






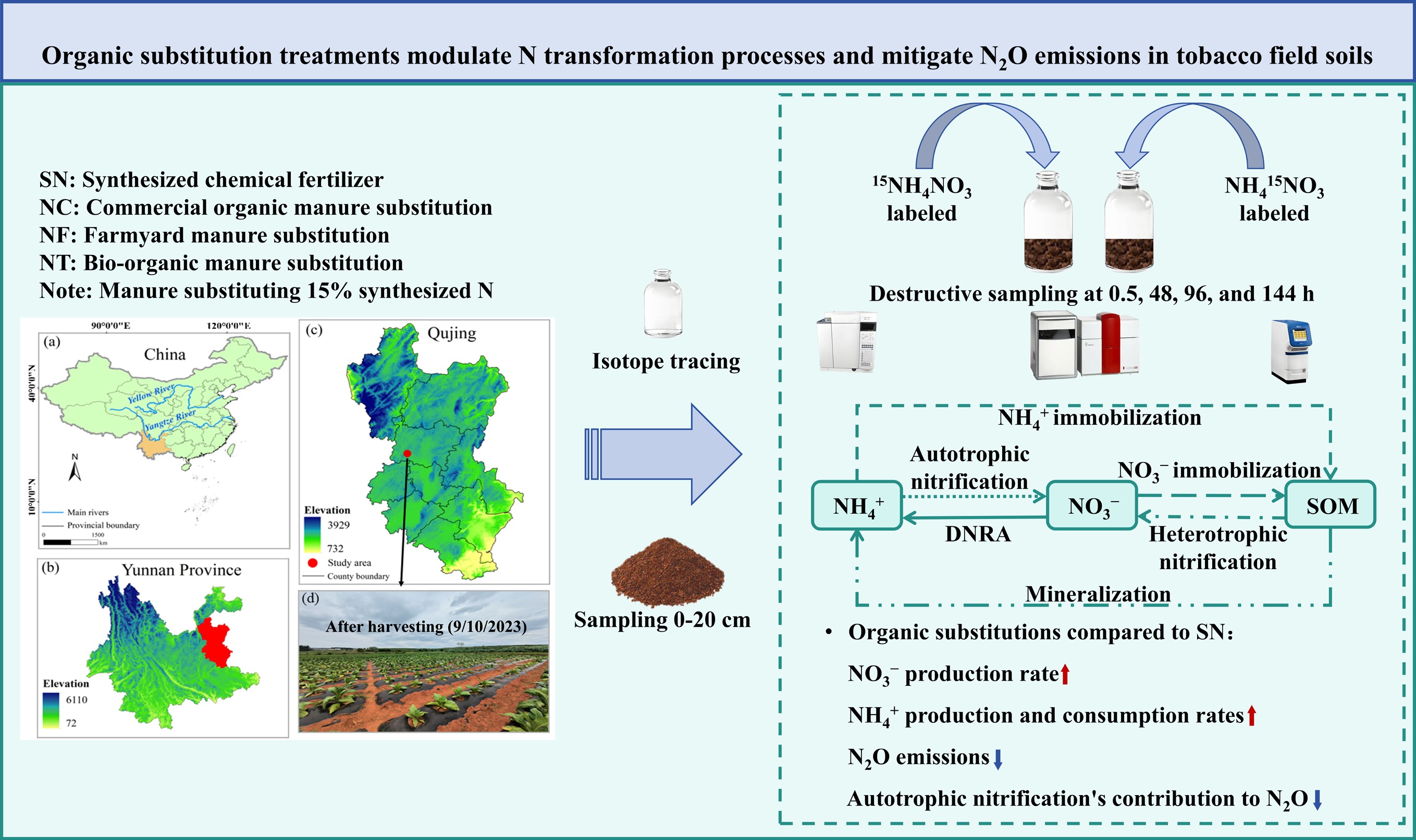



-
Since the 1980s, Chinese farmers have applied high nitrogen (N) fertilizer rates to overcome N limitation, but this has led to excessive N losses and increased nitrous oxide (N2O) emissions. With a 100-year global warming potential approximately 273-fold greater than that of carbon dioxide, N2O exacerbates global warming[1−3]. In this context, a key scientific and practical challenge for modern agriculture is to decrease reliance on mineral N fertilizers while sustaining crop productivity or limiting yield losses to an acceptable level, so as to reconcile food production demands with greenhouse gas mitigation and broader environmental protection objectives. In response, the Chinese Ministry of Agriculture launched the 'Zero Growth of Chemical Fertilizer' program in 2015, aiming to halt the growth of fertilizer use by 2020[4].
Among the options for reducing N inputs while improving efficiency, organic substitution—partially substituting chemical N fertilizer with organic manure—has been widely regarded as a more environmentally friendly approach[5]. It not only enhances agro-ecosystem sustainability but also exerts a pivotal influence on soil N transformations and utilization[6]. Organic N in manure, unlike N directly supplied by chemically synthesized fertilizers, is gradually released through microbial decomposition and mineralization, effectively maintaining the dynamic balance of soil N[7]. In nitrification, organic manures boost the activity of ammonia-oxidizing and nitrite-oxidizing microorganisms by increasing soil organic matter and N sources, thereby improving nitrate N (NO3−) production and soil N availability[8]. Additionally, organic manures enhance soil structure, promote anaerobic microsites, and supply denitrifying microorganisms with carbon (C) and suitable conditions that can increase denitrification rates. While enhanced denitrification may alleviate NO3− accumulation and leaching, it may simultaneously increase N2O emissions, raising concerns about the overall climatic impact[9,10]. Most previous studies have focused on staple crops such as rice and wheat. Here, we shift the focus to tobacco (Nicotiana tabacum L.), a crop that is highly sensitive to N supply and has distinctive nutrient-demand characteristics, with the aim of identifying and elucidating its specific patterns of soil N cycling under organic substitution.
Although numerous studies have examined how organic manures influence soil N transformations, these processes in red soils may differ significantly from those in other soil types because of their distinctive physicochemical characteristics. In southern China, red soils are typically low in organic matter and highly acidic, conditions that inhibit nitrification and slow the conversion of ammonium N (NH4+) to NO3−[11,12]. Additionally, red soils are rich in Fe-Al oxides, which interact with organic matter and NH4+, accelerating N2O production during denitrification and inhibiting the subsequent reduction of N2O to N2, ultimately intensifying N2O emissions[13,14]. Poor aeration in these soils also promotes anaerobic microsites that truncate denitrification at the N2O stage, resulting in higher N2O emissions and reduced fertilizer N use efficiency[15].
Tobacco is highly sensitive to both the form and timing of N supply, and both its productivity and leaf quality are closely coupled to N availability in the root zone. In many tobacco-growing regions of China, including Yunnan, tobacco is cultivated predominantly on acidic red soils with low organic matter content and limited buffering capacity[16]. Under such conditions, inappropriate N management can readily lead to soil acidification, poor structure, unbalanced NH4+/NO3− ratios, and elevated N2O emissions, ultimately constraining both N use efficiency and leaf quality[17]. The substitution of mineral fertilizer with organic manure in tobacco production entails both opportunities and challenges. On the one hand, it can improve soil physicochemical properties and biological activity, thereby enhancing soil quality and sustaining long-term nutrient supply[18]. On the other hand, the asynchronous release of N from organic manure over time may not coincide with crop N demand, and the increased availability of labile C and N may enhance ammonia volatilization and N2O emissions[19]. Organic manures have the potential to partially replace mineral N fertilizers in these systems. By supplying labile C, they can mitigate soil acidification, improve aggregate stability and pore structure, and stimulate microbial activity, thereby reshaping soil C–N coupling and N transformation rates in tobacco fields. Because tobacco roots may differentially prefer NH4+ vs NO3− depending on growth stage and environmental conditions, tobacco root exudation traits, together with regional cultivation systems, can jointly shape nitrifying and denitrifying communities, ultimately modulating N transformation rates and N2O emission pathways[20].
Soil N transformation processes are complex, and different organic substitution treatments exert distinct effects on these transformations. Bio-organic manure, owing to its microbial inoculants and suitable C/N ratio, can slow nitrification, modulate denitrification, and reduce N2O emissions[21]. Commercial organic manure, which undergoes industrial processing and fermentation, typically contains stable, plant-available N forms readily absorbed by crops, thereby mitigating soil N leaching and excessive N transformations and reducing N2O emissions[22]. Farmyard manure provides a long-term substrate for soil microorganisms due to its diverse, slow-releasing N sources, resulting in relatively low N2O emissions despite continuous denitrification[23]. However, it remains unclear how these contrasting organic substitution treatments differentially affect N transformation processes and N2O emission pathways in acidic red soils under tobacco cultivation, and whether there are specific thresholds or applicability ranges for each treatment in such systems. Given the prevalence of acidic red soils in Yunnan, systematically evaluating organic substitution treatments and identifying their applicability ranges and thresholds are indispensable for enhancing N use efficiency and developing N2O emission-reduction treatments.
This study was designed to elucidate how distinct organic substitution treatments influence gross N transformation and N2O emission pathways, and their underlying mechanisms. Using 15N labeling techniques (e.g., 15NH4NO3 and NH4 15NO3), we traced N transformation processes, determined transformation rates, and assessed the roles of autotrophic nitrification, heterotrophic nitrification, denitrification, and co-denitrification in N2O emissions. The specific objectives were to: (1) accurately measure gross N transformation rates in tobacco-cultivated soils; and (2) quantify the contributions of different pathways to N2O production. We hypothesize that: (1) different organic substitution treatments lead to varied gross N transformations, resulting in distinct NH4+ and NO3− production and consumption rates; and (2) different organic substitution treatments alter the relative importance of autotrophic nitrification, heterotrophic nitrification, denitrification, and co-denitrification as N2O production pathways, ultimately leading to treatment-dependent variation in N2O emissions.
-
The field study was carried out in Qujing City (25°40' N, 103°39' E), Yunnan Province. The site has a mean annual temperature of 14.5 °C and a mean annual precipitation of 1,218.5 mm. According to the International Soil Texture Classification System, the tobacco-planting soil is categorized as a silty clay loam, comprising 27.2% clay, 6.5% sand, and 66.3% silt. For detailed methods used to determine soil physicochemical characteristics, refer to the Supplementary Text S1.
The experiment followed a randomized block design comprising four nutrient management treatments, in which 15% of the mineral N input was substituted with organic manure, consistent with local conventional fertilization practices. Specifically, the four fertilization treatments were: mineral N fertilizer (SN); partial substitution of mineral N fertilizer with commercial organic manure (NC); partial substitution of mineral N fertilizer with farmyard manure (NF); and partial substitution of mineral N fertilizer with bio-organic manure inoculated with Trichoderma viride Pers. (NT). On May 2, 2023, tobacco plants of the 'Yunyan 100' variety were transplanted according to the local agricultural schedule, and harvesting was completed on September 10. The detailed fertilization program and field management practices are shown in Supplementary Tables S1 and S2. After harvest, soil was collected from the 0–20 cm cultivated layer and passed through a 2 mm stainless-mesh following manual removal of visible plant residues, roots, and gravel. Each soil sample was then split into two subsamples: one fraction was air-dried and stored at ambient laboratory temperature for physicochemical analysis, whereas the other portion was immediately frozen at −80 °C for subsequent 15N tracer experiment.
Laboratory 15N tracing incubation experiment
-
In this experiment, two sets of 15N-labeled treatments were applied to each soil sample: (1) 15NH4+ treatment by adding 15NH4NO3 solution (10.23 15N atom%), and (2) 15NO3− treatment by adding NH4 15NO3 solution (20.25 15N atom%). Two criteria were applied to set the tracer abundance: (1) the atom% 15N in the targeted soil nitrogen pool must be approximately 5%–10% at 0.5 h after labeling; and (2) by the end of the incubation, the atom% 15N in all labeled pools remained measurably above the natural isotopic background. We selected the tracer addition volume to comply with standard soil incubation protocols and the specifications of the 15N isotope dilution method. The detailed procedures followed those described by Duan et al.[24] and He et al.[25]. By monitoring 15N dynamics across distinct soil N pools, we isolated N dynamics from confounding field variability and directly assessed how different treatments influence the soil's capacity to incorporate newly applied inorganic N.
Fresh soil, equivalent to 20 g on an oven-dry basis, was weighed and transferred into 120 mL serum vials, then pre-incubated in the dark at 25 °C for 48 h. After this acclimation period, 1 mL of either 15NH4NO3 or NH4 15NO3 solution was added with a pipette, supplying 50 mg N kg−1 dry soil. Three independent replicates were prepared for each labeling treatment. Soil water content was regulated to 60% of field water-filled pore space, and vials were covered with perforated sealing, which limited evaporation while maintaining aerobic conditions. Incubations proceeded in the darkness at 25 °C, and destructive sampling was conducted at 0.5, 48, 96, and 144 h after tracer application. At each sampling time, soil NH4+ and NO3− were extracted with 2 M KCl to determine their concentrations and 15N abundance. Following extraction of inorganic N, the residual was rinsed with deionized water to eliminate remaining mineral N, subsequently oven-dried, and then utilized for determination of organic N concentration and 15N abundance. Gaseous samples were collected at 48, 96, and 144 h. Before collection, the vials were tightly sealed and purged with mineral air (22% oxygen and 78% nitrogen) for 6 h to standardize the gas composition. Gas samples were then taken to assess N2O concentration and 15N abundance. N2O concentration was analyzed using a gas chromatograph (Agilent 7890A, Agilent Ltd, China), and the 15N abundance of NH4+, NO3−, organic N, and N2O was analyzed with an isotope ratio mass spectrometer (Europa Scientific Integra, Crewe, UK)[25]. For details on calculating N transformation rates and cumulative N2O emissions contributions, see the Supplementary Text S1.
N cycling functional gene abundances
-
DNA was extracted from soil samples at the end of incubation using the Fast DNA SPIN Kit for Soil (MP Biomedicals, USA), and its concentration and purity were assessed with a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA), after which the extracts were diluted to 10 ng μL−1 and their concentrations further verified on a StepOnePlus Real-Time PCR System (Applied Biosystems, USA). The assays targeted genes associated with N2O emissions, including the archaeal and bacterial ammonia monooxygenase genes (AOA-amoA and AOB-amoA) associated with nitrification, and the denitrification-related genes nirS, nirK, fungal-nirK, nosZI, and nosZII. Each qPCR mixture had a final volume of 20 μL, consisting of 10 μL of SYBR Green master mix (TaKaRa Biotech, Dalian, China), 0.4 μL of ROX reference dye, 2 μL of template DNA, 6.8 μL of nuclease-free water, and 0.4 μL of each primer (forward and reverse)[24]. All amplifications were set up in triplicate for each sample, and a no-template control (nuclease-free water in place of DNA) was included. A standard curve was established using a plasmid harboring the target gene fragment, and a 10-fold serial dilution series (six points) was prepared for quantification. A single peak in the melting curve confirmed proper amplification. Copy numbers were log-transformed to normalize the data prior to statistical analysis. qPCR primers are detailed in Supplementary Table S3.
Data analysis
-
Statistical analyses were conducted using one-way analysis of variance (ANOVA), and treatment means were subsequently separated with Duncan's multiple range test at a significance threshold of p < 0.05 in IBM SPSS Statistics 27 (IBM, Chicago, Illinois, USA). Pairwise linear relationships among variables were examined using Pearson's correlation analysis. Graphs were created using Origin v2021 and ChiPlot (
www.chiplot.online ). The model's optimal solution was found using the fmincon function in MATLAB v2021. -
The one-way ANOVA results indicated that organic substitution treatments exerted significant effects on most soil physicochemical characteristics, except for pH and DON (Table 1). No measurable parameter differed significantly (p > 0.05) between the NC and NT treatments. Compared with SN, the NT treatment resulted in markedly elevated SOC and TN contents, while the C/N ratio remained statistically unchanged. In contrast, the NF treatment exhibited the lowest TN content and the highest C/N ratio, both of which differed significantly from those of NT and NC. NF also had the highest DOC content, significantly exceeding that of the SN treatment (p < 0.05). Overall, organic substitution treatments improved soil physicochemical characteristics, with each treatment displaying specific strengths in particular indicators.
Table 1. Soil physicochemical characteristics (mean ± SD) under different organic substitution treatments
Treatment pH SOC (g kg−1) TN (g kg−1) C/N ratio DOC (mg kg−1) DON (mg kg−1) SN 5.87 ± 0.02a 20.05 ± 2.25b 1.44 ± 0.09b 15.42 ± 2.06ab 21.29 ± 1.06b 24.37 ± 1.67a NC 6.41 ± 0.50a 22.83 ± 0.99ab 1.80 ± 0.16a 12.78 ± 1.44b 25.68 ± 0.13ab 28.93 ± 4.48a NF 6.45 ± 0.49a 23.57 ± 2.08ab 1.42 ± 0.05b 16.56 ± 1.33a 28.73 ± 1.06a 27.62 ± 0.58a NT 6.33 ± 0.52a 25.79 ± 0.20a 1.99 ± 0.14a 13.03 ± 0.84b 25.54 ± 4.26ab 25.98 ± 0.58a pH: soil potential of hydrogen; SOC: soil organic carbon; TN: total nitrogen; C/N ratio: SOC/TN ratio; DOC: dissolved organic carbon; DON: dissolved organic nitrogen; SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N. Significant differences among treatments are denoted by different letters (one-way ANOVA, Duncan's multiple range test, p < 0.05). N transformation rates
-
Table 2 presents the gross production and consumption rates of NH4+ and NO3−, along with their percentage contributions to the N transformation rates under different organic substitution treatments. Overall, gross NO3− production exceeded gross NH4+ production across all four fertilization treatments, while gross NH4+ consumption exceeded NO3− consumption. The gross NO3− production and NH4+ consumption rates ranked in the order: NF > NT > NC > SN, with NC, NF, and NT showing significantly higher values than SN, and significant differences were also observed among NC, NF, and NT (p < 0.05). Gross NO3− consumption rate followed the pattern: NT > NF > NC > SN, while gross NH4+ production rate followed the order: NT > NC > NF > SN. The NT treatment showed significantly elevated NO3− consumption and NH4+ production rates relative to the other treatments (p < 0.05).
Table 2. The average gross NH4+ and NO3− production and consumption rates, along with their percentage contributions to the total N pool production or consumption, under different organic substitution treatments at incubation times ranging from 0.5 to 144 h
N transformation rate SN (mg N kg−1
soil d−1)Contribution NC (mg N kg−1
soil d−1)Contribution NF (mg N kg−1
soil d−1)Contribution NT (mg N kg−1
soil d−1)Contribution Gross NO3− production 6.00 ± 0.04d 6.43 ± 0.14c 7.43 ± 0.13a 7.21 ± 0.04b Autotrophic nitrification 3.90 ± 0.13c 65.0% 4.07 ± 0.02c 63.4% 4.68 ± 0.14b 63.0% 5.39 ± 0.01a 74.8% Heterotrophic nitrification 2.10 ± 0.17c 35.0% 2.35 ± 0.15b 36.6% 2.74 ± 0.07a 37.0% 1.82 ± 0.03d 25.2% Gross NO3− consumption 2.88 ± 0.05b 3.14 ± 0.10b 3.18 ± 0.38b 3.58 ± 0.01a DNRA 0.32 ± 0.08c 11.1% 0.47 ± 0.05b 15.1% 0.52 ± 0.03b 16.5% 1.25 ± 0.02a 34.9% NO3− immobilization 2.54 ± 0.04a 88.1% 1.82 ± 0.10c 57.8% 2.19 ± 0.05b 68.7% 2.27 ± 0.05b 63.3% Gross NH4+ production 0.88 ± 0.04d 1.21 ± 0.04b 1.01 ± 0.03c 1.37 ± 0.01a DNRA 0.32 ± 0.08c 36.4% 0.47 ± 0.05b 39.3% 0.52 ± 0.03b 51.8% 1.25 ± 0.02a 90.9% Mineralization 0.56 ± 0.04b 63.6% 0.73 ± 0.03a 60.7% 0.49 ± 0.06c 48.2% 0.13 ± 0.00d 9.1% Gross NH4+ consumption 6.14 ± 0.10d 6.50 ± 0.03c 6.96 ± 0.02a 6.81 ± 0.02b Autotrophic nitrification 3.89 ± 0.14c 63.5% 4.07 ± 0.02c 62.6% 4.68 ± 0.14b 67.3% 5.39 ± 0.01a 79.2% NH4+ immobilization 1.30 ± 0.03a 21.0% 1.16 ± 0.12ab 17.9% 1.07 ± 0.05b 15.4% 1.17 ± 0.04ab 17.2% Nitrate nitrogen: NO3−; ammonium nitrogen: NH4+; dissimilatory nitrate reduction to ammonium: DNRA; SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N. Significant differences among treatments are denoted by different letters (one-way ANOVA, Duncan's multiple range test, p < 0.05). In NO3− production, autotrophic nitrification was the primary contributing process, accounting for 65.0%, 63.4%, 63.0%, and 74.8% of the gross NO3− production across all four fertilization treatments. It was also the main process for NH4+ consumption, contributing 63.5%, 62.6%, 67.3%, and 79.2% of the gross NH4+ consumption across all four fertilization treatments. Autotrophic nitrification in the NF and NT treatments was significantly elevated relative to that observed in the SN treatment.
DNRA and NO3− immobilization contributed over 70% to NO3− consumption, with NO3− immobilization as the main process. In contrast, DNRA was the primary source of NH4+ production in the NF and NT treatments, contributing 51.8% and 90.9%, respectively, while mineralization was the dominant process in the SN and NC treatments, contributing 63.6% and 60.7%, respectively.
N2O emission pathways
-
Figure 1a shows that the N2O emission rates remained dynamic at 60% of the field water-filled pore space, with a significant emission peak on the fourth day of the labeling experiment. Among the four fertilization treatments, the cumulative N2O emissions were, in descending order, SN > NF > NC > NT, with organic substitution treatments producing significantly lower emissions than the SN treatment (p < 0.05), whereas differences among the organic substitution treatments were not statistically discernible (Fig. 1b). Relative to the SN treatment, the organic substitution treatments also attenuated the cumulative N2O emissions from autotrophic nitrification. However, these treatments had varying effects on N2O emissions from denitrification and heterotrophic nitrification. NF and NC increased N2O emissions during denitrification, while NT decreased them. NC increased N2O emissions during heterotrophic nitrification, whereas NF and NT reduced them.
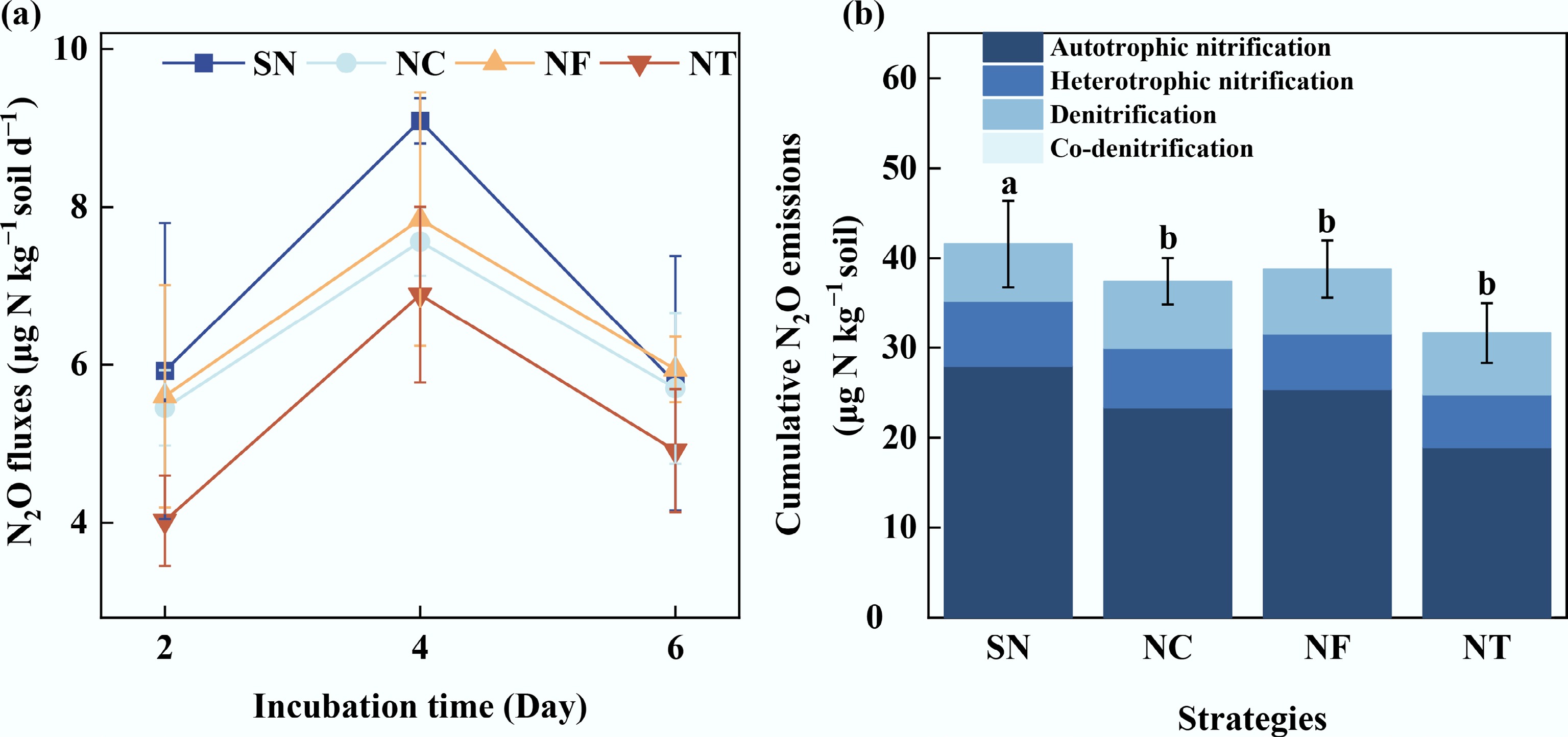
Figure 1.
(a) N2O fluxes, and (b) cumulative N2O emissions via four pathways in tobacco fields under organic substitution treatments. SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N. Significant differences among treatments are denoted by different letters (one-way ANOVA, Duncan's multiple range test, p < 0.05).
Throughout the incubation period, the relative contribution of autotrophic nitrification to N2O emission pathways increased over incubation time, whereas the proportional roles of heterotrophic nitrification and denitrification decreased (Fig. 2). On average, autotrophic nitrification accounted for 67.2%, 62.3%, 65.4%, and 59.7% of N2O emissions under the four fertilization treatments; heterotrophic nitrification contributed 15.3%, 16.6%, 15.8%, and 18.6%, respectively; and denitrification contributed 17.5%, 20.1%, 18.8%, and 21.7%, respectively. Co-denitrification made no significant contribution to N2O emissions.
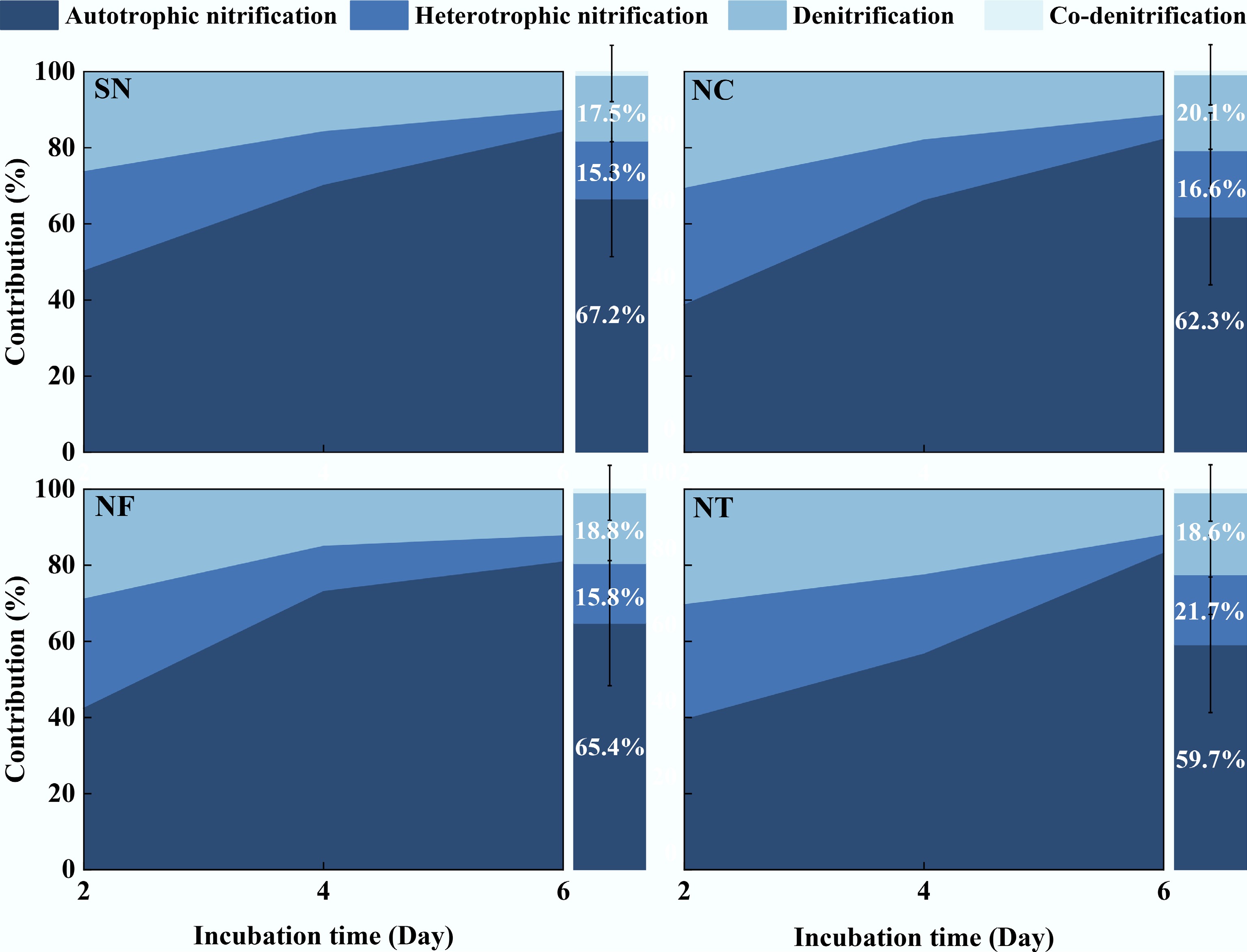
Figure 2.
N2O emission pathways and their average contributions in tobacco fields under organic substitution treatments. SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N.
Abundance of N2O-related functional genes
-
Organic substitution treatments significantly affected the copy numbers of nitrification and denitrification functional genes in tobacco soil (Fig. 3). Relative to the SN treatment, the NF treatment enhanced the abundance of the AOA-amoA gene by 2.7%, and nosZII gene abundance by 7.4% (p < 0.05). However, under the NC and NT treatments, nirK gene abundance decreased by 2.4% and 3.1%, respectively, relative to SN (p < 0.05). Under the three organic substitution treatments, NF consistently supported higher abundances of AOA-amoA, nirK, and nosZII than either NC or NT, and AOB-amoA gene abundance was also greater in NF than NT (p < 0.05). Additionally, nosZII gene abundance differed significantly among all four fertilization treatments (p < 0.05). No significant differences were detected for nirS, fungal-nirK, and nosZI gene abundances across the treatments.
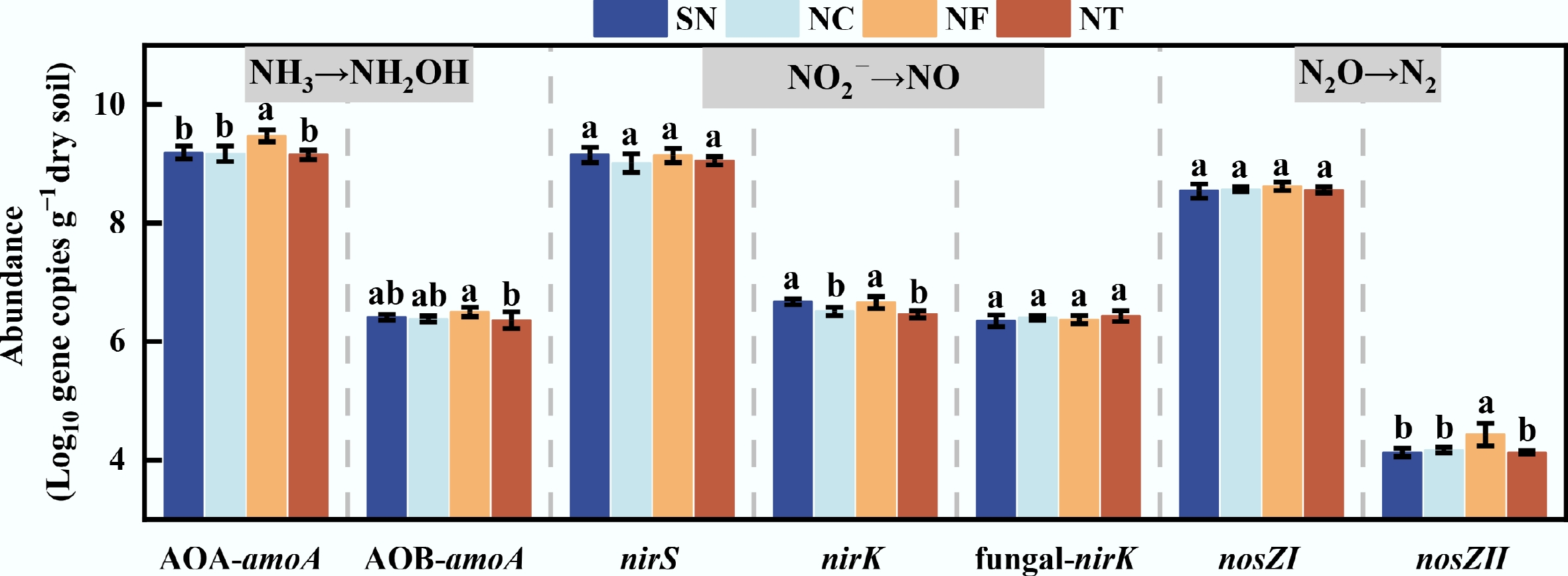
Figure 3.
Log10-transformed gene copy numbers of soil functional genes (AOA-amoA, AOB-amoA, nirS, nirK, fungal-nirK, nosZI, and nosZII) after 144 h of incubation under different organic substitution treatments. SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N. Significant differences among treatments are denoted by different letters (one-way ANOVA, Duncan's multiple range test, p < 0.05).
Inter-group correlation analysis
-
Soil biochemical characteristics influenced N transformation rates and N2O emission pathways in different patterns. During N transformations (Fig. 4a), DOC showed a significant negative relationship with both gross NO3− production and NH4+ consumption, whereas SOC and fungal-nirK were negatively associated with gross NO3− consumption and NH4+ production (p < 0.05). In contrast, the nirK gene exhibited a positive correlation with gross NO3− consumption and NH4+ production. In addition, the C/N ratio and nirS were positively related to gross NH4+ production (p < 0.01). Regarding N2O emission pathways (Fig. 4b), SOC, TN, and fungal-nirK were positively correlated with cumulative N2O emissions from autotrophic nitrification (p < 0.01), whereas the C/N ratio, nirS, and nirK genes were negatively correlated (p < 0.05).
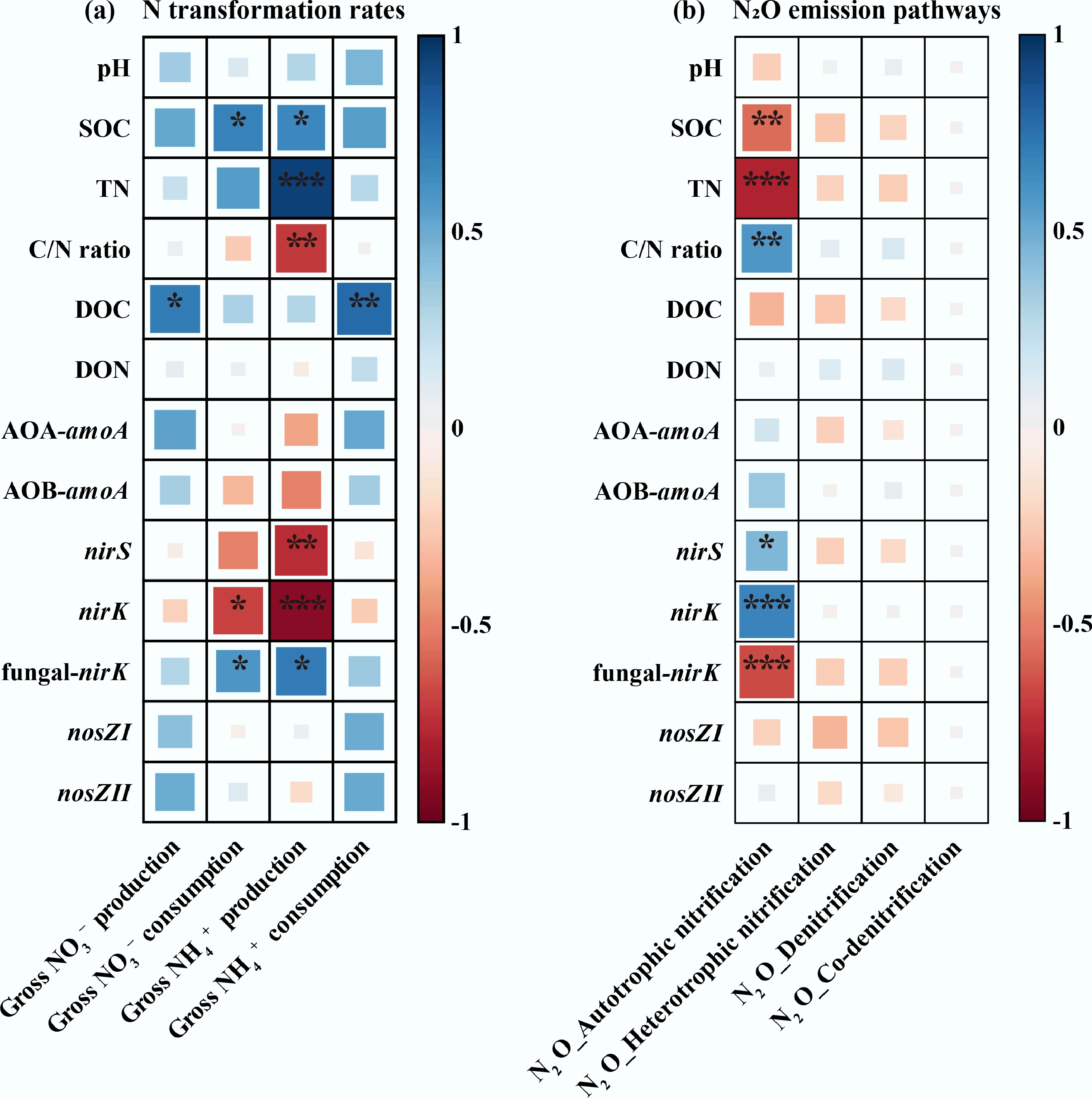
Figure 4.
Heatmap of Pearson's correlation matrix between soil biochemical characteristics and gross NH4+ and NO3− (a) production and consumption rates, and (b) cumulative N2O emissions via pathways in tobacco soils. SN: synthesized chemical fertilizer; NC: commercial organic manure substituting 15% synthesized N; NF: farmyard manure substituting 15% synthesized N; NT: bio-organic (Trichoderma viride Pers.) manure substituting 15% synthesized N. * indicates significance at p < 0.05; ** indicates significance at p < 0.01; *** indicates significance at p < 0.001.
-
Our study used a 15N tracer addition rate of 50 mg N kg−1 soil, which exceeds typical in situ inorganic N pools. This rate was chosen to ensure analytical precision for estimating gross N transformation rates, consistent with previous 15N tracer studies[24]. We acknowledge that this relatively high N input may stimulate microbial N processes and thereby lead to overestimation of absolute gross rates relative to field conditions; therefore, our results should be interpreted as potential, rather than strictly in situ rates. Nevertheless, because all treatments received the same tracer addition under identical conditions, any systematic bias should be consistent across treatments, and the observed treatment differences and main conclusions remain valid.
In tobacco fields in Yunnan, the gross rate of NO3− production exceeded that of NH4+ production, while the consumption of NH4+ was generally higher than the consumption of NO3− (Table 2). These patterns indicate an intensive nitrification process and a rapid turnover of the NH4+ pool, rather than a simple accumulation of mineral N. This pattern is closely linked to the ecological conditions of tobacco-growing regions. Tobacco is a typical nitrate-preferring crop, and its fertilization and quality control rely primarily on NO3− as the N source[26]. Once NH4+ is produced in the soil, this NH4+ is rapidly nitrified under the warm, well-aerated conditions of Yunnan, continuously replenishing the NO3− pool. Meanwhile, tobacco has a low tolerance for NH4+, and therefore maintaining high NH4+ levels in the rhizosphere is not desirable[27]. As a result, mineralization provides only limited NH4+, and NH4+ is quickly consumed through nitrification and microbial assimilation, so that its consumption exceeds its production. In contrast, NO3− is continuously replenished by nitrification and fertilization, and its supply often exceeds its removal via denitrification and leaching, leading to a net surplus of NO3−.
This investigation revealed that organic substitution treatments significantly affected N production and consumption processes, as evidenced by quantified N transformation rates. Relative to the SN treatment, treatments involving organic substitution produced substantially greater gross NO3− and NH4+ production and consumption. This indicates that the organic matter and associated microorganisms delivered by organic manures enhance soil N supply capacity by stimulating microbial biomass and metabolic activity, thereby accelerating the transformation of inorganic N pools[8,28]. The NT treatment showed the most significant improvement in gross NH4+ production and NO3− consumption rates, indicating that bio-organic manure inputs provide readily available C and N and enrich beneficial microbial communities, leading to faster production and consumption of NO3− and NH4+. Additionally, improvements in soil aggregation, porosity, moisture retention, and pH buffering create more favorable and stable micro-environments for microbial functioning, sustaining rapid transformation of these pools[29]. In contrast, the sole mineral inputs under SN treatment likely constrained microbial biomass through C limitation and soil acidification, dampening community activity and slowing N transformations.
Further examination of N transformation processes showed that autotrophic nitrification contributed more to NO3− production and NH4+ consumption, particularly under the NF and NT treatments, where nitrification rates were markedly elevated relative to those in the NC and SN treatments (Table 2). However, the copy numbers of AOA-amoA and AOB-amoA genes under NT treatment were statistically indistinguishable from those in the SN treatment (Fig. 3), suggesting that the enhancement of autotrophic nitrification results from higher ammonia-oxidizing rates per cell or per amoA gene copy, rather than from increased amoA gene abundance. Functional microbes and their metabolites in bio-organic manure help optimize soil microaerophilic niches and suppress pathogenic and heterotrophic microorganisms, thereby reducing competition for oxygen and NH4+. Collectively, these processes substantially strengthen the ammonia-oxidizing capacity of both AOA and AOB, thereby amplifying their overall role in NO3− production[30]. In the present study, despite the higher copy numbers of AOA-amoA and AOB-amoA genes observed under the NF treatment, their nitrification rates were lower than those measured under NT. This apparent discrepancy is probably attributable to the high organic matter content and incompletely decomposed residues in farmyard manure, which favored heterotrophic microbes and immobilization processes. As autotrophic nitrifiers preferentially utilize inorganic NH4+ rather than organic N, a slower mineralization and a stronger microbial competition for inorganic N could restrict their effective substrate supply[31]. Therefore, even with higher amoA gene abundance, ammonia oxidation rates in NF were not significantly enhanced.
The application of organic manures enriched the content of labile organic matter in the soil, thereby supplying ample C sources for microbial metabolism and promoting ammonia oxidation as well as the DNRA process[32]. Heterotrophic microorganisms contribute to NO3− production during organic matter decomposition, activating the native microbial community and increasing heterotrophic nitrifier diversity, which boosts heterotrophic nitrification rate[33]. Consequently, the combined enhancement of autotrophic and heterotrophic nitrification in treatments involving organic substitution markedly accelerated the transformation of NH4+ to NO3−. It is noteworthy that the NF and NT treatments altered the dominant source of NH4+ from organic N mineralization to DNRA. Under the NT treatment, DNRA contributed up to 90.9% of NH4+ production, far exceeding the levels reported for most aerobic soils (Table 2). However, this very high contribution ratio mainly reflects the relatively low gross NH4+ production and weak mineralization rate under NT, rather than an abnormally high DNRA rate; DNRA operated at a normal rate but accounted for most of the limited NH4+ generated. Mechanistically, the bio-organic manure likely stimulated nitrification, so that newly produced NH4+ was rapidly oxidized to NO3− and NH4+ accumulation was suppressed[34]. It also increased the bulk soil C/N ratio and DOC, which stoichiometrically favors NO3− reduction via DNRA over organic N mineralization as a source of NH4+[35]. Although the contribution rate of DNRA increased substantially, the DNRA transformation rate measured in this study (0.32–1.25 mg N kg−1 soil d−1) remained within the typical global range (0.18–1.30 mg N kg−1 soil d−1) and was consistent with existing data for Fujian red soil (0.70–2.38 mg N kg−1 soil d−1)[36,37], indicating that the DNRA transformation rate under our experimental conditions is reasonable at both regional, and global scales.
Variations in N2O emission pathways and magnitudes
-
N2O is a highly effective radiatively active gas, and its generation is primarily controlled by nitrification and denitrification processes[38]. In the present incubation, isotopic partitioning indicated that autotrophic nitrification is the main N2O emission pathway, accounting for over 60% of total emissions (Fig. 2). Compared to SN, all organic substitution treatments exhibited lower cumulative N2O emissions in this assay (Fig. 1). This result is consistent with previous field studies, which have concluded that organic substitution can significantly reduce N2O emissions[39]. Specifically, these treatments effectively reduce N2O emissions during autotrophic nitrification. A plausible explanation is that organic manures typically supply N in more slowly mineralizable forms, which reduce short-term N accumulation and prevent excessive soil N concentrations, inhibiting nitrification by autotrophic bacteria and slowing N2O production and emissions[40]. Under plant-free conditions, although the NT treatment resulted in a higher autotrophic nitrification rate, the application of bio-organic manure improved soil structure and aeration. As a result, nitrification occurs mainly under well-oxygenated, stable conditions, which reduces the fraction of N2O produced during nitrification[41]. At the same time, bio-organic manure increases SOC content and creates localized anaerobic microenvironments within soil aggregates. These localized anaerobic niches intensify denitrification activity and facilitate the further reduction of N2O to N2, thereby further consuming N2O that has already been formed. Accordingly, the contributions of heterotrophic nitrification and denitrification to N2O emissions were significantly reduced under NT, and the abundance of nirK decreased (Fig. 3). Because nirK encodes nitrite reductase in the denitrification pathway, its lower abundance indicates a reduced potential for denitrification-derived N2O[42]. Additionally, the microbial agents and appropriate C/N ratio in bio-organic manure may have promoted the dominance of competitive microorganisms during autotrophic nitrification, further reducing N2O emissions from heterotrophic nitrification and denitrification by inhibiting microbial activity during anaerobic N transformation[21]. The NF treatment reduced N2O emissions during heterotrophic nitrification and significantly increased nosZII gene abundance, promoting N2O reduction to N2 and effectively lowering N2O emissions[43]. In contrast, the NC treatment led to higher N2O emissions during heterotrophic nitrification and denitrification and significantly reduced nirK gene abundance, suggesting suppressed denitrifier activity. Under the NC treatment, soil exhibited a low C/N ratio and elevated DON (Table 1). Commercial organic manure, although formulated and relatively stable in terms of its bulk organic fraction, often contains a substantial proportion of readily available N and labile C, which can be quickly mineralized and denitrified[44]. This localized pulse of high DON combined with a relatively low C/N ratio in the decomposing manure can promote rapid N release and the formation of transient anaerobic microsites, thereby increasing N2O production, even though nirK gene abundance is relatively low. Thus, N2O emission pathways and intensity are closely linked to the fertilization treatment. Over-reliance on chemical N fertilizers increases N2O emissions, while organic substitution treatments can effectively regulate N2O emission pathways, achieving more eco-friendly fertilization goals.
Relationships among soil biochemical characteristics, N transformation rates, and N2O emission pathways
-
This study investigated the interaction of soil biochemical characteristics in N transformations in tobacco soils. Organic substitution treatments significantly improved soil physicochemical characteristics, particularly SOC and TN contents (Table 1). Compared with the SN treatment, NT increased the contents of SOC and TN, indicating that bio-organic manure, by supplying ample C and N, promoted organic matter accumulation and enhanced soil fertility[45]. This likely boosted microbial activity and N metabolism by facilitating N transformations[42]. Correlation analysis showed that SOC and TN contents were negatively associated with gross NH4+ production rate, but positively correlated with cumulative N2O emissions from autotrophic nitrification (Fig. 4). High SOC and TN enhanced microbial activity, leading to competitive consumption of NH4+ and oxygen by microorganisms, which decreased NH4+ production and increased N2O emissions[46]. Additionally, the C/N ratio was positively associated with gross NH4+ production rate, but negatively associated with cumulative N2O emissions from autotrophic nitrification. A high C/N ratio indicates more SOC and less TN, promoting the mineralization of organic N into NH4+. However, a high C/N ratio also implies N deficiency, weakening nitrification and reducing N2O emissions[47]. Although the NF treatment had the lowest TN content, its higher DOC and C/N ratio indicated that farmyard manure might contain more recalcitrant organic matter, reducing mineralization and potentially influencing N release and microbial preference for C sources[48]. This elevated C/N ratio could suppress rapid ammonium release, thereby affecting nitrification rates and ultimately decreasing N losses[49].
In addition, the abundances of nirS and nirK genes were associated with gross NH4+ production rate, but negatively related to cumulative N2O emissions from autotrophic nitrification. High expression of these genes indicated efficient bacterial denitrification, promoting the N cycle, increasing NH4+ production, and reducing N2O emissions[50]. Conversely, the abundance of fungal-nirK genes showed a negative correlation with gross NH4+ production rate, and a positive correlation with cumulative N2O emissions from autotrophic nitrification. High fungal-nirK gene abundance resulted in lower denitrification efficiency, reducing NH4+ production, and increasing N2O emissions[51].
In summary, organic substitution treatments improved soil nutrients and microecological stability by increasing both C and N reserves, adjusting the C/N balance, and enriching microbial C substrates. This strengthened N transformation processes and reduced N2O emissions, offering insights into the complexity of soil N cycling and its associated gaseous losses.
-
This study systematically examined how organic substitution treatments influence gross N transformation processes and N2O emission pathways in tobacco soils. The findings demonstrate that, relative to the SN treatment, organic substitution markedly enhanced NO3− formation and increased both the production and consumption rates of NH4+. Autotrophic nitrification dominated NO3− production and NH4+ consumption, particularly in the NF and NT treatments. Additionally, these treatments substantially reduced cumulative N2O emissions, although autotrophic nitrification remained the primary N2O emission source. In summary, organic substitution treatments modulate N transformation processes and significantly reduce N2O emissions, supporting sustainable agriculture and greenhouse gas mitigation. These findings provide crucial theoretical backing and practical guidance for adopting organic substitution treatments to enhance soil quality and reduce agricultural greenhouse gas emissions.
-
It accompanies this paper at: https://doi.org/10.48130/nc-0026-0008.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: Bingxue Wang: formal analysis, investigation, writing – original draft; Xiaopeng Deng: data curation, resources; Zhonglong Wang: investigation, data curation; Bin Wang: investigation, data visualization; Ruibao Wang: investigation, resources; Yali Zhang: investigation, writing – review & editing; Zhengqin Xiong: conceptualization, funding acquisition, supervision, writing – review & editing. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated and analyzed during the current study are not publicly archived but can be obtained by the corresponding author upon reasonable request.
-
This research was carried out with financial support from the Science and Technology Project of Yunnan Provincial Branch of China National Tobacco Corporation (2023530000241025) and from the National Natural Science Foundation of China (Grant No. 42377292).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Bingxue Wang, Xiaopeng Deng
Full list of author information is available at the end of the article. - The supplementary files can be downloaded from here.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang B, Deng X, Wang Z, Wang B, Wang R, et al. 2026. Effects of substituting mineral nitrogen fertilizer with organic manure on nitrogen transformations and nitrous oxide emissions from tobacco (Nicotiana tabacum L.) fields. Nitrogen Cycling 2: e021 doi: 10.48130/nc-0026-0008 

Effects of substituting mineral nitrogen fertilizer with organic manure on nitrogen transformations and nitrous oxide emissions from tobacco (Nicotiana tabacum L.) fields
- Received: 30 March 2026
- Revised: 23 April 2026
- Accepted: 30 April 2026
- Published online: 13 May 2026
Abstract: The partial substitution of mineral nitrogen fertilizer (SN) with organic manure, hereafter termed organic substitution, is increasingly regarded as an environmentally preferable and more sustainable nutrient management strategy. In the present research, we systematically investigated how three distinct organic substitution treatments—application of commercial organic manure (NC), traditional farmyard manure (NF), and a bio-organic manure inoculated with Trichoderma viride Pers. (NT)—influence gross nitrogen transformation processes and the pathways of nitrous oxide (N2O) production in Nicotiana tabacum L. (tobacco) fields, using plots receiving only SN as the reference treatment. Application of organic substitution treatments markedly enhanced soil nitrogen transformation processes. Relative to SN, these treatments stimulated nitrate (NO3−) production (6.43–7.43 mg N kg−1 soil d−1), increased ammonium (NH4+) production (1.01–1.37 mg N kg−1 soil d−1), and elevated NH4+ consumption (6.50–6.96 mg N kg−1 soil d−1). Autotrophic nitrification dominated NO3− production and NH4+ consumption, and was markedly enhanced under NF and NT treatments. Although organic substitution treatments increased nitrogen transformations and thereby reduced N2O emissions from autotrophic nitrification, autotrophic nitrification remained the dominant source of N2O. By modulating nitrogen transformation processes, organic substitution treatments significantly reduced overall N2O emissions, providing a basis for sustainable agricultural development and greenhouse gas mitigation.
-
Key words:
- Nicotiana /
- Organic manure /
- Gross nitrogen transformation /
- Nitrous oxide