









-
Angelica sinensis (Oliv.) Diels is a perennial herbaceous plant of the Apiaceae family. Its dried root is commonly used in Chinese herbal medicine, rich in bioactive compounds such as organic acids, polysaccharides, and essential oils[1,2]. It is well recognized for its functions of enriching blood, promoting blood circulation, regulating menstruation, relieving pain, and moistening the intestines. It is considered a 'sacred herb for blood disorders', and a 'sacred herb for gynecological conditions', in folk medicine[3]. A. sinensis thrives in cool, shady, and humid environments at altitudes of 2,200–3,000 m and is intolerant to high temperatures. Its production is concentrated in southeastern Gansu Province, which is the main production area of the Taoist medicinal herbs[4], as well as in Yunnan, Sichuan, Hubei, and Shanxi Provinces. Premature bolting in A. sinensis, a common root medicinal plant in the Apiaceae family, leads to increased root lignification and the decomposition of numerous bioactive components, which eventually reduce the yield and quality of the medicinal material [5,6].
NAC transcription factors (TFs) play a key role in the biosynthesis of lignin in plants. They regulate lignin biosynthesis and deposition by directly or indirectly modulating the expression of downstream target genes, including essential enzymes in the phenylpropanoid pathway. Additionally, NAC TFs are involved in cell wall formation, secondary growth, and stress responses[7]. For example, in some plants, NAC TFs directly activate key lignin biosynthesis genes (e.g., PAL, C4H, 4CL, CCR, and CAD). In Capsicum annuum, CaNAC76 selectively binds to the promoter of CaCAD1 to activate its transcription, thereby increasing lignin content and enhancing cold tolerance[8]. In rice, OsNAC28 directly activates CAD8B and positively regulates lignin accumulation[9], while OsNAC5 promotes lignin deposition by activating OsCCR10, thereby enhancing drought tolerance[10]. In the medicinal plant Angelica dahurica, recent studies have found that AdNAC20 has a unique dual regulatory function. It not only regulates the biosynthesis of lignin, but also negatively regulates the accumulation of coumarin, a medicinal component. Transcriptome analysis predicts that AdNAC20 can directly bind to the promoters of target genes containing SNBE-like motifs, such as MYB46, C3H, and CCoAOMT, which all have SNBE-like motifs, to regulate their transcription expression. This mechanism contributes to balancing lignification and secondary metabolism during A. dahurica premature bolting process[11]. Therefore, modulating NAC gene expression offers a promising approach for precisely controlling lignin content and composition, facilitating the breeding of stress-resistant varieties, and enhancing the potential applications of plants in bioenergy, papermaking, and agricultural quality improvement.
NAC TFs are known for their involvement in various biological processes, including plant growth, development, stress responses, and as metabolic pathways[12,13]. A notable number of research studies on NAC TFs have been reported in A. thaliana[14], Populus tomentosa[15], Camellia sinensis[16], Triticum aestivum[17], and O. sativa[18], as well as in medicinal plants such as Panax notoginseng[19], Eleutherococcus senticosus[20], and Panax ginseng[21]. However, the role of NAC TFs in regulating lignin biosynthesis in A. sinensis is still ambiguous. In this study, we identified the NAC TFs family in A. sinensis and determined the key candidate gene AsNAC043 through phylogenetic analysis. Additionally, we constructed an AsNAC043 overexpression vector and performed functional validation using a native callus transformation system in A. sinensis. Our objective was to investigate the effect of the AsNAC043 TF on lignin accumulation in A. sinensis roots, and to provide a foundation for elucidating the molecular regulatory mechanism of root lignification in this species. The findings of this study will enhance the theoretical understanding of secondary metabolism regulation in medicinal plants, and provide new strategies and targets for their genetic improvement.
-
A. sinensis plants were cultivated in She Block Township, Dongchuan District, Kunming City, Yunnan Province (latitude 26°15′7″ N, longitude 102°57′15″ E, altitude 3,254.46 m). The specimens were identified as A. sinensis (Oliv.) Diels (Apiaceae) by researcher Wenguang Yang (Kunming Institute of Botany, Chinese Academy of Sciences), and deposited at the Kunming Institute of Botany (voucher no. 20231019). The roots of A. sinensis bolting plants (BP) and unbolting plants (UBP) were collected in September 2022, quick-frozen in liquid nitrogen, and stored at −80 °C for spare use. Nicotiana benthamiana seeds were placed in sterile water and cold treated at 4 °C for 12 h before being sown into nutrient soil. Cultivation was accomplished in a greenhouse maintained at 23 °C. Once seedlings had develop four true leaves, they were transplanted with soil into pre-dug holes, and cultivation was continued in the 23 °C greenhouse.
Escherichia coli DH5α competent cells were purchased from Sangon Biotech (Shanghai) Co., Ltd. Agrobacterium tumefaciens: GV3101 competent cells were purchased from Beijing TransGen Biotech Co., Ltd. Plants expression vector pCAMBIA1300-GFP was maintained in our laboratory for future use.
Callus induction of A. sinensis
-
Sterile seedlings of A. sinensis were inoculated onto 1/2 MS medium (pH = 5.8) supplemented with 30 g/L sucrose, 4 g/L agar, 2 mg/L 3-indolebutyric acid (IBA), 0.2 mg/L kinetin (KT), and 0.5 g/L activated carbon to induce callus formation. The resulting callus tissue was then transferred to 1/2 MS medium (pH = 5.8) containing 20 g/L sucrose, 8 g/L agar, 2 mg/L IBA, and 0.5 g/L activated carbon to promote shoot formation. The induction of aseptic seedlings, callus, and shoot calli were all carried out in a culture room at 17 °C.
Identification of NAC family members and construction of expression profiling
-
Genome and annotation files of A. sinensis were retrieved from the NCBI database[22]. Transcriptome sequencing data were assembled based on the reference genome. The Pfam ID PF02365 for the NAC domain was downloaded from the UniProt database, and the corresponding Pfam feature file was obtained from Pfam (
http://pfam.xfam.org/search ). Candidate sequences containing the NAC domain were extracted using the 'HMM Search' tool in TBtools-II v2.154, with an E-value cutoff of < 1 × 10−5. in the domain scores. The CDS and protein sequences of NAC members were extracted using the 'Fasta Extract' tool in TBtools. Redundant or incomplete sequences were manually removed, retaining only those with complete NAC domains. Known functional NAC sequences from A. thaliana were downloaded from the TAIR database (www.arabidopsis.org/ ), and a phylogenetic tree was constructed using the neighbor-joining method. A heatmap was generated based on the normalized TPM values of 12 candidate NAC genes using the 'Heatmap' tool in TBtools.Physicochemical properties and chromosomal localization analysis of the AsNAC gene family
-
TBtools was employed to explore the physicochemical properties of the proteins encoded by the A. sinensis NAC gene family, along with amino acid number, molecular weight, theoretical pI, instability index, aliphatic index, and grand average of hydropathicity (GRAVY). Likewise, the WoLF PSORT online tool was used to predict subcellular localization. Chromosomal localization information for the AsNAC genes was obtained from the GFF3 genome annotation file in the NCBI database, which was then displayed using TBtools[23,24].
Analysis of conserved motifs, domains, and gene structure
-
Conserved motifs in the AsNAC protein sequences were identified using the MEME online tool (
https://meme-suite.org/meme/tools/meme ), with the number of motifs set to 10, and all other parameters set as default values (where the basic sequence width ranges from 6 to 50). Domain information obtained from the CDD database was integrated with motif data. A phylogenetic tree was constructed using the neighbor-joining method in MEGAX64. TBtools was used to visualize the exon-intron structures, conserved motif distributions, and domain architectures of the AsNAC proteins[25].RNA extraction and cloning of the AsNAC043 gene
-
Root tissue samples were collected from BP and UBP of A. sinensis. The tissue was ground in liquid nitrogen, and total RNA was extracted using the HiPure Plant RNA Mini Kit (Guangzhou Meiji Biotechnology Co., Ltd). RNA quality was assessed according to the manufacturer's instructions, and high-quality RNA samples were selected for subsequent experiments. cDNA was synthesized from the selected RNA using Hifair® AdvanceFast 1st Strand cDNA Synthesis Kit and stored at −20 °C. Based on the AsNAC043 gene sequence, specific gene cloning primers were designed (Supplementary Table S1). The coding sequence (CDS) of the AsNAC043 gene was cloned and analyzed by agarose gel electrophoresis. Results indicated that the cloned AsNAC043 gene ranged between 1,000 bp and 1,500 bp in length, consistent with the expected size. Subsequently, the gel was recovered (Supplementary Fig. S1).
Construction of plant expression vectors.
-
The expression vector pCAMBIA1300 was digested by the restriction enzyme BamHI. The digested plasmid was purified using the omega E.Z.N.A.® Cycle-Pure Kit column purification system. Under the catalysis of seamless cloning enzymes, the target gene was connected to the digested plasmid at 50 °C for 50 min. After the reaction, the recombinant product was transformed into the competent cells of E. coli DH5α. The activated cells were inoculated on LB solid medium containing 100 mg/L kanamycin, and cultured at 37 °C for 12−14 h. Choosing a colony for PCR verification, the PCR-positive colonies were placed in LB liquid medium containing kanamycin and cultured overnight at 37 °C and 200 rpm. Plasmids were extracted and sequenced. Finally, according to the sequencing results, it was determined whether the target gene has been successfully integrated into the vector.
Confocal laser scanning microscope (CLSM) analysis
-
The pC1300-GFP empty vector stored in the laboratory, and the pC1300-AsNAC043-GFP recombinant plasmid successfully sequenced, were transfected into competent cells of A. tumefaciens GV3101, which was then incubated at 30 °C and 200 rpm for 2−3 h. The activated bacteria was spread onto an LB solid medium containing 100 mg/L kanamycin and incubated at 30 °C for 48 h. A positive colony was chosen and inoculated into a 6 mL LB liquid culture medium containing kanamycin, and cultured at 30 °C and 200 rpm for 24 h. Then, 700 μL of the bacterial suspension was mixed with 700 μL of 50% glycerol and stored at −80 °C. The remaining culture was transferred to 30 mL LB medium containing kanamycin and amplified to an OD600, reaching 0.6−0.8. The bacterial cells were centrifuged at 1,000 rpm for 5 min, and the supernatant was discarded. The pellet was resuspended in a solution containing 10 mM MES, 1 mM MgCl2, and 150 μM acetosyringone (AS), and incubated for 0.5−3 h. The bacterial suspension was then gently injected into the lower epidermis of tobacco leaves using a needle without a tip. After injection, the tobacco plants were kept in the dark for 12−15 h, then transferred to suitable environmental conditions for 24 h[26]. The lower epidermis of the injected leaves was peeled off, placed on a slide, flattened, and stained with the nuclear dye DAPI. Fluorescence imaging was performed using a laser confocal microscope.
Agrobacterium tumefaciens-mediated stable transformation of A. sinensis callus
-
Ten microliter of A. tumefaciens stored in a −80 °C freezer and inoculate it into 6 mL of LB liquid medium containing kanamycin. It was then cultured at 30 °C and 200 rpm for 24 h, then transfer to 30 mL of LB medium containing kanamycin for culture expansion. When the OD600 reached 0.3−0.4, the bacterial cells were collected by centrifugation. Bacterial cells were collected by centrifugation and resuspended in an MS medium containing 100 μmol/L acetosyringone. After dark incubation for 30 min, 20-d-old A. sinensis callus was immersed in the bacterial suspension for 8−10 min, blotted dry on filter paper, and transferred to MS co-culture medium (pH = 5.8), containing 15 g/L sucrose, 7 g/L agar, 0.5 g/L activated carbon, and 100 μmol/L AS. After 2 d of dark co-culture, the callus was transferred to a selection medium consisting of 1/2 MS medium (pH = 5.8), supplemented with 20 g/L sucrose, 8 g/L agar, 2 mg/L IBA, 0.5 g/L activated carbon, 25 mg/L kanamycin, and 270 mg/L Timentin[27,28]. The medium was replaced weekly for 2 to 3 weeks until clear differentiation and growth of callus tissue were observed. Positive transformants were identified by PCR.
RT-qPCR analysis
-
Total RNA was extracted from positive A. sinensis callus tissue and reverse-transcribed into cDNA using the PrimeScript™ RT Reagent Kit with gDNA Eraser (RR047A, Takara). The cDNA was stored at −20 °C. RT-qPCR was performed using Novazene's AceQ qPCR SYBR Green Master Mix according to the manufacturer's instructions. Primers are listed in Supplementary Table S1. The AsEEF1G gene was used as an internal reference[29]. The reaction system was prepared according to the F488 SYBR qPCR Mix manual, with the following cycling program: 95 °C for 30 s, 40 cycles of 95 °C for 15 s, and 60 °C for 20 s; followed by 95 °C for 15 s, 60 °C for 1 min, and 95 °C for 15 s. Three biological replicates and four technical replicates were performed, excluding outliers. Relative gene expression was calculated using the 2−ΔΔCᴛ method.
Determination of lignin content
-
Comin lignin content determination kit (Cat No. MZS-2-G, Suzhou Keming Biotechnology Co., Ltd, China) was used to determine the lignin content in plant samples according to the kit instructions[30]. A 4:1 (v/v) mixture of acetic acid/acetyl bromide was mixed with approximately 5 mg of dry samples, thereafter incubated at 80 °C for 40 min, shaken every 10 min, and allowed to cool at room temperature. A 1:1 (v/v) mixture of 2 M NaOH/acetic acid was added to stop the reaction. A spectrophotometer (Hitachi SU1000) was employed to measure the extracted lignin from the supernatant of each sample at 280 nm. A standard curve (y = 0.0347x + 0.0068, R2 = 0.9889) was used to calculate the total lignin content. The calculation was performed with the formula: lignin (mg/g) = (0.0294 × [ΔA −0.0068])/W × T, where W represents the sample weight (g), T represents the dilution ratio, and ΔA represents the absorbance value obtained by applying the spectrophotometric values using the standard curve. The lignin percentage content (%) is then calculated as: lignin percentage (%) = (lignin content [mg/g]/1,000) × 100%, a unit conversion factor of 1,000 is applied (since 1 g = 1,000 mg)[31].
RNA-seq and DEGs analysis
-
Raw reads were processed with fastp (v0.18.0)[32] to remove adapter sequences, reads with > 10% unknown nucleotides (N), and reads with > 50% low-quality bases (Q ≤ 20). Clean reads were aligned to the reference genome using HISAT2.2.4[33] with default parameters. Gene annotation was performed by aligning sequences to the KEGG (
www.genome.jp/kegg/ ), NR, GO, and KOG databases using BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi ). Differentially expressed genes (DEGs) analysis was conducted with DESeq2, with DEGs defined as those with FDR < 0.05, and |log2 (fold change)| ≥ 2. Expression levels were quantified as TPM values using RSEM.Statistical analysis
-
This study employed a one-factor experimental design with three independent biological replicates per treatment group. Data are presented as mean ± standard deviation (SD), and one-way analysis of variance (ANOVA) was performed using GraphPad Prism 10.3 software to test for intergroup differences[34].
-
A total of 122 AsNAC family members were identified from the reference genome of A. sinensis. Sequence analysis indicated that the CDS lengths ranged from 351 to 3,195 bp, encoding proteins of 116 to 1,064 amino acids. Analysis of physicochemical properties using TBtools revealed that the molecular weights of the AsNAC proteins ranged from 13.6258 to 120.73788 kDa, and their theoretical isoelectric points varied from 4.35 to 9.96, indicating that this protein family exhibits both basic and acidic characteristics, with most members being predominantly acidic proteins (Supplementary Table S2). Using WoLF PSORT, subcellular localization predictions showed that 72.13% (88 out of 122) of AsNAC genes are localized in the nucleus, indicating a predominant role in regulating plant growth and development. The remaining members were distributed in the cytoplasm, chloroplasts, and peroxisomes. Chromosomal localization analysis revealed that the 122 AsNAC genes are distributed across all 11 chromosomes, with two additional genes located in scaffold regions (Supplementary Fig. S2).
Conserved motifs, domains, and gene structures of A. sinensis NACs
-
TBtools software was used to analyze conserved motifs, domains, and gene structures of 122 A. sinensis NAC proteins. Ten conserved motifs, designated motif 1 to motif 10, and consisting of 11 to 50 residues were identified (Supplementary Figs S3a, S4). Motif 6 was the most frequent and represents the core conserved motif of the family, whereas motif 9 was the least common. The composition and arrangement of motifs varied considerably among different evolutionary clades, although members within the same clade generally exhibited similar motif patterns, implying functional conservation. All 122 AsNAC proteins contained the typical no-apical-meristem (NAM) domain, with AS06G02092 additionally harboring a CRS1-Yhby domain (Supplementary Fig. S3b). Gene structure analysis showed that AsNAC genes contain 0−3 introns, and 1−9 exons (Supplementary Fig. S3c). Overall, genes within the same evolutionary branch displayed similar structural features, supporting the evolutionary conservation of the AsNAC family in A. sinensis.
Phylogenetic and expression pattern analysis of A. sinensis NACs
-
To investigate the phylogenetic relationships of the AsNAC family, a phylogenetic tree was constructed using 122 A. sinensis and 105 A. thaliana NAC protein sequences (Fig. 1a). The AsNAC members were classified into 18 subfamilies, all of which contained representatives from both species. In the OsNAC7 subfamily, genes such as AS02G00175, AS01G01899, and AS05G01615 clustered with A. thaliana ANAC043 and ANAC012. In the NAP subfamily, AS10G00848 and ANAC029 formed a distinct branch. This clustering pattern suggests that NAC proteins from the two species with high sequence similarity may perform analogous functions. Given that A. thaliana ANAC043 (in the OsNAC7 subfamily) is involved in lignin biosynthesis, we hypothesized that its orthologs in A. sinensis, including AS02G00175, may also regulate lignin synthesis. Expression analysis of 12 AsNAC genes from the OsNAC7 subfamily revealed that AS02G00175 was most significantly upregulated in BP[31] (Fig. 1b). Therefore, we have renamed AS02G00175 to AsNAC043 as a candidate gene for subsequent functional validation.
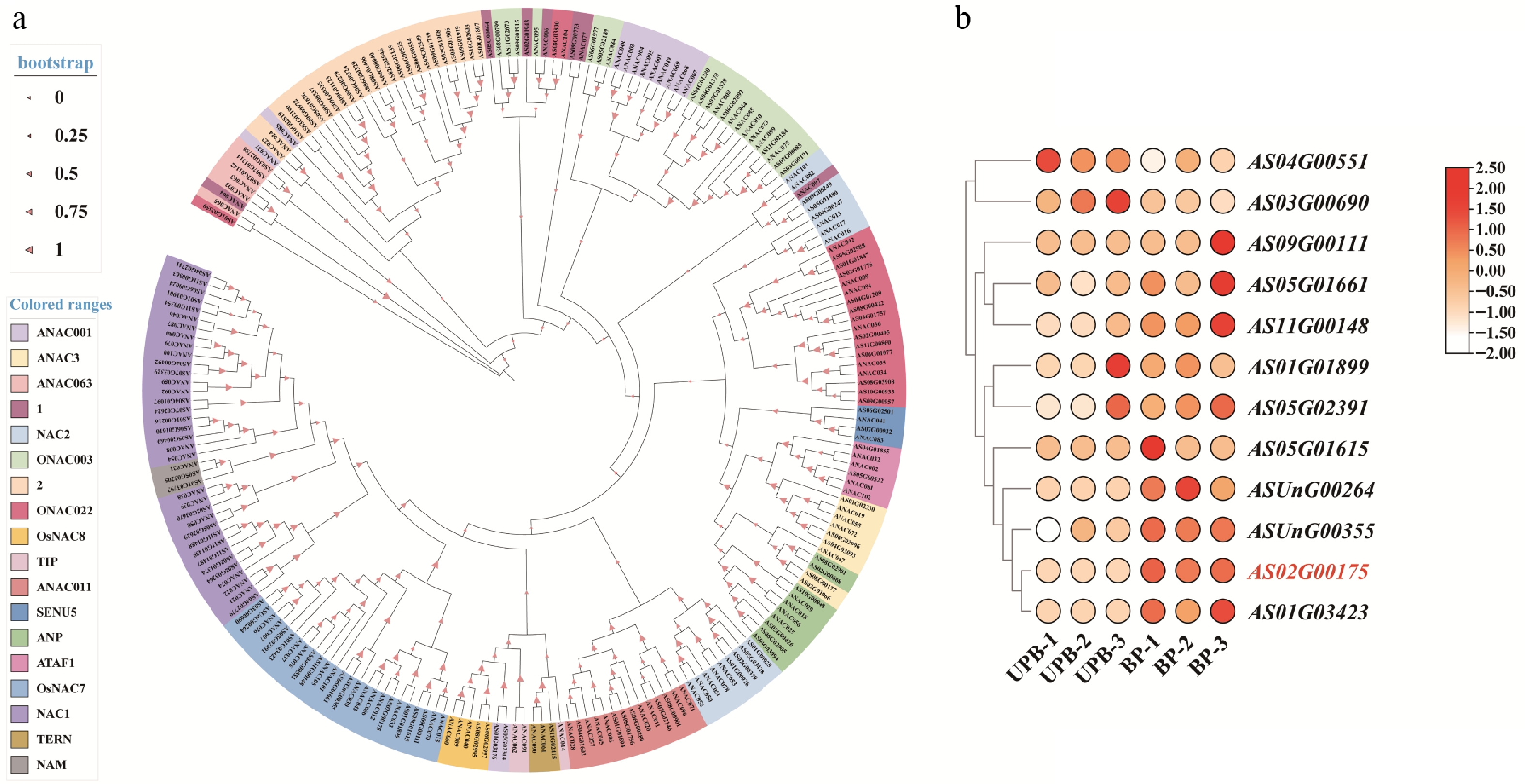
Figure 1.
Phylogenetic and expression analysis of NAC TFs in A. sinensis. (a) Phylogenetic tree of A. sinensis NAC TFs. (b) Expression pattern of OsNAC7 subfamily.
Cloning and subcellular localization of AsNAC043
-
In this study, the DNA sequence of AsNAC043 was cloned from A. sinensis. The full length of the sequence is 1,266 bp, encoding 422 amino acids, including five subdomains (A−E) at the N- terminal (Supplementary Fig. S5).
To further elucidate the subcellular localization of the AsNAC043, we used A. tumefaciens transient transformation. A. tumefaciens carrying the empty vector pC1300-GFP, and the fusion expression vector pC1300-AsNAC043-GFP were injected into N. benthamiana leaves, respectively, to realize the instantaneous expression of the target gene. Confocal imaging showed that GFP signals from the control were distributed throughout the cell, including the membrane and nucleus. In contrast, the AsNAC043-GFP fusion protein was exclusively localized in the nucleus (Fig. 2), consistent with its predicted function as a TF.
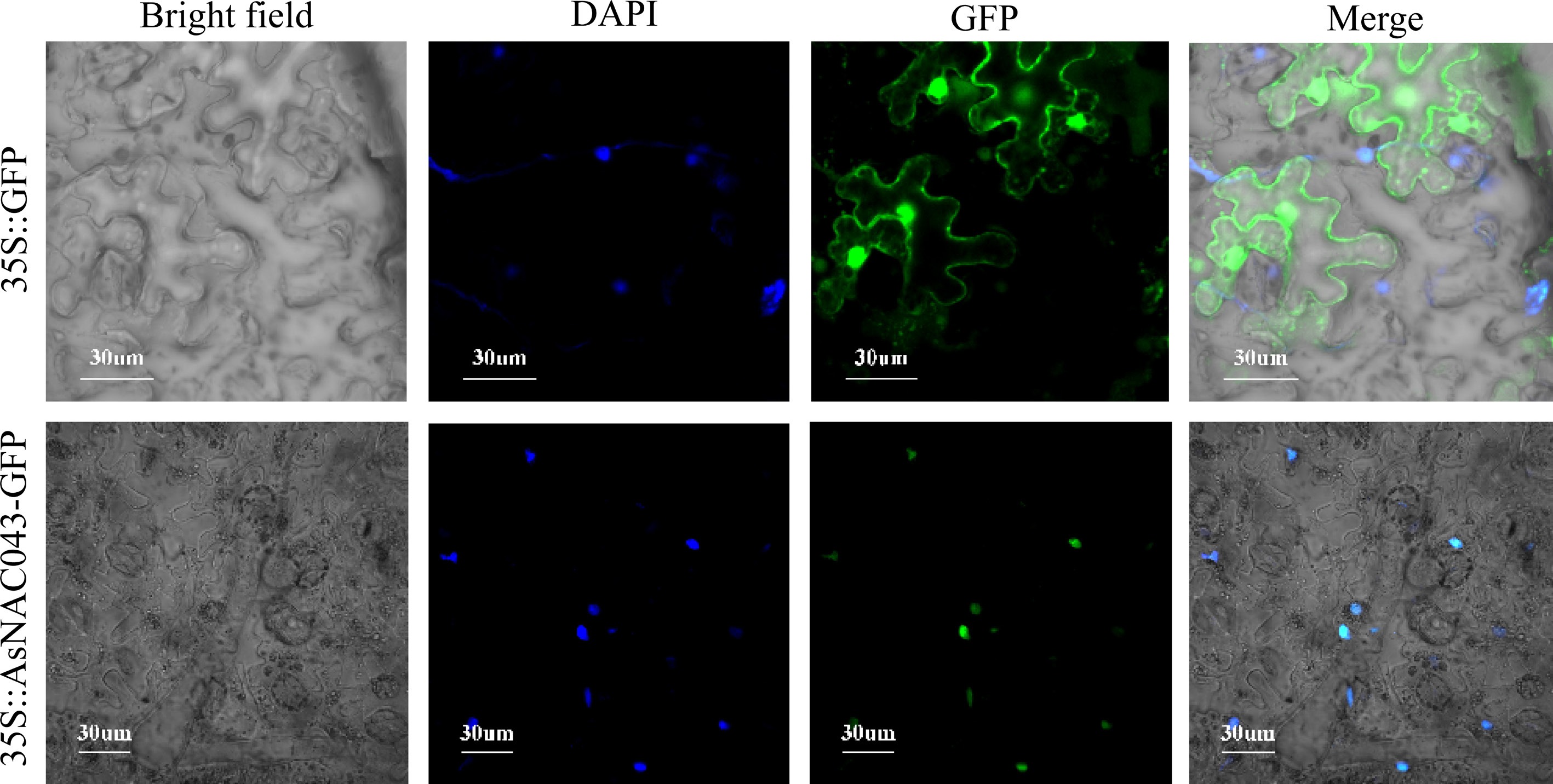
Figure 2.
Subcellular localization of AsNAC043 in lower epidermal cells of N. benthamiana leaves.
Functional characterization of AsNAC043 gene in A. sinensis
-
The AsNAC043 gene fragment was connected to the pC1300 vector and transformed. PCR analysis of the single colony confirmed that AsNAC043 was successfully recombined with the pC1300 vector. (Supplementary Fig. S6). The recombinant plasmid was introduced into A. tumefaciens GV3101 and used to transform A. sinensis callus.
After 3 to 4 weeks of subculture, transgenic lines (OE1, OE2, OE3) showed slower growth compared to wild-type (WT) plants. RT-qPCR analysis confirmed that AsNAC043 expression was significantly increased in the OE lines (Fig. 3a, b). Given the potential role of AsNAC043 in lignin biosynthesis, total lignin content was measured. UV spectrophotometric analysis revealed a significant increase in lignin content in the AsNAC043-overexpressing callus tissues compared to the WT (Fig. 3c).
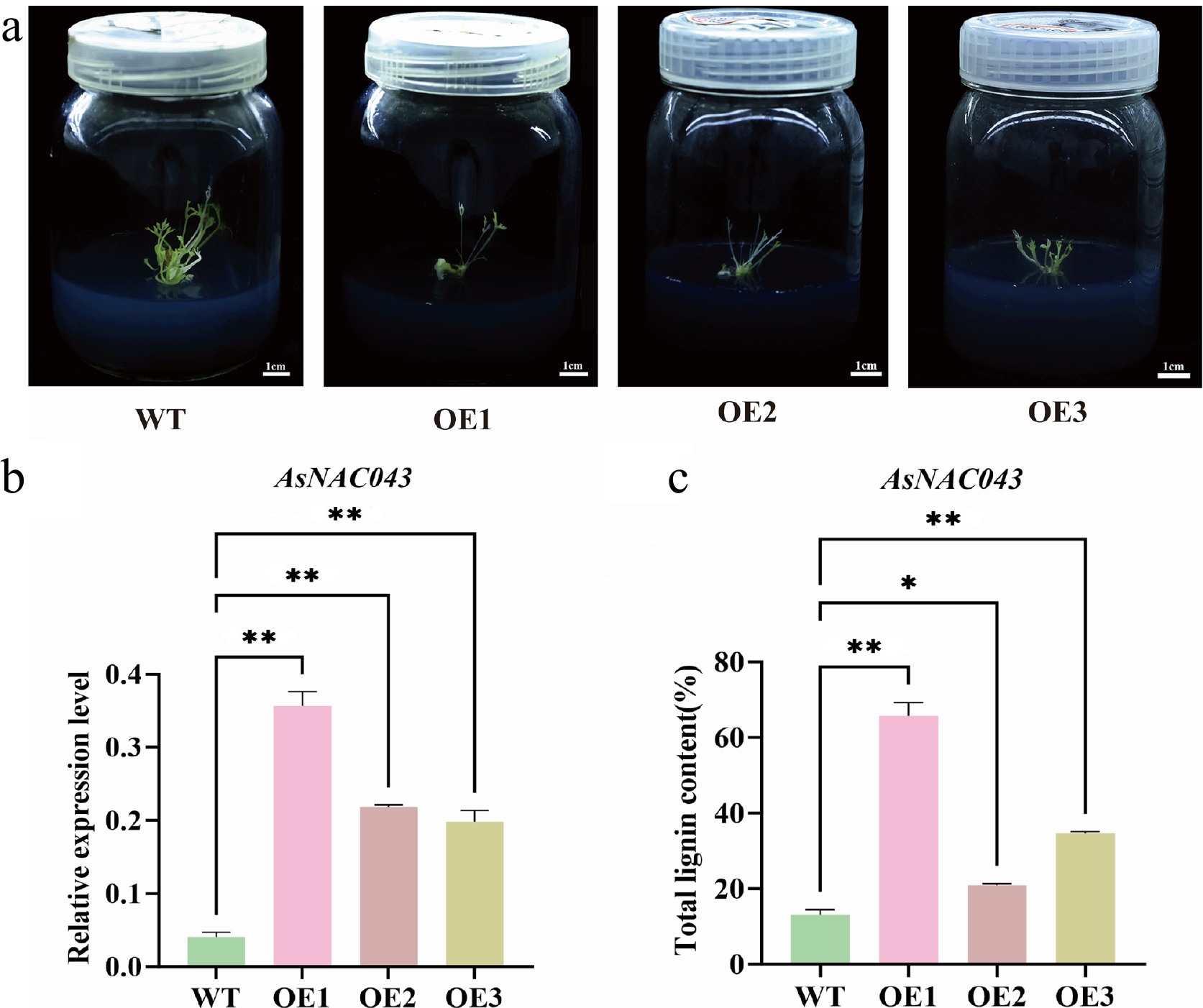
Figure 3.
Characterization of WT and AsNAC043-overexpressing transgenic lines (AsNAC043-OE). (a) Phenotypes of WT and AsNAC043-OE in A. sinensis plants. OE1, OE2, and OE3 represent independent transgenic lines. (b) RT-qPCR analysis of AsNAC043 expression in WT and transgenic lines (OE1, OE2, OE3). (c) Lignin content in WT and transgenic plants. An asterisk indicates a significant difference (* p < 0.05; ** p < 0.01).
RNA-seq analysis of AsNAC043-overexpressing plants
-
To explore the global transcriptional changes induced by AsNAC043 overexpression, RNA-seq was performed on WT and OE callus tissues with three biological replicates per group. Principal component analysis (PCA) clearly separated the WT and OE samples, and correlation heatmaps indicated high reproducibility among replicates (Fig. 4a, b). A total of 6,885 DEGs were identified, including 2,594 upregulated and 4,291 downregulated genes (Fig. 4c, d, Supplementary Table S3). GO enrichment analysis assigned the DEGs in terms of biological process, molecular function, and cellular component categories. KEGG pathway analysis indicated that the DEGs were predominantly enriched in pathways related to secondary metabolite biosynthesis, phenylpropanoid biosynthesis, metabolic pathways, and flavonoid biosynthesis (Fig. 4e, f).
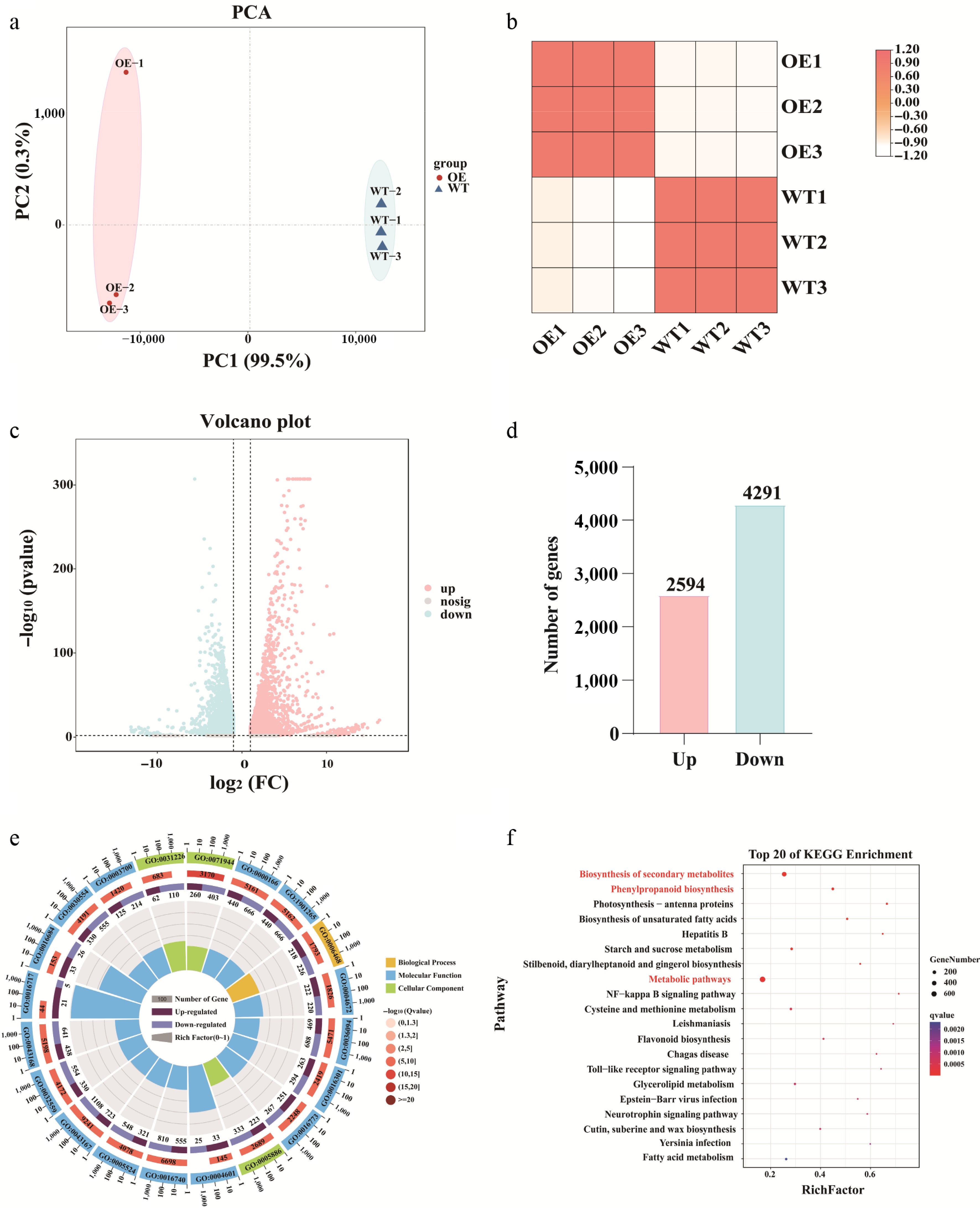
Figure 4.
Differential gene expression and enrichment analysis of WT and AsNAC043-overexpressing transgenic lines (AsNAC043-OE). (a) Principal component analysis (PCA). (b) Sample correlation analysis. (c) Volcano plot of differential expressed genes. (d) Statistics of differentially expressed genes. (e) GO functional enrichment analysis. (f) KEGG pathway enrichment analysis.
Effects of AsNAC043 overexpression on lignin synthesis-related genes and protein interaction networks
-
Transcriptome data were used to analyze the expression of AsNAC043 and key lignin biosynthesis genes. Results indicate that the expression level of the AsNAC043 gene is significantly upregulated in the OE line, as well as key lignin pathway genes such as AsCCR (AS07G02794), and AsCADs (AS01G01636, AS01G00171, AS06G03766). Phenylpropanoid pathway genes, including AsPALs (AS06G01386, AS05G03296), and As4CLs (AS06G02531, AS07G02833), were also upregulated. In contrast, genes involved in ferulic acid biosynthesis (AsCOMT, AS01G03183), and flavonoid synthesis (AsCHI, AS03G00525) were downregulated. Among TFs, AsMYB (AS09G01066) was upregulated, while AsMYB (AS10G02150) and AsF5H (AS04G00561) were downregulated (Fig. 5a).
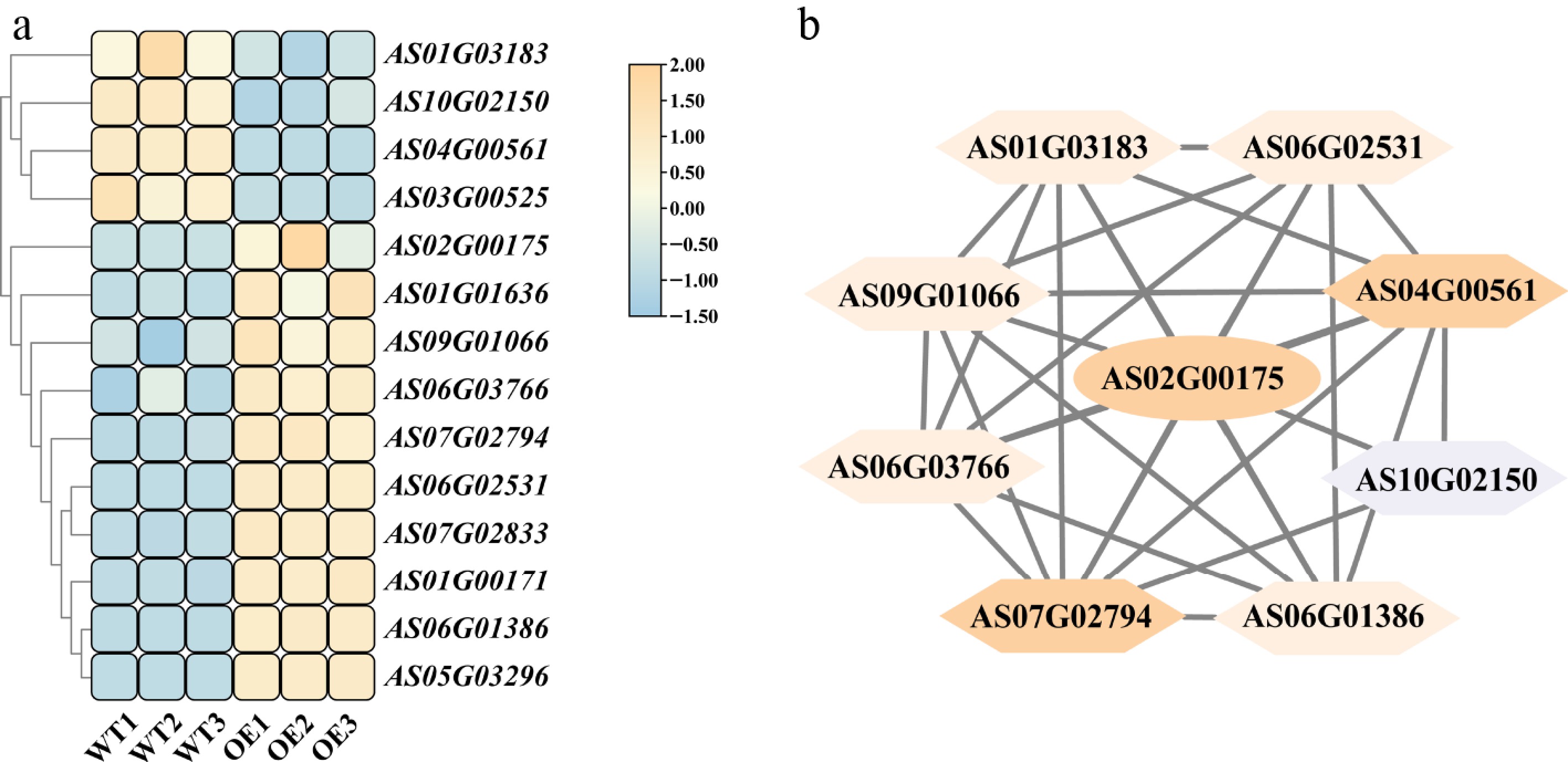
Figure 5.
Effects of AsNAC043 overexpression on the expression of lignin biosynthesis-related genes and protein interaction network analysis. (a) Heatmap of key lignin biosynthesis gene expression in the WT and AsNAC043-OE lines. (b) Analysis of the Interaction network among proteins encoded by key genes in lignin biosynthesis. AsNAC043 (AS02G00175); AsPALs (AS06G01386, AS05G03296); AsCADs (AS01G00171, AS06G03766, AS01G01636); As4CLs (AS06G02531, AS07G02833); AsCCR (AS07G02794); AsCOMT (AS01G03183); AsCHI (AS03G00525); AsF5H (AS04G00561), and AsMYBs (AS09G01066, AS10G02150).
Protein interaction network analysis revealed that MYB TFs (AS09G01066, AS10G02150) interact with key lignin biosynthesis genes such as CCR, CADs, COMT, 4CL, PAL, and F5H. AsNAC043 also showed strong associations with AsMYBs, AsF5H, and AsCCR, suggesting close regulatory relationships and potential synergy in the regulation of lignin biosynthesis (Fig. 5b).
-
Recently, with the expanding market demand for root and rhizome medicinal ingredients, many plants are prone to premature bolting and flowering during cultivation due to germplasm degradation, nutrient accumulation, and external environmental influences. Since the medicinal properties of these plants are predominantly found in their roots or rhizomes, premature flowering disrupts nutrient allocation, accelerates root lignification, and severely compromises both the quantity and quality of the crude drug[6]. Previous studies have demonstrated a close association between TFs and lignin biosynthesis in plants (Supplementary Table S4). For instance, a NAC TF (LOC107435239) was identified in Ziziphus jujuba Mill that upregulates F5H expression, thereby promoting lignin accumulation[35]. Lignification in Pyrus fruit is regulated by PpNAC187, where the overexpression of PpNAC187 upregulates CCR and COMT gene expression, thereby increasing lignin content[36]. Overexpression of OsNAC17 in O. sativa induces lignin-related gene expression, promotes lignin accumulation in leaves and roots, and enhances drought tolerance[37]. Similarly, in Eucalyptus grandis[38] and Apium graveolens[39], heterologous or homologous overexpression of NAC genes effectively activates the lignin synthesis pathway, thereby increasing lignin accumulation levels in plants.
This study identified 122 members of the AsNAC family in A. sinensis. Subcellular localization predictions indicated that 72.13% of AsNAC proteins are nuclear-localized, aligning with the canonical role of NAC TFs in transcriptional regulation. The localization of a subset of AsNAC proteins to the cytoplasm, chloroplasts, and peroxisomes suggests potential roles in signal transduction or organelle-specific functions, possibly mediated by dynamic subcellular redistribution. Such diversity in localization has also been observed in NAC families of other plants, including Miscanthus sinensis[40], highlighting the functional complexity of this gene family. The 122 AsNAC genes were predominantly distributed across 11 chromosomes, with two genes located in scaffold regions. This distribution may reflect evolutionary occurrences such as segmental or tandem duplications, which could potentially lead to functional diversification. All AsNAC proteins possess the conserved NAM domain[41,42], thereby validating the reliability of our identification. The presence of an additional CRS1-Yhby domain in some members may confer specialized functions, warranting further investigation.
To further identify key NAC members involved in lignification, we performed phylogenetic and expression analysis and identified AsNAC043, an ortholog of A. thaliana ANAC043[43]. Using A. tumefaciens-mediated transformation, we successfully overexpressed AsNAC043 in A. sinensis callus. RT-qPCR confirmed significantly increased AsNAC043 expression in transgenic lines (OE1, OE2, OE3) compared to WT, establishing a reliable system for functional studies. Lignin content assays revealed a significant elevation in total lignin within AsNAC043-OE calli, indicating that AsNAC043 positively regulates lignin biosynthesis. Protein interaction network analysis revealed that AsNAC043 not only interacts with genes associated with lignin biosynthesis, but also exhibits particularly strong associations with MYB TFs. Previous studies have demonstrated that Pyrus WRKY46 can directly activate the transcription of NAC187, thus promoting the expression of the lignin biosynthesis gene CCR and enhancing lignin accumulation[44]. It is also found that PbNAC47 forms complexes with PbAGL7 and PbMYB73, forming complexes that synergistically activate target genes and promote lignin biosynthesis and secondary wall thickening in stone cells[45]. Overexpression of CgNAC043 in Citrus grandis directly activates the promoters of CgMYB46, CgCCoAOMT, and CgC3H, upregulates CgMYB58 expression, and enhances lignin accumulation in vacuoles[43]. Therefore, it can be inferred that AsNAC043 may also form regulatory complexes with MYB TFs (e.g., AS06G02257, and AS04G00182) to synergistically enhance the expression of lignin-related genes and improve regulatory efficiency.
To elucidate the molecular mechanisms underlying AsNAC043-mediated lignin biosynthesis, we conducted RNA-seq analysis of WT and AsNAC043-OE callus. Principal component analysis clearly separated the WT and OE groups, revealing 6,885 DEGs, which signifies that AsNAC043 overexpression substantially alters the transcriptome. GO and KEGG enrichment analyses showed that DEGs were significantly enriched in pathways related to secondary metabolite biosynthesis, phenylpropanoid biosynthesis, metabolic pathways, and flavonoid biosynthesis, suggesting that AsNAC043 modulates lignin synthesis through these metabolic pathways. Specifically, the expression of key lignin biosynthesis genes AsCCR and AsCADs, along with phenylpropanoid pathway genes AsPALs and As4CLs, was significantly upregulated in OE lines. In contrast, genes involved in ferulic acid biosynthesis (AsCOMT) and flavonoid synthesis (AsCHI) were downregulated. This result indicates that AsNAC043 may positively regulate the phenylpropanoid pathway to redirect metabolic flux toward the lignin biosynthesis branch while simultaneously inhibiting the flavonoid biosynthesis pathway, a competing metabolic route. The downregulation of AsCOMT may be related to the stage-specific characteristics of lignin biosynthesis and the functional bias of metabolic branches.
Overall, AsNAC043 acts as a positive regulator of lignin biosynthesis in A. sinensis, likely through direct activation of lignin pathway genes, and suppression of alternative metabolic pathways. These insights enhance our understanding of the transcriptional network controlling lignin biosynthesis in A. sinensis, and establish a theoretical foundation for improving medicinal quality by genetic engineering. Future work should focus on validating direct target genes of AsNAC043 and exploring its potential roles in plant development and stress responses.
-
In conclusion, this study successfully identified and characterized the NAC TFs family in A. sinensis, with a particular focus on AsNAC043. Our findings demonstrate that AsNAC043 localizes to the nucleus and functions as a positive regulator of lignin biosynthesis. Overexpression of AsNAC043 in transgenic calli led to significantly increased lignin content and altered expression of key genes in the lignin biosynthesis pathway. Transcriptome analysis further revealed that AsNAC043 regulates multiple metabolic pathways, including phenylpropanoid biosynthesis, while repressing competing pathways such as flavonoid biosynthesis. These results not only provide important insights into the molecular mechanisms underlying lignin accumulation in A. sinensis, but also offer valuable genetic targets for future breeding strategies aimed at improving medicinal quality through reduced root lignification.
This work was supported by the National Natural Science Foundation of China (82460743), the Major Special Science and Technology Project of Yunnan Province (202502AS100012; 202403AK140082), Yunnan Characteristic Plant Extraction Laboratory (2022YKZY001), Yunnan Province Youth Talent Support Program (XDYC-QNRC-2022-0219), the National Key Research and Development Program of China (2023YFC3504800), and Program of Shanghai Academic/Technology Research Leader (23XD1423500).
-
The authors confirm their contributions to the paper as follows: experimentation, data analysis, mapping, manuscript drafting and revision: Zhang JJ, Xiang CF, Li LS, Wang J, He JD; revised the manuscript: Li MF, Faruque MO, Yang S; conceived and designed the experiments, and wrote and revised the manuscript: Xiao Y, Zhao Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
- Supplementary Table S1 All primers used in the experiments.
- Supplementary Table S2 Analysis of the physical and chemical properties of AsNACs family.
- Supplementary Table S3 Differentially expressed genes between and OE lines.
- Supplementary Table S4 Mechanisms of lignin regulation mediated by NAC in different plants.
- Supplementary Fig. S1 Gel electrophoresis of AsNAC043 gene amplification.
- Supplementary Fig. S2 Chromosomal localization of the A. sinensis AsNACs family.
- Supplementary Fig. S3 Phylogenetic relationships, conserved motifs, domains and gene structures of the A. sinensis AsNACs family.
- Supplementary Fig. S4 Visualization of 10 conserved motifs in the AsNAC family.
- Supplementary Fig. S5 Multiple alignment of amino acid sequences from AsNAC043.
- Supplementary Fig. S6 PCR detection for the construction of AsNAC043 overexpression vector.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang JJ, Xiang CF, Li LS, Wang J, He JD, et al. 2026. The NAC043 transcription factor from Angelica sinensis (Oliv.) Diels regulates lignin biosynthesis. Medicinal Plant Biology 5: e012 doi: 10.48130/mpb-0026-0005 

The NAC043 transcription factor from Angelica sinensis (Oliv.) Diels regulates lignin biosynthesis
- Received: 07 December 2025
- Revised: 02 February 2026
- Accepted: 11 March 2026
- Published online: 26 May 2026
Abstract: Angelica sinensis (Oliv.) Diels is a valuable medicinal plant whose dried roots are widely used in traditional Chinese medicine for blood enrichment and nourishment. NAC transcription factors (TFs) are known to play central roles in plant growth, secondary metabolism, and lignin biosynthesis. Here, we identified 122 AsNAC TFs from the A. sinensis genome. Phylogenetic analysis using NAC proteins from Arabidopsis thaliana, combined with expression profiling, identified AsNAC043 as a key candidate regulator. Subcellular localization confirmed that AsNAC043 localizes specifically to the nucleus, consistent with its function as a TF. We subsequently constructed an AsNAC043-overexpressing transgenic line in A. sinensis callus. Overexpression of AsNAC043 resulted in a significant increase in lignin content. Transcriptomic analysis further revealed upregulation of several key lignin biosynthesis genes, including AsCCR and AsCADs. Our results demonstrate that AsNAC043 may promote lignin accumulation in A. sinensis by activating the expression of core lignin biosynthesis genes. These findings provide insight into the molecular mechanism of AsNAC-mediated lignin biosynthesis, and offer valuable targets and theoretical support for breeding high-quality A. sinensis varieties.
-
Key words:
- Angelica sinensis /
- Lignin biosynthesis /
- Transcription factor /
- NAC