









-
Laccase (LACs, EC 1.10.3.2) is a type of multi-copper oxidase belonging to the blue copper protein superfamily. Its active centre binds four copper ions to form copper oxidase domains[1], enabling it to catalyse the oxidative degradation of various aromatic and non-aromatic compounds (such as phenols, lignin, anilines, thiols, and arylamines)[2]. During catalysis, copper ions absorb and transfer electrons, reducing oxygen to water and generating substrate-derived free radicals that drive the oxidation reaction[3]. Laccases are widely distributed in bacteria, fungi, insects, and plants[4]. They were first discovered by the Japanese scientist Yoshida in the lacquer tree (Rhus vernicifera) of the family Anacardiaceae in China[5]. The plant laccase gene family constitutes a medium-sized multigene family[6]. Its primary functions include catalysing lignin biosynthesis, secondary cell wall formation, and the polymerization of phenolic compounds, thereby not only regulating plant growth and development but also playing a key role in enhancing resistance to both biotic and abiotic stresses[7−9].
The plant laccase multigene family (LACs) has been identified in numerous species, including Arabidopsis thaliana[10], Citrus sinensis[11], Pyrus[12], Gossypium[13], Populus[14], and Oryza sativa[15], among others. Although the number and types of LAC gene family members vary considerably among these different plants, the family is evolutionarily conserved overall, sharing similar domains and conserved active reaction sites, and performing analogous functions. Most plant laccases are involved in the oxidation of monolignols, which contribute to the formation of higher-order lignin structures. In Arabidopsis thaliana, which contains 17 LAC family members, AtLAC2, AtLAC4, and AtLAC17 are believed to participate in lignin biosynthesis, thereby regulating plant growth and development[16−18]. In Brachypodium distachyon, the double knockout mutant of LAC8 and LAC5 results in a significant decrease in lignin content and a marked increase in saccharification efficiency, without compromising plant integrity[19]. In Populus deltoides, a laccase (PdLAC2) is associated with the oxidation of phenolic compounds and the binding of flavonoids that interact with lignin[20]. In cotton, the GhLAC15 gene enhances resistance to Verticillium wilt by increasing cell wall lignin content[21]. In rice (Oryza sativa), OsLAC10 enhances copper (Cu) stress tolerance in Arabidopsis, potentially by promoting root lignification to prevent excessive copper uptake, indicating a role in improving plant resistance to heavy metal stress[15]. In sweet orange, cold treatment induces differential regulation of 21 CsLACs, accompanied by increased laccase activity and lignin content in leaves; CsLACs can be induced by various hormones and abiotic stresses[22]. These findings collectively highlight that plant laccases are not merely structural enzymes, but are dynamic components of the plant's stress response machinery[14,23].
Despite this accumulation of knowledge in other species, the specific biological functions of laccases in mango (Mangifera indica L.), an economically important fruit tree of the Anacardiaceae family, remain largely unexplored. This research gap is particularly critical given the current environmental challenges in mango cultivation. Mango is predominantly grown in low-latitude tropical and subtropical regions, where it is frequently exposed to intense environmental stressors, most notably enhanced UV-B radiation (280−315 nm) resulting from stratospheric ozone depletion. Such elevated UV-B radiation can severely compromise plant cellular integrity, photosynthesis, and ultimately fruit quality and yield[24−26]. Therefore, unravelling the molecular mechanisms underlying mango adaptation to UV-B stress has become an important scientific question.
The theoretical foundation for the involvement of laccases in the UV-B response lies in the protective nature of the secondary metabolites they synthesize. Plant laccases catalyze the polymerization of lignin and phenolic compounds[17], which act as vital physiological screens by accumulating in the epidermal cell walls to absorb deleterious UV radiation[27]. Furthermore, these phenolic products are potent antioxidants capable of scavenging reactive oxygen species (ROS) induced by UV-B exposure, thereby preventing oxidative damage to cellular macromolecules[28,29]. Additionally, plant responses to abiotic stresses, including UV-B radiation, are regulated by endogenous phytohormones, particularly abscisic acid (ABA) and methyl jasmonate (MeJA)[30]. These hormones serve as central signalling molecules that rapidly accumulate under stress conditions and trigger the expression of downstream defense-related genes[31]. However, it remains unclear whether mango laccases participate directly in UV-B stress tolerance, and whether their activity is regulated through ABA- or MeJA-mediated signalling pathways.
To address these scientific questions, a comprehensive genome-wide analysis is necessary to understand how the MiLAC gene family responds to environmental radiation. In this study, we utilised genomic data from mango and Arabidopsis to identify MiLACs, subsequently analysed their physicochemical properties, phylogeny, chromosomal distribution, and promoter cis-acting elements. By integrating transcriptomic data with real-time quantitative polymerase chain reaction (RT-qPCR) validation, we aim to elucidate the expression patterns of MiLAC family members under enhanced UV-B radiation.
-
The experimental site is located at the Shengchang Village Mango Orchard, Haitang District, Sanya City, Hainan Province (18°25' N, 109°46' E). The average UV-B radiation intensity in this area is 83.47 kJ·m−2·d−1; the annual average precipitation is about 1,700 mm, and the annual average temperature is about 25 °C. The soil is red sandy loam. The 'Tainong No. 1' mango (Mangifera indica L. cv. Tainongyihao) trees are 17 years old and have naturally rounded canopies, planted at a spacing of 4 m × 5 m. The orchard and tree canopies have good ventilation and light penetration. The mango whole-genome sequence and corresponding annotation data were downloaded from the NCBI Datasets resource (
www.ncbi.nlm.nih.gov/datasets/taxonomy/29780 ). The Arabidopsis thaliana (L.) Heynh. laccase (AtLAC) gene family sequences were retrieved from the TAIR database (www.arabidopsis.org ).Experimental design
-
Previous findings from our research group indicated that artificially simulated enhanced UV-B radiation at 96 kJ·m−2·d−1, equivalent to a 15% increase over natural levels, significantly reduced per-plant yield and deteriorated fruit quality in 'Tainongyihao' mango[32]. Therefore, the experimental setup involved installing four 40 W UV-B lamps (λ = 313 nm) uniformly in a cross pattern 40 cm above the canopy to provide enhanced UV-B radiation in the treatment group. The control group received natural sunlight only. The experiment was arranged in a single-tree plot design with five replicates. Field trials were conducted over two consecutive fruit growth cycles: from November 16, 2023 (30 d after flowering) to January 6, 2024 (91 d after flowering), and from March 10, 2024 (30 d after flowering) to May 11, 2024 (92 d after flowering). During each growth cycle, fruit samples were collected at 30, 40, 50, 60, 72, 77, and 91 d after flowering in 2023, and at 30, 40, 51, 61, 71, 81, and 92 d after flowering in 2024 for subsequent physiological and biochemical analyses. The lamps were switched on daily from sunrise to sunset to provide irradiation. The enhanced UV-B treatment was suspended during overcast or rainy weather by turning off the lamps.
Sampling and sample pre-treatment
-
Five fruits of median size were selected from the middle section on each of the four sides of the canopy periphery of every sample tree as reference fruits for sampling. Fruit samples were collected once at the initiation of the field-enhanced UV-B radiation treatment, and subsequently at 10-d intervals. Sampling was performed according to the size and colour development of the reference fruit. In the field, the peel and pulp were rapidly separated, immediately flash-frozen in liquid nitrogen, transported back to the laboratory, and stored in a −80 °C ultra-low temperature freezer for subsequent use.
Identification of the LAC gene family in mango
-
Using the protein sequences of the AtLAC gene family members as a reference, a local BLAST analysis was performed against the mango protein database using TBtools software, yielding 54 protein sequences as candidate mango laccases (MiLACs). Hidden Markov models (HMMs) for the Cu-oxidase domain families (Cu-oxidase: PF00394, Cu-oxidase-2: PF07731, and Cu-oxidase-3: PF07732) were downloaded from InterPro (
www.ebi.ac.uk/interpro )[33] and used as queries to search the protein dataset using HMMER 3.0. Candidate genes were initially identified using the NCBI online CD-search tool (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). To ensure accurate identification, the candidate MiLAC genes were further validated using the SMART database (http://smart.embl-heidelberg.de ), and redundant sequences as well as proteins lacking the conserved domains were removed, yielding the final set of MiLAC gene family members.Protein characterisation of LAC genes
-
The physicochemical properties of MiLAC proteins—including amino acid length, molecular weight (MW), and theoretical isoelectric point (pI)—were predicted using the online tool ProtParam (
http://web.expasy.org/protparam ). Subcellular localization was predicted with WoLF PSORT (https://wolfpsort.hgc.jp ). Signal peptides were predicted using the SignalP 5.0 online server (https://services.healthtech.dtu.dk/service.php?SignalP-5.0 ).Gene structures and conserved motifs analysis
-
The filtered MiLAC protein sequences were submitted to the NCBI Batch Web CD-Search tool (
www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) to identify conserved domains. The corresponding protein sequences were then analysed using MEME (https://meme-suite.org/meme/tools/meme ) to detect conserved motifs, with the maximum number of motifs set to 10. All results were visualised using TBtools.Phylogenetic relationship of the LAC gene family in mango
-
A multiple sequence alignment was used to determine the evolutionary relationship among mango LAC genes and their homologs in A. thaliana using the MUSCLE algorithm, based on a total of 71 protein sequences (54 from Mangifera indica and 17 from Arabidopsis thaliana). Phylogenetic analysis was performed in MEGA 11 using the Maximum Likelihood (ML) method based on the identified protein sequences from different species, with bootstrap resampling set to 1,000 replicates to assess node support. The resulting phylogenetic tree was subsequently visualised using the online tool Chiplot (
https://chiplot.online ).Chromosomal mapping and Ka/Ks ratio evaluation
-
Intraspecies synteny analysis in mango was conducted using TBtools. Based on whole-genome annotation files, gene density across the genome and the chromosomal distribution of MiLAC genes were determined. Using the genome assembly (whole-genome sequence) file, chromosomal GC content and gap regions were calculated. The Ka and Ks substitution rates of homologous gene pairs were estimated using the natural gradient descent (NG) method, and the resulting Ka/Ks ratios were used to assess selective pressure.
Identification of cis-elements in the promoter region of LAC genes
-
Based on the mango gene family sequence information, the 2,000-bp upstream promoter regions of MiLACs genes were extracted using TBtools. These promoter sequences were then submitted to PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) for cis-acting regulatory element prediction, and the results were visualised with TBtools.Screening of differentially expressed genes and qPCR validation
-
Based on previously published transcriptome data from our group[34] (the mango transcriptome dataset is available in the NCBI database under accession number PRJNA1245353), two highly expressed MiLAC genes were selected as key targets for investigation during 2023–2024 (Table 1). Gene-specific qPCR primers were designed using the NCBI Primer-BLAST online tool (
www.ncbi.nlm.nih.gov/tools/primer-blast ; accessed December 1, 2024), and primer synthesis was performed by Shanghai Shengxin Biotechnology Co., Ltd. (Shanghai, China).Table 1. Basic information on MiLAC proteins.
Gene ID Gene name Number of
amino acidMolecular
weightpI Instability index Aliphatic index Grand average of
hydropathicitySignal
peptideSubcellular localization LOC123201611.1 MiLAC1 556 60,749.63 9.08 35.24 88.29 −0.041 YES chlo LOC123205456.1 MiLAC2 584 64,386.08 9.52 29.79 83.75 −0.076 YES chlo LOC123229717.1 MiLAC3 1147 126,257.25 5.19 34.44 80.51 −0.092 NO plas LOC123224994.1 MiLAC4 572 63,281.91 9.1 27.84 89.3 0.014 YES chlo LOC123209589.1 MiLAC5 578 64,417.75 8.48 36 78.58 −0.149 YES extr LOC123192083.1 MiLAC6 572 63,621.29 10.07 27.3 86.89 −0.091 YES chlo LOC123209528.1 MiLAC7 553 61,191.25 8.47 36.03 89.51 −0.056 YES vacu LOC123216306.1 MiLAC8 572 63,469.53 6.37 39.47 84.84 −0.043 YES chlo LOC123228608.1 MiLAC9 566 63,672.79 6.67 31.23 83.14 −0.127 YES plas LOC123229715.1 MiLAC10 574 64,038.58 6.59 33.01 76.74 −0.194 YES pero LOC123229713.1 MiLAC11 567 63,081.24 5.7 31.88 79.58 −0.146 YES vacu LOC123229041.1 MiLAC12 489 54,460.42 5.51 32.9 79.71 −0.18 NO cyto LOC123192537.1 MiLAC13 555 60,981.79 9.05 25.26 88.52 −0.045 YES extr LOC123198235.1 MiLAC14 568 63,437.73 6.37 34.2 77.2 −0.171 YES vacu LOC123229003.1 MiLAC15 561 62,323.26 6.07 33.51 82.51 −0.098 YES vacu LOC123201553.1 MiLAC16 557 60,936.06 9.09 33.74 90.09 −0.023 YES chlo LOC123228569.1 MiLAC17 582 65,830.37 5.57 38.25 78.83 −0.237 YES nucl LOC123206493.1 MiLAC18 568 63,338.6 6.37 35.12 77.2 −0.183 YES vacu LOC123229720.1 MiLAC19 568 63,373.65 6.54 33.88 76.51 −0.18 YES vacu LOC123203686.1 MiLAC20 561 62,591.07 6.43 35.67 76.45 −0.15 YES pero LOC123201619.1 MiLAC21 555 60,688.79 9.33 35.32 88.13 −0.055 YES chlo LOC123204379.1 MiLAC22 582 64,553.83 9.01 33.26 78.61 −0.161 YES chlo LOC123229716.1 MiLAC23 570 63,427.87 6.75 33.8 79.33 −0.175 YES chlo LOC123229257.1 MiLAC24 563 63,486.69 8.41 34.99 82.4 −0.168 YES chlo LOC123229178.1 MiLAC25 568 63,154.61 6.53 33.09 77.04 −0.177 YES chlo LOC123229004.1 MiLAC26 560 62,105.23 6.2 33.31 82.84 −0.088 YES vacu LOC123228609.1 MiLAC27 561 62,326.46 6.02 34.23 83.39 −0.077 YES vacu LOC123206108.1 MiLAC28 572 63,240.64 8.74 31.83 87.9 −0.034 YES chlo LOC123201844.1 MiLAC29 578 63,603.97 9.14 28.91 83.49 −0.063 YES chlo LOC123229714.1 MiLAC30 549 60,833.73 6.05 35.5 77.58 −0.215 NO cyto LOC123227135.1 MiLAC31 574 63,105.94 7.63 31.52 81.86 −0.051 YES chlo LOC123229712.1 MiLAC32 561 62,131.03 5.69 34.2 83.58 −0.074 YES vacu LOC123229362.1 MiLAC33 582 65,859.48 5.35 40.64 83.38 −0.204 YES nucl LOC123229040.1 MiLAC34 568 63,650.32 8.78 35.14 76.02 −0.275 NO vacu LOC123192675.1 MiLAC35 574 63,699.53 6.4 31.04 76.92 −0.073 YES chlo LOC123230046.1 MiLAC36 580 65,597.04 5.53 37.77 78.97 −0.261 YES extr LOC123198865.1 MiLAC37 561 62,316.52 7.69 35.58 84.3 −0.121 YES chlo LOC123201999.1 MiLAC38 566 62,256.27 8.69 30.64 85.92 −0.021 YES vacu LOC123229061.1 MiLAC39 564 62,158.85 4.83 35.2 78.67 −0.105 YES extr LOC123228568.1 MiLAC40 582 65,548.8 5.33 38.46 79.54 −0.267 YES nucl LOC123229436.1 MiLAC41 578 63,779.86 9.04 29.33 80.93 −0.097 YES chlo LOC123216462.1 MiLAC42 499 55,377.93 6.45 36.53 79.66 −0.117 NO cyto LOC123229209.1 MiLAC43 569 62,757.79 5.44 35.49 78.51 −0.093 NO extr LOC123206286.1 MiLAC44 552 61,357.32 7.66 36.69 86.5 −0.106 YES vacu LOC123229039.1 MiLAC45 569 63,648.25 6.1 35.15 79.02 −0.189 YES chlo LOC123229164.1 MiLAC46 569 62,698.66 5.52 35.51 79.02 −0.099 NO extr LOC123209471.1 MiLAC47 557 61,890.37 8.69 29.79 89.89 −0.036 YES chlo LOC123202218.1 MiLAC48 464 51,228.21 9.68 24.53 79.44 −0.176 NO cyto LOC123230053.1 MiLAC49 569 63,418 6.17 32.88 76.1 −0.202 YES E.R. LOC123229109.1 MiLAC50 569 63,430.15 6.62 32.22 76.1 −0.193 YES chlo LOC123216139.1 MiLAC51 566 62,425.18 8.43 32.58 83.22 −0.071 YES chlo LOC123208638.1 MiLAC52 589 65,038.69 8.96 32.73 87.57 −0.038 NO extr LOC123229718.1 MiLAC53 237 25,893.89 4.58 26.69 77.76 −0.093 NO cyto LOC123230055.1 MiLAC54 406 44,523.01 5.25 37.31 76.4 −0.138 NO cysk chlo: chloroplast; plas: plasma membrane; extr: extracellular; vacu: vacuole; cyto: cytoplasm; nucl: nucleus; pero: peroxisomes; E.R.: endoplasmic reticulum; cysk: cytoskeleton. Total RNA was extracted from mango pulp using the SteadyPure Plant RNA Extraction Kit (Cat. No. AG21024; Hunan Accurate Biotechnology Co., Ltd., Changsha, China). First-strand cDNA synthesis was carried out using the Evo M-MLV Reverse Transcription Premix Kit (Cat. No. AG11728; Hunan Accurate Biotechnology Co., Ltd.) on a BIO-RAD T100™ thermal cycler, following the manufacturer's instructions. Quantitative PCR was performed using the SYBR® Green Premix Pro Taq HS qPCR Kit (Cat. No. AG11701; Hunan Accurate Biotechnology Co., Ltd.) on a qTOWER3 real-time PCR system (Analytik Jena AG, Jena, Germany). The thermal cycling protocol was as follows: 95 °C for 10 min, followed by 40 cycles of 95 °C for 5 s, 60 °C for 30 s, and 72 °C for 30 s. All experiments were performed with three biological replicates and three technical replicates. The mango Actin gene was used as the internal reference, and relative transcript levels were calculated using the 2−ΔΔCᴛ method. Primer sequences are listed in Supplementary Table S1.
Detection method of hormone levels in mango flesh
-
The contents of abscisic acid (ABA) and methyl jasmonate (MeJA) were determined using Enzyme-Linked Immunosorbent Assay (ELISA) kits (Catalog No: YJ077235, YJ077224; Shanghai Enzyme-linked Biotechnology Co., Ltd.) according to the manufacturer's instructions. A standard curve was generated using the standard solutions provided in the kit (R2 > 0.9900). After colour development, the optical density (OD) was measured at a wavelength of 450 nm using a microplate reader (Multiskan FC, Thermo Fisher). The concentrations of ABA and MeJA in the samples were calculated based on the standard curve and expressed as ng/g fresh weight (FW) and pmol/g.
Data processing and statistical analysis
-
Statistical analysis was performed using SAS software. A Student's t-test was used to assess the significance of differences between treatment and control groups (p < 0.05 was considered statistically significant, and p < 0.01 was considered highly significant). ANOVA was used to analyse variance in dynamic changes, and Duncan's method was used for multiple comparisons at different time points.
-
A total of 54 MiLAC genes were identified in the mango genome, and they were named MiLAC1 to MiLAC54 based on the naming convention used for Arabidopsis genes. In this study, the protein lengths, molecular weights, isoelectric points, instability indices, fat coefficients, hydrophilicity, signal peptides, and subcellular localisation information of the 54 members of the mango laccase gene family were summarised, as shown in Table 1.
The protein lengths of the mango laccase gene family members range from 237 to 1,147 amino acids, with most proteins approximately 560 amino acids in length. The longest protein is MiLAC53, and the shortest is MiLAC3. There is a certain degree of variation in protein length among the different members, which may be related to the functional diversity of the members. As shown in Table 1, the molecular weights of the MiLAC proteins range from 25,893.89 to 126,257.25 Da, with theoretical isoelectric points ranging from 4.58 to 10.07. The instability indices of the proteins range from 24.53 to 40.64. Except for MiLAC33, all other MiLAC proteins have instability indices below 40, indicating that the proteins in this gene family generally have stable structures. The fat coefficients range from 76.02 to 90.09, and the hydrophilicity values range from −0.275 to 0.014. Except for MiLAC4, whose protein is hydrophobic, all other 53 MiLAC proteins are hydrophilic. Moreover, out of the 54 MiLAC genes, 43 contain signal peptides, while 11 genes (MiLAC3, MiLAC12, MiLAC30, MiLAC34, MiLAC42, MiLAC43, MiLAC46, MiLAC48, MiLAC52, MiLAC53, and MiLAC54) do not. This suggests that most members of this gene family are secretory proteins lacking transmembrane domains. Subcellular localization analysis (Table 1) revealed that the 54 MiLACs are unevenly distributed across various cellular locations, including the extracellular space, cytoplasm, nucleus, plasma membrane, endoplasmic reticulum, chloroplast, peroxisome, and cytoskeleton. Among them, 26 are localised to the chloroplasts, 12 to the vacuole, seven to the extracellular space, five to the cytoplasm, three to the nucleus, two to the peroxisome, two to the plasma membrane, and one each in the cytoskeleton and endoplasmic reticulum.
Phylogenetic tree construction of the MiLAC gene family
-
To investigate the phylogenetic relationships and classification of the MiLAC family members, a phylogenetic tree was constructed using the amino acid sequences of 54 MiLACs and 17 AtLACs family members, based on protein homology (Fig. 1). The results revealed that the MiLAC and AtLAC genes were divided into five subfamilies. Among them, AtLACs were distributed across subfamilies II, III, IV, and V, while MiLACs were primarily distributed in subfamily I, with 23 members; 16 members were located in subfamily V, seven in subfamily II, five in subfamily III, and three in subfamily IV. LACs within the same subfamily are likely to share similar biological functions.
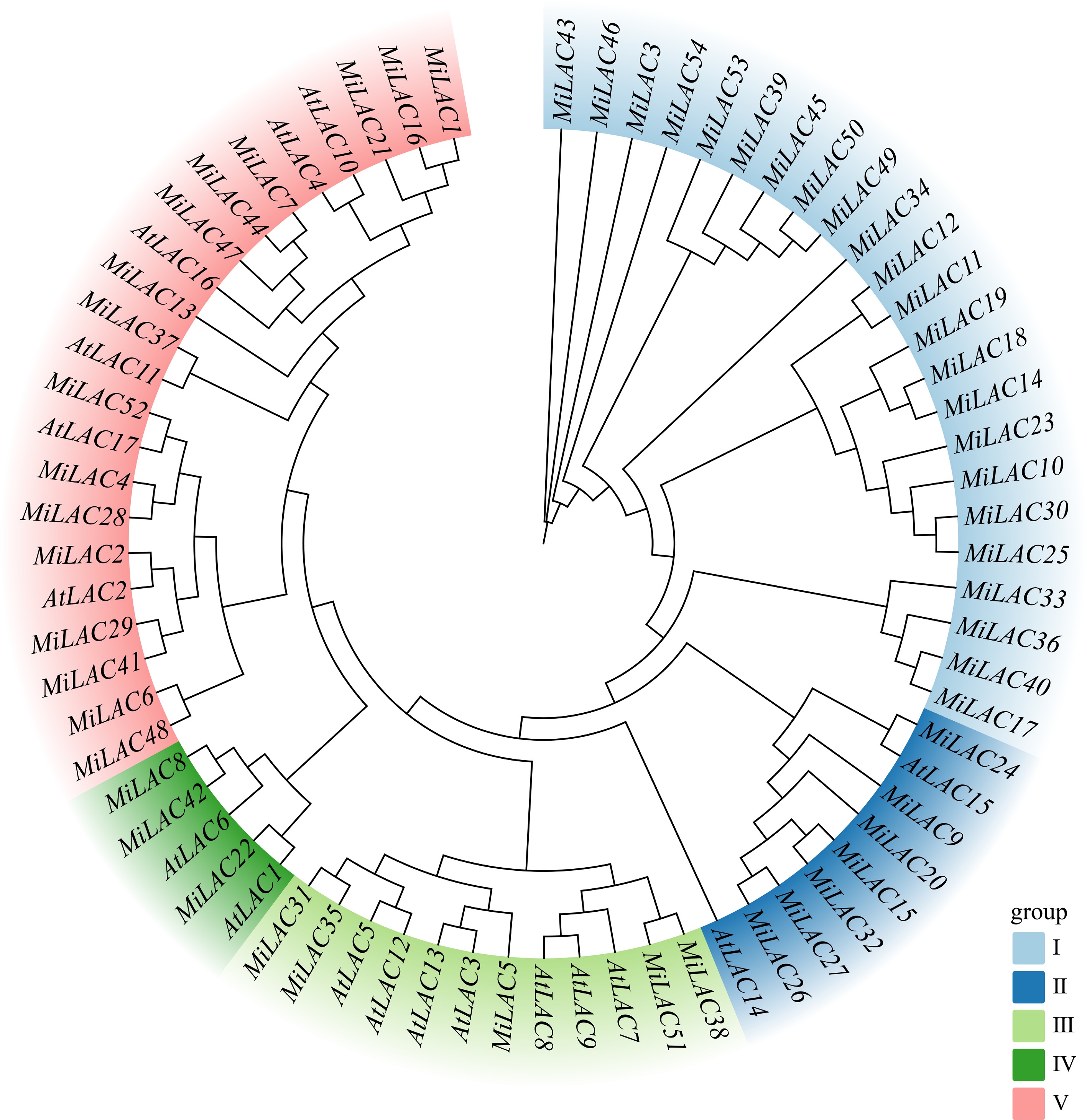
Figure 1.
Phylogenetic tree of MiLACs.
Analysis of conserved domains in MiLACs structural proteins
-
Using the MEME online tool, 10 conserved motifs were identified in the protein sequences of all MiLAC family members, and the conserved sequences of these motifs were summarised (Supplementary Table S2). Additionally, the structure of the 54 MiLACs genes was analysed using TBtools software (Fig. 2). Among the 54 MiLACs, Motif 1, Motif 7, and Motif 2 were present in 52 MiLACs, Motif 3, Motif 5, and Motif 10 were found in 53 MiLACs, and Motif 4 was present in 50 MiLACs. Meanwhile, Motif 9, Motif 8, and Motif 6 were conserved across in all 54 protein sequences. The length of each conserved motif ranged from 15 to 50 amino acids, indicating a high degree of conservation in the protein domains of this gene family. The conserved motifs were further verified using the Pfam database, which revealed that three motifs correspond to three distinct copper oxidase domains (Cu_oxidase, Cu_oxidase_2, and Cu_oxidase_3). Specifically, Motif 1 fully corresponds to the Cu_oxidase_3 domain, Motif 2 matches the Cu_oxidase domain, and Motif 3 constitutes the Cu_oxidase_2 domain. The number of introns in the MiLACs genes ranged from two to 14, with 25 genes containing five introns. The coding regions of MiLAC3, MiLAC47, MiLAC7, and MiLAC44 exhibited higher variability, with these regions separated by relatively long introns. Variations in the number and distribution of exons and introns across different MiLAC genes may be associated with the functional differentiation of these genes.

Figure 2.
Gene structure and motif analysis of MiLAC.
MiLACs chromosomal localization, synteny analysis, and selective pressure analysis
-
In this study, the chromosomal localization map of the MiLAC genes and the syntenic relationships between the members were plotted based on their positional information (Fig. 3). The MiLAC genes are distributed across 12 chromosomes and one scaffold fragment. No MiLAC genes were located on chromosomes 3, 4, 6, 7, 8, 13, 14, or 17. The overall distribution of the gene family on the chromosomes is dispersed, but some members are more closely clustered. The highest concentration of MiLACs was found on chromosome 11, with 27 genes, followed by chromosome 2 with seven genes, chromosome 18 with six, chromosome 1 with three, and chromosome 5 with two genes, and on chromosome 5, two MiLACs were clustered. The remaining chromosomes each contained a single MiLAC gene.
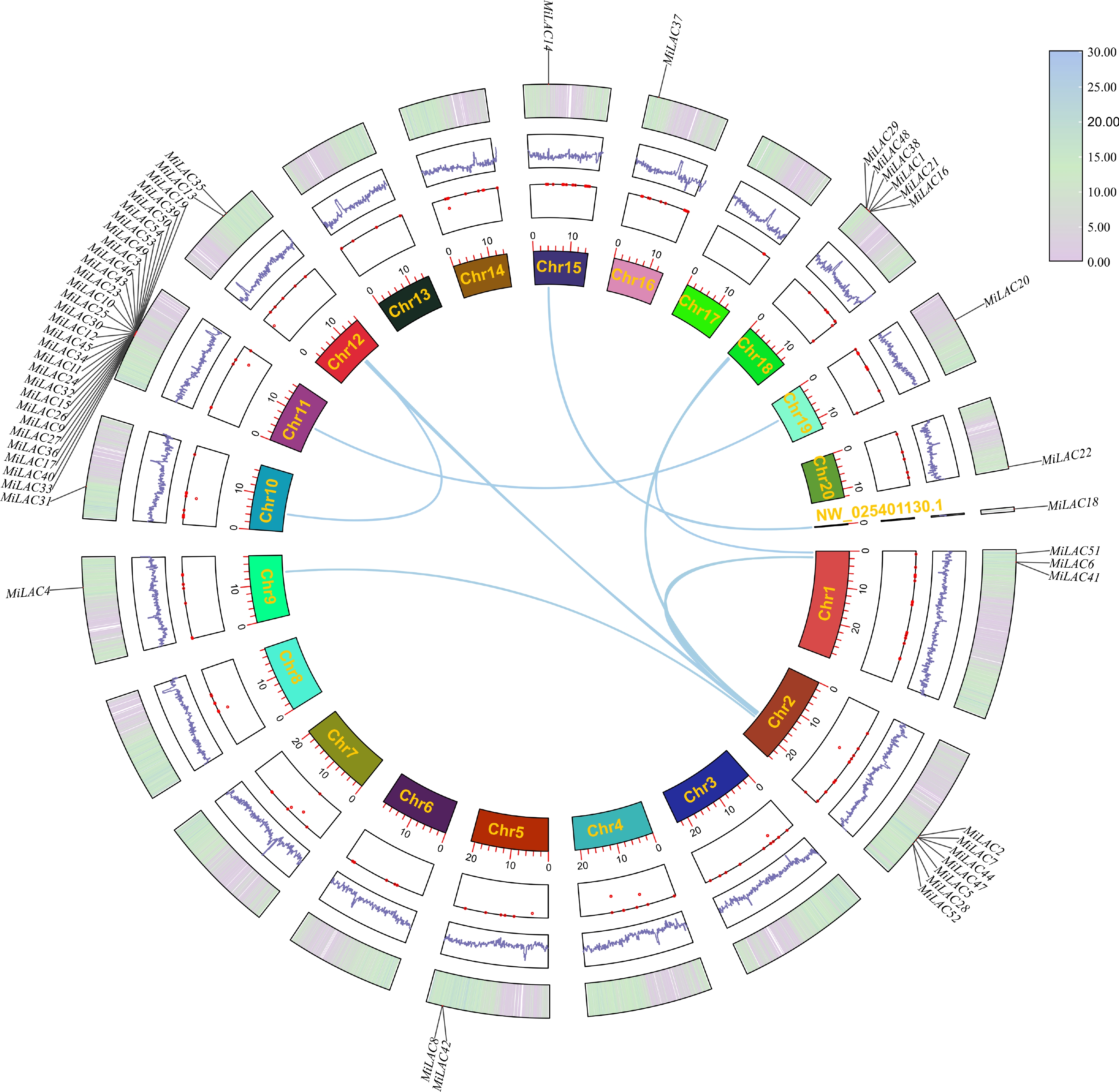
Figure 3.
Chromosome distribution and collinearity analysis of the MiLACs.
Synteny analysis within the mango genome revealed 45 MiLAC genes (83%) involved in tandem duplication and were distributed across 5 chromosomes: Chr1, Chr2, Chr5, Chr11, and Chr18. A total of 10 pairs of homologous relationships were identified among 16 MiLACs, distributed across nine chromosomes and one scaffold fragment. Furthermore, most members exhibit multiple syntenic relationships with different members, suggesting that segmental duplication is the main driving force behind the evolutionary expansion of the MiLAC gene family. These pairs of syntenic genes have likely undergone whole-genome duplication events or have extremely similar gene functions and roles. As shown in Table 2, the Ka/Ks ratios between homologous gene pairs are all well below one, indicating that the MiLAC genes have mainly undergone purifying selection pressure during their evolutionary process.
Table 2. Ka and Ks analysis of MiLACs replication gene pairs.
Seq_1 Seq_2 Ka Ks Ka/Ks MiLAC6 MiLAC2 0.205375182 1.971231992 0.104186206 MiLAC2 MiLAC29 0.186898577 2.325797136 0.080358933 MiLAC28 MiLAC4 0.050855744 0.360717481 0.140984973 MiLAC6 MiLAC5 0.423715485 − − MiLAC5 MiLAC35 0.216972756 1.462202414 0.148387633 MiLAC7 MiLAC13 0.265466731 1.557947571 0.170395163 MiLAC14 MiLAC18 0.002294457 0.017957249 0.127773291 MiLAC27 MiLAC20 0.133210788 0.511023074 0.260674703 MiLAC31 MiLAC35 0.058176459 0.272809991 0.213249005 MiLAC51 MiLAC38 0.080871157 0.579130917 0.139642272 '−': the differences between sequences are too large, resulting in low similarity. Cis-acting element analysis of MiLAC promoter regions
-
The 2,000 bp upstream regions of the 54 MiLAC genes were analysed to identify potential regulatory motifs. After removing uncharacterized sequences, the identified elements were classified into three major categories: abiotic stress-related elements, plant hormone signalling-related elements, and growth and development-related elements (Fig. 4).
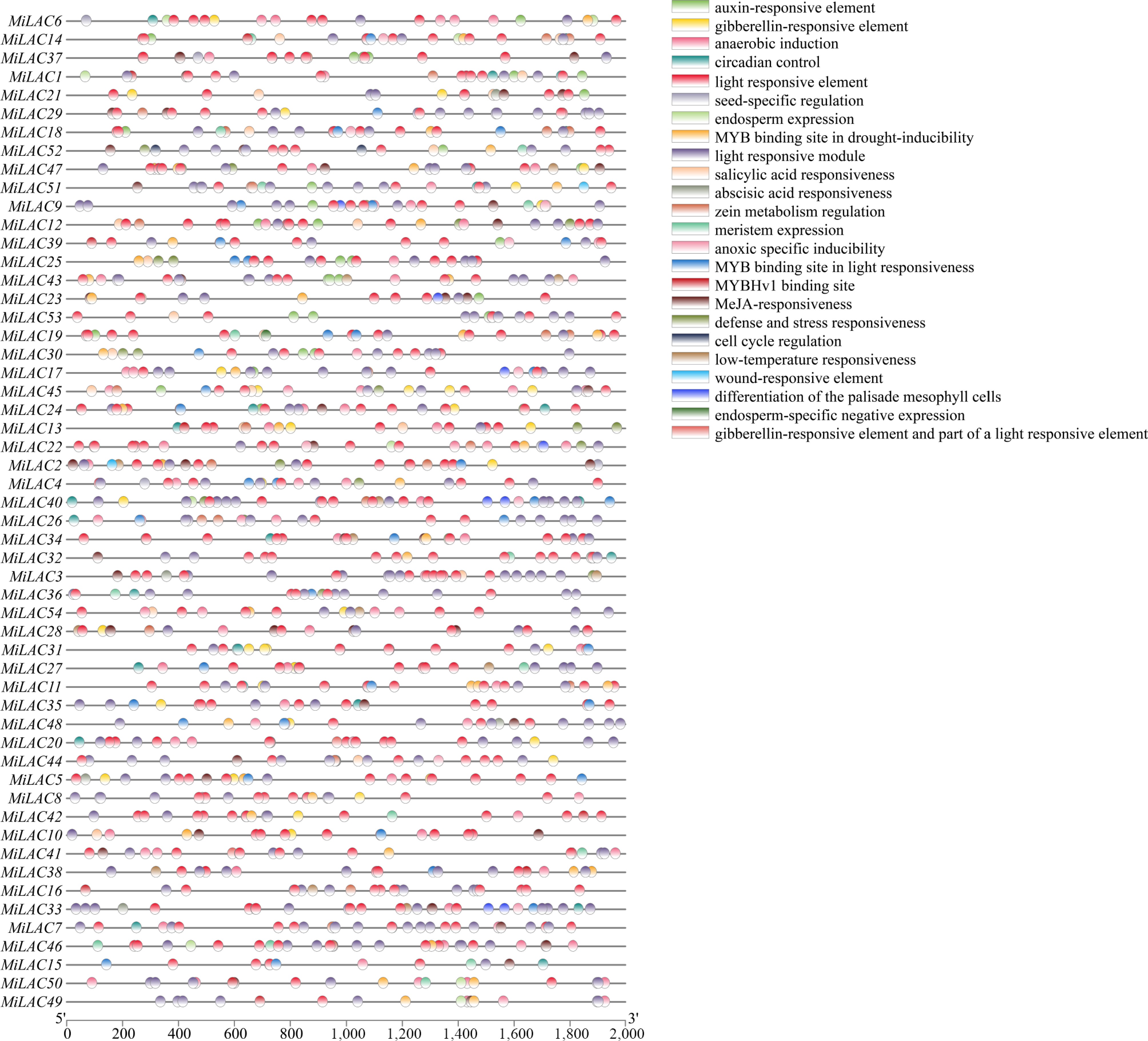
Figure 4.
Analysis of cis-acting elements of MiLACs promoter.
In the stress-related group, five elements were found, corresponding to light response, antioxidant defence, anaerobic conditions, stress and adversity response, and drought and low-temperature response. The results indicate that each MiLAC promoter region contains varying numbers of light-responsive elements, including TCCC-motif, TCT-motif, GATA-motif, GA-motif, AE-box, chsCMA1a, GT1-motif, I-box, Box 4, G-Box, AAAC-motif, 4cl-CMA2b, L-box, MRE, ATCT-motif, Box II, chsCMA2a, 3-AF1 binding site, CAG-motif, AT1-motif, ACA-motif, ACE, ATC-motif, chs-Unit 1 m1, Gap-box, GTGGC-motif, LAMP-element, TCCC-motif, TCT-motif, and Sp1.
Particularly noteworthy is the G-box element, a core regulatory motif involved in both light signalling and hormonal pathways, which was detected in all 54 MiLAC promoters. The widespread presence of these light-responsive elements suggests that MiLAC expression may be subject to intricate regulation by light signals, thereby contributing to plant adaptation to varying light conditions and stress responses. Additionally, antioxidant response elements (AREs), known to play a key role in reactive oxygen species scavenging and the maintenance of redox homeostasis, were identified in the promoter regions of 42 MiLACs. This is especially relevant given that enhanced UV-B radiation is known to trigger a surge in intracellular reactive oxygen species. The convergence of both light-responsive and antioxidant regulatory motifs within the MiLAC promoters points to a possible dual role for these genes in mediating UV-B stress tolerance. Anaerobic-induced enhancer-like elements (GC-motif) were present in 10 MiLAC promoter sequences, defence and stress response elements (TC-rich repeats) were identified in 19 MiLAC promoters, drought stress response elements (MBS) were observed in 33 MiLAC promoters, and cold stress response elements (LTR) were detected in 14 MiLAC promoters. Regarding plant hormones, five types of hormone-related elements were identified: abscisic acid (ABA) response elements (ABRE) were present in 39 MiLAC promoters; auxin response elements (TGA) were found in 22 MiLAC promoters; gibberellin response elements (GARE-motif, P-box, TATC-box) were present in 27 MiLAC promoters; methyl jasmonate (MeJA) response elements (CGTCA-motif, TGACG-motif) were identified in 30 MiLAC promoters; and salicylic acid (SA) response elements (TCA-element, SARE) were present in eight MiLAC promoters. Notably, the most common elements were the ABRE (ABA) and CGTCA-motifs, and the TGACG-motif (MeJA), with MiLAC3 and MiLAC50 containing the highest number of cis-acting elements, which are primarily ABA and MeJA response elements. This highlights the significant role of the MiLAC family in hormone regulation.
Additionally, six growth and development-related elements were identified, including meristem-specific expression elements (CAT-box) in 14 MiLAC promoters; circadian rhythm regulation elements (circadian) in 20 MiLAC promoters; seed-specific elements (RY element) in five MiLAC promoters; zein metabolism elements (O2-site) in 21 MiLAC promoters; MYBHv1 binding sites (CCAAT-box) in nine MiLAC promoters; and endosperm-specific expression elements (GCN4) in eight MiLAC promoters. The dominance of the O2-site element suggests that MiLACs may be involved in developmental processes. Cis-acting element analysis revealed that the MiLAC family is extensively involved in light morphogenesis, antioxidant responses, hormone signalling, and other growth and development mechanisms. The richness and diversity of these elements suggest that MiLACs play a crucial role in mango's adaptation to environmental stimuli and developmental regulation.
Expression analysis of MiLACs under UV-B stress and qRT-PCR validation
-
To investigate the impact of enhanced UV-B treatment on mango pulp, the expression levels of MiLAC genes associated with the lignin biosynthesis pathway were analysed and compared with the control group. Based on transcriptome data and the lignin synthesis pathway (phenylpropanoid biosynthesis, ko00940), a differential gene expression heatmap was generated (Supplementary Fig. S1). In both 2023 and 2024, MiLAC3 and MiLAC50 were selected as representative family members for quantitative real-time PCR (qRT-PCR) validation over two consecutive years. The results (Fig. 5) showed that, at 50 d post-flowering in 2023 and 61 d post-flowering in 2024, the expression levels of MiLAC3 and MiLAC50 were significantly higher in the treated group compared to the control group, which is consistent with the transcriptome analysis. These results indicate that enhanced UV-B radiation treatment increases the expression of key MiLAC genes, thereby enhancing LAC activity and subsequently promoting lignin accumulation in the mango pulp. This process may help mitigate oxidative damage to the pulp, providing a protective effect against reactive oxygen species.
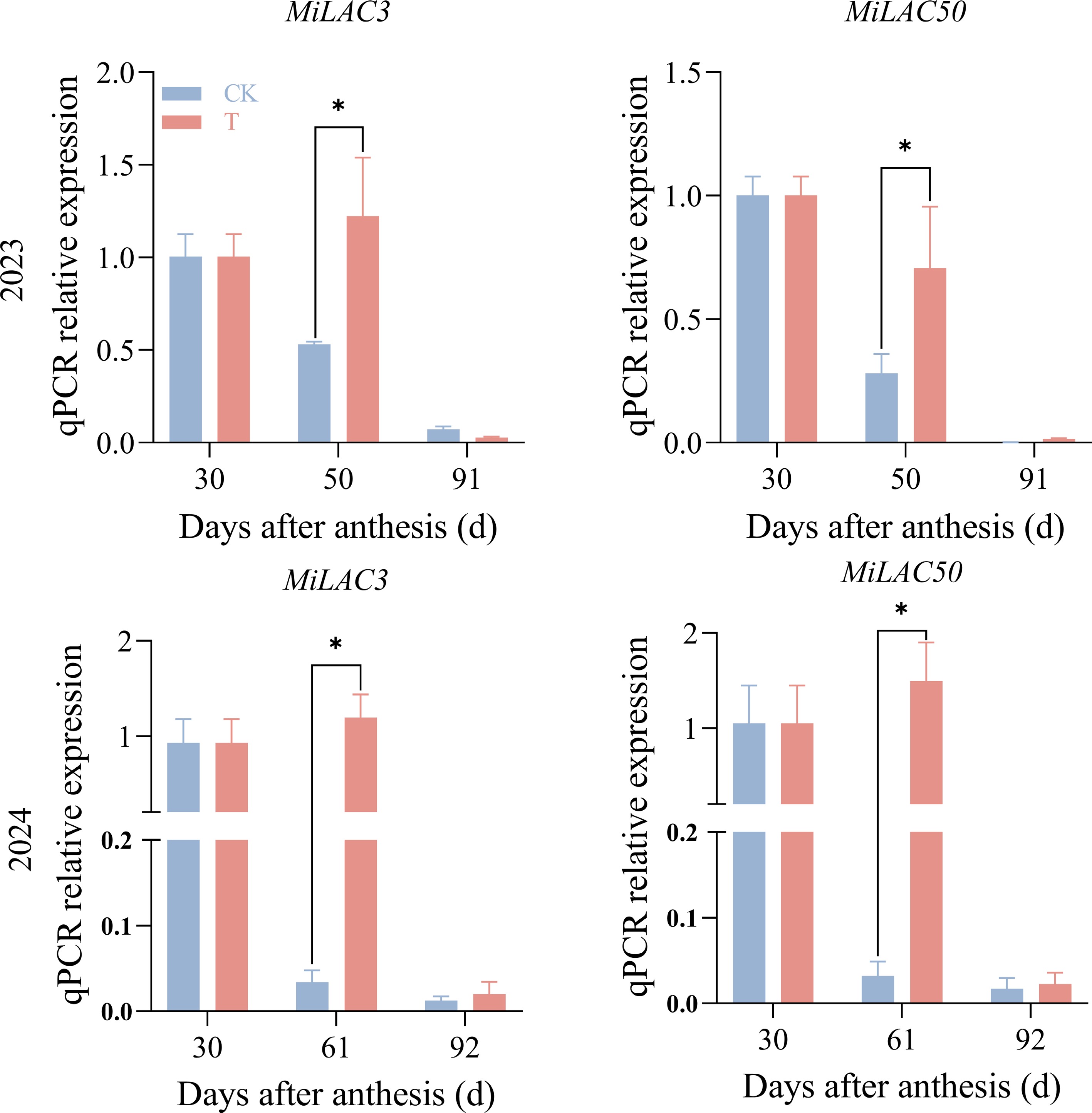
Figure 5.
q-PCR results of MiLACs under enhanced UV-B radiation treatment. Note: the two images in the first row are from 2023, and the two images in the second row are from 2024.
Effects of enhanced UV-B radiation treatment on ABA and MeJA levels in mango pulp
-
The dynamic changes in hormone levels between the treatment and control groups in mango pulp were consistent both within and between years, with significant differences observed at different sampling time points. Over the 2 years, the trends in hormone levels at the same sampling time point were generally similar between the treatment and control groups (Fig. 6).
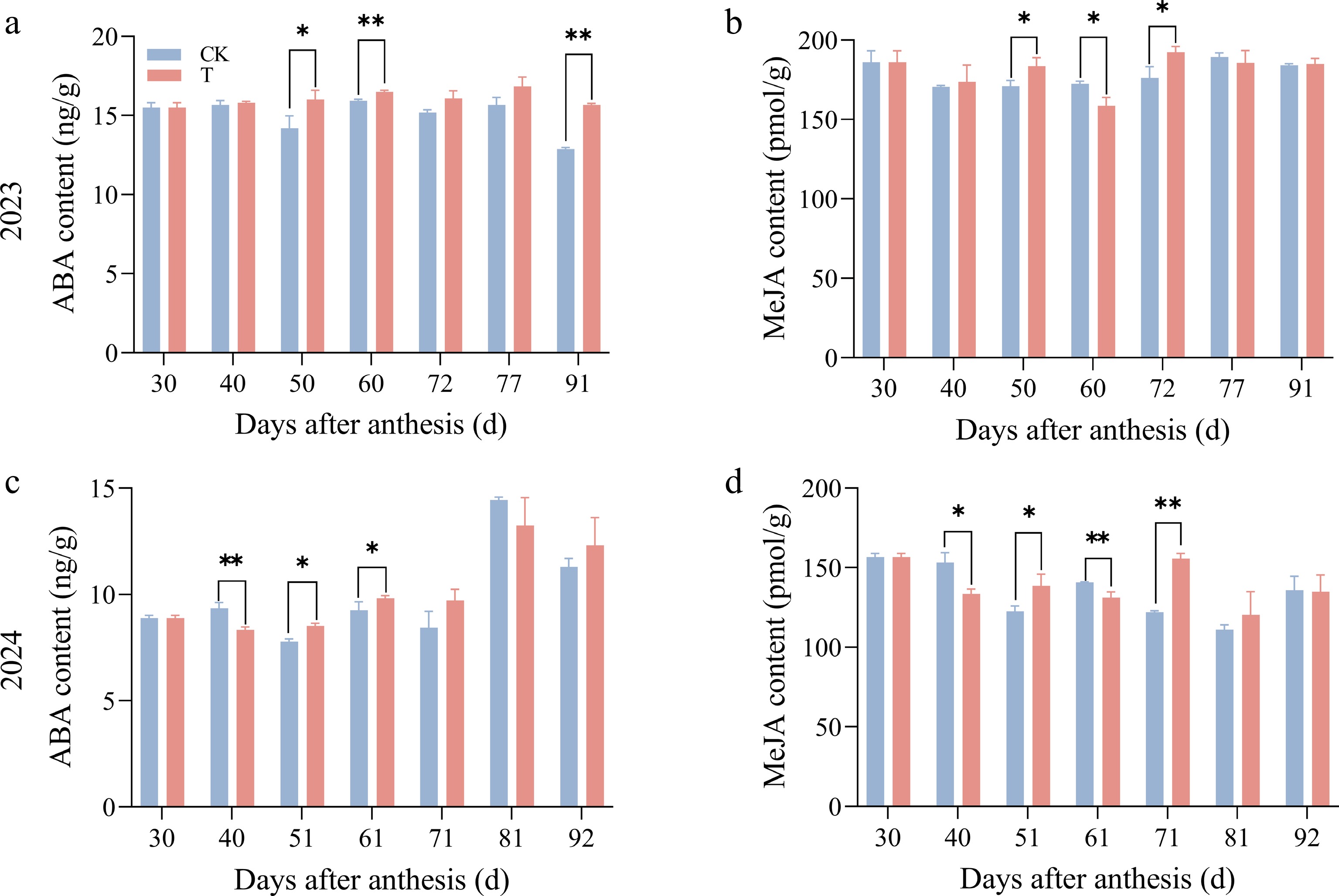
Figure 6.
Effect of enhanced UV-B radiation treatment on ABA and MeJA content changes in the pulp of 'Tainongyihao' mango.
ABA content tended to be higher in the treatment group overall, with significant differences observed at 51 and 61 d after flowering. However, at 40 and 81 d after flowering in 2024, ABA levels were significantly lower in the treatment group compared to the control. MeJA content was higher in the treatment group during the mid-stage of rapid fruit expansion. The main interannual variation was observed at 40 d after flowering in 2024; MeJA content was significantly lower in the treatment group, while no significant difference was observed at the same period in 2023. For both hormones, no significant differences were found between the treatment and control groups at the remaining sampling time points. In summary, despite subtle year-to-year fluctuations in hormone levels between the treatment and control groups, the overall trends over the 2 years were consistent. Together with the finding that MiLAC3 and MiLAC50 promoters contain the most ABA and MeJA response elements and exhibit upregulated expression, this suggests that enhanced UV-B radiation treatment may upregulate MiLAC3 and MiLAC50 expression by increasing ABA and MeJA levels in mango pulp.
-
In this study, a total of 54 MiLACs were identified. Comparative analysis with species such as Arabidopsis thaliana, strawberry, sugarcane, potato, Populus davidiana × P. bolleana, sweet orange, canola, cotton, sorghum, and Eucalyptus grandis revealed variations in the number of gene family members. These differences may be attributed to variations in genome size and gene duplication events during the evolutionary processes of these species[35]. The MiLAC gene family was classified into five subfamilies, a clustering pattern consistent with findings in species such as strawberry[36] and potato[37]. Notably, Subfamily I contained the largest number of MiLAC members, whereas Arabidopsis thaliana LACs were entirely absent from this subgroup. This stark contrast strongly suggests that Subfamily I represents a mango-specific expansion branch. Such an expansion reflects distinct duplication events and subsequent functional divergence experienced by mango during its evolution. Gene duplication serves as the primary driver of gene family expansion. Our analysis indicates that both tandem and segmental duplications have played pivotal roles in the evolution of MiLACs, not only facilitating the genesis of novel genes but also driving functional diversification[38]. The uneven distribution of MiLACs across different chromosomes may be attributed to tandem or whole-genome duplication events; furthermore, such a distribution pattern may be intrinsically linked to specific biological functions during mango fruit growth and development. Despite these genetic variations, MiLACs exhibit a high degree of evolutionary conservation. Members within the same subfamily share similar exon-intron structures and conserved motifs, implying that they likely possess analogous biological functions.
Analysis of cis-acting elements in the promoter regions further corroborates this conservation and provides crucial clues regarding their regulatory networks. Similar to LACs in peach[39], apple[40], and amaranth[41], the promoter regions of MiLACs are enriched with light-responsive elements, highlighting the genetic conservation of LACs across different plant species in response to light signaling. Furthermore, the MiLACs promoters contain a diverse array of hormone and stress-responsive elements. Elements responsive to phytohormones such as abscisic acid (ABA), methyl jasmonate (MeJA), and salicylic acid (SA) are particularly prevalent. These hormones frequently act as pivotal secondary messengers that bridge the perception of abiotic stress to the activation of downstream defense genes[42]. Consistent with research on potato LACs[37], the presence of these elements in MiLACs promoters suggests they may be induced under various stress conditions, thereby participating in the regulation of antioxidant functions. Among them, the promoters of MiLAC3 and MiLAC50 displayed the greatest abundance of cis-elements, being enriched with ABA-responsive and MeJA-responsive elements, respectively.
Importantly, the temporal dynamics of ABA and MeJA contents (Fig. 6) closely paralleled the expression patterns of MiLAC3 and MiLAC50, with both hormones and transcripts peaking during the middle to late fruit development stages. This correlation, observed across both years despite interannual absolute differences, supports the idea that endogenous ABA and MeJA functionally induce MiLAC3 and MiLAC50 expression in a concentration-dependent manner, linking hormone accumulation to laccase-mediated lignin biosynthesis.
To synthesize these molecular findings with physiological adaptations, we propose a coherent regulatory network linking UV-B radiation, hormone signaling, and lignin biosynthesis. Previous research by our group demonstrated that enhanced UV-B radiation treatment significantly improves the resistance of mango fruit to environmental stress. The underlying mechanism begins with the perception of UV-B, which rapidly promotes the accumulation of endogenous phytohormones, notably ABA, SA, and MeJA, in the pulp during the mid-stage of rapid fruit expansion[26]. These stress-responsive hormones then act as signaling molecules that specifically bind to the corresponding enriched cis-acting elements within the promoters of specific laccase genes. This binding event directly triggers the transcriptional upregulation of these genes, which aligns perfectly with our observation that the promoters of MiLAC3 and MiLAC50 are exceptionally rich in ABA and MeJA response elements.
Following this hormone-mediated gene expression, the increased abundance of MiLAC enzymes catalyzes the polymerization of phenolic precursors, driving the synthesis of lignin. Indeed, our earlier work indicated that the lignin content in fruit treated with enhanced UV-B radiation was significantly higher than in the control group. Lignin accumulation aids in dissipating the stress effects caused by UV-B radiation by acting as a physical screen and mitigates cellular damage by scavenging reactive oxygen species, thus enhancing the fruit's overall adaptability to adversity[26]. Consequently, these results support that enhanced UV-B radiation induces ABA and MeJA accumulation, which in turn activates the expression of MiLAC3 and MiLAC50 via promoter binding. The upregulated laccases then promote lignin accumulation during mango fruit maturation, ultimately strengthening the fruit's structural and biochemical tolerance to UV-B stress.
-
The mango LAC gene family consists of 54 members, exhibiting both conservation and evolutionary divergence. Segmental duplication appears to be a major evolutionary mechanism driving the expansion of this gene family. Most members contain hormone and stress response elements, suggesting that they play regulatory roles in response to stress and hormonal signalling. Under enhanced UV-B radiation treatment, mango fruit likely upregulates the expression of MiLAC3 and MiLAC50 by increasing the levels of ABA and MeJA, which in turn promotes lignin synthesis, enhances resistance to reactive oxygen species (ROS) damage, and alleviates the harmful effects of UV-B radiation stress, thereby improving resistance to enhanced UV-B stress. These findings provide a theoretical foundation for future functional studies of MiLACs and offer valuable insights for developing breeding strategies and cultivation techniques to enhance mango's resistance to enhanced UV-B radiation stress.
-
The authors confirm their contributions to the paper as follows: study conception and design: Gao Y, Zhou K, Xiong Y; data collection: Gao Y, Tahir H; analysis and interpretation of results: Gao Y, Tahir H, Xiong Y, Zhou K; draft manuscript preparation: Gao Y; supervision, project administration, funding acquisition: Zhou K. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available from the corresponding author upon reasonable request.
-
This work was financially supported by the National Natural Science Foundation of China (No. 32160677), the Natural Science Foundation of Hainan Province, China (No. 321RC465), and the Hainan University Mango Research System.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/tp-0026-0019.
-
Received 22 January 2026; Accepted 6 May 2026; Published online 5 June 2026
- Supplementary Table S1 Primer sequences for different genes.
- Supplementary Table S2 Conserved motif for the protein of the MiLAC.
- Supplementary Fig. S1 Expression patterns of LAC genes expressed (DEGs) involved in lignin biosynthesis in mango pulp.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao Y, Tahir H, Zhou K, Xiong Y. 2026. Genome-wide identification of the laccase phylogenetic family in mango and expression analysis in response to enhanced UV-B radiation treatment. Tropical Plants 5: e018 doi: 10.48130/tp-0026-0019 

Genome-wide identification of the laccase phylogenetic family in mango and expression analysis in response to enhanced UV-B radiation treatment
- Received: 22 January 2026
- Revised: 27 April 2026
- Accepted: 06 May 2026
- Published online: 05 June 2026
Abstract: Laccases are copper-containing polyphenol oxidases involved in plant growth, development, and stress responses. In this study, members of the mango laccase multigene family (MiLACs) were identified and characterised, and their expression patterns under enhanced UV-B radiation were analysed to provide a basis for further functional studies. A total of 54 MiLAC genes were identified in the mango genome and mapped to 12 chromosomes and one scaffold. The encoded proteins ranged from 237 to 1,147 amino acids, with most being approximately 560 aa in length. Most MiLAC proteins were predicted to be stable and hydrophilic; 43 contained signal peptides, and most were predicted to localise to chloroplasts. Phylogenetic analysis grouped the MiLAC family into five subgroups, whose members shared similar gene structures. Segmental duplication appeared to be a major driver of family expansion. In addition, all syntenic gene pairs showed Ka/Ks ratios far below 1, indicating that MiLACs have mainly undergone purifying selection during evolution. MiLAC proteins mainly contained conserved blue copper protein domains with three copper oxidase domains. Promoter analysis revealed numerous light-, hormone-, and stress-responsive cis-elements, with MiLAC3 and MiLAC50 containing the most ABA- and MeJA-responsive elements. Expression analysis showed that MiLAC3 and MiLAC50 were significantly upregulated under enhanced UV-B treatment, accompanied by increased ABA and MeJA contents, suggesting that these genes may contribute to UV-B stress tolerance in mango.
-
Key words:
- Mango /
- Laccases /
- Phylogenetic analysis /
- Enhanced UV-B radiation /
- Expression analysis