






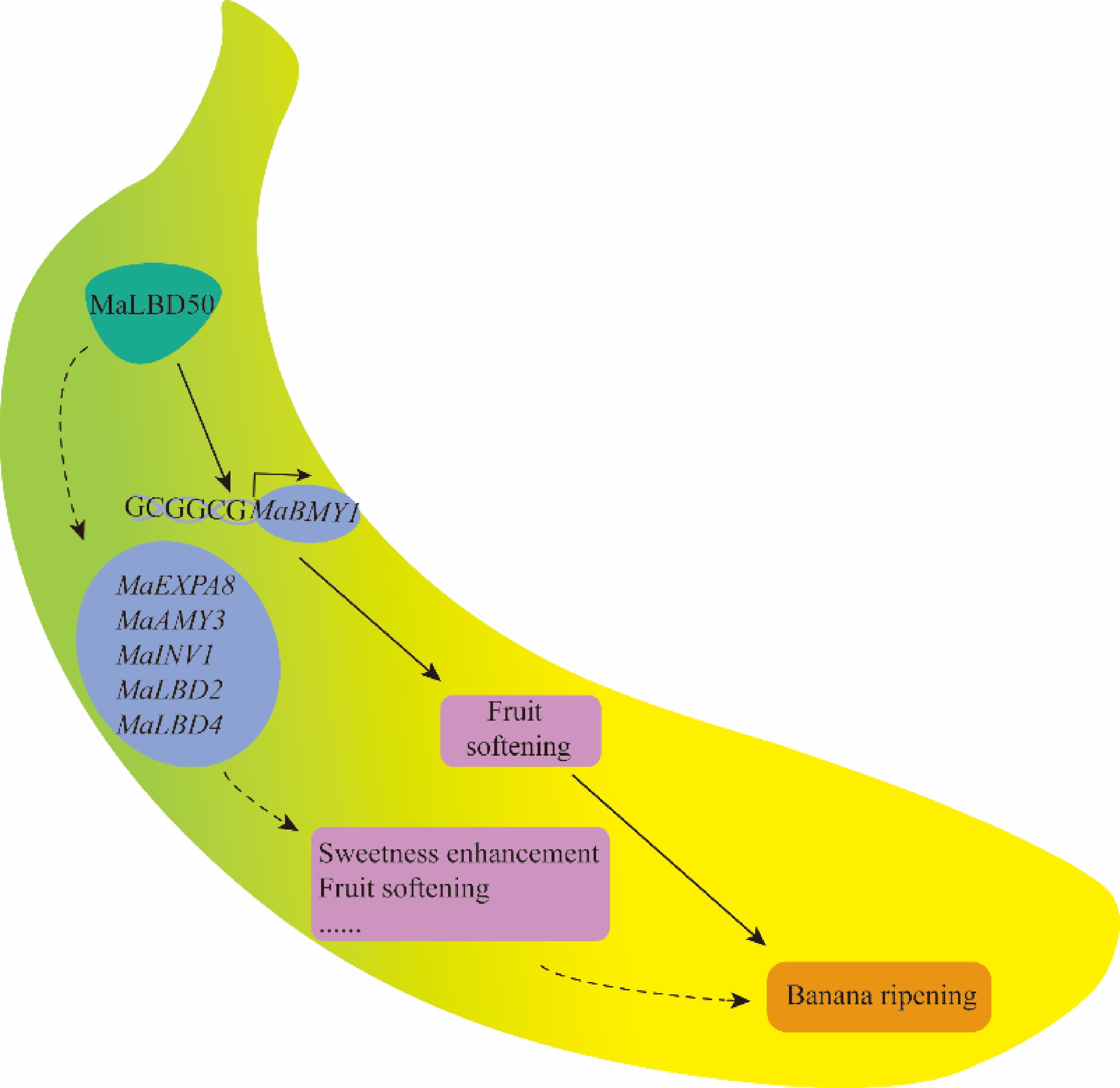



-
Bananas (Musa spp.) are the most widely traded and consumed fresh fruit worldwide and also serves as a staple food in several countries in Africa, Oceania, and the Americas[1]. As a climacteric fruit, its ripening is accompanied by coordinated changes in taste, firmness, aroma, and color, which are determined by both the accumulation of substances during fruit development and their subsequent conversion during postharvest ripening[2]. For example, the degradation and modification of cellulose and pectin lead to fruit softening. Starch synthesis and accumulation during development, followed by its rapid hydrolysis during ripening, provide substrates for the accumulation of soluble sugars, and the balance between sugar metabolism and transport ultimately determines sweetness. Each of these pathways is driven by key enzymes, including amylases, sucrose synthase, pectin methylesterase, lipoxygenase, and alcohol acyltransferase, whose activities are fundamentally controlled by the spatiotemporal expression of their encoding genes.
The expression of these enzyme-coding genes is regulated by complex transcriptional networks that act at multiple levels, including transcription, translation, and post-translational modification, with transcriptional regulation playing a central role[3−5]. In recent years, numerous transcription factors and regulatory genes that modulate banana's ripening rate, fruit composition, and postharvest physiological disorders have been identified[2,6−11]. The fruitENCODE project identified three transcriptional feedback circuits that govern ethylene-dependent ripening[12]. In banana, a dual-loop regulatory model has been proposed to explain why ripening, once initiated, cannot be inhibited by 1-methylcyclopropene (1-MCP), as an independent MaNAP1–MaMADS1 loop sustains ethylene biosynthesis. Based on this work, integrated analyses of DNase I hypersensitive site sequencing (DNase-seq), chromatin immunoprecipitation sequencing (ChIP-seq), and RNA sequencing (RNA-seq) data revealed that the MaNAP1–MaMADS1 module coordinates ethylene signaling with epigenetic regulation to control peel softening and pedicel abscission during ripening[6]. In addition, MaNAC1/19/29/42/169 regulate fruit ripening by forming homo- or heterodimers, thereby integrating ethylene biosynthesis, sucrose accumulation, and cell wall degradation into a core transcriptional network[7,11,13−15].
Starch is the primary energy source for cellular respiration and provides the prerequisite energy supply for the climacteric respiration and the ethylene-induced ripening of banana fruit[2,10,16−21]. During postharvest ripening, starch is degraded to soluble sugars by α-amylase (AMY), β-amylase (BMY), and isoamylase (ISA) under complex transcriptional regulation[2]. Transcription factors such as MaMADS36, MaEIL2, MabHLH6, and MabZIP21 activate the expression of these enzymes' genes, thereby facilitating starch–sugar conversion and promoting fruit ripening[18,20−23]. In contrast, MaAP2a, MaMYB3, MaMYB44, and MaMYB73 act as negative regulators by repressing these genes to delay fruit ripening[17,24,25]. Although these findings underscore the complexity of the transcriptional regulation of starch degradation in banana, many questions remain unresolved, particularly whether lateral organ boundaries domain (LBD) transcription factors participate in this process.
The LBD gene family comprises plant-specific transcription factors, named after the lateral organ boundaries (LOB) gene first identified in Arabidopsis thaliana[26]. Typical LBD proteins contain a highly conserved LOB domain at the N-terminus, consisting of a zinc finger-like motif (CX2CX6CX3C), a glycine–alanine–serine motif (GAS block), and a leucine zipper-like motif (LX6LX3LX6L)[27,28]. In A. thaliana, on the one hand, AtLBD regulates the formation of lateral root primordia by modulating auxin signaling; on the other hand, it indirectly leads to abnormal starch accumulation by inhibiting nitrogen assimilation[29,30]. In rice (Oryza sativa), OsLBD genes show dynamic spatiotemporal expression patterns during reproductive development, with certain members participating in pollen cell walls' formation and endosperm starch biosynthesis[31]. In poplar (Populus spp.), LBD proteins contribute to wood formation by regulating secondary growth[32]. These findings show that LBD transcription factors are involved in diverse biological processes across species[29,31,33−35]. Given that LBD transcription factors have been shown to play roles in starch metabolism in both rice and A. thaliana, and considering that banana fruit ripening is characterized by the massive conversion of starch to soluble sugars, we hypothesized that certain LBD members may play previously unrecognized roles in this process. However, the size and evolutionary characteristics of the LBD gene family in banana, as well as its regulatory mechanisms in banana ripening, remain largely unknown.
In this study, we systematically identified members of the LBD gene family by examining the banana genome and analyzed their physicochemical properties, chromosomal distribution, phylogenetic relationships, and synteny. Integrated multiomics analyses identified MaLBD50 as a candidate gene associated with banana fruit ripening. Yeast one-hybrid (Y1H) assays, dual-luciferase reporter (DLR) assays, and transient expression assays demonstrated that MaLBD50 promotes starch degradation and fruit ripening by directly activating MaBMY1. Collectively, these findings broaden our understanding of the roles of LBD transcription factors in fruit ripening and provide a promising target gene and theoretical foundation for the molecular breeding of banana.
-
Two banana cultivars were used. Specifically, 'Baxijiao' (M. acuminata, AAA group, cv. Cavendish) was used for all sequencing experiments (RNA-seq, DNase-seq, DAP-seq) and gene expression analyses, all of which were performed using pulp tissue. 'Haigong' (Musa spp., AA group, Inarnibal subgroup) was used only for transient transformation experiments on LBD genes. Fruits were harvested at the mature green stage from a commercial plantation in Hainan Province, China. Healthy fruits with a uniform size and maturity and without mechanical damage or disease symptoms were selected. The fruits were surface-sterilized by immersion in 0.1% (v/v) Sporgon solution for 5 min.
Tobacco (Nicotiana benthamiana) plants were grown in a growth chamber at 22 °C under long-day conditions, and 4-week-old plants were used for infiltration.
Identification and bioinformatic analysis of the LBD gene family in banana
-
The hidden Markov model (HMM) file corresponding to the LBD domain (PF03195) was searched against the banana (M. acuminata, DH-Pahang v4) protein data[36]. The domain integrity of candidate genes was verified using the NCBI-CDD (
www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ) databases. Physicochemical properties, including the molecular weight (MW), theoretical isoelectric point (pI), instability index, aliphatic index, and grand average of hydropathicity (GRAVY), were analyzed using the ProtParam tools of the Expasy web platform (https://web.expasy.org/protparam ). The chromosomal localization of MaLBDs was visualized by TBtools[37]. Multiple sequence alignment and visualization of LBD proteins were performed using DNAMAN software. The phylogenetic tree of LBD proteins from banana, Arabidopsis, rice, and grape (Vitis vinifera) was constructed using the neighbor-joining (NJ) method with 1,000 bootstrap replicates, the Poisson model, and complete deletion. MCScanX software was used to analyze synteny blocks among banana, Arabidopsis, and rice, and the results were visualized by TBtools.Chrogmatin accessibility analysis and prediction transcription factor-binding motifs and cis-acting elements in the MaLBD promoter
-
The chromatin accessibility analysis was performed using published DNase-seq data[12]. In brief, the procedure was as follows. Nuclei were extracted from banana pulp and digested with DNase I, and fragmented DNA was size-selected to 150–300 bp. Libraries were constructed using the TruePrep DNA Library Prep Kit (Vazyme) and sequenced on the Illumina NovaSeq 6000 platform (2 × 150 bp). After trimming, the raw reads were aligned to the banana reference genome using Bowtie2 (version 2.2.5). Subsequently, SAMtools (version 1.7) was used to filter out reads with a mapping quality below 30, nonuniquely mapped reads, and reads mapped to organelle genomes. Polymerase chain reaction (PCR) duplicates were removed using Picard (version 3.1.1), and peak calling was performed with MACS2 (version 2.2.7.1). To identify consistent peaks between biological replicates, irreproducible discovery rate (IDR, version 2.0.4.2) analysis was conducted. Peak annotation was performed using ChIPseeker (version 1.30.3), with the promoter region defined as 2 kb upstream of the start codon (ATG). The PlantCARE (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html ) and JASPAR (https://jaspar.elixir.no ) databases were used to predict cis-acting elements and transcription factor-binding motifs in the MaLBD promoter region, respectively.RNA-seq data analysis
-
For data analysis, raw data were processed using Trim Galore (version 0.6.7) to remove adapter sequences and low-quality reads, and quality control was performed using FastQC (version 0.11.9). Ribosomal RNA contamination was removed using Bowtie2 (version 2.2.5). The remaining clean reads were aligned to the banana reference genome using HISAT2 (version 2.1.0). Sorting and indexing of BAM files were performed using SAMtools (version 1.7), and gene counts were quantified using the featureCounts function within the Rsubread package (version 2.8.1). Differential expression analysis was conducted using DESeq2 (version 1.34.0). Differentially expressed genes (DEGs) were identified on according to the following parameters: q-value ≤ 0.01 and |log2fold change (FC)| ≥ 1.
Agrobacterium-mediated transient transformation of banana
-
The full-length coding sequence (CDS) of MaLBD50 was cloned into the pEXT06 vector to generate the OE-MaLBD50 construct. An approximately 300-bp specific fragment of MaLBD50 was cloned into the pTRV2 vector to generate the RNAi-MaLBD50 construct. The recombinant vectors were transformed into Agrobacterium tumefaciens strain EHA105 harboring the pSoup helper plasmid (Weidi Bio, catalog number AC1012). Agrobacterium cells grown to the logarithmic phase were resuspended in an infection buffer (10 mM 2-[N-morpholino] ethanesulfonic acid [MES], 10 mM MgCl2, 200 μM acetosyringone) and incubated in the dark for 2 h. The bacterial suspension was injected into the distal ends of banana fruits using a 1-mL syringe. After 2 d in the dark, the fruits were treated with 0.4% ethephon, and then phenotypic changes were recorded.
Determination of starch content
-
The starch content of banana pulp was determined using a spectrophotometric method. Briefly, 0.1 g of banana pulp powder was defatted with petroleum ether and washed with ethanol to remove soluble sugars. The residue was then dissolved in dimethyl sulfoxide (DMSO) in a boiling water bath. Subsequently, the solution was subjected to enzymatic hydrolysis with amylase at 40 °C for 30 min to convert starch into glucose. The supernatant was collected and mixed with a chromogenic reagent. The reaction mixture was incubated at 40 °C in the dark for 20 min, and the absorbance was measured at 510 nm. Starch content was calculated on the basis of a glucose standard (0.1 mg/mL).
Quantitative real-time PCR
-
Total RNA from banana fruit was isolated using the hot borate method as described previously[38]. Briefly, fruit powder was added to a lysis buffer prewarmed to 90 °C and then incubated at 42 °C with gentle rotation for 2 h. Potassium chloride was added to a final concentration of 160 mM and incubated on ice for 1 h to precipitate proteins. To precipitate RNA, LiCl was added to a final concentration of 0.2 M, and the samples were incubated on ice overnight. The next day, the RNA pellet was dissolved in 100 mM Tris-HCl (pH 7.5) and then reprecipitated with 0.2 M KAc (pH 5.5) to remove polysaccharides and other insoluble contaminants. The RNA was washed twice with ethanol, and its concentration and quality were determined.
Total RNA was reverse-transcribed into cDNA using HiScript IV 1st Strand cDNA Synthesis Kit (Vazyme, catalog number R412-02). Quantitative real-time PCR (qRT-PCR) was performed using ChamQ Universal SYBR qPCR Master Mix (Vazyme, catalog number Q711-03) on a QuantStudio 6 Flex Real-Time PCR System. Relative gene expression levels were calculated using the 2−ΔΔCᴛ method, with MaRPS2 serving as the internal reference gene.
DAP-seq library construction and data analysis
-
Genomic DNA (gDNA) was extracted from banana pulp using the cetrimonium bromide (CTAB) method[6]. The gDNA was fragmented by sonication, and DNA fragments ranging from 150 to 250 bp were recovered to construct the DNA library. DAP-seq library construction was performed according to a previously described method with minor modifications[6,38,39]. Briefly, The CDS of MaLBD50 was cloned into the HaloTag vector to express Halo-MaLBD50 protein in vitro using a wheat (Triticum aestivum) germ cell-free expression system (Promega, catalog number L4130). The Halo-MaLBD50 protein was incubated with magnetic beads at room temperature for 1 h to conjugate the protein to the beads. Subsequently, the prefragmented DNA library was incubated with the beads at room temperature for 1 h to allow MaLBD50 binding, followed by phosphate-buffered saline (PBS) washes to remove nonspecific binding. Bound DNA was eluted by heating at 98 °C for 10 min and amplified by PCR. The PCR products were purified and sequenced on an Illumina NovaSeq 6000 platform.
Raw DAP-seq data were filtered and subjected to quality control, followed by alignment to the banana reference genome. Peak calling was performed using MACS2 (version 2.2.7.1). Motif enrichment analysis was conducted using HOMER (version 4.11). For DNA affinity purification (DAP) quantitative PCR (qPCR) analysis, specific primers were designed, based on the signal peak positions identified by DAP-seq. The DAP-seq library was used as the template, with the input DNA library serving as the control.
Yeast one-hybrid assay
-
The CDS of MaLBD50 was cloned into the pB42AD vector. Approximately 500 bp of the promoter sequence of MaBMY1 was individually cloned into the pLacZi vector. The pB42AD-MaLBD50 plasmids were co-transformed with the pLacZi-ProMaBMY1 plasmid into Saccharomyces cerevisiae strain EGY48 and plated on a synthetic dextrose (SD)/-Trp/-Ura dropout medium. Positive clones were spotted onto the SD/-Trp/-Ura medium supplemented with X-Gal for Y1H binding analysis.
Dual-luciferase reporter assay
-
The CDS of MaLBD50 was cloned into the pSuper1300 vector to generate the 35S-MaLBD50, and the promoter sequence of MaBMY1 was cloned into the pGreenII 0800-LUC vector to generate ProMaBMY1-LUC. These recombined constructs were transformed into A. tumefaciens strain GV3101 harboring the pSoup helper plasmid (Weidi Bio, catalog number AC1002). Agrobacterium cells were grown to an optical density at 600 nm (OD600) of 0.8 and then resuspended in an infection buffer (10 mM MES, 10 mM MgCl2, 200 μM acetosyringone). Agrobacterium suspensions harboring 35S-MaLBD50 and ProMaBMY1-LUC were mixed and infiltrated into tobacco leaves using a needleless syringe. A mixture of Agrobacterium carrying the empty pSuper1300 vector and ProMaBMY1-LUC served as the negative control. Plants were kept in the dark for 1 d and then transferred to light conditions for 2 d. Luciferase activity was visualized using a live plant imaging system. The primers used in this study are listed in Supplementary Table S1.
Data analysis
-
All experiments were performed with at least three biological replicates. Data are presented as the mean ± standard deviation (SD). Statistical analysis and graphing were performed using GraphPad Prism 8.0. Differences between two groups were analyzed using Student's t-test. A p-value < 0.05 was considered to be statistically significant, and p < 0.01 was considered to be highly significant.
Accession numbers
-
The genes and their accession numbers used in this study are listed as follows: MaLBD2 (Macma4_01_g14910), MaLBD4 (Macma4_02_g14730), MaLBD16 (Macma4_03_g24120), MaLBD17 (Macma4_03_g28560), MaLBD19 (Macma4_03_g32340), MaLBD20 (Macma4_03_g32950), MaLBD21 (Macma4_04_g03960), MaLBD22 (Macma4_04_g07000), MaLBD23 (Macma4_04_g14370), MaLBD24 (Macma4_04_g14720), MaLBD30 (Macma4_04_g37560), MaLBD32 (Macma4_04_g41410), MaLBD50 (Macma4_07_g25060), MaAMY3 (Macma4_08_g04000), MaBMY1 (Macma4_05_g10290), MaEXPA8 (Macma4_03_g09830), MaLINV1 (Macma4_08_g23570), AtLBD10 (AT2G23660), AtLBD16 (AT2G42430), AtLBD37 (AT5G67420), OsLBD31 (LOC_Os10g07510), OsLBD4 (LOC_Os0g45750), OsLBD35 (LOC_Os03g41600), VvLBD11 (VIT_07s0129g00330).
-
In total, 77 MaLBD transcription factors were identified in the banana genome and designated MaLBD1 to MaLBD77 according to their chromosomal positions (Fig. 1a). They were unevenly distributed across 11 chromosomes, with chromosome 4 harboring the most genes and chromosome 1 the fewest (Fig. 1a). The physicochemical properties of MaLBD proteins were analyzed and organized (Supplementary Table S2). The theoretical molecular weights of MaLBDs ranged from 9.87 to 111.50 kDa. Among them, 43 MaLBDs were predicted to be basic (pI > 7), but 34 were acidic (pI < 7). Hydropathy analysis showed that most MaLBDs were characterized as unstable and hydrophilic proteins (GRAVY < 0), only MaLBD10, -15, -26, -36, -43, -46, and -75 were predicted to be hydrophobic.
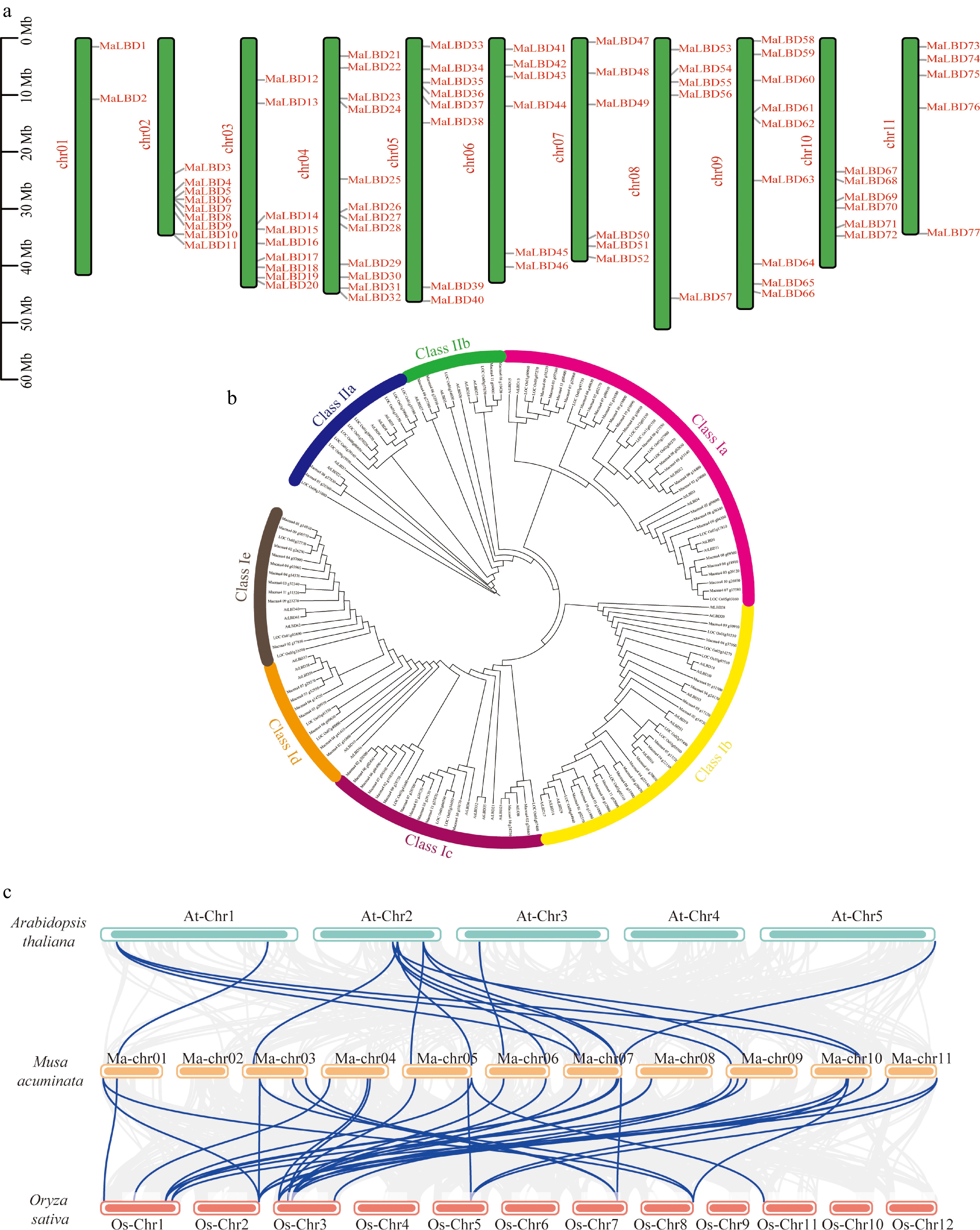
Figure 1.
Identification of MaLBD gene family members. (a) Chromosomal positions of MaLBD genes in banana. The vertical axis indicates chromosome length, chr01–11 denote chromosome numbers. (b) Phylogenetic tree for banana, A. thaliana, and rice LBD proteins. Differently colored rings represent distinct subclasses (Class Ia, Class Ib, Class Ic, Class Id, Class Ie, Class IIa, and Class IIb). (c) Synteny analysis of LBD genes among banana, A. thaliana, and rice. Gray lines indicate genome-wide syntenic gene pairs, and blue lines indicate syntenic gene pairs within the LBD gene family. Blue, yellow, and orange represent the chromosomes of A. thaliana, banana, and rice, respectively.
To investigate the evolutionary relationships of LBD transcription factors across species, we constructed a NJ phylogenetic tree using LBD proteins from banana (77), A. thaliana (41), and rice (35) (Fig. 1b). All LBD proteins were classified into two classes (Class I and Class II), with Class I further divided into five subclasses (Ia–Ie) and Class II into two subclasses (IIa and IIb). Synteny analysis was performed between banana and the other two species (Fig. 1c). In total, 15 homologous LBD gene pairs were identified between banana and A. thaliana, and 33 pairs between banana and rice (Supplementary Table S3). Among them, rice chromosome 3 exhibited the strongest collinearity with banana LBD genes. Moreover, banana and rice shared more than twice as many syntenic LBD gene pairs as banana and A. thaliana, indicating a stronger conservation of the LBD gene family between these two monocot species.
Analysis of chromatin accessibility, transcription factor binding motifs, and cis-acting elements in MaLBD promoters
-
To investigate the open chromatin regions of MaLBDs' promoters and their potential regulatory factors during fruit ripening, chromatin accessibility and transcription factor binding site analyses were performed (Fig. 2a). Among 77 MaLBDs, 9 genes (MaLBD4, MaLBD19, MaLBD20, MaLBD21, MaLBD22, MaLBD23, MaLBD24, MaLBD32, and MaLBD50) showed chromatin accessibility in their promoter regions at Stage 4 (S4, fully ripe). Within these accessible regions, binding motifs corresponding to 30 transcription factor families were identified, including AP2/ERF, WRKY, MYB, and bZIP.
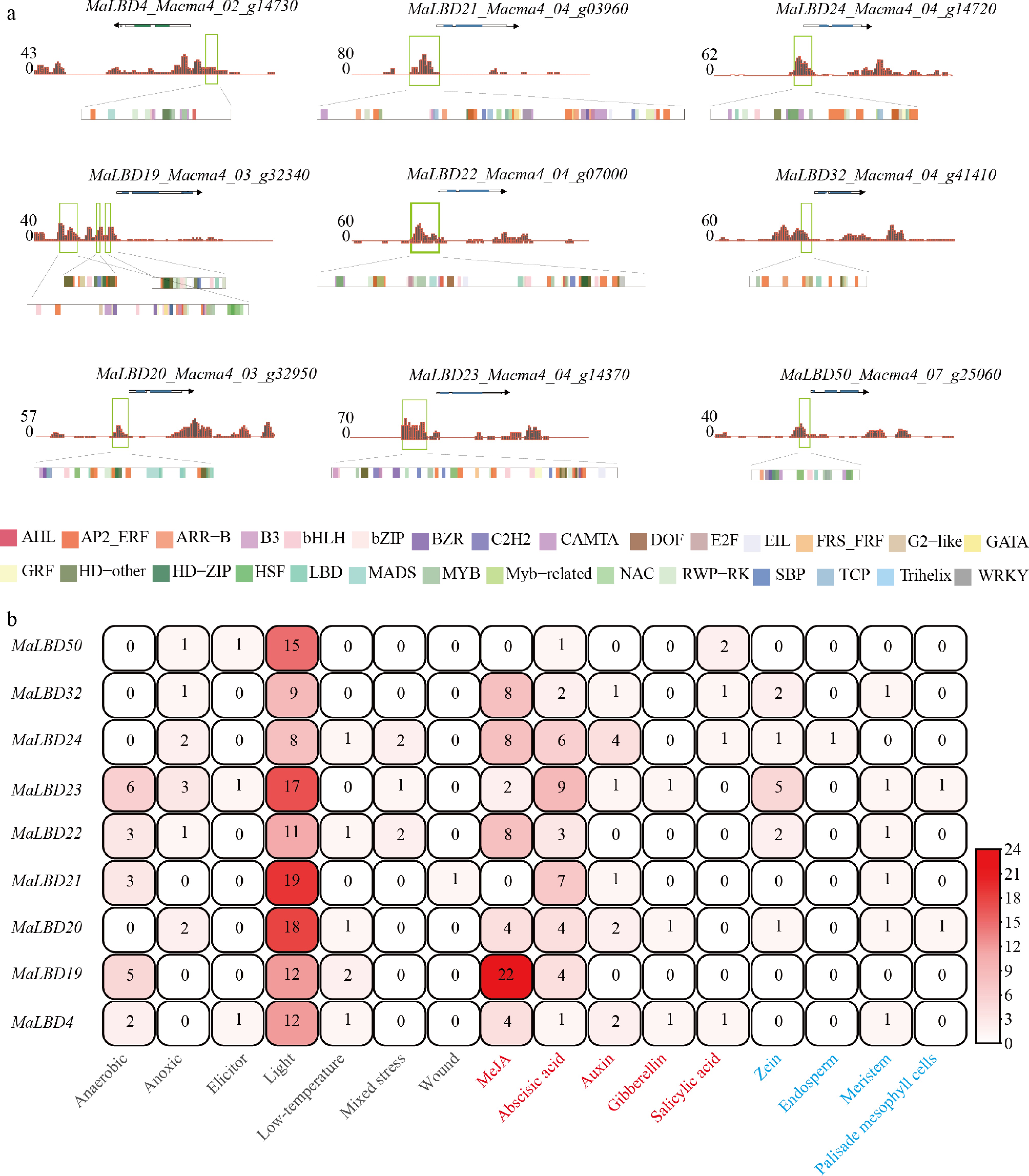
Figure 2.
Analysis of MaLBD gene promoters. (a) Chromatin accessibility and transcription factor-binding motif analysis. Brown peaks indicate DNase-seq signal intensity (open chromatin). Colored rectangles represent predicted transcription factor-binding motifs (color-coded by family). (b) Heatmap of cis-acting element numbers in MaLBD gene promoters. The color scale indicates the number of elements.
The promoter regions of nine MaLBD genes were further analyzed using the PlantCARE database to identify cis-acting elements (Fig. 2b; Supplementary Table S4). In total, 16 types of cis-acting elements were identified and categorized into three groups: Environmental response elements, phytohormone response elements, and tissue- or development-specific elements. Among the 16 types of cis-acting elements, light-responsive elements were the most abundant and were present in all MaLBD promoters. In the case of hormone-related elements, abscisic acid-responsive elements were the most common in MaLBD's accessible promoter regions. Together, these results suggest that MaLBD transcription factors may contribute to fruit ripening in banana through light- and abscisic acid (ABA)-mediated signaling pathways.
Differential expression analysis of MaLBD genes during banana fruit ripening
-
In this study, we evaluated the expression levels of MaLBD genes in banana pulp during the fruit development and ripening stages by analyzing RNA-seq data from a public database (fruitENCODE project)[12]. According to the experimental records of the original publication, the four stages are characterized as follows: S1 (fruit set, ~30 d after flowering, small green fruit with dense pulp); S2 (immature, ~60 d after flowering, larger green fruit, pulp begins to accumulate starch); S3 (mature green, ~90 d after flowering, full-size green fruit, pulp firm with a high starch content); S4 (fully ripe, 5–7 d after ethylene treatment, yellow peel, soft and sweet pulp with starch largely converted to sugars). Overall, MaLBD genes showed relatively low basal expression levels at S1, with MaLBD7 being the most highly expressed. Compared with S1, the expression levels of MaLBD2, MaLBD3, MaLBD19, MaLBD20, MaLBD21, MaLBD22, and MaLBD23 were significantly upregulated at S2 (immature), suggesting that these genes may be involved in fruit development and expansion. Furthermore, when we compared S3 (mature green) with S4 (fully ripen), nine MaLBD genes (MaLBD2, MaLBD4, MaLBD16, MaLBD17, MaLBD23, MaLBD24, MaLBD30, MaLBD50, and MaLBD75) exhibited significantly higher expression levels at S4 (Fig. 3a; Supplementary Table S5).
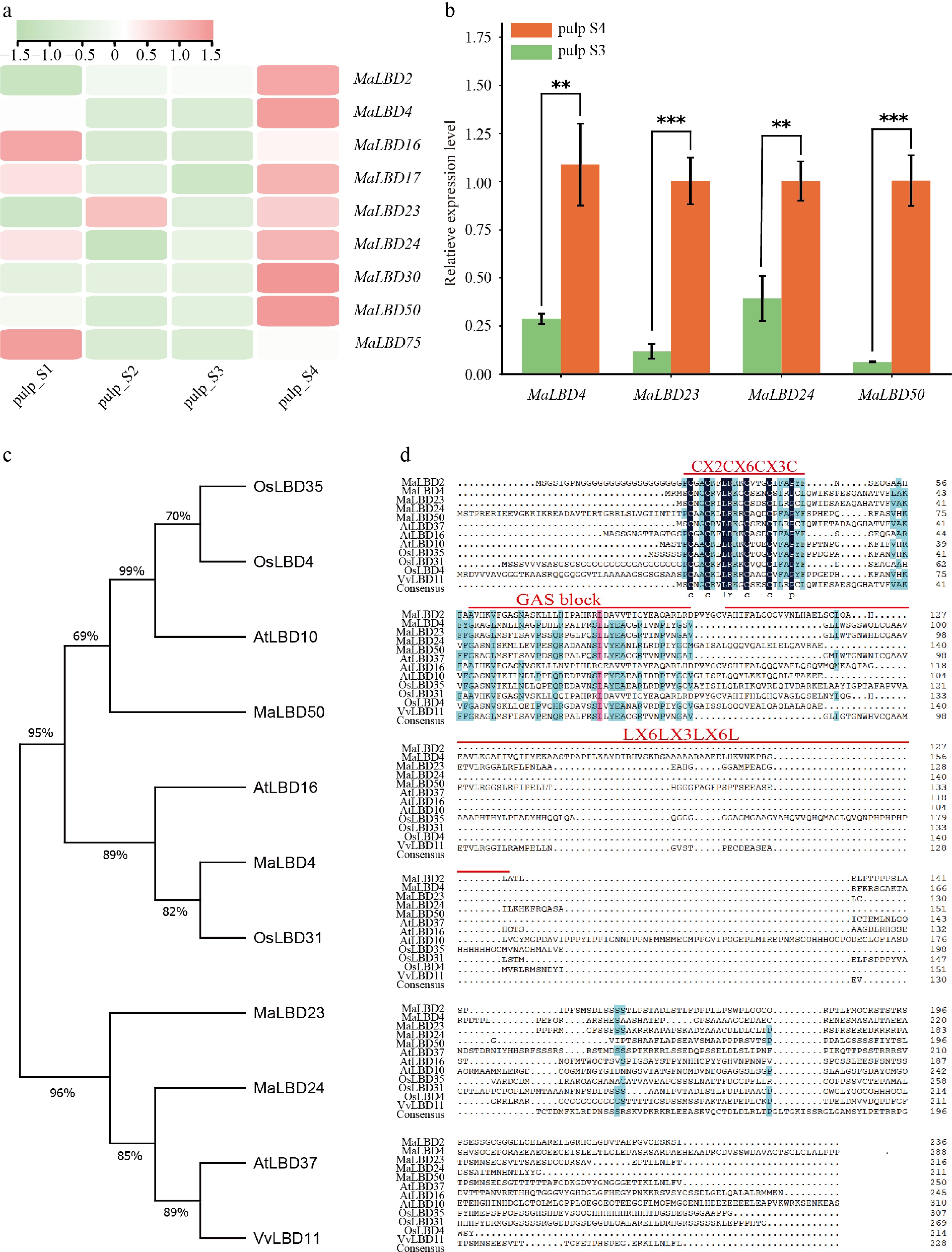
Figure 3.
Expression level of MaLBD genes during banana fruit ripening. (a) Expression heatmap of selected MaLBD genes during banana fruit development and ripening. Stage 1 (S1, fruit set), small green fruit; S2 (immature), larger green fruit; S3 (mature green), full-size green fruit, firm pulp; S4 (fully ripe), yellow peel, soft pulp. Detailed phenotypic descriptions are provided in the main text. (b) Expression levels of MaLBD4, MaLBD23, MaLBD24, and MaLBD50 at S3 and S4 as determined by qRT-PCR. Student's t-test was used for statistical analysis. ** p < 0.01, *** p < 0.005. Data are presented as the mean ± SD (n = 3). (c) Phylogenetic tree of MaLBD4, MaLBD23, MaLBD24, and MaLBD50 from Musa acuminata; AtLBD10, AtLBD16, and AtLBD37 from Arabidopsis thaliana; OsLBD31, OsLBD4, and OsLBD35 from Oryza sativa; and VvLBD11 from Vitis vinifera. (d) Multiple sequence alignment of these LBD proteins.
In addition, we intersected these nine highly expressed MaLBD genes with the nine genes showing promoter chromatin accessibility (MaLBD4, MaLBD19, MaLBD20, MaLBD21, MaLBD22, MaLBD23, MaLBD24, MaLBD32, and MaLBD50) at S4, and identified four candidate LBD transcription factors (MaLBD4, MaLBD23, MaLBD24, and MaLBD50) potentially involved in the regulation of ripening. The expression patterns of these four genes were subsequently validated by qRT-PCR (Fig. 3b), Compared with the preclimacteric levels at S3, the expression levels of these four genes were significantly upregulated at S4, which showed high consistency with the RNA-seq results. Furthermore, amino acid sequence alignment demonstrated that the MaLBD4, MaLBD23, MaLBD24, and MaLBD50 proteins contain the conserved C-block, GAS motif, and leucine zipper-like block (Fig. 3d), confirming their identity as typical LBD family members.
MaLBD50 promotes banana fruit ripening
-
To investigate the regulatory functions of MaLBD genes in banana fruit ripening, Agrobacterium-mediated transient overexpression and silencing assays were performed in banana pulp for MaLBD4, MaLBD24, and MaLBD50. MaLBD23 (also known as MaLOB41) was excluded from this experiment, as its role in promoting fruit ripening has been previously reported[6]. Phenotypic analysis revealed that MaLBD50 acts as a positive regulator of ripening (Fig. 4a). Consistent with its increased expression during ripening, transient overexpression of MaLBD50 significantly accelerated fruit ripening compared with the control, whereas it was delayed in MaLBD50-silenced bananas. To validate the efficiency of the transient transformation, qRT-PCR was performed. The results showed that expression levels of MaLBD50 were upregulated approximately 3.3-fold in the overexpression (OE) lines and significantly downregulated in the silenced lines compared with the control, confirming successful manipulation of MaLBD50 expression (Fig. 4b). Consistent with these gene expression changes, starch content analysis showed a 32.6% decrease in the MaLBD50-OE pulp compared with the control, but it increased by 9.9% in the silenced pulp (Fig. 4c). Taken together, these results demonstrate that MaLBD50 positively promotes banana fruit ripening through enhancing starch hydrolysis.
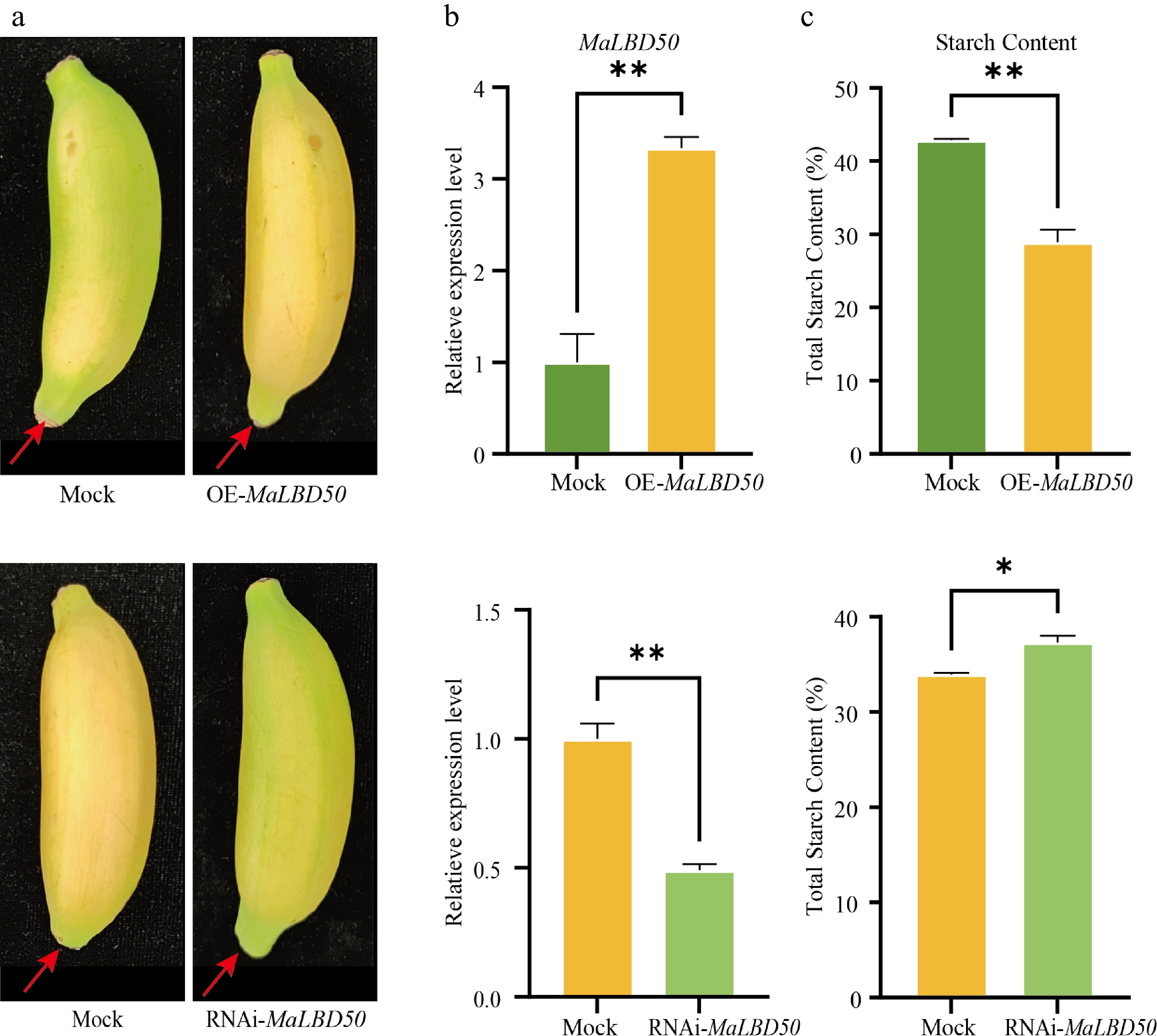
Figure 4.
Effects of MaLBD50 transient transformation on banana fruit ripening. (a) Phenotypes of banana fruits after transient transformation of MaLBD50. Red arrows indicate the injection site of the infiltration solution. (b) Expression levels of MaLBD50 after transient transformation as determined by qRT-PCR. (c) Starch content in the pulp of banana after transient transformation of MaLBD50. Student's t-test was used for statistical analysis. * p < 0.05, ** p < 0.01. Data are presented as the mean ± SD (n = 3).
Genome-wide identification of MaLBD50-targeted genes during banana ripening
-
To further elucidate the regulatory mechanism of MaLBD50 during banana fruit ripening, DAP-seq was used to identify the genome-wide binding sites of MaLBD50 (Supplementary Table S6). Motif enrichment analysis revealed that MaLBD50 specifically recognizes the motif 5'-CGGCGGCGGCGG-3' (Fig. 5a), which contains a classical LBD binding motif 5'-(G)CGGC(G)-3', as reported previously[40]. The high consistency of enriched motifs between two independent biological replicates confirmed the reliability of the DAP-seq data. Genomic distribution analysis showed that MaLBD50's binding sites were unevenly distributed, with 28.39% of the peaks located in gene promoter regions (Fig. 5b). By merging data from the two replicates, 7,813 high-confidence target genes were identified in total (Fig. 5c). Among these, 194 genes were significantly upregulated and 268 were significantly downregulated at the transcriptional level during ripening (Fig. 5d; Supplementary Table S7).
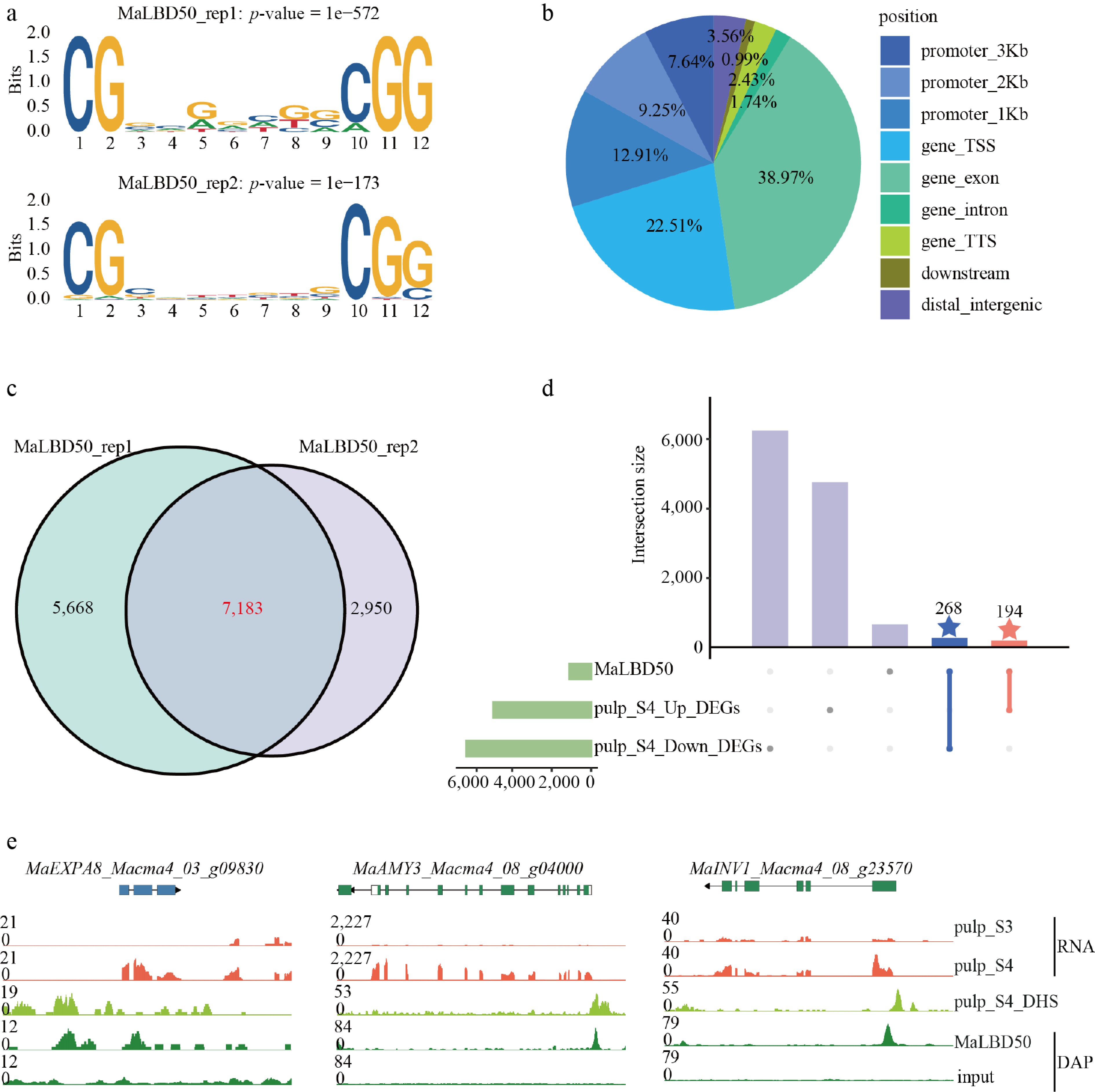
Figure 5.
Regulatory model of MaLBD50 during banana fruit ripening. (a) Binding motif of MaLBD50 revealed by DAP-seq data; rep1 and rep2 represent two biological replicates. (b) Peak distribution of MaLBD50 DAP-seq data. (c) Venn diagram of MaLBD50 target genes identified from two biological replicates. (d) UpSet plot showing overlaps between MaLBD50 target genes and upregulated (Up_DEGs) or downregulated (Down_DEGs) differentially expressed genes in ripening pulp (Stage 4, S4). (e) Dynamic changes in expression and chromatin accessibility of MaLBD50 target genes (MaEXPA8, MaAMY3, and MaINV1) in pulp during banana fruit ripening.
By integrating DAP-seq, DNase-seq, and RNA-seq data, three key ripening-related genes were identified as potential targets of MaLBD50: MaEXPA8, MaAMY3, and MaINV1. These genes exhibited significantly upregulated expression levels and high chromatin accessibility in their promoter regions at S4. Functionally, MaEXPA8 encodes an expansin involved in cell walls loosening and softening; MaAMY3 encode amylases that facilitate starch hydrolysis; and MaINV1 encodes an invertase affecting sucrose accumulation. Integrative visualization revealed that MaLBD50 may systematically drive the ripening transition from S3 to S4 by coordinately regulating starch degradation, cell wall remodeling, and sugar metabolism pathways (Fig. 5e).
Open chromatin regions and MaLBD50-binding sites were also detected in the promoters of MaLBD2 and MaLBD4 (Supplementary Fig. S1). This suggests that MaLBD50 may not only influence fruit ripening by directly regulating downstream ripening marker genes but also forms a transcriptional network that coordinately controls the ripening process.
MaLBD50 promotes starch degradation by regulating MaBMY1
-
In addition to the three genes mentioned above, this study also identified a β-amylase gene, MaBMY1, which is regulated by MaLBD50. This gene is upregulated during the S4 fruit ripening stage, and its promoter region contains a chromatin-accessible region (Fig. 6a). DAP-qPCR analysis further revealed that MaLBD50 is significantly enriched in the chromatin-accessible region of the MaBMY1 promoter (Fig. 6b).
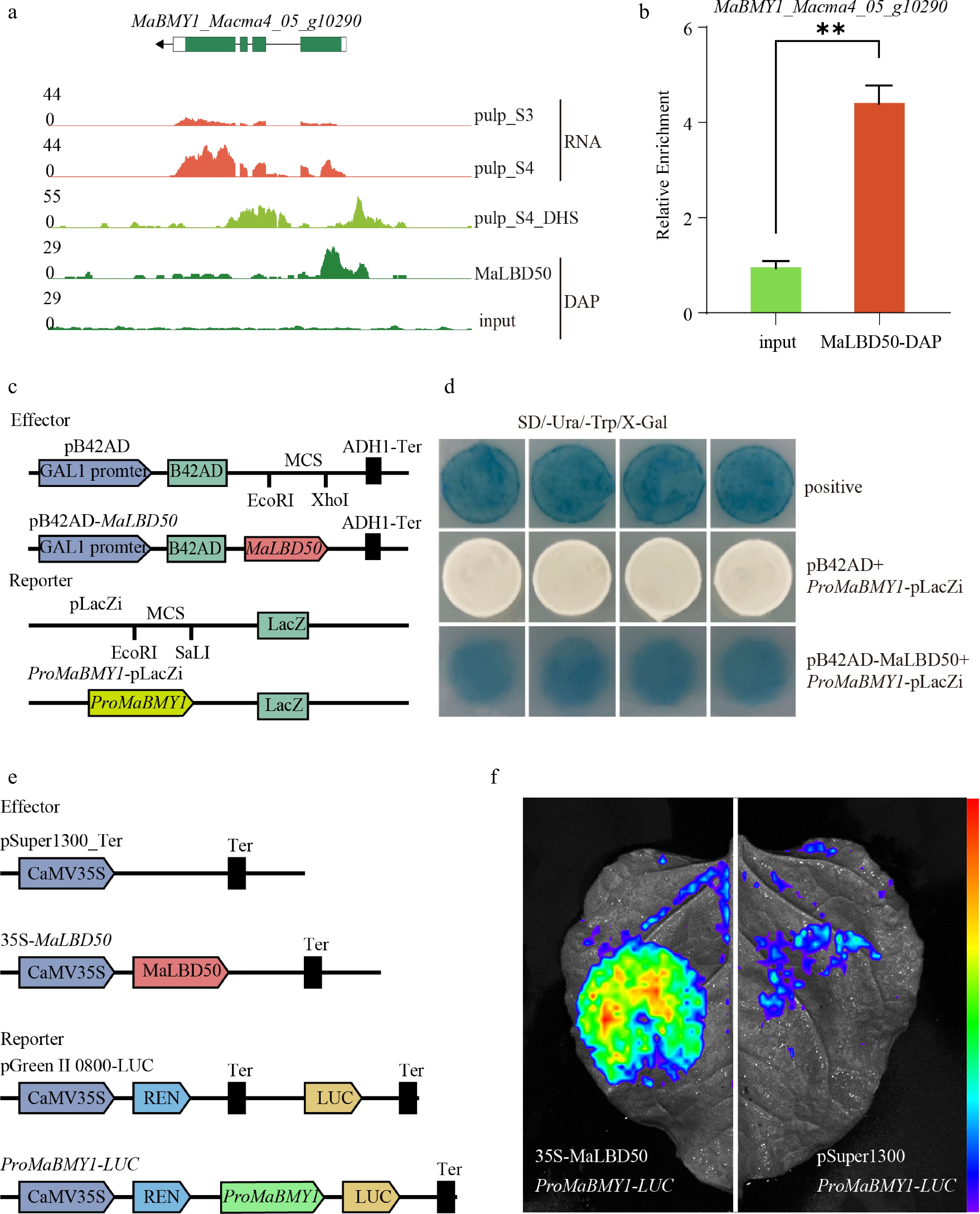
Figure 6.
Regulation of MaBMY1's expression by MaLBD50. (a) Expression levels, promoters' chromatin accessibility, and MaLBD50-binding sites of MaBMY1. (b) Enrichment of MaLBD50 binding to the MaBMY1 promoter as determined by DAP-seq qPCR. (c) Schematic diagram of yeast Y1H assays and vector construction. (d) Validation of MaLBD50 binding to the promoters of MaBMY1 by the Y1H assay. (e) Schematic diagram of the DLR reporter vector constructs. (f) Luciferase (LUC) activity detected by live imaging in tobacco leaves. Student's t-test was used for statistical analysis. ** p < 0.01. Data are presented as the mean ± SD (n = 3).
Y1H assays were conducted to verify the direct binding of MaLBD50 to the promoters of MaBMY1 genes. The CDS of MaLBD50 was fused to the GAL1 activation domain in the pB42AD vector, and the promoter fragments of MaBMY1 were cloned into the pLacZi vector (Fig. 6c). Yeast cells co-transformed with pB42AD-MaLBD50 and ProMaBMY1-pLacZi exhibited robust growth and turned blue on synthetic complete (SC)-Trp-Ura medium supplemented with X-Gal. In contrast, cells co-transformed with the empty vector pB42AD and the ProMaBMY1-pLacZi construct showed no β-galactosidase activity. These results demonstrate that MaLBD50 specifically binds to the promoter regions of MaBMY1 (Fig. 6d). We then performed the DLR assay to verify the transcriptional regulatory activity of MaLBD50 on MaBMY1. Strong luminescence signals were detected in tobacco leaves co-transformed with the effector 35S-MaLBD50 and the reporter ProMaBMY1-LUC (Fig. 6f). These results provide further evidence that MaLBD50 directly binds to the MaBMY1 promoter and significantly activates its transcription. Consistent with this, starch content was significantly reduced in the MaLBD50-OE lines, whereas the MaLBD50-RNAi lines exhibited the opposite phenotype (Fig. 4c).
In conclusion, by combining molecular mechanistic insights with functional validation, this study systematically elucidates the core regulatory role of MaLBD50 as a key transcription factor in banana fruit ripening. These findings provide a theoretical basis and potential gene targets for the precise regulation of fruit ripening and the molecular breeding of banana varieties with improved shelf life.
-
The LBD family represents a class of plant-specific transcription factors that primarily regulate vegetative growth processes, such as lateral root development, leaf morphogenesis, and inflorescence development[29−34]. In this study, we screened the MaLBD gene family through genome-wide analysis and identified MaLBD50 as a core transcription factor regulating banana fruit ripening. Recent studies have also highlighted the roles of LBD transcription factors in fruit development and ripening[33,35,41,42]. In tomato (Solanum lycopersicum), the transcription factor SlMYC2 activates SlLBD40's expression and forms a protein complex with SlLBD40 to synergistically activate the SlEXPA5 gene, thereby promoting cell expansion and fruit size[35]. In grape, VvLBD11 and VvLBD28 reduce anthocyanin accumulation via three distinct pathways: Repressing the expression of anthocyanin biosynthetic genes, disrupting the MYB-bHLH-WD40 transcriptional complex, and blocking transporter function[33]. Additionally, PpLOB1 positively regulates peach (Prunus persica) fruit softening by directly activating the pectin lyase genes PpPL1 and PpPL15[41]. In this study, MaLBD50 promotes fruit ripening and starch degradation by directly binding to the promoters of MaAMY3 and MaBMY1, and specifically activates MaBMY1's expression. This functional divergence may be an adaptation to the distinct ripening physiology of banana as a starch-rich climacteric fruit. Besides MaLBD50, other transcription factors such as MaERF012, MaERF95L, MaERF96L, MaABI5-like, and MabZIP21 also regulate starch degradation in banana through diverse mechanisms[8,10,19,20,43,44]. For instance, the MaBEL1–MaABI5-like–MaEBF1 complex synergistically activates genes involved in cell wall and starch degradation, thereby promoting fruit softening[8,16]. Additionally, MaMPK6-3 phosphorylates MabZIP21, enhancing its binding affinity and transcriptional activation of MaGWD1 and MaAMY3[20,44]. Collectively, these findings contribute to a complex transcriptional regulatory network governing starch degradation during banana fruit ripening.
Integrated multiomics analysis has emerged as a powerful approach for dissecting biological problems[4,45,46]. In soybean (Glycine max), a genome-wide transcriptional regulatory network (SoyGRN) was constructed by integrating the DAP-seq profiles of 148 transcription factors with assay for transposase-accessible chromatin using sequencing (ATAC-seq), ChIP-seq, and RNA-seq data[45]. In apple (Malus domestica), combined ATAC-seq and RNA-seq analyses demonstrated that five DOF transcription factors (MdOBP1-like1/2, MdADOF1-like, MdOBP2-like, and MdOBP3-like) positively regulate sugar and acid accumulation by directly activating the expression of specific metabolic target genes[4]. In tea (Camellia sinensis) plants, the integration of ATAC-seq, RNA-seq, and DAP-seq data uncovered the molecular mechanisms underlying heterosis in oolong tea[46]. In this study, the high chromatin accessibility within the promoter regions of MaLBD50's target genes at ripening stage provided the structural basis for its binding. Furthermore, the observation that 28.39% of DAP-seq binding peaks were located in promoter regions further confirms the close association between chromatin's accessibility and transcription factor recruitment.
Fruit ripening is a complex biological process that involves synergistic changes in color, texture, flavor, and aroma, necessitating the coordinated action of multiple transcription factors. In the present study, our integrated analysis suggests that MaLBD50 may coordinate multiple ripening-associated pathways; however, direct evidence is currently confined to its regulation of starch degradation via MaBMY1. The roles of MaLBD50 in fruit softening and flavor formation remain to be directly quantified, and no color change-related genes were identified as its potential targets in the DAP-seq dataset. Nevertheless, we observed that MaLBD50 directly binds to the promoters of MaLBD2 and MaLBD4, implying the existence of an intrafamily regulatory network. For instance, MaSPL16 promotes fruit ripening by directly activating MaNAC029 (MaNAP1)[14]. Similarly, MaEIL4 activates the expression of MaMADS36, and MaMADS36 subsequently activates the expression of MaACS7, creating a positive feedback loop of ethylene signaling that accelerates ripening[22]. Conversely, MaSINAT5 negatively regulates the MaABI5-like–MaERF113 module by promoting the ubiquitin-mediated degradation of MaERF113, thereby suppressing the expression of starch and chlorophyll degradation genes and delaying fruit ripening[8]. These findings suggest that the MaLBD50 regulatory module revealed in this study likely operates within a larger network, forming hierarchical or parallel regulatory relationships with NAC, MYB, SPL, and other family members—a possibility that warrants further investigation.
As a key positive regulator of banana fruit ripening, MaLBD50 represents a promising target for molecular breeding. Using clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated protein technique to knockout or knockdown MaLBD50 expression could effectively delay fruit ripening. Conversely, overexpression of this gene may accelerate ripening, promoting starch degradation and sugar accumulation. Similar strategies have been successfully applied in various fruits: CRISPR editing of FaPG1 in strawberry (Fragaria × ananassa) significantly improved fruit firmness[47]; knockout of SlPL and SlPG in tomato delayed softening process[48]; and editing of ClNAC68 in watermelon (Citrullus lanatus) reduced the accumulation of sugar. However, the pleiotropy of transcription factors' regulatory networks requires us to consider potential side effects during their application[49]. Therefore, safer strategies include using tissue-specific or inducible promoters to drive gene editing, or using promoter editing to achieve fine-tuned regulation. Furthermore, allele mining based on natural variation is another important direction; for instance, an insertion–deletion variation in the promoter region of apple's MdNAC18.1 has been shown to be significantly associated with ripening time[50].
-
In summary, this study systematically identified the banana LBD transcription factor family and revealed the molecular mechanism by which MaLBD50 promotes fruit ripening via direct activation of the starch degradation gene MaBMY1. In addition, it revealed the potential regulatory effects of MaLBD50 on other ripening-associated genes, including MaEXPA8, MaAMY3, and MaINV1, thereby expanding our understanding of LBDs' functions in fruit ripening. By integrating DAP-seq, DNase-seq, and RNA-seq data, we established a link among transcription factors, chromatin accessibility, and gene expression, providing a paradigm for dissecting the epigenetic regulation of fruit ripening. Future research should focus on (1) whether MaLBD50 is subject to epigenetic modifications, (2) the synergistic regulatory networks between MaLBD50 and other transcription factors, and (3) the development of precision regulation strategies based on gene editing. These endeavors will deepen our understanding of the molecular mechanisms governing fruit ripening and provide theoretical support and technical tools for breeding new banana varieties with high quality and extended shelf life.
-
During the preparation of this manuscript, the authors used Deepseek V3.2 to assist in improving the clarity and readability of the language. All content generated was carefully reviewed and revised by the authors as necessary, and the authors take full responsibility for the final content of the article.
-
The authors confirm their contributions to the paper as follows: project administration: Lü P, Chen Z, Huang J; study conception and design, draft manuscript preparation and revision: Jiang Y, Li T; experimental work and data analysis: Jiang Y, Cai D, Li T, Dong D, Fan X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The work was supported by the National Natural Science Foundation of China (32402517, 32372666), the Natural Science Foundation of Hainan Province (326JCQN0983), the Project of State Key Laboratory of Tropical Crop Breeding (NKLTCBCXTD26 and NKLTCB-RC202402), and the earmarked fund for China Agriculture Research System (CARS-31).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at https://doi.org/10.48130/tp-0026-0026.
-
Received 16 April 2026; Accepted 26 May 2026; Published online 17 June 2026
-
# Authors contributed equally: Youfeng Jiang, Tianxiang Li
- Supplementary Table S1 Primer information.
- Supplementary Table S2 Physical and chemical properties of proteins encoded by MaLBD family genes.
- Supplementary Table S3 Collinear gene pair.
- Supplementary Table S4 The expression patterns of MaLBD genes during banana fruit development (stages S1−S2) and ripening (stages S3−S4).
- Supplementary Table S5 Cis-acting element information.
- Supplementary Table S6 Statistics of DAP-seq libraries.
- Supplementary Table S7 Upset analysis of MaLBD50 target genes and differentially expressed genes in fruit pulp at ripening stage.
- Supplementary Fig. S1 Identification of MaLBDs genes targeted by MaLBD50.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jiang Y, Li T, Cai D, Fan X, Dong D, et al. 2026. MaLBD50 directly activates MaBMY1 to promote starch degradation during banana fruit ripening. Tropical Plants 5: e021 doi: 10.48130/tp-0026-0026 

MaLBD50 directly activates MaBMY1 to promote starch degradation during banana fruit ripening
- Received: 16 April 2026
- Revised: 15 May 2026
- Accepted: 26 May 2026
- Published online: 17 June 2026
Abstract: The lateral organ boundaries domain (LBD) transcription factor family plays critical roles in plant growth and development. However, its function in banana (Musa spp.) fruit ripening remains poorly understood. Here, we performed a genome-wide identification of the LBD gene family in banana and investigated the function and regulatory mechanism of MaLBD50 during fruit ripening. Transient overexpression of MaLBD50 accelerated banana fruit ripening and enhanced starch degradation, whereas transient silencing produced the opposite effects. By integrating RNA sequencing (RNA-seq), DNase I hypersensitive site sequencing (DNase-seq), and DNA affinity purification sequencing (DAP-seq) data, we identified four ripening-associated genes (MaBMY1, MaAMY3, MaEXPA8, and MaINV1) as putative MaLBD50 targets, these genes showed higher expression level and increased promoter chromatins' accessibility at the ripening stage. MaLBD50 was shown to directly bind to and activate the expression of MaBMY1, using yeast one-hybrid and dual-luciferase assays. Collectively, these results indicate that MaLBD50 functions as a positive regulator of banana fruit ripening by promoting the expression of ripening-related genes, highlighting MaLBD50 as a potential target for molecular breeding in bananas.
-
Key words:
- Banana /
- LBD /
- Fruit ripening /
- Starch degradation /
- Chromatin accessibility /
- DAP-seq