fungal pathogens that affect yield, quality, and product safety during both pre- and post-harvest stages[8]. Favorable environmental conditions, genotype susceptibility, and improper post-harvest handling can promote fungal contamination. Among these pathogens, Fusarium is a large and diverse genus of ubiquitous filamentous fungi, distributed worldwide in soil, air, water, and plant tissues. Species of Fusarium are among the most economically important fungi, acting as major plant pathogens that cause significant yield losses in agricultural crops; producing mycotoxins that contaminate food and feed which pose serious health risks to humans and livestock, and are opportunistic pathogens of humans and animals, particularly in immunocompromised individuals. Control of Fusarium diseases remains challenging due to their broad distribution, long-term survival in soil and plant debris, and their ability to overcome host resistance[9,6].
Plant diseases, particularly root and crown rots and vascular wilts, are caused by members of the Fusarium solani species complex (FSSC), and affect a variety of plants such as soybeans, potatoes, cucurbits, peas, sweet potatoes, Chinese roses, and other legumes[10−13]. Moreover, Fusarium species are reported to be some of the most destructive pathogens affecting C. sativa, especially during the root and vegetative growth phases, leading to decreased crop quality and potential total yield loss[8]. For instance, brown rot of flower buds, foliar blight (which is characterized by mycelium covering inflorescences and leaves), and foliar and flower blight (which affects both flowers and leaves) are among the symptoms caused by several Fusarium species. These symptoms underscore the possibility of serious impacts on the plant's reproductive systems and overall health[14−16]. Furthermore, symptoms of F. oxysporum infection include internal stem decay, crown rot, and yellowing. In addition, it may cause pith browning, brown rot of the roots, root necrosis, chlorosis, and irregular wilt. Notably, it also causes damping-off in cuttings, which negatively affects seedling survival[16−19]. Similarly, F. solani and F. proliferatum are known to cause pith necrosis, crown infections, and tissue decay, all of which reduce plant health and productivity. At present, C. sativa productivity and health are seriously threatened by these Fusarium species, especially during crucial stages of growth[8,14,20].
Our ongoing work focuses on fungal pathogens associated with C. sativa (hemp) cultivated in Northern Thailand. During our investigation, we have found more than 10 pathogenic fungi, including one species that was found to be different from extant ones, and deemed to be a novel species. The objectves of this study were: (1) to characterize the novel isolate through detailed morphological descriptions; (2) to determine its phylogenetic position within the genus Fusarium by analysing DNA sequences from regions of the tef-1α, rpb2, and cal genes; and (3) to access its pathogenicity. The resulting data provides a framework for accurate quarantine screening and field diagnostics, supporting improved management of Fusarium in hemp production.
-
Symptomatic parts of C. sativa (hemp) stems were collected from an outdoor plantation of Mae Sa Noi Village, Mae Rim District, Chiang Mai Province, Northern Thailand (18°51'50.06" N, 98°50'45.57" E; elevation 1,285 m/4,216 ft). Ten symptomatic stems were randomly collected from the site. The samples were individually kept at 4 °C in sterile polyethene bags and processed in the laboratory within 24 h of collection. The single-spore isolation method on water agar (WA) was used to obtain pure cultures for subsequent molecular work, following Senanayake et al.[21]. All pure cultures were maintained at the Sustainable Development of Biological Resources Laboratory, Faculty of Science, Chiang Mai University, Chiang Mai, Thailand (SDBR-CMU), and the Kunming Institute of Botany Culture Collection, Kunming, China (KUNCC). One-month-old cultures on potato dextrose agar (PDA) were used to prepare dried fungal cultures, which were deposited as herbarium specimens in the Herbarium of the Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai, Thailand (CMUB).
Morphological study
-
The morphological analysis of Fusarium isolates was conducted as described by Wang et al.[22]. Cultural characteristics, including colony shape, colour, and odour, were noted during a 7-d incubation in the dark at 30 °C on PDA, oatmeal agar (OA), yeast extract agar (YEA), V8 juice agar (V8), and synthetic nutrient-poor agar (SNA). Colony characteristics were further observed after incubation on PDA at 30 °C for 15–30 d until sporulation. Phenotypic characteristics such as colony shape, size, colour, exudates, colony margins, spore colour and size were observed, recorded, and photographed using a Nikon ECLIPSE Ni-U compound microscope equipped with a Nikon DS-Ri2 camera[23]. For each anatomical structure (such as conidia, conidiophores, conidiogenous cells, and chlamydospores), at least 50 measurements were made using the Tarosoft (R) Image Framework program.
DNA extraction, PCR amplification, and sequencing
-
Genomic DNA from fungal mycelia was extracted using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux, Hangzhou, China) based on manufacturer's protocols. The translation elongation factor-1 alpha (tef1-α), RNA polymerase second largest subunit (rpb2) and calmodulin (cal) were amplified by the polymerase chain reaction (PCR) using the primer pairs EF1/EF2 (EF1: 5'-A TGGGTAAGGAGGACAAGAC-3', EF2: 5'-GGAGGTACCAGTGATCATGTT-3')[24,25]; RPB2-5f/RPB2-7cR (RPB2-5f: 5'-GGGGWGAYCAGAAGAAGGC-3', RPB2-7cR: 5'-CCCATRGCTTGYTTRCCCAT-3')[26], and Cal-228F/Cal-2Rd (Cal-228F: 5'-GAGTTCAAGGAGGCCTTCTCCC-3', Cal-2Rd: 5'-TGRTCNGCCTCDCGGATCATCTC-3')[25]. PCR amplification was performed in a 25 μL reaction volume, containing 12.5 μL Master Mix (mixture of EasyTaqTM DNA Polymerase, dNTPs, and optimised buffer; Beijing TransGen Biotech Co., Ltd., Chaoyang District, Beijing, China), 8.5 μL of double-distilled water (ddH2O), 2 μL DNA template, and 1 μL each forward and reverse primer (10 μM). The PCR thermal cycle program used for gene amplification is shown in Table 1. Purification of PCR products and DNA sequencing were conducted by TsingKe Company (Kunming City, Yunnan Province, China).
Table 1. The PCR thermal cycle programs for gene amplification used in this study.
Locus Primer name PCR procedures Ref. tef1-α EF1/EF2 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 50 s, elongation at 72 °C for 1 min, and final extension at 72 °C for 10 min [24,25] rpb2 RPB2-5f/
RPB2-7cR95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 1 min, annealing at 52 °C for 2 min, elongation at 72 °C for 1.5 min, and final extension at 72 °C for 10 min [26] cal Cal-228F/
Cal-2Rd94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 59 °C for 30s, extension at 72 °C for
1 min and final step at 72 °C for 10 min[25] Phylogenetic analyses
-
The reverse and forward sequences were combined and examined using BioEdit v7.2.5[27]. The sequences were BLASTn searched in GenBank (
https://blast.ncbi.nlm.nih.gov/ ) to identify taxa with the highest level of similarity (Table 2). The sequence alignment was completed using the MAFFT online server (https://mafft.cbrc.jp/alignment/server/index.html )[28] and minor modifications were performed in BioEdit v7.2.5[27] as needed. Maximum likelihood analysis (ML) was done on the CIPRES Science Gateway v3.3 (www.phylo.org/portal2 )[29] using RAxML-HPC2 on XSEDE (8.2.12)[30] with the GTRGAMMA substitution model with 1,000 bootstrap iterations. The best-fit substitution models were evaluated using MrModeltest v2.3[31]. Bayesian Inference (BI) analysis was carried out using MrBayes on XSEDE v3.2.7a[32−34] using the same platform[29]. Six Markov chains were run continuously for 10 million generations and automatically terminated when the standard deviation of split frequency exceeded 0.01. The trees were sampled every 100th generation, and the first 25% of the sampled trees were eliminated during the burn-in phase of the analysis, which was assessed using Tracer v1.7[35]. The phylograms were shown and changed using FigTree v1.4.0[36] and Adobe Illustrator version 24.3 (Adobe Systems, San Jose, CA, USA).Table 2. GenBank accession numbers of the Fusarium incarnatum-equiseti species complex (FIESC) used in phylogenetic analysis.
Taxon Strain GenBank assession number tef1-α rpb2 cal Fusarium aberrans CBS 131385T MN170445 MN170378 MN170311 F. aberrans CBS 131387 MN170446 MN170379 MN170312 F. arcuatisporum LC12147 MK289584 MK289739 MK289697 F. brevicaudatum NRRL 43638 GQ505665 GQ505843 GQ505576 F. bryceae BRIP 74865c PP209369 PP209368 — F. caatingaense URM 6779 LS398466 LS398495 — F. camptoceras CBS 193.65 MN170450 MN170383 MN170316 F. cateniforme CBS 150.25T = ATCC 11853 MN170451 MN170384 MN170317 F. caulendophyticum GUCC 191050.1 = CGMCC3.25474T OR043881 OR043826 OR043731 F. caulendophyticum GUCC 191050.2 OR043882 OR043827 OR043732 F. caulicola GUCC 191051.1 OR043883 OR043828 OR043733 F. citri CGMCC 3.19467T MK289617 MK289771 MK289668 F. citrullicola SDBR-CMU422 OP020920 OP020928 OP020924 F. clavus CBS 394.93 = BBA 64265 = NRRL 25795 GQ505597 GQ505775 GQ505509 F. clavus CBS 126202T MN170456 MN170389 MN170322 F. clavus CBS 130395 = NRRL 34032 = UTHSC 98- 2172 GQ505635 GQ505813 GQ505547 F. clavus CBS 119881 = MRC 8412 MN170457 MN170390 MN170323 F. clavus NRRL 32871 = FRC R-9561 GQ505619 GQ505797 GQ505531 F. coffeatum CBS 635.76T = BBA 62053 = NRRL 208 MN120755 MN120736 MN120696 F. coffeatum CBS 430.81 = NRRL 28577 MN120756 MN120737 MN120697 F. compactum NRRL 36323 GQ505648 GQ505826 GQ505560 F. concolor NRRL 13459 GQ505674 GQ505852 — F. croceum CBS 131777 MN170463 MN170396 MN170329 F. curculicola PPRI 20458T = PREM 61345 MF787266 MN605062 — F. duofalcatisporum CBS 384.92T = NRRL 36448 GQ505652 GQ505830 GQ505564 F. duofalcatisporum CBS 264.50 = NRRL 36401 GQ505651 GQ505829 GQ505563 F. equiseti CBS 245.61 = NRRL 20697 — JX171595 — F. equiseti NRRL 26419 = CBS 307.94 = BBA 68556T GQ505599 GQ505777 GQ505511 F. equiseti NRRL 36136 = CBS107.07, IMI 091982 GQ505644 GQ505822 GQ505556 F. equiseti NRRL 36466 = CBS 414.86 GQ505653 GQ505831 GQ505565 F. extenuatum LLC1501 OP487158 OP486728 OP486039 F. fasciculatum CBS 131382T MN170473 MN170406 MN170339 F. fasciculatum CBS 131383 MN170474 MN170407 MN170340 F. fasciculatum CBS 131384 MN170475 MN170408 MN170341 F. fecundum LC15875 OQ125250 OQ125544 OQ125281 F. flagelliforme NRRL 36269 GQ505645 GQ505823 GQ505557 F. flavens SDBR-CMU869 = KUNCC 25-20161T PX380329 PX380327 PX380330 F. goeppertmayerae BRIP 64547d OQ626868 — — F. gracilipes NRRL 43635T GQ505662 GQ505840 GQ505573 F. guilinense CBS 161.25 MN170448 MN170381 MN170314 F. guilinense MFLUCC 24-0626 PV394837 PV394827 PV297811 F. hainanense LC11638 MK289581 MK289735 MK289657 F. hainanense MFLUCC 24-0630 PV394834 PV394828 PV297812 F. heslopiae BRIP 74746a PP209371 PP209370 — F. humuli CGMCC 3.19374 = CQ1039T MK289570 MK289724 MK289712 F. humuli CQ1048 MK289571 MK289725 MK289713 F. incarnatum CBS 132.73 MN170476 MN170409 MN170342 F. ipomoeae CGMCC 3.19496 = LC12165 = M0111T MK289599 MK289752 MK289704 F. ipomoeae LC12166 MK289600 MK289753 MK289706 F. irregulare LC7188 MK289629 MK289783 MK289680 F. jinanense LC15878 OQ125131 OQ125521 OQ125271 F. khuzestanicum IRAN 4863C PP858510 PP858506 PP858502 F. khuzestanicum SCUA-Af-108–2 PP858512 PP858508 PP858504 F. kotabaruense Indo172 LS479445 LS479859 LS479429 F. kotabaruense MFLUCC 24-0633 PV394836 PV394831 PV297813 F. lacertarum NRRL 20423 = ATCC 42771 = CBS 130185T = IMI 300797 GQ505593 JX171581 GQ505505 F. lacertarum NRRL 36123 = CBS102300, BBA 70843 GQ505643 GQ505821 GQ505555 F. longicaudatum CBS 123.73T = ATCC 24370 = IMI 160825 = NRRL 25477 MN170481 MN170414 MN170347 F. longifundum NRRL 36372 GQ505649 GQ505827 GQ505561 F. luffae LC12167 MK289601 MK289754 MK289698 F. makinsoniae BRIP 64547a OQ626867 — — F. melonis SDBR-CMU424 OP020922 OP020930 OP020926 F. mianyangense LC15879 OQ125232 OQ125510 OQ125335 F. mianyangense MFLUCC 24-0625 PV394834 PV394829 PV297814 F. monophialidicum NRRL 54973 MN170483 KC808362 MN170349 F. mucidum CBS 102395 MN170485 MN170418 MN170351 F. multiceps NRRL 43639 GQ505666 GQ505844 GQ505577 F. nanum CGMCC 3.19498 = LC12168 = GXGL14-2T MK289602 MK289755 MK289651 F. nanum LC1384 MK289611 MK289764 MK289661 F. nanum LC1385 MK289612 MK289765 MK289662 F. nanum LC1516 MK289613 MK289766 MK289663 F. neoscirpi NRRL 26922 GQ505601 GQ505779 GQ505513 F. neosemitectum CBS 189.60 MN170489 MN170422 MN170355 F. nothincarnatum LC18436 OQ125147 OQ125509 OQ125290 F. oryzicola IRAN 4864C PP858509 PP858505 PP858501 F. oryzicola SCUA-Af-125–2 PP858511 PP858507 PP858503 F. pernambucanum MUM 1862T = URM 7559 LS398489 LS398519 — F. pernambucanum URM 6810 LS398485 LS398515 — F. pernambucanum MUM 1864 = URM 7554 LS398486 LS398516 — F. pernambucanum SDBR-CMU744 PX233235 — PX233226 F. pernambucanum SDBR-CMU745 PX233236 — PX233227 F. persicinum CBS 479.83 MN170495 MN170428 MN170361 F. radicigenum GUCC 197371.1 OR043907 OR043851 OR043752 F. radicigenum GUCC 197425.1 OR043908 OR043852 OR043753 F. radicigenum GUCC 197221.1 = CGMCC3.25478T OR043909 — OR043754 F. rhinolophicola KUMCC 21–0449 OR026001 OR025917 OR022061 F. ruthhalliae BRIP 72406h OP627085 OP627084 — F. scirpi NRRL 36478 GQ505654 GQ505832 GQ505566 F. serpentinum CBS 119880 MN170499 MN170432 MN170365 F. sulawesiense Indo138 LS479443 LS479855 LS479422 F. sulaweaiense MFLUCC 24-0629 PV394835 PV394830 PV297816 F. sylviaearleae BRIP 72816b OR269444 OR269438 — F. tanahbum-buense Indo176 LS479448 LS479863 LS479432 F. tangerinum JZG-2022b OP487189 OP486758 OP486067 F. toxicum CBS 406.86 MN170508 MN170441 MN170374 F. weifangense LC18333 OQ125107 OQ125515 OQ125276 F. wereldwij-sianum NL19-94.009 MZ921850 MZ921718 MZ921538 The new taxon is indicated in bold. The ex-type strains are indicated by T. Pathogenicity tests
Fungal inoculum preparation
-
The pathogenicity test was performed using the Fusarium flavens (SDBR-CMU869 = KUNCC 25-20161) for Koch's postulates. The fungal isolate was grown on PDA plates and incubated at 25 ± 2 °C for one week. To prepare for the suspension, 15 mL of sterile distilled water (DW) was added to PDA plates containing F. flavens. The spores were gently dislodged using a sterile rod, the resulting liquid was filtered through four layers of sterile gauze, and collected in a 50 mL tube. The filtrate was then centrifuged at 900 rpm, 4 °C for 10 min. After discarding the supernatant, the pellet was resuspended in 20 mL of sterile distilled water, centrifuged again, and decanted. The final pellet was suspended in 15 mL of sterile distilled water, and the spore concentration was determined using a hemocytometer under the Olympus light microscope (Olympus CH30RF200, Japan), then adjusted to 106 spores·mL−1.
Detached leaf and stem assay procedure
-
Before being tested, asymptomatic plant tissues (leaves and stems) were collected from a C. sativa (hemp) plantation (45-day-old plants), kept in sterile plastic bags, and carried to the laboratory. Leaves and stems were sterilized with a 0.2% sodium hypochlorite (NaOCl) solution (v/v) for 3 min, washed three times with DW, and air-dried. After that, a uniform wound (3 pores, 1 mm in width) was made at the equator of each leaf and stem using an aseptic needle. To fulfil Koch's postulates, inoculations were performed using a mycelial plug and a spore suspension. A 5-mm-diameter plug of F. flavens was taken from the actively growing edge of the fungal colonies and placed onto the surface of each healthy plant tissue. Sterile PDAC (potato dextrose agar supplemented with chloramphenicol) plugs served as negative controls. The inoculated tissues were positioned on a sterile plastic net inside sterile 90 mm petri dishes, which contained moistened sterile paper towels soaked with 1 mL of DW to maintain humidity. The 10 µL of spore suspension was evenly dropped into healthy plant tissues[14]. Three replications of each treatment were performed. The samples were incubated at 25 ± 2 °C for 3–7 d until disease symptoms appeared. After the appearance of symptoms, the plates were examined for fungal growth and symptom progression.
Detached seedlings assay procedure
-
To perform a pathogenicity test on hemp seedlings, spore suspensions were prepared from cultures of F. flavens as previously described. Hemp seeds were surface-sterilized with 2.5% NaOCl solution for 5 min, washed three times with DW, and then germinated on moist filter paper in sterile plastic boxes for 3–5 d at 25 ± 2 °C. The seedlings were transferred into pots (7 cm high × 8 cm wide) containing a sterilized mixture of vermiculite : perlite : peat moss (1:1:2) for 2 weeks under greenhouse conditions. Fifteen two-week-old hemp seedlings were gently washed with DW to remove debris and then soaked in the spore suspensions, while the control seedlings were soaked in DW for 30 min. After inoculation, each seedling was transplanted into a pot (14.5 cm high × 14 cm wide) containing 800 g of sterilized soil[14]. The plants were cultivated at ambient temperatures ranging from 25 to 34 °C and substrate moisture content within a range of 58% to 81% for 30 d. The pot experiment was conducted in a greenhouse located at the Faculty of Science, Chiang Mai University, Thailand. Throughout the experiment, plants were monitored for wilt symptoms, such as slow growth and yellowing. The level of disease was assessed based on plant growth rates and categorized into four classes: mild (1%–25%), moderate (26%–50%), severe (51%–75%), and very severe (76%–100%). Disease incidence (DI) was calculated as the percentage of diseased plants using the formula: DI (%) = (Number of diseased plants/Total number of plants) × 100. Each isolate was tested in triplicate to ensure consistency and reliability.
To fulfill Koch's postulate, fungi associated with disease symptoms were re-isolated from infected detached tissues and seedlings. The re-isolated fungi were identified based on morphological characteristics and DNA sequence analysis, thus fulfilling Koch's postulates.
-
The BLAST results revealed that our isolate had the highest similarity to members of the Fusarium incarnatum-equiseti species complex. Therefore, this complex was selected for detailed phylogenetic analysis. The combined sequence dataset of tef1-α, rpb2, and cal comprised 96 Fusarium strains in F. incarnatum-equiseti species complex (FIESC), and one outgroup taxon, F. concolor (NRRL 13459) (Fig. 1). The aligned dataset consisted of a total 2,954 characters with gaps (tef1-α: 1–623 bp, rpb2: 624–2,356 bp, cal: 2,357–2,954 bp). The best-scoring maximum likelihood (RAxML) tree was selected to represent the relationships among taxa, with a final likelihood value of –15,909.427308. The matrix contained 963 distinct alignment patterns, with 24.92% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.260395, C = 0.258686, G = 0.244464, and T = 0.236456; substitution rates were AC = 1.412250, AG = 4.053456, AT = 1.124038, CG = 0.659120, CT = 8.715739, and GT = 1.000000; gamma distribution shape parameter α = 0.305373, Tree Length = 1.140524. For Bayesian inference (BI) analysis, the best-fitting model of each locus and the final average standard deviation of split frequencies were determined as follows: the average standard deviation of split frequencies was 0.009952, while the best-fit substitution models were GTR + I + G for tef1-α, GTR + I + G for rpb2, and SYM + G for cal. Similar tree topologies were acquired from ML and BI analyses. In the phylogenetic tree (Fig. 1), the new species, F. flavens (SDBR-CMU869 = KUNCC 25-20161), formed an independent branch basal to F. caulendophyticum (CGMCC 3.25474 and GUCC 191050), and F. weifangense LC 18333 with 72% ML and 1.00 BYPP support (Fig. 1).

Figure 1.
Phylogram constructed by maximum likelihood (ML) analyses of the combined tef1-α, rpb2, and cal sequence dataset of Fusarium. Bootstrap support valued for ML and Bayesian posterior probabilities (BYPP) equal to or greater than 70%, and 0.90, are shown above the nodes as ML/BYPP. The tree is rooted to F. concolor (NRRL 13459). The newly generated strain is indicated by black bold. T indicates the ex-type strains.
Taxonomy
-
Fusarium flavens. Ei, Phookamsak, Monkai & S. Lumyong, sp. nov. (Fig. 2)
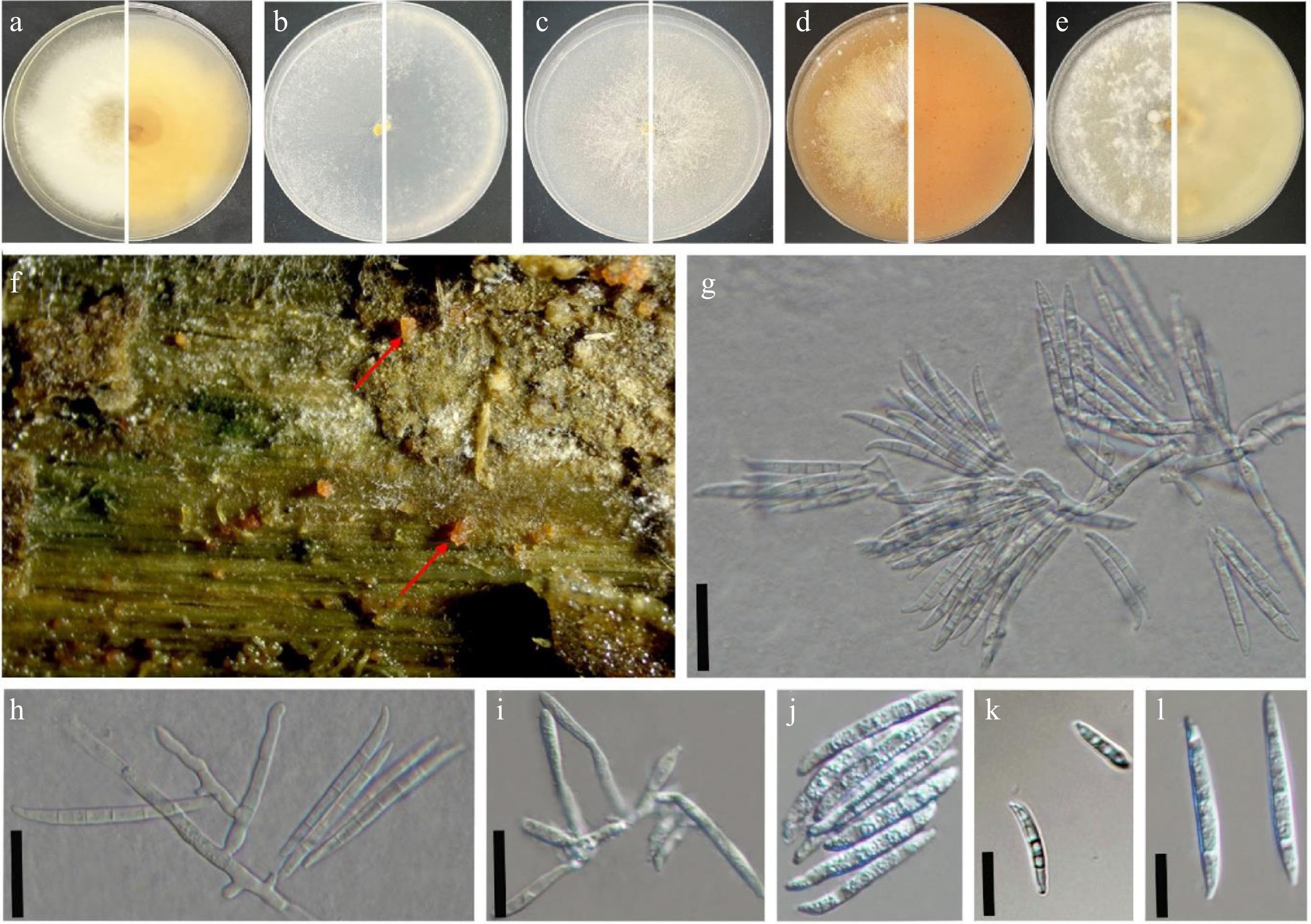
Figure 2.
Fusarium flavens (SDBR-CMU869 = KUNCC 25-20161, ex-type). (a)–(e) Seven-day-old culture on PDA, SNA, YEA, V8 juice agar, and OA incubated at 30 °C (top and reverse). (f) Natural symptoms of stem disease on Cannabis sativa (red arrow indicates the symptoms). (g), (h) Sporodochial conidiophores. (i) Phialides. (j)–(l) Macroconidia found on PDA media incubated at 30 °C. Scale bars: (g)–(i) = 100 μm; (k), (l) = 10 μm.
Index Fungorum number: IF 904383.
Etymology—name refers to the yellowish colour of cultures on the PDA and V8 juice agar.
Holotype—(CMUB 40081) isolated from Cannabis sativa (hemp) stems, Chiang Mai, Thailand.
Pathogenic fungus occurring on the stems of Cannabis sativa (hemp) in Northern Thailand. Sexual morph: not observed. Asexual morph: Sporodochia not observed. Conidiophores macronematous and developed on the aerial mycelium. Aerial phialides subulate to subcylindrical, smooth- and thin-walled, 6–33 × 2–5 µm (
$\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Fusarium flavens (SDBR-CMU869 = KUNCC 25-20161) grew in darkness at 30 °C after 7 d on PDA, SNA, YEA, V8, and OA. Colonies on PDA reached 80 mm in diam. after 7 d of incubation, with abundant mycelial growth, the colony surface turned to creamy white, reverse pale yellow. Colonies on SNA reached 82 mm in diam. after 7 d of incubation, colonies are membranous to woolly, white, with abundant sporulation on the surface, giving a powdery appearance; reverse colourless. Colonies on YEA reached 82 mm in diam. after 7 d of incubation, white, with sparse to moderate aerial mycelium, slightly raised center, and the same reverse colour. Colonies on V8 juice agar medium reached 80 mm in diam. after 7 d of incubation, pale orange with floccose mycelium, reverse saffron yellow. Colonies on OA reached 83 mm in diam. after 7 d of incubation, white to orangish-white, dense and woolly, resembling a cotton-like mass, reverse creamy white colour (Fig. 2).
Material examined—Thailand, Chiang Mai Province, Mae Rim District, Mae Sa Noi Village, unhealthy stems of hemp (Cannabis sativa, Cannabaceae), 18°51'50.06" N, 98°50'45.57" E; elevation 1,285 m/4,216 ft, 12 December 2023, Toe Swe Zin Ei, PS4 (CMUB 40081, holotype), ex-type, SDBR-CMU869 = KUNCC 25-20161.
Notes: According to nucleotide BLAST search, the tef1-α, rpb2, and cal sequences of the new isolate Fusarium flavens (SDBR-CMU869 = KUNCC 25-20161) showed the highest similarity to Fusarium sequences available in GenBank. Details of the closest BLAST matches and percentage identities are presented in Table 3. Our phylogeny depicts a close association of F. flavens with F. humuli, as well as F. weifangense and F. caulendophyticum (Fig. 1). The nucleotide comparison among our new isolate (SDBR-CMU869 = KUNCC 25-20161), and the ex-type strains of F. caulendophyticum, F. weifangense, and F. humuli indicated reasonable nucleotide differences to justify the establishment of a novel species as recommended by Jeewon & Hyde[37] (Table 4).
Table 3. Nucleotide BLAST similarity results among several similar strains of Fusarium flavens.
Genes Percentage similarity across similar species tef1-α F. citri
NRRL 52765 (98.24%)
JF740839Fusarium sp.
FS5 (98.10%)
JQ244848Fusarium sp.
CMW55738 (97.94%)
MZ966178rpb2 F. humuli
hnxryc2 (98.82%)
OR257587F. humuli
JXRC 11 (98.71%)
MZ824673F. humuli
GL 17-3 (98.71%)
OQ512810cal F. humuli
LC18763 (99.27%)
OQ125277F. humuli
LC18553 (99.27%)
OQ125278F. humuli
CMML20-13 (98.99%)
OL331025Table 4. Nucleotide differences among closely related strains of Fusarium flavens.
Compared strains Gene region/locus tef1-α rpb2 cal F. humuli
(CQ 1039, ex-type)2.8%
(18/629 bp)
MK2895701.1%
(10/874 bp)
MK2897240.5%
(3/531 bp)
MK289712F. weifangense
(LC18333, ex-type)3.4%
(19/551 bp)
OQ1251072.1%
(15/751 bp)
OQ1255151.4%
(8/548 bp)
OQ125276F. caulendophyticum
(CGMCC3.25474, ex-type)2.9%
(10/650 bp)
OR0438812.0%
(19/932 bp)
OR0438261.6%
(10/595 bp)
OR043731Fusarium caulendophyticum, described from healthy stems of Rosa roxburghii in Guizhou Province, China[38], possesses irregularly branched conidiophores with lateral and terminal mono or polyphialides. In contrast, F. flavens develops simpler, subulate to subcylindrical aerial phialides, with larger conidiogenous cells (6–33 × 2–5 vs 10–24 × 2–3 μm) and larger macroconidia (21–47 × 2–5 μm vs 16–27 × 2.5–4 μm),[38] compared with those in F. caulendophyticum.
Fusarium weifangense, isolated from symptomatic tissues of Triticum aestivum in Shandong Province, China[39], forms densely branched sporodochial conidiophores with apical whorls of 3 to 4 phialides. In contrast, F. flavens produces macronematous aerial conidiophores with subulate to subcylindrical phialides and smaller conidia, compared with those in F. weifangense (21–47 × 2–5 µm vs 26.5–49.4 × 4.1–7.1 µm)[39].
Fusarium humuli, described from leaves of Humulus scandens in Jiangsu Province, China[22], produces pale orange sporodochia with densely packed, verticillately branched conidiophores bearing small subulate to subcylindrical phialides (6.3–11.9 × 2–3.4 µm). In contrast, F. flavens forms only aerial conidiophores with larger phialides (6–33 × 2–5 µm) and larger conidia (21–47 × 2–5 μm vs 21–35 × 2–3 µm)[22]. Based on these morphological differences and phylogenetic evidence, F. flavens (SDBR-CMU869 = KUNCC 25-20161) is introduced here as a novel species.
Pathogenicity test on detached tissues
-
The pathogenicity test of F. flavens (SDBR-CMU869 = KUNCC 25-20161) on hemp was carried out using a detached leaves and stems assay (Fig. 3a–h). After inoculation with the fungal mycelium plug, the symptoms of brown lesions developed rapidly and spread onto the leaves and stem surface after 3 d of incubation (Fig. 3b, d). In contrast, control leaves and stems receiving treatment with sterile PDAC plugs remained healthy and symptom-free, as shown in (Fig. 3a, c). In the spore suspension inoculation method, brown lesions and wilting developed and spread across the surface of the leaves and stems (Fig. 3f, h). Conversely, the control leaves and stems receiving DW remained healthy and showed no disease symptoms (Fig. 3e, g).

Figure 3.
Pathogenicity tests on detached hemp leaves, stems, and seedlings. (a) Control leaves, and (c) stems, inoculated with a PDAC plug. (b), (d) Symptoms after being inoculated with Fusarium flavens (SDBR-CMU869 = KUNCC 25-20161) mycelium plug for 7 d. (e) Control leaves, and (g) stems, inoculated with distilled water (DW). (f), (h) Symptoms after being inoculated with F. flavens (SDBR-CMU869 = KUNCC 25-20161) spore suspension for 7 d. Inoculation of hemp seedling control with (i) DW, compared with (j) that inoculated with F. flavens (SDBR-CMU869 = KUNCC 25-20161) spore suspension. (k) The roots of a plant inoculated with DW (control) shows no symptoms. (l) The roots of a plant inoculated with F. flavens (SDBR-CMU869 = KUNCC 25-20161) spore suspension shows root rot symptoms (30 d).
Pathogenicity test on detached seedlings
-
In the hemp seedling test, Fusarium flavens was also evaluated for its ability to cause disease (Fig. 3i–l). Four weeks after inoculation, seedlings inoculated with F. flavens exhibited slower growth compared to uninoculated controls. Slow development of symptoms was observed in approximately 73% of the seedling inoculated with F. flavens, respectively. The average plant height of control and inoculated plants is presented in Table 5. The remaining plants showed reduced and stunted development (Fig. 3i, j [right], and l), while the control, non-inoculated, saline-watered seedlings continued to grow healthily (Fig. 3i, j [left] and k). The results of isolated pathogenic strains of F. flavens caused varying symptoms in infected leaves, stems, and seedlings. The fungi were re-isolated from infected plant parts for Koch's postulate assays, and were confirmed based on their morphological characteristics that this fungus was the disease-causative agent.
Table 5. Comparison of plant height between control and inoculated plants.
Treatment Number of plants (n) Plant height
($\overline{\rm x} $ ± SD) (cm)Disease incidence (%) Control plant 15 83.82 ± 5.67 no symptoms Inoculated plant 15 56.13 ± 9.03 73% Disease severity was evaluated based on plant growth performance. Growth reduction (%) relative to the control was calculated and reported in the Results section. -
The identification of Fusarium species, particularly within the Fusarium incarnatum–equiseti species complex (FIESC), requires a combined taxonomic approach integrating both morphological and molecular phylogenetic analyses. Morphological traits, such as colony growth, spore type, size, and septation are important but often insufficient due to overlapping characteristics and cryptic variation among closely related species[22]. Ribosomal DNA markers (ITS and LSU) are widely used, but generally lack the resolution to distinguish among cryptic taxa[40−43]. Therefore, protein-coding genes, including tef1-α, rpb2, and cal, are regarded as more informative and are increasingly adopted as standard markers for species delimitation in Fusarium[38,44].
In practice, accurate identification involves combining morphological examination with multigene phylogenetic analysis, which provides robust species boundaries and resolves hidden diversity. This approach is particularly important in FIESC, which contains numerous cryptic species and newly described taxa[38,44]. In the present study, Fusarium isolates were recovered from hemp stem rot in Northern Thailand. Based on both morphology and multigene phylogeny (tef1-α, rpb2, and cal), one isolate was determined to represent a novel species, F. flavens (SDBR-CMU869 = KUNCC 25-20161), in accordance with established taxonomic practices[38,44]. This study aimed to isolate and identify fungal pathogens associated with C. sativa, and to confirm their pathogenicity by fulfilling Koch's postulates using detached plant tissues and attached seedlings. The causal agent of stem rot, F. flavens, was found on hemp stems. Several Fusarium species are significant pathogens of C. sativa in North America. For instance, F. falciforme has been identified as the causal agent of root rot in the United States[45]. In Israel, soil-borne species such as F. oxysporum, F. proliferatum, and F. solani have been associated with disease development in cannabis[14]. Additionally, F. proliferatum has been documented as the cause of stem rot in Canada[46]. During our taxonomic investigation of pathogenic fungi, several Fusarium species were identified based on morphological characteristics, nucleotide pairwise comparisons, and phylogenetic evidence. One of them, F. flavens, a novel species, is reported here for the first time on C. sativa. The novel species, F. flavens was found to be phylogenetically related to F. caulendophyticum and F. weifangense (Fig. 1). However, the phylogenetic placement of F. flavens received relatively moderate bootstrap support (Fig. 1), indicating the need for further investigation. However, the independent lineage and the nucleotide differences support the establishment of a new species. In this study, pathogenicity was confirmed using both detached tissues and seedling tests, demonstrating that the F. flavens isolate obtained from hemp stems can also cause both stem and root rot symptoms. In pathogenicity tests, brown lesions and wilting developed in detached tissues, while seedling inoculations resulted in root rot symptoms. The severity varied among test conditions, indicating that the pathogenic potential of this isolate may be influenced by environmental factors.
In Thailand, Fusarium species have been documented to affect a variety of crops. For example, F. proliferatum, F. thapsinum, and F. verticillioides have been found on sorghum[47]. Additionally, F. oxysporum has been reported in banana farms in Chiang Rai, causing significant crop losses and posing a serious threat to banana production[48]. Moreover, two species of Fusarium, F. citrullicola and F. melonis, have been identified from rot lesions on watermelon and muskmelon fruits[49]. Recent studies have also discovered three novel causal agents of muskmelon fruit rot: F. compactum, F. jinanense, and F. mianya[50]. A recent study by Samarakoon et al.[51] described F. pernambucanum as a causal agent of bud rot of hemp in northern Thailand. There is a need to do further research on this genus in Thailand, given its potential to cause diseases in many economically important crops.
-
In conclusion, these findings highlight the presence and potential impact of Fusarium species in C. sativa cultivation in Chiang Mai Province, Northern Thailand. The confirmation of their pathogenicity provides a valuable basis for future research for effective disease management, and the development of resistant cultivars. Overall, this study expands our understanding of the fungal diversity and pathogenic threats affecting C. sativa in Thailand and underscores the need for ongoing monitoring and integrated management strategies to reduce potential economic losses in hemp production.
-
The authors confirm contributions to the paper as follows: study conception and design: Ei TSZ, Phookamsak R, Monkai J, Jatuwong K, Aiduang W, Lumyong S; data collection: Ei TSZ, Phookamsak R; Monkai J, Jatuwong K, Aiduang W; analysis and interpretation of results: Ei TSZ, Phookamsak R; Monkai J, Jatuwong K, Aiduang, Hongsanan S, Khuna S; draft manuscript preparation Ei TSZ, Phookamsak R, Jeewon R, Monkai J, Jatuwong K, Aiduang, Hongsanan S, Khuna S, Lumyong S. All authors reviewed the results and approved the final version of the manuscript.
-
All sequences generated in this study were submitted to GenBank.
-
This study was supported by the Faculty of Science, Chiang Mai University for providing tuition and fees scholarship. The authors extend their gratitude to the Biology Experimental Center, Germplasm Bank of Wild Species, Kunming Institute of Botany, and the Chinese Academy of Sciences for providing the molecular laboratory facilities. Dr. Shaun Pennycook is thanked for the nomenclatural review. Toe Swe Zin Ei sincerely acknowledges the Kunming Institute of Botany, University of Chinese Academy of Science (UCAS) partical scholarship, (Grant No. E33M681261). Prof. Saisamorn Lumyong acknowledges the 'Thailand and National Research Council of Thailand (N42A650198)' and this research was partly supported by the International Research Fellowship (Visiting Professor Program), Chiang Mai University (Grant No. 031/2568). Rungtiwa Phookamsak sincerely acknowledges the Talent Introduction Plan of Kunming Institute of Botany, Chinese Academy of Sciences, Yunnan Revitalization Talent Support Program 'Young Talent' Project (Grant No. YNWR-QNBJ-2020-120), Yunnan Revitalization Talent Support Program: High-end Foreign Expert Project (Grant No. XDYC-GDWZ-2024-0016), Independent Research of Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. Y537731261), Yunnan Provincial Department of Human Resources and Social Security, Yunnan Province Foreign Expert Project (Grant No. 202505AO120002) and Yunnan Intelligence Union Program for Young Scientists, Yunnan, China (Grant No. 202503AM140005). The University of Mauritius and CAS President's International Fellowship Initiative (PIFI) for visiting scientist (Grant No. 2026PVB0047) are also acknowledged. The New Drug Discovery and Evaluation Center for international cooperation and disciplinary innovation ('111 Center') at Zunyi Medical University and College of Science, King Saud University (Saudi Arabia) are also acknowledged.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ei TSZ, Phookamsak R, Monkai J, Jatuwong K, Aiduang W, et al. 2026. Morpho-phylogenetics and pathogenicity tests reveal a new species of Fusarium infecting Cannabis sativa L. (hemp) in Northern Thailand. Studies in Fungi 11: e011 doi: 10.48130/sif-0026-0011 

Morpho-phylogenetics and pathogenicity tests reveal a new species of Fusarium infecting Cannabis sativa L. (hemp) in Northern Thailand
- Received: 03 November 2025
- Revised: 16 February 2026
- Accepted: 05 March 2026
- Published online: 17 April 2026
Abstract: Cannabis sativa L., widely known as hemp or cannabis, has been cultivated for centuries and also legalized in several countries around the world. In Thailand, permits to use cannabis only for medical and licensed purposes are allowed, and hemp (tetrahydrocannabinol [THC] < 0.2%), is legal for general use. The rapid expansion of hemp cultivation has increased the crop's susceptibility to fungal diseases. Species of the genus Fusarium are major phytopathogens, although they may also exist as endophytes. During a survey of pathogenic fungi in Northern Thailand, diseased stems of C. sativa were collected, from which a novel Fusarium species, F. flavens sp. nov., was isolated. Identification was performed using both morphological features and analyses of DNA sequence data. Phylogenetic trees were constructed based on analyses of combined dataset of the translation elongation factor 1-α (tef1-α), RNA polymerase II second largest subunit (rpb2), and calmodulin (cal) gene regions. A combined taxonomic approach of morphological comparisons and multi-locus phylogenetic analyses confirmed the distinctiveness of the species. Pathogenicity was assessed by inoculating spore suspensions and mycelium plugs on hemp leaves, stems, and seedlings. Typical symptoms were reproduced, including leaf and stem wilting and root rot. Re-isolation and re-identification of the same fungus confirmed Koch's postulates. Based on this comprehensive approach, F. flavens was identified as the causal agent of stem and root rot in hemp. This study highlights the significance of F. flavens as a novel pathogen of hemp and emphasizes the importance of integrating morphology, molecular phylogenetics, and pathogenicity tests to clarify its role in hemp diseases.
-
Key words:
- Nectriaceae /
- New species /
- Pathogenicity test /
- Stem and root rot pathogen /
- Taxonomy