






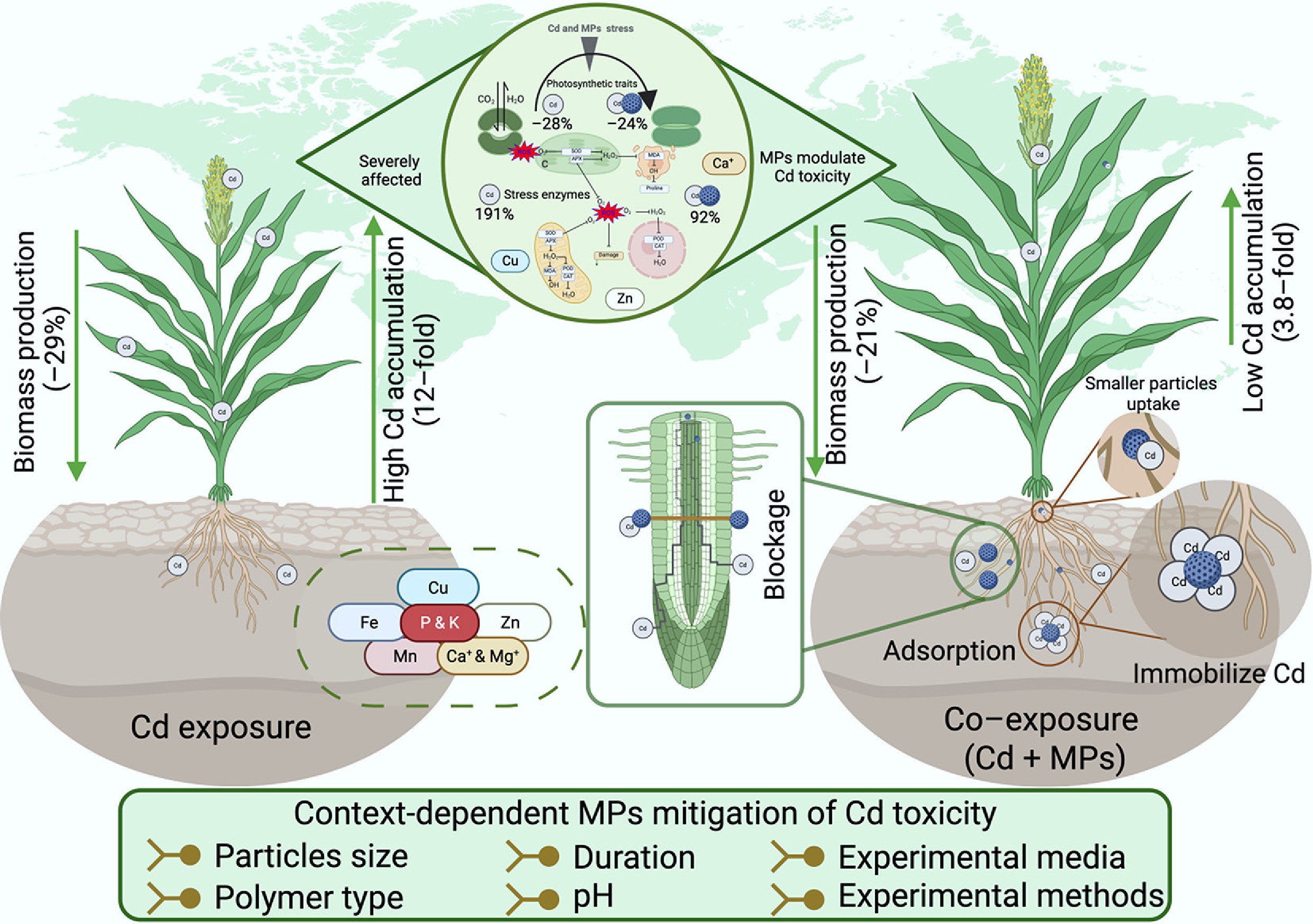



-
Cadmium (Cd) is one of the most hazardous heavy metals, posing persistent environmental and food safety concerns due to its long-term stability, high bioaccumulation potential, and toxicity[1]. Even at low concentrations, Cd readily accumulates in soils and plants[2], where it disrupts growth, induces oxidative stress, and threatens human health. It has no known biological function in plants and engages in complex interactions within soil–plant systems[3]. In China, a national soil pollution survey reported that Cd concentrations exceeded safety thresholds in approximately 7% of farmland soils[4], largely due to natural geochemical background and anthropogenic activities such as mining, metal smelting, and wastewater irrigation[5]. These findings highlight the urgent need to mitigate Cd accumulation in agricultural soils to ensure sustainable land use and crop production.
Microplastics (MPs) are now recognized as ubiquitous contaminants in terrestrial ecosystems, including agricultural lands[6]. Due to their high specific surface area and diverse functional groups, MPs can bind metal pollutants, influencing their transport and fate in soils[7,8]. In farmlands where MPs and Cd often co-occur[9,10], MPs can significantly alter Cd bioavailability, mobility, uptake, and toxicity in plants. Several studies have reported that MPs' adsorption of Cd reduces its mobility and toxicity[10−13]. For example, smaller MP particles have been shown to downregulate Cd transporter proteins such as IRT1 (LOC_Os03g46470) and NRAMP4 (LOC_Os01g31870) in rice[14], and therefore limit the Cd bioaccumulation[15]. MPs exposure may also promote lignin or suberin deposition in root endodermal cells, strengthening apoplastic barriers and restricting Cd movement from roots to shoots[16−18]. Additionally, MPs adhering to root surfaces can block adsorption sites, reducing direct Cd contact with root tissues[10,19]. In some cases, co-exposure to polystyrene (PS) MPs and Cd decreased Cd accumulation and alleviated toxicity in other plants[11,12].
However, the capacity of MPs to reduce Cd toxicity is highly context-dependent, influenced by particle size, polymer type, dose, plant species, and environmental conditions. For example, co-exposure to polyethylene (PE) and Cd has been shown to enhance Cd mobility, increase plant accumulation, and amplify toxicity[20]. In particular, smaller particles, such as nanoplastics (NPs), have higher surface area-to-volume ratios, enabling stronger sorption of Cd ions and potential penetration into root tissues[21]. Depending on binding affinity and transport behavior, this can either mitigate or enhance toxicity. MP dose also plays a nonlinear role: low PS concentrations (0.5% w/w) increased Cd uptake in roots, whereas higher concentrations (2% w/w) reduced Cd content in both roots and shoots[11,22]. Polymer type is equally critical: PE increased Cd accumulation in maize shoots[23], whereas biodegradable polylactic acid (PLA) reduced shoot Cd uptake under the same conditions. PS has been reported to enhance photosynthesis and antioxidant defenses under Cd stress in pakchoi[11], whereas PE exacerbated toxicity in lettuce[20]. These contrasting outcomes are largely driven by differences in polymer composition, particle morphology, and surface reactivity[24].
Given these inconsistencies, synthesizing existing evidence is essential for a comprehensive assessment of MPs–Cd interactions in agroecosystems. We hypothesize that MPs can modulate Cd toxicity, as an antagonistic effect in plants, and that the magnitude of this modulation is governed by MP characteristics (e.g., particle size and polymer type) as well as soil environmental factors such as soil pH. This study presents a global-scale meta-analysis of 4,243 pairwise observations across diverse plant species to quantify the effects of Cd alone and in combination with MPs on plant morphology, photosynthetic performance, biochemical activity, and Cd accumulation. We also identify key MPs-related characteristics and soil environmental factors that govern the magnitude and direction of Cd toxicity modulation. Our findings provide robust evidence for understanding MPs–Cd interactions and for guiding risk assessment and management strategies in co-contaminated agricultural systems.
-
Data for this study were extracted from peer-reviewed articles published before February 2025. Relevant publications were identified through systematic searches of Web of Science (
https://apps.webofknowledge.com ), Scopus (www.scopus.com ), Google Scholar (https://scholar.google.com ), and ResearchGate. Search terms included combinations of keywords related to 'plastic', 'microplastic' or 'nanoplastic, 'cadmium', 'plant' or 'crop', 'morphological or physiological response', 'biochemical indicators', 'accumulation', 'types', and 'soil' or 'hydroponic'.Following the Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines, a rigorous search, screening, and selection process was conducted to compile the most relevant and reliable datasets (Supplementary Fig. S1). Studies were included if they met all of the following criteria: (1) Reported at least one of the following response variables: plant morphological traits (root length, shoot length, root biomass, shoot biomass), photosynthetic traits (total chlorophyll, chlorophyll a, chlorophyll b, photosynthetic rate), antioxidant and oxidative stress indicators (superoxide dismutase [SOD], peroxidase [POD], catalase [CAT], hydrogen peroxide [H2O2], malondialdehyde [MDA]), protein content (PC), and Cd accumulation in roots and shoots. (2) Provided experimental details including MP polymer type, doses of both MPs and Cd, particle size, experimental duration, growth medium, and initial media pH of MPs, Cd, or their combinations. (3) A paired design was employed between treatment (MPs, Cd, or their combinations) and control, while studies with additional confounding factors were excluded. (4) For studies reporting multiple harvest times or different growth media, each dataset was treated as an independent observation if it met all inclusion criteria.
A total of 60 articles met the inclusion criteria, yielding 4,243 paired experimental observations of eight plant species or groups, i.e., Oryza sativa, Triticum aestivum, Zea mays, Lactuca sativa, Brassica, Lolium, Glycine max, and other plants (Supplementary Fig. S1). Due to the limited number of observations, other plant species (Solanum lycopersicum, Sorghum bicolor, Arachis hypogaea, Fragaria ananassa, Ipomoea batatas, Solanum americanum, Lantana camara, Phragmites australis, Arabidopsis thaliana, Amaranthus tricolor, and Taraxacum) were pooled into one group. For each study, the mean value, standard deviation (SD), and sample size for each response variable were extracted for both treatment and control groups. Data were obtained directly from text and tables or digitized from figures and supplementary materials using WebPlotDigitizer (V5;
https://automeris.io/WebPlotDigitizer/ ). A comprehensive dataset was then established, in which plant variables were classified into four major categories and 17 subcategories according to their biological characteristics (Supplementary Fig. S2). The geographic distribution of the selected studies showed that most experiments were conducted in China, which emerged as the leading research region.To assess the MP-mediated modulation of Cd toxicity, this meta-analysis compared the effect sizes between single-exposure (Cd only) and co-exposure (Cd + MPs) groups. To further identify how MP characteristics drive Cd toxicity, MPs were categorized into two size groups (> 100 μm and ≤ 100 μm; Supplementary Fig. S2). MP concentrations reported in different units (e.g., g kg−1, mg kg−1, mg L−1, g L−1, w/w) were standardized to mg kg−1, and particle sizes to micrometers (μm). By polymer type, MPs were categorized as PS, PE, PLA, polyvinyl chloride (PVC), polypropylene (PP), and other polymers. Exposure duration was grouped into three timeframes (< 15 d, 15–30 d, and > 30 d; Supplementary Fig. S2). Initial media pH was classified as acidic, neutral, or alkaline. Growth media were categorized as soil, hydroponic, foliar, or petri dish, and experimental conditions as incubation or greenhouse.
Meta-analysis
-
The effect size (lnRR) was calculated as the natural log response ratio using the openMEE software, following the approach of Hedges et al.[25] and Gurevitch et al.[26]:
$ \mathrm{ln}RR=\mathrm{ln}\left(\dfrac{{X}_{\mathrm{t}}}{{X}_{\mathrm{C}}}\right)=\mathrm{ln}\left({X}_{\mathrm{t}}\right)-\mathrm{ln}\left({X}_{\mathrm{c}}\right) $ (1) where, Xt and Xc represent the mean values of the treatments and control groups, respectively. A positive lnRR (lnRR > 0) indicated a stimulatory effect, whereas a negative lnRR (lnRR < 0) indicates an inhibitory effect. The corresponding error variance (v) of lnRR was calculated as:
$ v=\dfrac{S_{\mathrm{c}}^{2}}{{n}_{\mathrm{c}}X_{\mathrm{c}}^{2}}+\dfrac{S_{\mathrm{t}}^{2}}{{n}_{\mathrm{t}}X_{\mathrm{t}}^{2}} $ (2) where, St and Sc represent the SD of the treatments and control group, respectively; nt and nc represent the corresponding sample sizes. If the standard error (SE) was reported instead of SD, it was converted using:
$ \mathrm{SD}=\mathrm{SE}\times\sqrt{\mathrm{Number}~\mathrm{of}~\text{replicates}} $ (3) The weighting factor (w) and mean effect size were calculated as:
$ w=\dfrac{1}{v} $ (4) $ \mathrm{ln}RR_{\mathrm{mean}}=\dfrac{\sum_{ }^{ }(\mathrm{ln}RR_i\times w_i)}{\sum_{ }^{ }w_i} $ (5) where, wi is the weight, and lnRRi is the effect size from the ith comparison.
The response ratio (RR) for each variable was further expressed as a percentage change induced by MPs and Cd exposure, using the following equation:
$ {\text{%}} ~\mathrm{change}=\left({\mathrm{e}}^{RR}-1\right)\times 100 $ (6) Heterogeneity among studies was quantified using Cochran's Q statistic, I2 index, and τ2 (between–study variance). Cochran's Q was calculated as:
$ Q=\sum{w}_{i}{\left({\ln RR}_{i}-{\ln RR}_{\text{mean}}\right)}^{2} $ (7) where, wi are the fixed-effect weights.
The significance of heterogeneity was assessed using a chi-square test:
$ P=1-CHISQ\_ CDF(Q,n-1) $ (8) where, n is the number of effect sizes.
The proportion of total variability attributable to real heterogeneity rather than sampling error was expressed using Higgins' I2 statistic:
$ {I}^{2}=\max \left\{0, \dfrac{Q-(n-1)}{Q}\right\}\times 100$ (9) Values of I2 = 25%, 50%, and 75% were interpreted as low, moderate, and high heterogeneity, respectively.
This transformation facilitates straightforward interpretation of the magnitude and direction of MPs, Cd, and their combination effects on plants.
All statistical analyses were performed using the OpenMEE software with the 'meta'[27] and 'metafor'[28] packages in R (version 4.3.1). RF model was performed using packages in R (version 4.3.1) with the 'ranger' package. Data visualization was conducted using the ggplot2 package in R and GraphPad Prism (Version 10.1.10). BioRender (
www.biorender.com ) was used to create the graphical abstract and Fig. 7. The overall pooled effect was estimated using a first-order meta-analysis with 95% confidence intervals (CI). -
Exposure to MPs, Cd, or their combination, on average, generally exerted negative effects on plant species compared with CK, with a few exceptions such as Glycine max and Lolium perenne. Notably, Oryza sativa, Zea mays, Brassica, and Triticum aestivum exhibited pronounced reductions in root length, root biomass, shoot biomass, and photosynthetic traits, with declines ranging from 10% to 60% relative to CK (Fig. 1). In addition, Cd accumulation in both root and shoot tissues increased significantly, by more than threefold across most species, with Glycine max being the only exception.
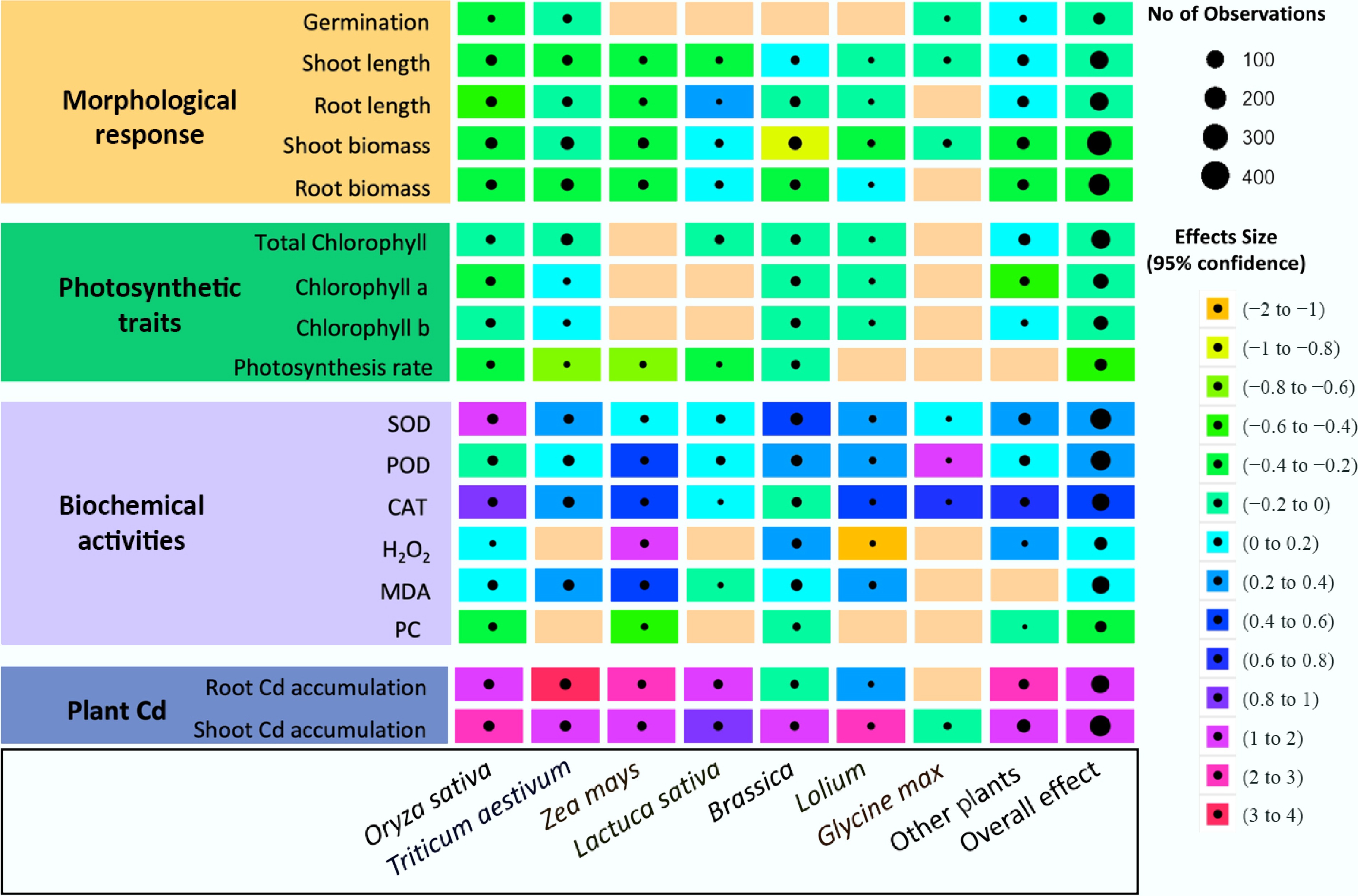
Figure 1.
The overall effects of Cd, MPs, and their combined exposures on plant responses relative to control in terms of morphological parameters, photosynthetic traits, biochemical activities, and plant Cd accumulation across different species. Rows represent specific traits, while columns correspond to plant species, family, and the overall effect. Effect sizes (± 95% CI) are indicated by a color gradient, ranging from strongly inhibitory (orange, –2 to –1) to strongly promotive (magenta, 3 to 4). Circle size denotes the number of contributing observations, and blank cells indicate the absence of reported data.
When the three treatments (MPs, Cd, and co-exposure) were evaluated separately (Supplementary Figs S3–S5), Cd exposure alone caused the most severe effects, although some species, such as Glycine max, were less affected. Oryza sativa, Zea mays, and Triticum aestivum were particularly sensitive, with root and shoot lengths and biomass reduced by 23%–54%. Photosynthetic performance was also strongly impaired, with Oryza sativa and Zea mays showing declines of 27%–57%. Cd accumulation increased markedly, with root concentrations rising 10–56-fold (excluding Triticum aestivum and Brassica) and shoot concentrations increasing 4–33-fold (Supplementary Fig. S3). Additionally, MPs exposure reduced root length, root biomass, and shoot biomass of Oryza sativa, Zea mays, and Brassica by 6%–32% (Supplementary Fig. S4).
Under co-exposure conditions, however, the negative effects on Oryza sativa, Zea mays, and Triticum aestivum were less pronounced compared with Cd alone. Reductions in root length, shoot length, root biomass, and shoot biomass ranged from 12% to 34%. Photosynthetic traits were also less impaired, with Oryza sativa and Zea mays declining only by 21%–40%. Likewise, Cd accumulation under co-exposure was not substantially higher than under Cd alone, with root concentrations increasing by 1–27-fold (excluding Triticum aestivum and Brassica) and shoot concentrations by 1–6-fold across all species (Supplementary Fig. S5). These results suggest that MPs mitigated Cd toxicity under co-exposure, indicating an antagonistic interaction between MPs and Cd.
Since different species generally responded in a similar manner to MPs, Cd, and their combination, we subsequently averaged the dataset across all eight species to emphasize treatment-level comparisons between Cd alone and co-exposure (Cd + MPs). This approach facilitated a more detailed assessment of the antagonistic effects of MPs on Cd toxicity, while also helping to elucidate the underlying mechanisms responsible for these interactions.
Plant morphological traits
-
Exposure to MPs or Cd alone significantly reduced plant morphological traits. Cd alone decreased shoot length by 24%, root biomass by 42%, and shoot biomass by 47%, while MPs alone caused smaller reductions of 3%, 13%, and 24%, respectively. Under co-exposure, MPs alleviated Cd toxicity to some extent, with reductions in shoot length, root biomass, and shoot biomass limited to 14%, 26%, and 39%, respectively (Fig. 2). Overall, Cd exposure caused the largest reduction in morphological traits (–0.342; 95% CI: –0.3840 to –0.2999). MPs alone produced a smaller decline (–0.1108; 95% CI: –0.1379 to –0.0836), whereas co-exposure resulted in an intermediate reduction (–0.2122; 95% CI: –0.2464 to –0.1779) (Supplementary Fig. S6a). These patterns indicate an antagonistic effect between MPs and Cd for all three morphological traits.
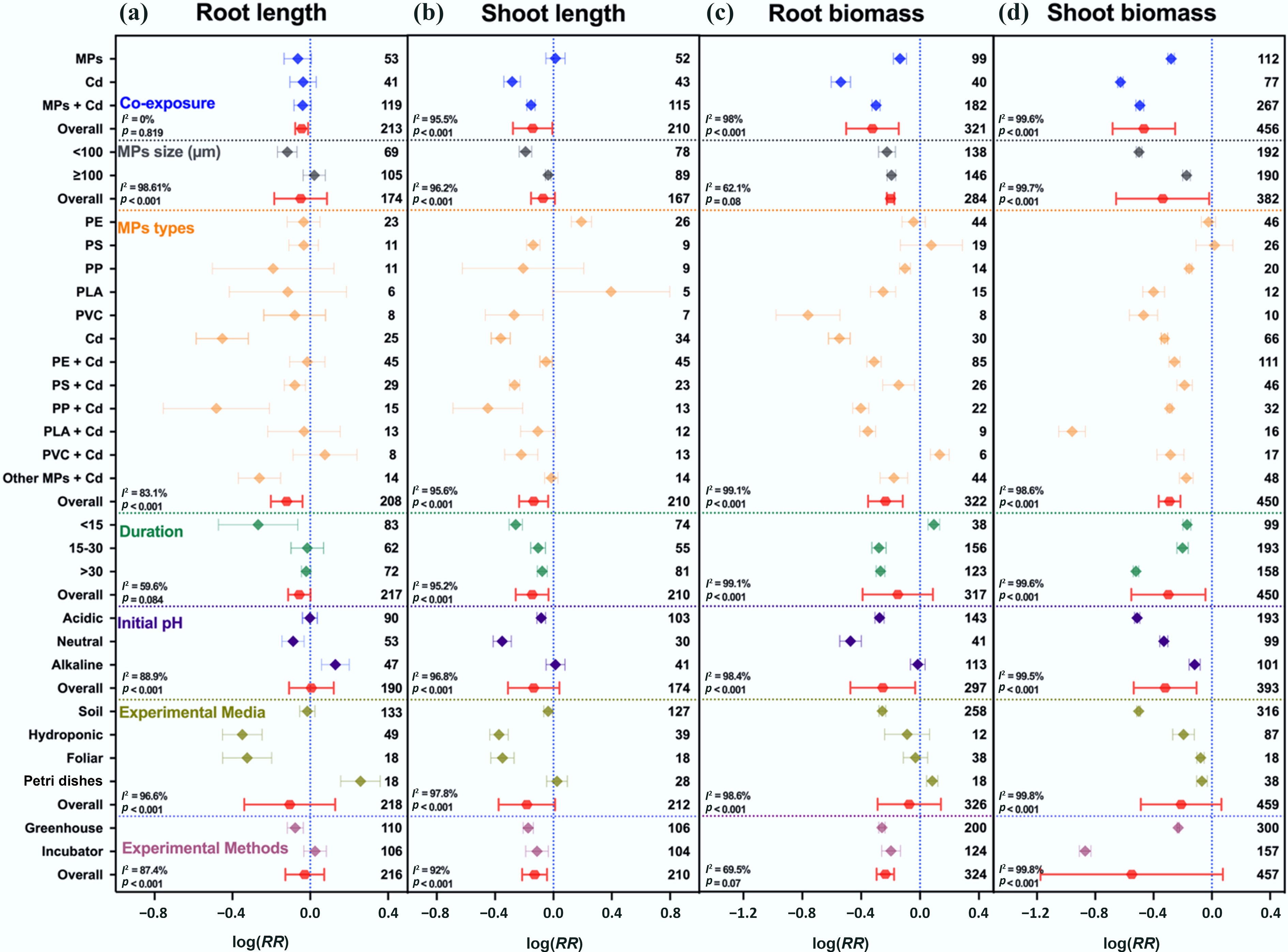
Figure 2.
Interactive effects of MPs, Cd, and co-exposures on plant morphological traits. (a) Root length, (b) shoot length, (c) root biomass, and (d) shoot biomass. Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot. Response ratios are arranged by exposure type, MP particle size, polymer types, stress exposure duration, initial media pH, growth media, and experimental methods, which are classified by different colors. In each plot, the number associated with each data point represents the number of pairwise observations. For each plot (a)–(d), color represents different groups of the meta-analytical model effect: blue (single and co-exposure of MPs and Cd), gray (MP size in µm), orange (polymer types), green (duration in days), purple (initial media pH), indigo (growth media), maroon (experimental methods), and red indicates the overall effect across groups.
The magnitude of this antagonistic effect was influenced by several factors. The particle size of MPs played a critical role: smaller particles (< 100 μm) caused more pronounced reductions in root and shoot length and biomass (17%–39%) compared with larger particles (≥ 100 μm), which caused only 5%–18% reductions (Fig. 2). Polymer type also affected responses; PP (average –33%) and PLA (average –26%) co-exposed with Cd showed stronger adverse effects across all measured growth traits. The experimental medium also modulated responses. In hydroponic and foliar systems, root and shoot length decreased by 28%–31%, while biomass losses were smaller (3%–18%). Dose-dependent effects of co-exposure showed that higher MP dose (> 100 mg kg–1) mitigated Cd-induced inhibition of plant length (–2% to –11%) compared to lower MP dose with Cd (≤ 100 mg kg–1; –18% to –24%) (Supplementary Fig. S6b). The Random Forest (RF) model further identified the most important variables for growth parameters. For root and shoot length (Supplementary Fig. S7), experimental media (18% each), and particle size (14% and 13%, respectively) were the strongest predictors. For root biomass, size (20%) was the dominant factor, followed by dose (16%), and pH (14%).
Photosynthetic performance
-
Cd exposure alone caused substantial declines in photosynthetic traits, reducing photosynthetic rate and chlorophyll contents (total, a, and b) by 32%, 12%, 61%, and 12%, respectively. Co-exposure with MPs slightly mitigated Cd effects, with reductions of 24%, 10%, 56%, and 8%, respectively (Fig. 3). Similarly, overall, Cd and MPs reduced photosynthetic traits up to –0.1379 (95% CI: –0.1689 to –0.1069), and –0.3757 (95% CI: –0.4360 to –0.3153), respectively, while co-exposure of both decreased by –0.1377 (95% CI: –0.1719 to –0.1035) (Supplementary Fig. S8a), indicating an overall antagonistic interaction.
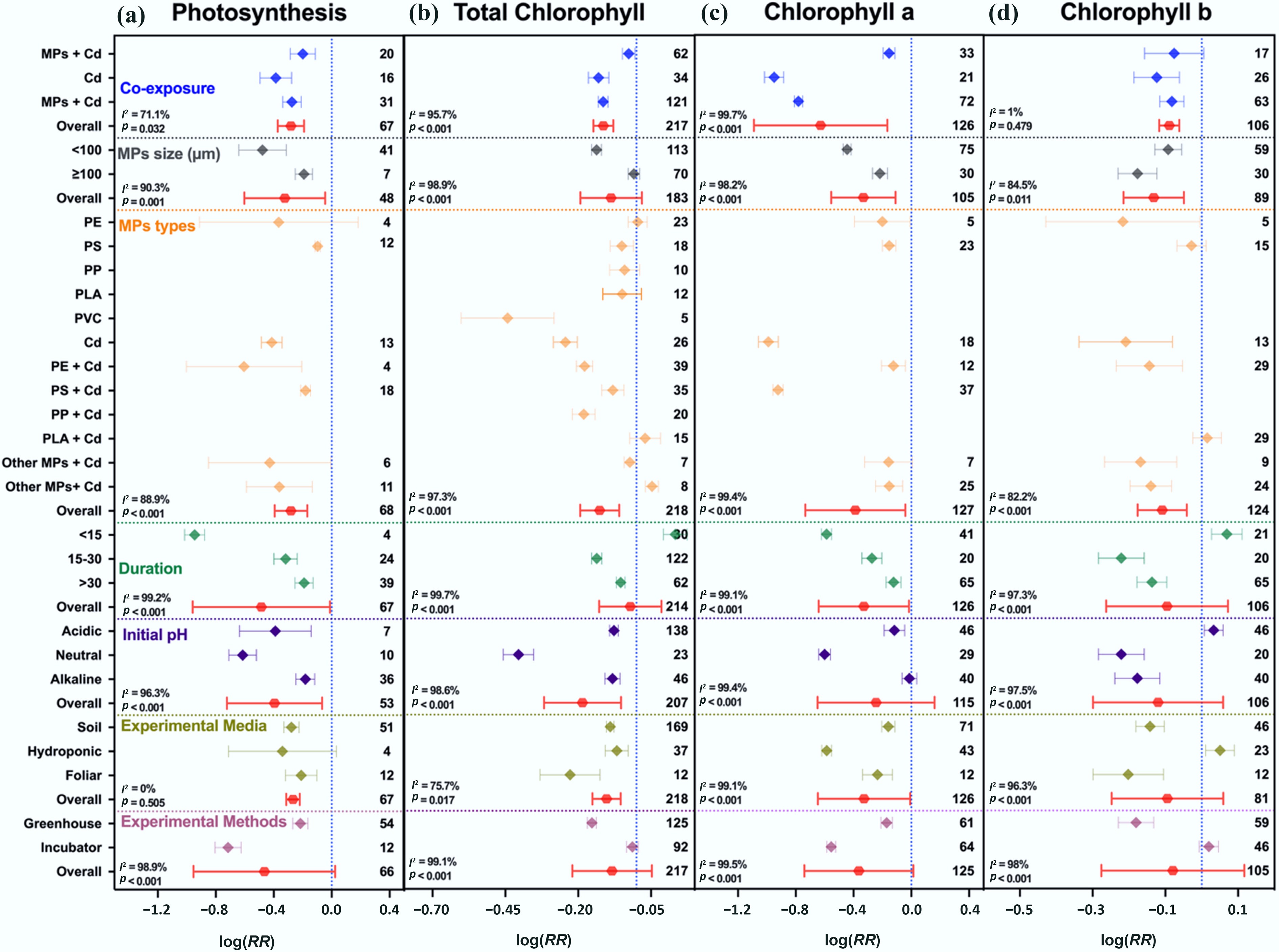
Figure 3.
Responses of plant photosynthetic traits to single and co-exposure of MPs and Cd. (a) Photosynthetic rate, (b) total chlorophyll, (c) chlorophyll a, and (d) chlorophyll b. Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot. Response ratios are arranged by exposure type, MPs particle size, polymer types, duration of stress exposure, initial media pH, growth media, and experimental methods, which are classified by different colors. For each plot (a)–(d), color represents different groups of the meta-analytical model effect: blue (single and co-exposure of MPs and Cd), gray (MP size in µm), orange (polymer types), green (duration in days), purple (initial media pH), indigo (growth media), maroon (experimental methods), and red indicates the overall effect across groups.
The degree of modulation depended on multiple factors. Regarding the polymer type, PVC with Cd alleviated toxicity more effectively, with an average reduction of 17%, whereas PE with Cd caused more severe declines (average 22%). Temporal patterns showed that the greatest damage occurred within the first 15 d of exposure, particularly for photosynthetic rate (–61%) and total chlorophyll content (–14%). Dose-dependent effects were observed, as higher MP dose (≥ 100 mg kg–1) with Cd limited reductions in photosynthetic traits (–10% to –21%), compared to stronger declines under lower MP dose with Cd (< 100 mg kg–1; –13% to –58%) (Supplementary Fig. S8b).
Environmental conditions further modulated the combined effects. Neutral pH conditions caused the strongest declines in photosynthetic rate (–46%), total chlorophyll (–33%), chlorophyll a (–45%), and chlorophyll b (–20%), whereas alkaline conditions mitigated these effects. Exposure medium also influenced outcomes: soil-grown plants showed smaller average declines in photosynthetic traits (–15%) than plants under foliar exposure (–20%), or hydroponic systems (–19%).
Biochemical activity
-
Exposure to either MPs or Cd alone significantly enhanced oxidative stress responses in plants. CAT activity increased by 27% and 52%, MDA by 16% and 30%, and H2O2 by 3.4 and 3.5-fold, respectively. PC declined by 28% under MPs and 49% under Cd. Under co-exposure, MPs modulated Cd-induced oxidative stress, resulting in a moderate increase in CAT (44%), MDA (27%), and H2O2 (1.5-fold), and a smaller reduction in PC (24%) compared to Cd alone (Figs 4 and 5). Overall, oxidative damage was high under Cd (0.5348; 95% CI: 0.4027–0.6669) and MP exposure (0.5463; 95% CI: 0.3998–0.6927), whereas co-exposure yielded less damage (0.2389; 95% CI: 0.1481–0.3297) (Supplementary Figs S9a and S10a).
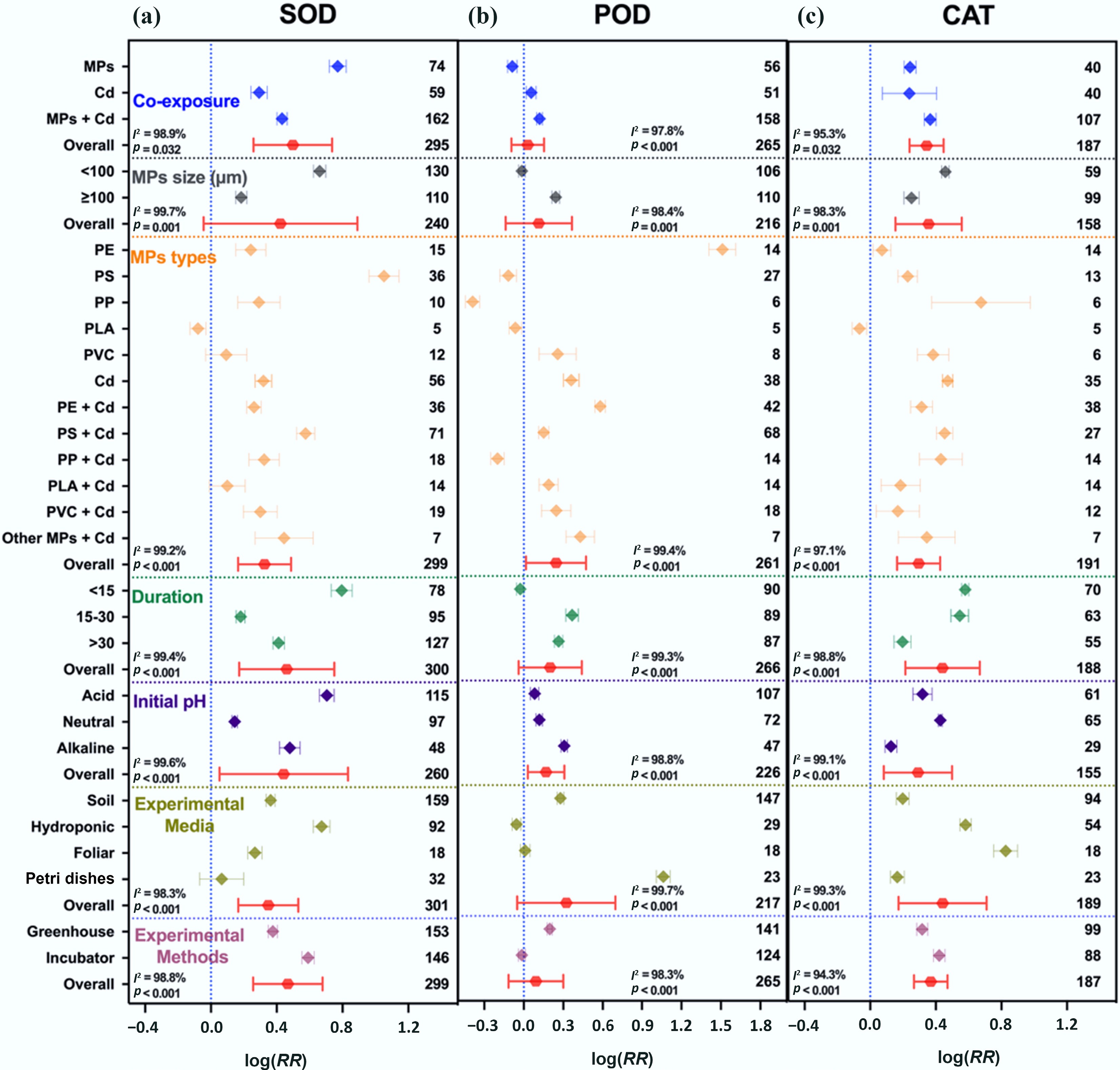
Figure 4.
Single and co-exposure of MPs and Cd modulate the antioxidant enzymatic activities. (a) Superoxide dismutase (SOD), (b) peroxidase (POD), and (c) catalase (CAT). Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot. Response ratios are arranged by exposure type, MP particle size, polymer types, duration of stress exposure, initial media pH, growth media, and experimental methods, which are classified by different colors. For each plot (a)–(c), color represents different groups of the meta-analytical model effect: blue (single and co-exposure of MPs and Cd), gray (MP size in µm), orange (polymer types), green (duration in days), purple (initial media pH), indigo (growth media), maroon (experimental methods), and red indicates the overall effect across groups.
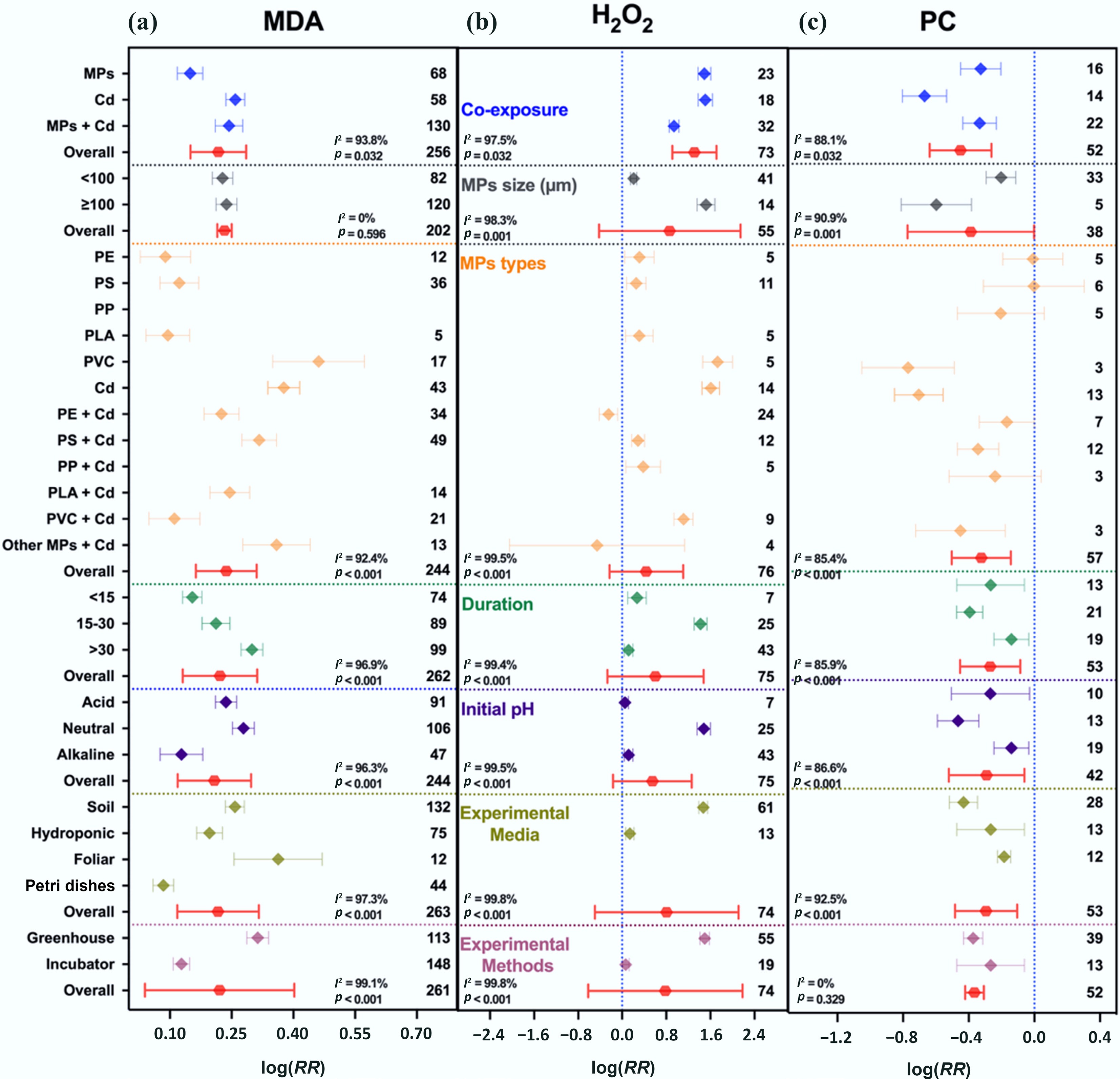
Figure 5.
Responses of oxidative enzymes and protein contents to MPs, Cd, and their co-exposure. (a) Malondialdehyde (MDA), (b) hydrogen peroxide (H2O2), and (c) protein contents (PC). Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot. Response ratios are arranged by exposure type, MP particle size, polymer types, duration of stress exposure, initial media pH, growth media, and experimental methods, which are classified by different colors. For each plot (a)–(c), color represents different groups of the meta-analytical model effect: blue (single and co-exposure of MPs and Cd), gray (MP size in µm), orange (polymer types), green (duration in days), purple (initial media pH), indigo (growth media), maroon (experimental methods), and red indicates the overall effect across groups.
The intensity of biochemical responses varied with MP particle size, exposure duration, growth medium, and pH (Figs 4 and 5). Smaller MPs (< 100 μm) caused stronger activation of antioxidant enzymes, with SOD, CAT, and MDA increasing by 94%, 58%, and 26%, respectively, compared with larger MPs (≥ 100 μm), which induced smaller increases (SOD 20%, CAT 29%, MDA 25%). Exposure duration also influenced oxidative responses; MPs and Cd co-exposure induced oxidative damage of 24% after 15–30 d, with the highest oxidative stress (35% increase) occurring after > 30 d, indicating a time-dependent intensification of damage. In the dose-dependent effect, higher MP dose (≥ 100 mg kg–1) with Cd enhanced SOD and POD activities while reducing superoxide radicals, indicating mitigation of Cd-induced oxidative stress compared to lower MP dose (< 100 mg kg–1) (Supplementary Figs S9b and S10b).
Growth medium influenced the biochemical responses to stresses induced by MPs and Cd. The soil-based system exhibited the highest oxidative damage, with MDA and H2O2 increasing by 29% and 3.3-fold, and antioxidant enzyme activities rising by 21%–43%, alongside a 35% reduction in PC (Figs 4 and 5). In hydroponic systems, antioxidant enzymes (SOD and CAT) showed the largest increases (78%–96%). Neutral pH caused the most serious oxidative damage (MDA +32%, H2O2 +3.4-fold) with increased antioxidant activities (13% to 53%), while acidic and alkaline (5.5% to 27%) conditions attenuated oxidative damage.
Cd uptake, accumulation, and translocation
-
Single Cd exposure caused a 19-fold increase in root Cd and a 5.6-fold increase in shoot Cd. Co-exposure with MPs markedly reduced Cd accumulation, limiting it to 4.5-fold in roots and 1.6-fold in shoots (Fig. 6), indicating that MPs can reduce Cd bioavailability and uptake in plants, accompanied by enhanced photosynthetic traits, and antioxidant and oxidative activities that supported plant growth and development (Fig. 7).
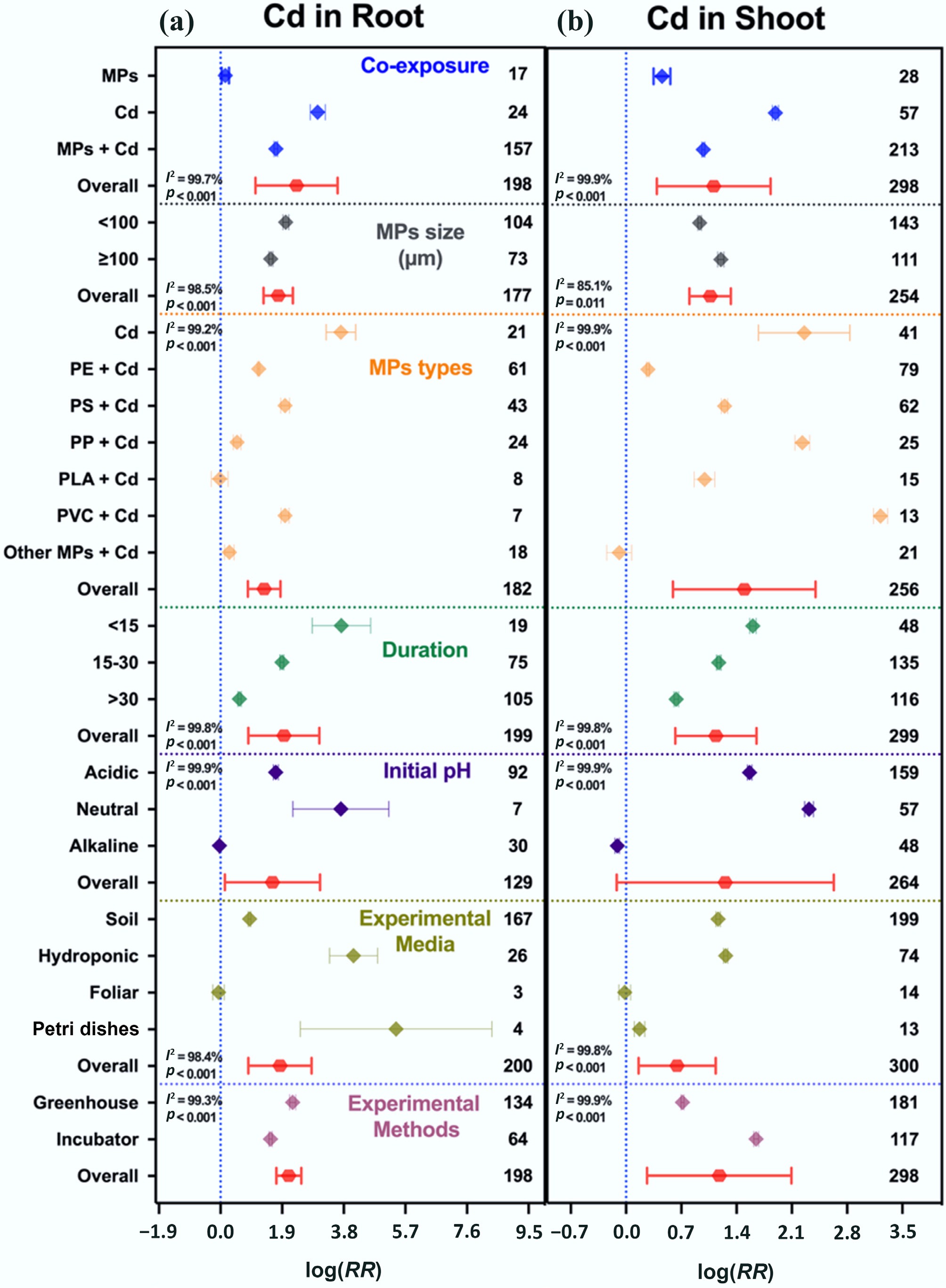
Figure 6.
Cd accumulation in plants under single and co-exposure of MPs and Cd. (a) Cd accumulation in roots, and (b) Cd accumulation in shoots. Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot. Response ratios are arranged by exposure type, MP particle size, polymer types, duration of stress exposure, initial media pH, growth media, and experimental methods, which are classified by different colors. For each plot (a), (b), color represents different groups of the meta-analytical model effect: blue (single and co-exposure of MPs and Cd), gray (MP size in µm), orange (polymer types), green (duration in days), purple (initial media pH), indigo (growth media), maroon (experimental methods), and red indicates the overall effect across groups.
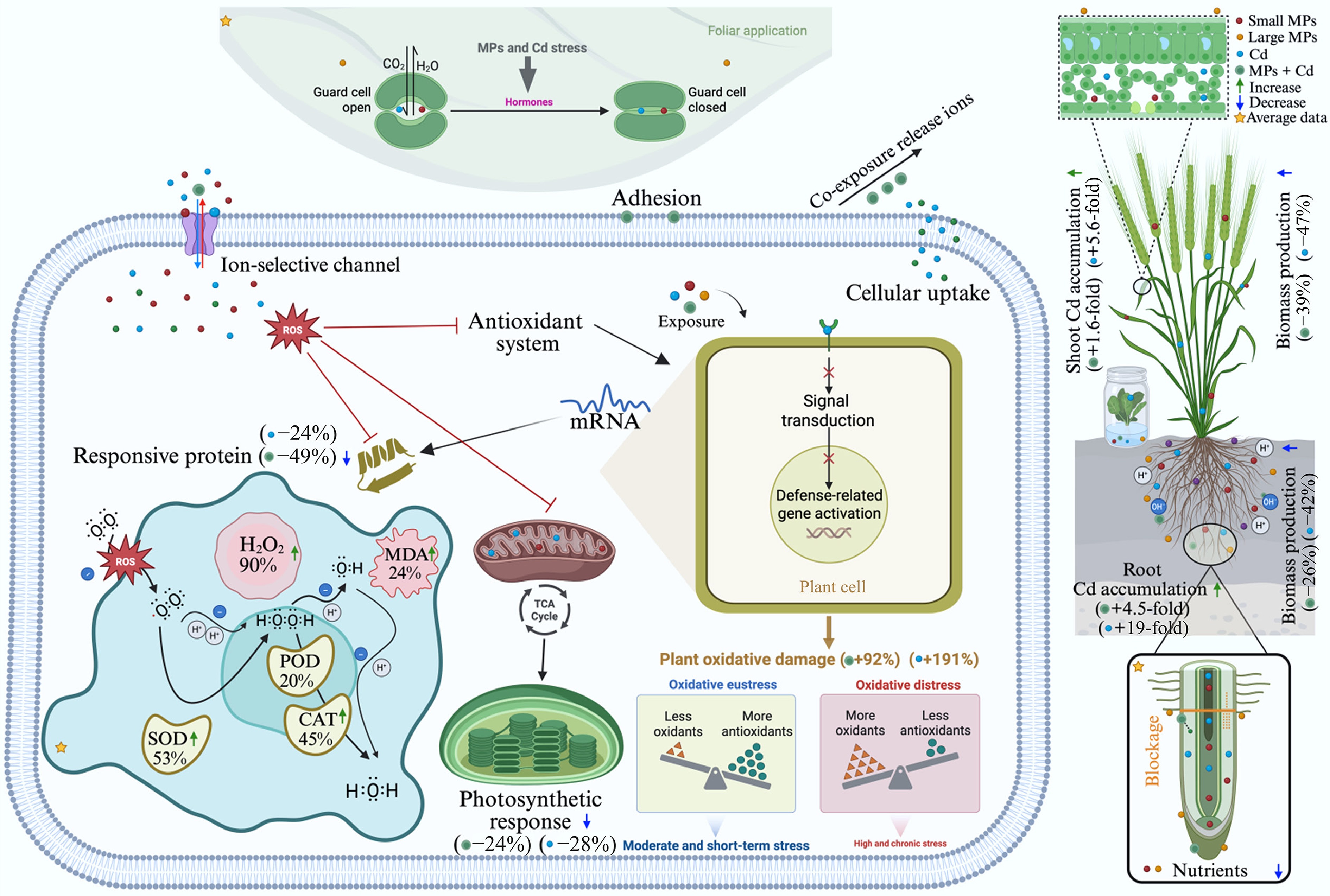
Figure 7.
Mechanisms of MPs and Cd effects on plant growth, photosynthetic performance, Cd uptake and translocation in crop systems, and plant defensive actions against single and co-exposure stresses. Cd can enter plants through both root and above-ground tissues. MPs and soil particles cause physical blockage and adsorption, which may influence plants' uptake and translocation of Cd and nutrients. Plant defense systems mainly involve antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), along with indicators of oxidative damage (malondialdehyde [MDA] and hydrogen peroxide [H2O2]). The value in parentheses represents the response ratio of observed parameters. Positive and negative values indicate the enhanced or decreased effects on plant response.
The particle size of MPs affected Cd distribution between root and shoot tissues. Smaller MPs (< 100 μm) increased root accumulation (6.5-fold) but limited shoot translocation (1.5-fold), whereas larger MPs (≥ 100 μm) reduced root accumulation (3.7-fold) but increased shoot translocation (2.3-fold). Polymer type also influenced the uptake of Cd between root and shoot tissues. PVC caused the highest Cd retention in roots (6.3-fold) and shoots (24-fold), whereas PP facilitated greater shoot translocation (8.3-fold) despite low root retention (0.6-fold). Short-term exposure (<15 d) led to extremely high root accumulation (40-fold) but moderate shoot translocation (3.9-fold), while long-term exposure (≥ 30 d) substantially reduced Cd uptake (0.8-fold in both tissues), suggesting desorption or detoxification over time.
Dose-dependent effect also modulated the Cd accumulation in roots and shoots. Co-exposure of MPs with low Cd dose (< 10 mg kg−1) increased Cd accumulation by 1–4-fold, while at higher Cd dose (≥ 10 mg kg−1) with MPs, the accumulation increased moderately by 3–5-fold compared to Cd alone (8–18-fold). This indicates stronger antagonistic effects of MPs at higher Cd dose, effectively mitigating Cd accumulation in roots and shoots (Supplementary Fig. S11b). Growth media also shaped Cd dynamics: hydroponic systems showed much higher root (59-fold) and shoot Cd accumulation (2.5-fold) than soil systems (1.5- and 2.2-fold, respectively). The RF model also highlighted the most influential moderators (Supplementary Fig. S7). For root Cd accumulation, experimental media (31%) was the most important factor, followed by plant species (18%) and dose (12%). For shoot Cd uptake, plant species (18%) and polymer type (16%) were the primary determinants.
Disruption of nutrient homeostasis
-
Nutrient homeostasis, essential for plant growth and productivity, was significantly disrupted by MP and Cd exposure (Fig. 8). Cd alone reduced iron (Fe), zinc (Zn), manganese (Mn), and potassium (K) by 34%, 13%, 37%, and 10%, respectively, while increasing Cu and Ca by 4.8-fold and 8%. Co-exposure with MPs altered these patterns. For example, Fe, Mn, magnesium (Mg), phosphorus (P), and K declined by 40%, 42%, 13%, 9%, and 6%, respectively, while Cu, Zn, and calcium (Ca) increased by 2.6-fold, 37%, and 44%, respectively. These results indicate that MPs can intensify Cd-induced depletion of some essential nutrients while enhancing the uptake of others.
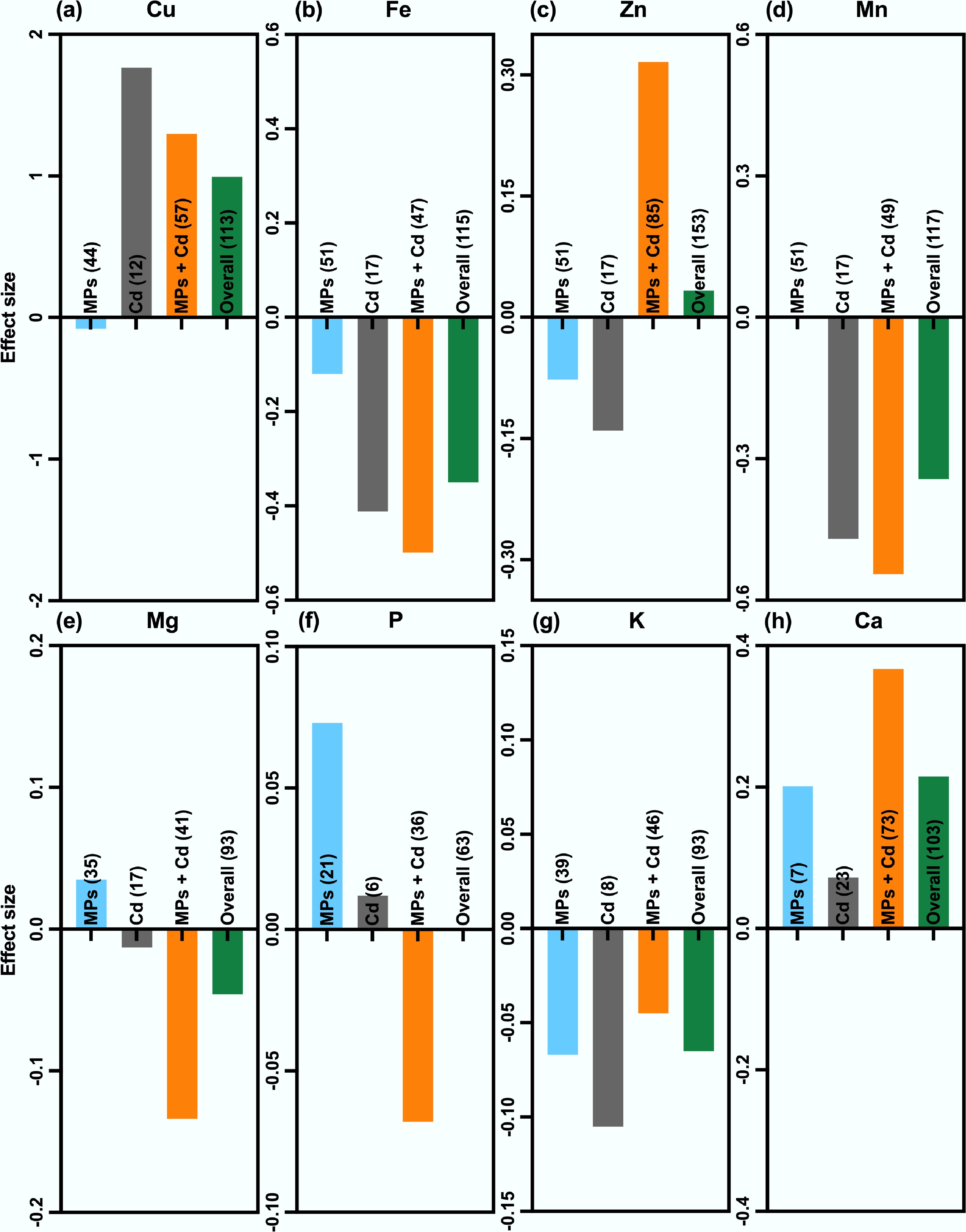
Figure 8.
Meta-analysis of MPs, Cd, and their co-exposure effects on essential plant nutrient uptake. Results are presented for eight mineral nutrients, including: (a) copper (Cu), (b) iron (Fe), (c) zinc (Zn), (d) manganese (Mn), (e) magnesium (Mg), (f) phosphorus (P), (g) potassium (K), and (h) calcium (Ca) under MP exposure (light blue), Cd exposure (gray), and co-exposure of MPs and Cd (orange), along with the overall effect (green). Values represent the mean effect size ± 95% CI. Positive values (above the baseline) indicate enhanced nutrient uptake, while negative (below the baseline) values indicate inhibition.
This nutrient modulation capacity varied with MP size, exposure time, and growth medium. Smaller MP particle size (< 100 µm) caused stronger disruptions in Mg, P, and Mn, while larger MP particle size (≥ 100 µm) enhanced Cu, Zn, and Ca accumulation (Supplementary Fig. S12). Short-term exposure (≤ 15 d) greatly increased Cu but depleted Zn and Mn, whereas longer exposures (> 30 d) improved Fe and Mn, indicating partial recovery of nutrient balance (Supplementary Fig. S13). Greenhouse conditions promoted nutrient accumulation, while incubator systems generally caused nutrient losses (Supplementary Fig. S14).
Microbial diversity alteration
-
Exposure to Cd reduced soil microbial diversity, decreasing Chao1 by 8.42%, Shannon by 0.80%, Simpson by 3.73%, and ACE by 0.80%, indicating loss of species richness and evenness. In contrast, MPs alone had minimal impact, with Chao1 decreasing by 7.69% and Shannon nearly unchanged (0.10%). Notably, co-exposure of MPs and Cd decreased Chao1 by 4.50%, and Shannon increased by 4.60%, suggesting that MPs can partially protect microbial communities against Cd-induced stress. Overall, Cd was toxic to soil microbes, while MPs moderated this effect (Fig. 9).
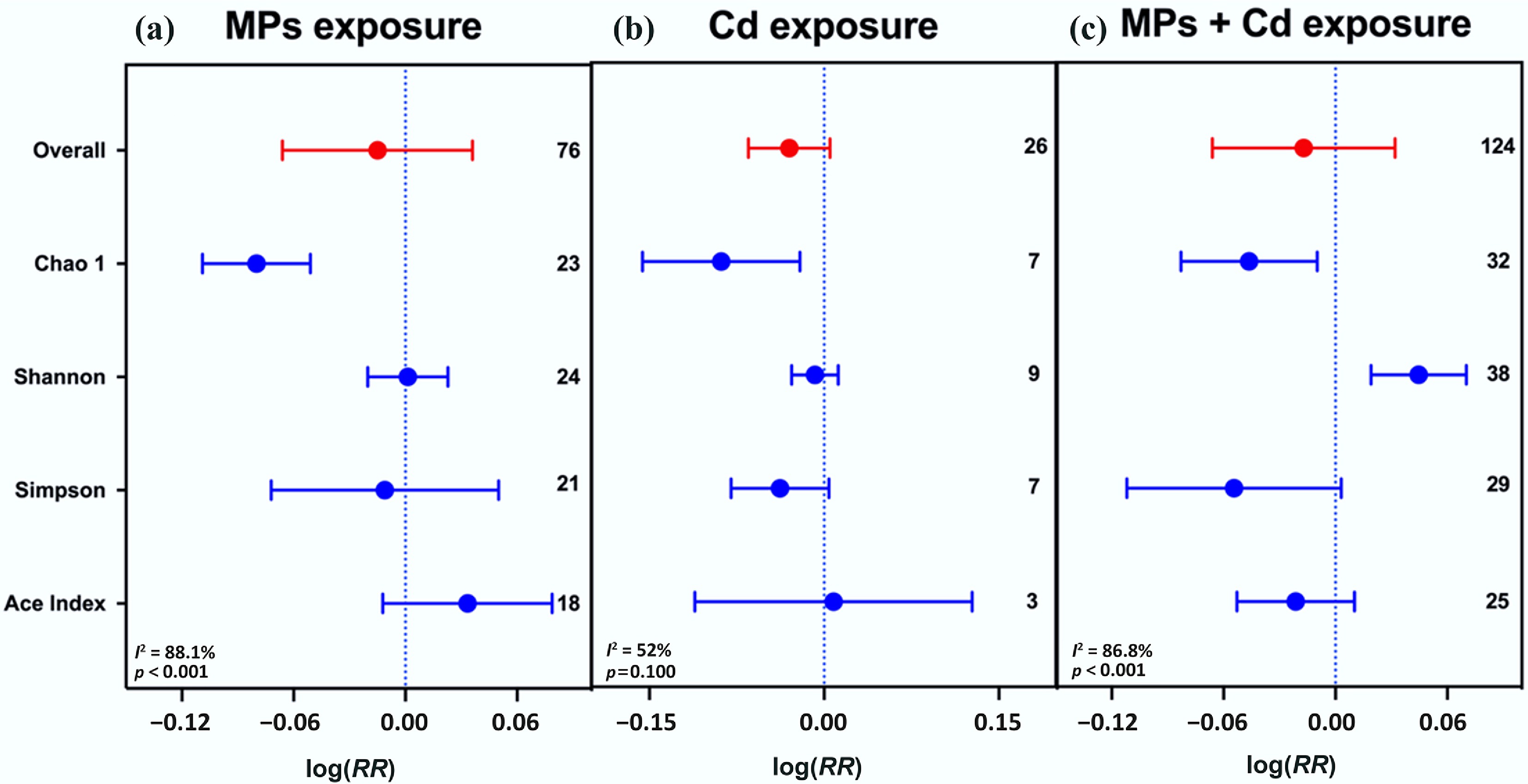
Figure 9.
Single and co-exposure of MPs and Cd affect soil microbial diversity. (a) MP exposure, (b) Cd exposure, and (c) co-exposure of MPs and Cd. Values are the mean effect size ± 95% CI. The right sidebar has a positive effect, the left side has a negative effect, and the bar overlapping zero indicates that the effect is not significant for each plot.
-
Our results confirm that Cd exposure alone exerts severe toxic effects on plant morphology and photosynthetic activity, whereas under co-exposure conditions, MPs partially reduced these negative impacts (Figs 2 and 3). This antagonistic effect is primarily explained by alterations in Cd bioavailability and uptake (Fig. 6). MPs act as sorbents, with their weathered or oxidized surfaces containing reactive functional groups (–COOH, –OH, –C=O) that strongly bind Cd ions through electrostatic attraction, complexation, or ion exchange[10,19,29,30]. These sorption processes immobilize Cd, decreasing the pool of free Cd ions in soil solution and thereby reducing plant uptake. Consistent with our findings, Cd accumulation decreased in roots and shoots under co-exposure of MPs and Cd compared to Cd alone (Fig. 6). PE and PS MPs have been shown to adsorb Cd more efficiently, attenuating metal toxicity[10,19]. A previous study also evidenced the antagonistic effect in biomass responses: Cd alone reduced root and shoot biomass by ~25%, whereas co-exposure with MPs lessened reductions to 10%–21% in roots, and 8%–18% in shoots[11]. In contrast, several studies have reported a synergistic effect between MPs and heavy metals, where co-exposure significantly decreased plant growth and development[5,31]. This variation in plant response can be attributed to factors such as the type and size of MPs, the specific heavy metals involved, the plant species, pH levels, and the experimental media used. Collectively, these results support the conclusion that surface adsorption of Cd by MPs is a critical driver of reduced Cd toxicity[32].
The mechanisms by which MPs adsorb Cd include several processes: physical pore filling, molecular interactions between MPs and heavy metals, complexation via surface functional groups, electrostatic interactions, chemical bonding between bipolar ions, and hydrophobic distribution[33]. Various types of MPs exhibit different Cd adsorption behaviors, with MPs having grooves, pores, and cracks, which promote adsorption in polymeric, complex, and colloidal forms on their surfaces[34]. The surface structure of MPs, affected by factors such as weathering and wear, increases their ability to attract charges and adsorb metal cations[35]. Over time, MP aging can increase the surface oxygen-containing groups, such as C–O, C=O, and –OH, thereby enhancing adsorption capacity[36,37].
A second mechanism involves the physical barrier effect of MPs in the rhizosphere. Larger MP particles generally cannot penetrate root tissues but can adhere to root surfaces, clog pore spaces, and disrupt ion exchange processes[10,19]. The current results showed that MPs reduce Cd bioaccumulation and also restrict the uptake of essential nutrients (Figs 6 and 8). Such dual effects mean that while Cd uptake is lowered, co-exposure can still result in an additional form of abiotic stress caused by nutrient limitation[38]. This highlights the complexity of MPs–Cd interactions, as MPs can shield plants from Cd to some extent, but at the cost of impaired nutrient balance, creating trade-offs in plant physiological responses (Fig. 2).
The role of MPs in mitigating Cd toxicity is further supported by changes in the plant antioxidant defense system. Cd exposure triggered a strong increase in oxidative stress biomarkers (CAT, H2O2, and MDA), reflecting elevated reactive oxygen species (ROS) accumulation and lipid peroxidation, which compromise membrane integrity and disrupt metabolism[39,40]. By contrast, co-exposure with MPs resulted in smaller increases in these indicators (Figs 4 and 5), suggesting alleviated oxidative stress. The antagonistic effect was particularly visible in MDA levels, which were significantly higher under Cd alone compared to co-exposure, demonstrating that MPs reduce ROS-driven membrane damage[13,41]. Plants respond to Cd stress by activating antioxidant enzymes such as SOD, POD, and CAT, which detoxify ROS by sequentially converting superoxide radicals to H2O and O2[42]. In line with previous findings, Cd alone sharply elevated antioxidant activity (SOD, POD, and CAT increased by 58%–382%), whereas co-exposure with MPs caused only moderate increases (41%–231%)[41]. This moderate activation suggests that MPs help stabilize redox homeostasis, preventing excessive oxidative damage and minimizing downstream disruptions to photosynthesis and growth (Fig. 7).
Furthermore, the modulation of nutrient balance under MP–Cd co-exposure suggests that MPs can reduce Cd toxicity by stabilizing nutrient homeostasis. Our results show that the increase in Ca, Zn, and Cu likely plays a protective role, as Ca competes with Cd for transport sites and enhances membrane integrity, while Zn and Cu act as cofactors for antioxidant enzymes, strengthening defenses against Cd-induced oxidative stress (Fig. 8)[43−45]. By offsetting Cd-driven depletion of Fe, Mn, and Mg, MP-mediated nutrient adjustments may buffer physiological disruptions, thereby supporting photosynthetic activity and plant growth under combined exposure. Taken together, these results show that MPs reduce Cd toxicity in plants by reducing Cd bioavailability through adsorption, creating barrier effects in the rhizosphere, and moderating oxidative stress responses and restoring nutrient homeostasis.
Exposure to Cd significantly reduced soil microbial diversity, as evidenced by declines in Chao1 and Simpson indices. This decrease suggests a loss of both species richness and evenness, which is consistent with previous studies showing that Cd toxicity can disrupt microbial community structure by inhibiting microbial growth and reducing alpha diversity[46,47]. Soil microbial communities play a significant role in Cd transformation through processes such as bioaccumulation, degradation, and the modification of Cd bioavailability[48]. Disruption of these communities can exacerbate the environmental risks associated with Cd contamination.
MPs alone had a relatively minor effect on microbial diversity, with only slight reductions in Chao1. Previous studies have reported that PE-MPs have a less pronounced effect on microbial diversity and functional gene abundance[49]. It may be due to MPs providing labile carbon sources that boost microbial activity and increase soil microbial network complexity[50,51]. However, under co-exposure to MPs and Cd, Cd toxicity was alleviated, and the microbial community was less affected than under Cd exposure alone, suggesting that MPs may play a protective role in minimizing the adverse effects of Cd. This finding is supported by previous studies showing that MPs can adsorb Cd, thereby reducing its bioavailability in soil and altering microbial communities[52]. In contrast, other studies have reported that co-exposure to MPs and Cd negatively affects microbial communities[53]. These contrasting results suggest that MPs may modulate metal bioavailability through physical and chemical interactions, influencing metal speciation and either mitigating or enhancing toxicity to microorganisms. However, the effects of co-exposure to MPs and Cd remain uncertain and warrant further investigation to better understand their ecological implications.
Another possible mechanism involves the structure–function relationships within microbial communities, particularly the role of microbial functional groups in mediating Cd toxicity. Cd-resistant bacteria, which possess mechanisms such as efflux pumps and intracellular sequestration, can reduce Cd bioavailability in soils[47,54]. MPs may influence the abundance and activity of these functional groups by providing habitats that favor Cd-resistant microbes or by altering soil physicochemical conditions that enhance their resistance mechanisms. In addition, plant growth-promoting rhizobacteria (PGPR), which enhance plant tolerance to Cd through processes such as phosphate solubilization and phytohormone production[55,56], may also be affected by MPs. MPs could either promote PGPR proliferation by offering refuge from environmental stress or disrupt their beneficial interactions with plant roots.
Drivers of MP characteristics on Cd toxicity
-
The ability of MPs to modulate Cd toxicity is highly context-dependent and varies with particle size, polymer type, exposure duration, soil environmental pH, and growth medium (Supplementary Fig. S7). A critical evaluation of these factors highlight the complexity of plant responses and provide mechanistic insights into the observed variability.
Smaller MPs (< 100 µm) intensified Cd toxicity by aggravating morphological impairments, photosynthetic inhibition, and biochemical disturbances compared to larger MPs (≥ 100 µm) (Figs 3−5). The greater surface area of smaller MPs enhances Cd adsorption, while their nanoscale fractions can penetrate root tissues, traverse apoplastic or symplastic pathways, and translocate via the xylem to aerial tissues[57,58]. Once inside, they disrupt nutrient balance, photosynthesis, and cellular redox status[59]. In contrast, larger MPs are rarely taken up by root tissue; instead, they adhere to root surfaces, limiting Cd bioavailability, and thus reducing overall toxicity[6,38]. The current study confirms a size-dependent effect of MPs, i.e, smaller MPs enhance heavy metal bioaccumulation and oxidative stress in plants, whereas larger-sized plastics primarily influence external interactions. This size-dependent behavior may partly explain their differential impacts on plant growth and development (Fig. 2).
Experimental pH strongly influenced Cd mobility and plant responses. Neutral to acidic conditions exacerbated Cd toxicity by enhancing solubility, uptake, and oxidative damage (Figs 3, 5−7), and under these conditions, MPs can further intensify toxicity by maintaining Cd in soluble forms or facilitating its transport to root surfaces. In contrast, alkaline conditions alleviated toxicity by promoting Cd precipitation into insoluble forms[60,61]. Acidic environments also reduce soil surface charge, thereby increasing Cd binding to root exchange sites and facilitating uptake[62,63]. These findings underscore the need to evaluate pH-dependent interactions in contaminated soils, as remediation strategies (e.g., liming) could significantly alter Cd-MPs dynamics.
Time-dependent responses indicated that short-term exposure (< 15 d) induced substantial Cd accumulation in roots (~40-fold), accompanied by limited translocation to shoots (~3.9-fold). However, with prolonged exposure (> 30 d), Cd accumulation declined markedly (~0.8-fold) (Fig. 6). This suggests that plants progressively activate defense mechanisms (Fig. 4), including synthesis of metal-chelating peptides (phytochelatins, metallothioneins), enhanced antioxidant activity, and vacuolar compartmentalization of Cd to mitigate toxicity[64]. The RF model variation confirmed that the toxicity observed under short-term exposures may not accurately represent long-term adaptive responses, emphasizing the importance of time-scale considerations in risk assessment.
Growth medium conditions further modulated toxicity expression. Hydroponic media provided a simplified environment but amplified toxicity, as MPs and Cd readily interfered with nutrient and water uptake[22]. Soil media, with greater buffering capacity and microbial activity, moderated Cd availability but still allowed significant accumulation under high contaminant loads[65]. In soils, microbial communities and organic matter can either immobilize Cd or alter MP surface chemistry, creating complex interactions that shape plant responses[42,66]. Notably, both media showed that toxicity was mediated not only through oxidative stress (Fig. 5) but also through impaired nutrient absorption and ion homeostasis (Supplementary Figs S12–S14)[38].
Environmental conditions can influence Cd toxicity by regulating its adsorption onto MP surfaces. Factors such as Cd speciation, retention time, and soil properties play important roles in determining the adsorption capacity of MPs[34,67]. In natural environments, processes such as rainfall and solar radiation promote the aging of MPs. Aged MPs typically exhibit rougher, more porous, and wrinkled surfaces, which enhance their capacity to adsorb heavy metals compared with pristine MPs[68]. This enhanced adsorption is attributed to increased surface area, the formation of additional adsorption sites, and the introduction of oxygen-containing functional groups during aging[69]. Moreover, natural soil components, including sand, gravel, and clay particles, can accelerate the physical aging of MPs through abrasion processes[70]. As a result, aged MPs in soil generally exhibit stronger heavy metal adsorption, indicating that the degree of aging plays a critical role in regulating metal adsorption-desorption dynamics. This may partly explain why co-exposure to MPs reduces Cd toxicity relative to Cd alone, while in some cases promoting plant growth and development.
Furthermore, the RF model highlighted that experimental conditions, particle characteristics, soil pH, plant species, growth media, and duration collectively drive variability in plant growth and Cd accumulation (Supplementary Fig. S7). These key variables should be prioritized in future studies and considered in environmental management strategies aimed at mitigating Cd contamination in agricultural systems.
Implications and perspectives
-
The evidence from this study highlights that MP–Cd complexes are not governed by a single factor but emerge from the combined effects of particle characteristics, environmental conditions, and plant adaptive responses. This complexity means that the risks associated with MPs and Cd must be evaluated in site-specific contexts that account for the soil environment and crop physiology. Neglecting these combined effects may lead to misleading risk evaluations. For instance, while some MPs can immobilize Cd and reduce its uptake into crops, nanoscale plastics (< 1 µm) may instead increase Cd bioavailability and translocation, raising potential food safety concerns. Such context-dependent duality emphasizes the importance of developing tailored risk assessment frameworks for co-contaminants.
Beyond Cd, MPs can act as vectors for diverse contaminants, including pesticides, antibiotics, and other heavy metals, thereby influencing contaminant transport, persistence, and bioavailability in soils. Their presence may synergistically enhance the immobilization of toxicants in some cases, while in others, they may compete with soil surfaces or organic matter for adsorption sites, altering contaminant equilibria. These interactions expand the scope of MPs beyond being passive pollutants, positioning them as active participants in shaping the fate and effects of multiple contaminants in agroecosystems.
Taken together, these findings underscore that remediation strategies must account for MPs when managing hazardous substances in agricultural soils. Approaches such as soil pH regulation, organic amendments, and microbe-assisted remediation may achieve greater effectiveness when explicitly accounting for the presence of MPs and their physicochemical characteristics. Future research should prioritize field-based validation under realistic agricultural conditions, especially considering aged plastics colonized by biofilms, as these better reflect environmental conditions. Additionally, emphasis should be placed on multi-contaminant interactions, since MPs may simultaneously interact with Cd, Pb, pesticides, antibiotics, and organic pollutants. Advancing understanding of these combined effects will be critical for accurate risk assessment and for the design of effective remediation and management strategies.
Study limitations
-
This meta-analysis provides new insights into the toxicity of Cd and the modulatory role of MPs; however, several limitations should be acknowledged. The dataset is geographically biased, with most studies conducted in Asia, and far fewer originating from other regions. Given the wide variation in agroecological zones, soil properties, and crop species across regions, this lack of coverage limits the global applicability of the findings.
Another limitation is that the majority of studies were performed under controlled laboratory or greenhouse conditions, which cannot fully replicate the complexity of field environments. Natural soils are subject to heterogeneous physical, chemical, and biological interactions, including fluctuating pH, variable organic matter, and dynamic microbial communities, all of which strongly influence the behavior of MP–Cd complexes.
Considerable variability in experimental parameters, such as polymer type, particle size, doses, exposure duration, and plant developmental stage, introduces heterogeneity that obscures mechanistic generalizations. Moreover, limited information on soil physicochemical properties and plant–microbe interactions constrains the ability to evaluate key processes governing Cd mobility, uptake, and detoxification pathways.
Finally, although our analysis indicates that Cd bound to MPs may reduce immediate plant uptake, this does not eliminate long-term risks. Cd retained in soil matrices can persist and potentially remobilize under changing environmental conditions, representing a delayed hazard to agroecosystems and food safety.
Future research should aim to expand geographical representation, validate findings under realistic field conditions, standardize methodological approaches, and investigate long-term interactions under multifactorial stress scenarios. Addressing these gaps will enhance the predictive accuracy of MP–Cd risk assessments and improve their relevance for sustainable soil management.
-
This global meta-analysis advances a mechanistic framework for understanding how MPs modulate Cd toxicity in plants. Our findings confirm the well-established detrimental impacts of Cd on plant growth, photosynthesis, protein content, and oxidative balance, while also demonstrating that co-exposure with MPs can alleviate these effects. The antagonistic role of MPs appears to derive from multiple mechanisms, including sorption of Cd onto MP surfaces, physical barrier effects at the root–soil interface, modulation of antioxidant defenses, and alterations in nutrient homeostasis. Importantly, the extent of MP-mediated reduction is highly context-dependent, varying with particle size, polymer type, exposure duration, and soil pH. Larger particles primarily act as external sorbents, reducing Cd toxicity, whereas smaller particles penetrate more easily and exacerbate disruptions in nutrient balance. Similarly, alkaline soil conditions diminish Cd bioavailability through enhanced MP–Cd complexation and precipitation of insoluble Cd forms. Beyond soil–plant systems, the findings carry broader implications for environmental governance. As MPs can act as carriers for pesticides, antibiotics, and other heavy metals, their presence must be incorporated into global strategies for crop improvement, soil remediation, plastic regulation, and multi-pollutant management. This highlights the need to move beyond single-pollutant frameworks toward integrated, multi-contaminant governance that addresses the coupled challenges of plastic pollution and agrochemical contamination.
-
It accompanies this paper at: https://doi.org/10.48130/newcontam-0026-0013.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: Imran Azeem: writing an original draft, methodology, data collection, formal analysis, proofreading, and conceptualization; Zhen Shi: writing an original draft, methodology, proofreading; Li Xiong: writing an original draft, methodology, proofreading; Jawad Ullah: writing an original draft, methodology, proofreading; Farooq Shah: writing an original draft, methodology, proofreading; Wei Wu: conceptualization, investigation, editing, review, and funding acquisition. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
This study was financially supported by Postdoctoral Research Projects in Hainan Province (Grant No. RZ2500009203).
-
The authors declare that they have no conflict of interest.
-
Full list of author information is available at the end of the article.
- The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Azeem I, Shi Z, Xiong L, Ullah J, Shah F, et al. 2026. Microplastic-mediated modulation of Cd toxicity: evidence from a global meta-analysis. New Contaminants 2: e016 doi: 10.48130/newcontam-0026-0013 

Microplastic-mediated modulation of Cd toxicity: evidence from a global meta-analysis
- Received: 29 January 2026
- Revised: 27 March 2026
- Accepted: 21 April 2026
- Published online: 09 May 2026
Abstract: Microplastics (MPs) and cadmium (Cd) contamination in agroecosystems pose significant threats to crop productivity and food safety. The adverse effects of Cd alone on plant growth and physiology are well established, while the mechanisms by which MPs modulate Cd toxicity remain poorly understood. We conducted a global meta-analysis comprising 4,243 pairwise observations to quantify the effects of Cd alone and in combination with MPs on plant performance, with a focus on MP-mediated modulation of Cd toxicity. Across global-scale studies, Cd exposure was found to significantly reduce biomass production (–29%), photosynthetic performance (–28%), and protein content (–49%), while markedly increasing oxidative damage (+191%) and Cd accumulation in plant tissues (up to 12-fold). Co-exposure with MPs generally reduced Cd toxicity relative to Cd alone; the reductions in biomass (–21%) and photosynthetic activity (–24%) were less severe, while the increases in oxidative damage (+92%) and Cd accumulation (+3.8-fold) were also less pronounced. The observed antagonistic effect is likely attributable to the sorption of Cd onto MPs' surfaces and the formation of physical barriers at the root–soil interface, restricting Cd uptake and translocation in plants. The role of MPs in modulating Cd toxicity is further evidenced by the antioxidant defense system and nutrient balance. In addition, the capacity of MPs to modulate Cd toxicity is highly context-dependent, varying with particle size, polymer type, and soil pH. For example, larger particles reduced toxicity by –11% vs –23% for smaller ones, likely because they act mainly as external sorbents. Alkaline pH further lowered Cd bioavailability and plant uptake, likely through MPs–Cd complexation and Cd precipitation as hydroxides or carbonates. These findings demonstrate that MPs can markedly alter Cd toxicity, and offer critical evidence to improve risk assessment of co-contaminated soils, and to guide targeted management strategies for sustainable agriculture and global food security.
-
Key words:
- Cadmium /
- Microplastics /
- Accumulation /
- Phytotoxicity /
- Meta-analysis