








-
Soil salinization represents a substantial challenge to global agricultural productivity. In China, saline soils constitute 4.88% of the total arable land, ranking among the most severe abiotic stress factors affecting crop cultivation[1]. The development of saline soils is influenced by various factors, including the high salt content inherent in parent soil materials, an imbalance where evaporation surpasses precipitation, and suboptimal irrigation techniques. These conditions contribute to the excessive accumulation of sodium salts in the soil, leading to osmotic stress, ion toxicity, and oxidative damage in plants. Consequently, plant growth is markedly hindered, manifesting as stunted development, a reduction in leaf number, diminished yields, and, in extreme cases, total crop failure[2]. In coastal tidal flat regions, prolonged exposure to oceanic climatic influences has exacerbated soil salinization in island reef areas. This progressive degradation has severely impeded regional agricultural expansion and posed limitations on vegetable cultivation. Affected by this, the vegetable supply of the island residents depends on the mainland, but the island reefs in the South China Sea are in an area of frequent typhoons, and transportation is often interrupted. It is of great significance to select the vegetables suitable for the high-salt soil of the island reefs in the South China Sea to ensure the living standard of the island residents.
Gynura divaricata, a widely distributed wild vegetable in Hainan province, is scientifically classified as Gynura japonica and belongs to the Gynura genus within the Asteraceae family. This species is a perennial herbaceous plant[3]. The historical presence of Gynura divaricata in Hainan province dates back to the wartime period of the 19th century. Notably, it served as a primary food source for soldiers stationed at the Wuzhi Mountain revolutionary base, which led to its designation as the 'Revolutionary Vegetable'[4]. The edible and medicinal properties of Gynura divaricata extend across its roots, stems, leaves, flowers, and fruits, positioning it as a significant component of traditional Chinese herbal medicine. Historically, this plant has been acknowledged for its therapeutic potential in the treatment of scurvy, detoxification, cancer prevention, and antioxidative applications[5]. Contemporary pharmacological research has further explored its bioactive properties, revealing that multiple compounds found in Gynura divaricata exhibit efficacy in mitigating and managing ailments such as hypertension, hyperglycemia, hyperlipidemia, and diabetes[6−8].
A vegetable resembling Gynura divaricata was previously identified on the reefs and islands of the South China Sea and was designated as Sansha Gynura divaricata. This novel variety exhibits several advantageous traits: its edible components possess an enhanced texture due to increased succulence, it neither flowers nor produces fruit, maintains an extended vegetative growth phase, and demonstrates remarkable resilience to environmental stress. Adaptation to the extreme conditions of Sansha—characterized by high temperatures, humidity, and salinity—allows it to thrive throughout the year without seasonal constraints, facilitating multiple harvests. Given its growth in saline atmospheric conditions on the reefs and islands of the South China Sea, Sansha Gynura divaricata may have undergone epigenetically regulated genetic modifications[9]. Although its high salt tolerance is evident, research concerning its salt tolerance mechanisms and nutritional composition remains insufficient. Therefore, to further investigate island reef vegetable resources and assess the salt tolerance capacity of Sansha Gynura divaricata, gradient NaCl treatments were administered to evaluate its growth and physiological responses, thereby determining its threshold for salt tolerance. Furthermore, the nutritional profile of its edible parts was analyzed. This study establishes a theoretical foundation and serves as a reference for the large-scale cultivation of Sansha Gynura divaricata on the South China Sea islands, offering significant implications for overcoming the challenges associated with vegetable production in these saline-affected environments.
-
The experimental materials, Gynura divaricata, were sourced from Wuzhishan city, Hainan Province, while the Sansha Gynura divaricata germplasms were acquired from Sansha city, Hainan Province, China. The experiment was carried out within a temperature-controlled greenhouse under natural light facility at the College of Coastal Agriculture, Guangdong Ocean University, Guangdong, China. Stem segments, each measuring 5 cm in length and comprising a pair of axillary buds along with two nodes, were excised from vigorously growing Sansha Gynura divaricata seedlings. These segments underwent an initial washing with tap water three times, followed by surface sterilization using 75% ethanol for 5 min. Subsequently, they were rinsed thrice with distilled water. The prepared segments were then placed into a cultivation substrate formulated from an equal proportion (1:1) of nutrient soil and vermiculite. After undergoing a 2-week culture period in a medium supplemented with 1/2 Hoagland nutrient solution, seedlings exhibiting uniform and healthy growth were selected for transplantation. Each pot was filled with 2 kg of the previously mentioned substrate, and three seedlings were transplanted into each pot. After transplantation, standard irrigation and fertilization protocols were applied to the Sansha Gynura divaricata seedlings. The experimental procedures commenced 23 d following transplantation.
Experimental design
-
Four salinity gradient treatments were established as follows: (1) Control (CK) with 1/2 Hoagland nutrient solution containing 0 mmol·L−1 NaCl; (2) 100 mM treatment with 1/2 Hoagland nutrient solution supplemented with 100 mmol·L−1 NaCl; (3) 200 mM treatment with 1/2 Hoagland nutrient solution containing 200 mmol·L−1 NaCl; and (4) 400 mM treatment with 1/2 Hoagland nutrient solution supplemented with 400 mmol·L−1 NaCl. Each treatment was conducted in quadruplicate, with 12 plants assigned to each condition, resulting in a total of 48 plants. Throughout the duration of the salt stress experiment, quantitative irrigation was administered every 3 d using the respective saline solutions to mitigate salt buildup and sustain stable salinity levels. After 23 d of salt exposure, samples were collected for morphological assessment. Additionally, specimens designated for nutritional and physiological analyses, RNAseq, and metabolite analyses were promptly flash-frozen in liquid nitrogen and preserved at −80 °C until further examination.
Determination of major nutrients in Sansha Gynura divaricata
-
The parameters assessed in this study encompassed moisture, ash, protein, mineral elements, vitamins, and flavonoids. The methodologies employed for determination were as follows: GB5009 was utilized for the quantification of Moisture (GB 5009.3-2016).ash (GB 5009.4-2016), potassium and sodium (GB 5009.91-2017). Phosphorus (GB 5009.87-2016). Calcium (GB 5009.92-2016),copper, zinc, iron, magnesium, and manganese were all conducted using their respective flame atomic absorption spectrometry methods. These were based on the following standards: GB 5009.13-2017 for copper, GB 5009.14-2017 for zinc, GB 5009.90-2016 for iron, GB 5009.241-2017 for magnesium, and GB 5009.242-2017 for manganese. Protein content was analyzed via the automatic nitrogen analyzer method (NY/T2419-2013). The concentrations of vitamins B1, B2, B6, B7, niacin, folic acid, and pantothenic acid were determined through the ultra-performance liquid chromatography method[10]. Additionally, high-performance liquid chromatography was applied for the detection of gallocatechin, catechin, epicatechin, delphinidin chloride, ellagic acid, glycyrrhizin, quercetin, luteolin, and kaempferol[11].
Determination of morphological and physiological parameters of Sansha Gynura divaricata
-
Following a 23-d period of salt stress exposure, the height and root length of Sansha Gynura divaricata were recorded utilizing a metric ruler, while the fresh weight of the entire plant was quantified with a 1/10,000 electronic balance. Three independent replicates were maintained for each measurement. For enzymatic and biochemical analyses, 0.1 g of fresh plant tissue was weighed and homogenized in 1 mL of extraction buffer (0.05 M phosphate buffer, pH 7.8) using a freezing grinder. The homogenates were centrifuged at 12,000 rpm for 20 min at 4 °C, and the resulting supernatant was used as the enzyme source. Superoxide dismutase (SOD) activity was evaluated using the nitroblue tetrazolium (NBT) method[12], whereas peroxidase (POD) activity was quantified following the guaiacol method[13]. Malondialdehyde (MDA) content was analyzed through the thiobarbituric acid (TBA) method[14]. Catalase (CAT) activity was determined in accordance with the method outlined by Ekinci et al.[15], and soluble protein (SP) content was measured employing the Coomassie Brilliant Blue G-250 staining method[16].
RNA isolation, library construction, and sequencing
-
The four sample groups were designated as follows: L0 (leaves under 0 mM NaCl treatment), L2 (leaves under 200 mM NaCl treatment), R0 (roots under 0 mM NaCl treatment), and R2 (roots under 200 mM NaCl treatment). Each group comprised three biological replicates, with each replicate consisting of four individual seedlings. Total RNA was extracted using the RNA extraction kit (Tiangen, China, DP437). The quality of the RNA was assessed with an Agilent 2100 bioanalyzer, and qualified samples were then sent to the Guangzhou Gene Denovo Honour Biotechnology Co., Ltd for library construction and library sequencing on the NovaSeq 6000 platform. After data generation, the OmicShare online website (
www.omicshare.com ) was used for analysis and plotting. The KEGG online website (www.kegg.jp ) was utilized for KEGG pathway annotation and enrichment analysis of differentially expressed genes.Metabolite extraction and LC/MS analysis
-
The sample grouping setup for metabolome assays is the same as RNAseq: each group comprised six biological replicates, with each replicate consisting of four individual seedlings. A 100 mg sample was used for metabolite extraction and the extract was subsequently injected into the LC-MS assay system for analysis. The original data were screened using CD 3.3 software and compared with mzCloud (
www.mzcloud.org ), mzVault, and Masslist databases, leading to the identification and relative quantification of each metabolite. Metabolites were classified as differential if they met the criteria of VIP > 1, p-value < 0.05, and FC ≥ 2 or FC ≤ 0.5 between comparison groups. The OmicShare online website (www.omicshare.com ) was used for enrichment analysis and plotting.Data analysis
-
The experimental data were processed using Excel 2021 and SPSS 27.0 software. Subsequently, one-way analysis of variance (ANOVA) was conducted. Finally, the data was visualized using GraphPad Prism 9 software.
-
In this experiment, Gynura divaricata from Wuzhishan, Hainan, was designated as the CK sample, while Sansha Gynura divaricata was utilized as the research material. Stem segments containing two internodes with uniform morphology were selected in January and planted at a 45° angle in the high-salinity coral sand soil of Yongxing Island. Upon sample collection in March, it was observed that Gynura divaricata had reached the flowering stage, whereas Sansha Gynura divaricata remained in the vegetative growth phase, indicating distinct developmental differences. Comparatively, Gynura divaricata exhibited yellowish-green leaves, with some plants already in bloom, while Sansha Gynura divaricata displayed robust vegetative growth without flowering (Fig. 1). Analyzing individual plants revealed that Sansha Gynura divaricata possessed more tender stems and leaves, thereby contributing to a larger edible portion. Gynura divaricata typically flowers between March and April, after which plant aging occurs, leading to a decline in the quality of edible parts. In contrast, Sansha Gynura divaricata neither flowers nor produces fruit, sustains vigorous vegetative growth, and retains superior vegetable quality. Consequently, Sansha Gynura divaricata exhibits increased yield potential, reduced cultivation expenses, and enhanced vegetable quality. These attributes suggest that Sansha Gynura divaricata is more suitable for cultivation on South China Sea islands, offering both varietal and technical support for the demonstration and promotion of vegetable cultivation in high-salinity coral soils in this region.

Figure 1.
Gynura divaricate and Sansha Gynura divaricate grown on Yongxing island in Sansha city. The stem segments of common Gynura divaricate and Sansha Gynura divaricate with the same shape and two internodes were planted on Yongxing Island of Sansha. In March, the plants were photographed; (a) Sansha Gynura divaricate, (b) Gynura divaricate, and (c) branches of Sansha Gynura divaricate and common Gynura divaricate.
Nutritional components of Sansha Gynura divaricata
-
The nutritional composition of Sansha Gynura divaricata is presented in Table 1. Per 100 g of edible portions (tender stems and leaves), the water content is measured at 95.67 g, ash content at 4.33 g, and protein content at 1.26 g. Sansha Gynura divaricata is characterized by its thick, fleshy leaves and high juice content. Its protein content is moderate—lower than that of high-protein spinach, but comparable to that of common vegetables such as celery and lettuce. In terms of vitamin composition, the plant contains a comprehensive profile of B vitamins, indicating high nutritional value. Among these, folate is the most abundant, followed by vitamin B6 and biotin (B7), while thiamine (B1), riboflavin (B2), niacin, and pantothenic acid are present in relatively lower amounts. Compared with spinach, celery, and lettuce, Sansha Gynura divaricata exhibits higher folate and riboflavin levels than lettuce, and a higher vitamin B6 content than both celery and lettuce. Dietary intake of B vitamins is known to play a significant role in human health, including cancer prevention. Regarding mineral elements, Sansha Gynura divaricata is exceptionally rich. These minerals can be categorized into two groups: those that constitute intracellular components, such as phosphorus (P), zinc (Zn), and copper (Cu); and those that regulate plant physiological activities, including magnesium (Mg), calcium (Ca), and potassium (K). Among these, K and Ca are the most abundant, followed by P, sodium (Na), and Mg, with manganese (Mn), iron (Fe), Zn, and Cu present in progressively lower concentrations. Notably, the K content is more than three times that of Na, with a K/Na ratio of 3.07, classifying Sansha Gynura divaricata as a typically high-K, low-Na plant. This property aligns with its traditional medicinal use in managing hypertension and hyperlipidemia. Furthermore, the tender stems and leaves of Sansha Gynura divaricata are abundant in flavonoid compounds. A total of nine flavonoids were identified, among which epigallocatechin exhibited the highest concentration at 460.29 mg·100 g−1, followed by epicatechin, while other flavonoids were present in comparatively lower amounts.
Table 1. Nutrient composition table of Sansha Gynura divaricate.
Vitamins and other ingredients Value Mineral elements Value Flavonoids Value Water (g·100 g−1) 95.67 ± 0.45 Ca (mg·100 g−1) 3,330 ± 104.1 GC (mg·100 g−1) 460.29 ± 78.41 Ash content (g·100 g−1) 4.33 ± 0.45 P (mg·100 g−1) 1,917 ± 57.09 Cianidanol (mg·100 g−1) 0.79 ± 0.27 Protein (g·100 g−1) 1.26 ± 0.31 K (mg·100 g−1) 3,829 ± 291.33 EC (mg·100 g−1) 122.67 ± 10.91 VB1 (μg·100 g−1) 4.92 ± 1.82 Na (mg·100 g−1) 1,246 ± 231.55 Delphinidin chloride (mg·100 g−1) 3.77 ± 0.98 VB2 (μg·100 g−1) 14.11 ± 6.2 Mg (mg·100 g−1) 472 ± 2 EA (mg·100 g−1) 2.05 ± 0.77 Niacin (μg·100 g−1) 10.44 ± 1.32 Fe (mg·100 g−1) 28.55 ± 0.43 Glycyrrhizin (mg·100 g−1) 0.48 ± 0.01 Pantothenic acid (μg·100 g−1) 11.81 ± 5.79 Zn (mg·100 g−1) 3.68 ± 0.08 QR (mg·100 g−1) 1.71 ± 0.25 VB6 (μg·100 g−1) 27.9 ± 2.05 Cu (mg·100 g−1) 1.49 ± 0.21 Luteolin (mg·100 g−1) 0.81 ± 0.29 VB7 (μg·100 g−1) 17.61 ± 2.96 Mn (mg·100 g−1) 26.3 ± 0.46 Kaempferol (mg·100 g−1) 3.94 ± 4.77 FA (μg·100 g−1) 65.99 ± 18.63 VB1: vitamin B1; VB2: vitamin B2; VB6: vitamin B6; VB7: vitamin B7; FA: folic acid; Ca: calcium; P: phosphorus; K: potassium; Na: sodium; Mg: magnesium; Fe: ferrum; Zn: zinc; Cu: copper; Mn: manganese; GC: gallic catechin; EC: epicatechin; EA: ellagic acid; QR: quercetin. The data were expressed as mean ± SE of three replicates. The different letters at the top of the column indicate significant differences at p < 0.05. Analysis of salt tolerance in Sansha Gynura divaricata
-
To evaluate the salt tolerance of Sansha Gynura divaricata seedlings, growth parameters—including plant height, root length, fresh weight, and water content—were assessed under varying salt concentration gradients. As illustrated in Fig. 2, the growth of Sansha Gynura divaricata was negatively affected by salt stress, exhibiting an overall decline in plant height, root length, fresh weight, and water content as soil salinity increased. Under 100 mM and 200 mM salt treatments, plant height, root length, and fresh weight declined by 7%, 20%, and 27%, and by 22.6%, 33.5%, and 59%, respectively, compared to the CK group, whereas water content did not show a statistically significant reduction at the p < 0.05 level. Observation of Fig. 2a revealed that the leaves of Sansha Gynura divaricata remained a deep green color under both 100 and 200 mM NaCl treatments. Despite a significant reduction in plant height and root length, these parameters were sustained at relatively favorable levels. This data suggests that the plant maintains robust growth and high vitality, demonstrating its capacity to regulate and adapt to a 200 mM NaCl concentration. However, at 400 mM salt concentration, plant height, root length, and fresh weight per plant underwent significant reductions, while water content exhibited a slight decline. Specifically, growth parameters decreased by 38%, 43%, 74%, and 2.7%, respectively, compared to CK, although the plants maintained a normal external morphology.
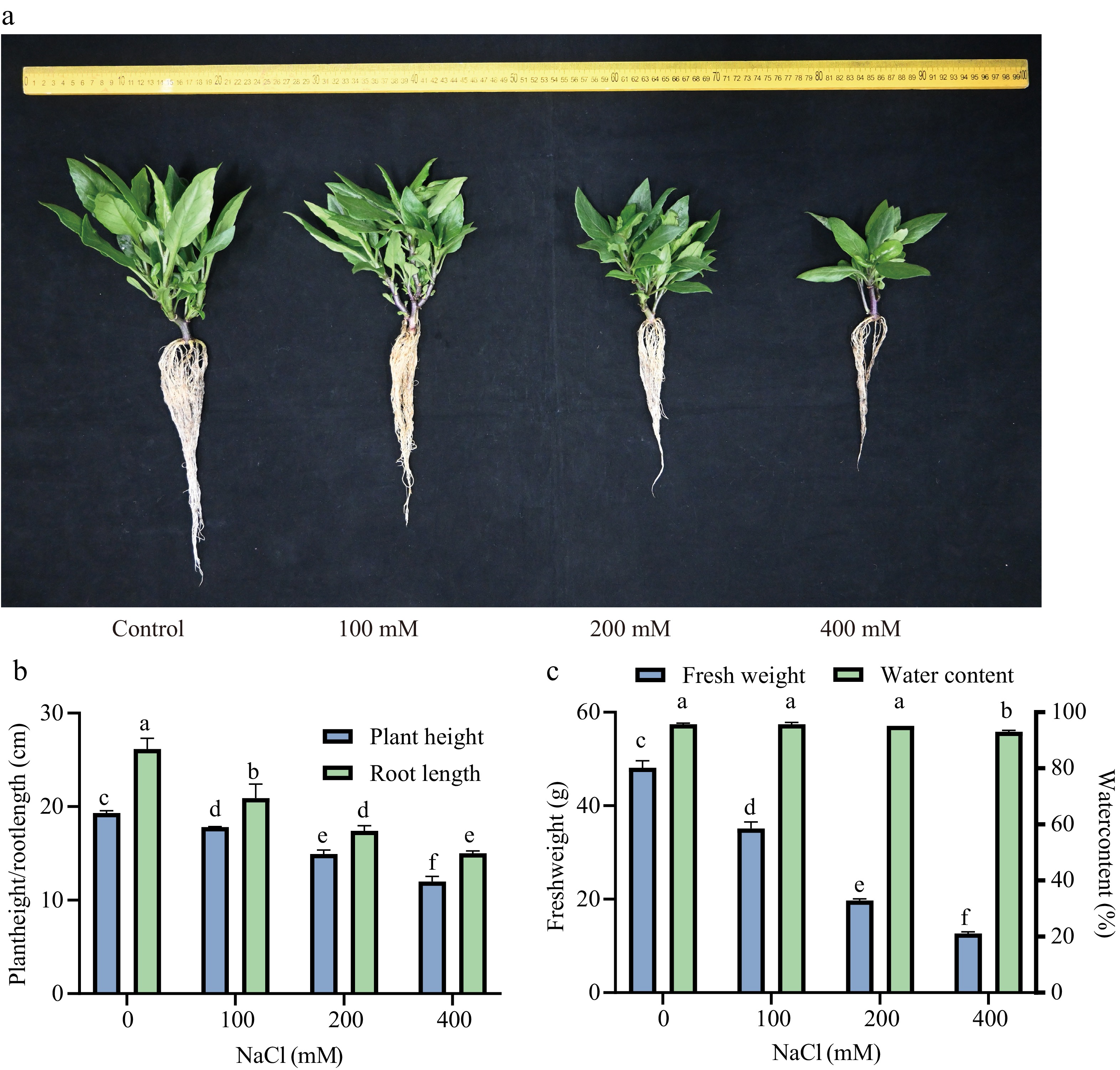
Figure 2.
Effect of salt stress on the growth of Sansha Gynura divaricate. The seedlings cultured for 2 weeks were treated with 0, 100, 200, and 400 mM NaCl. After 23 d, the seedlings were sampled and (a) photographed. (b) Plant height and root length, and (c) fresh weight and water content were measured. The data were expressed as mean ± SE of three replicates. The different letters at the top of the column indicate significant differences at p < 0.05.
Effects of salt stress on K+ and Na+ contents in Sansha Gynura divaricata seedlings
-
Under salt stress conditions, plants undergo ionic stress, primarily reflected in cellular ion imbalances caused by Na+ accumulation and K+ deficiency, which disrupts fundamental metabolic and physiological processes. In extreme cases, this imbalance can result in cell death. The impact of varying salt stress concentrations on K+ and Na+ contents in Sansha Gynura divaricata seedlings was analyzed (Fig. 3). A progressive increase in Na+ content was observed in leaves, with markedly higher levels compared to roots (Fig. 3a). Relative to the CK, the Na+ content increased by factors of 3.00, 6.28, and 8.81, respectively. In roots, Na+ content exhibited an initial increase, followed by a decline, and then a subsequent rise, with respective growth rates of 1.01, 0.82, and 0.86 times that of CK. Consequently, the accumulation rate of Na+ in leaves was substantially greater than in roots. The K+ content exhibited a similar trend in both leaves and roots, characterized by an initial decline, a subsequent increase, and a final reduction (Fig. 3b). In leaves, the percentage decreases in K+ content relative to the CK were 33%, 16.5%, and 36%, whereas in roots, the reductions were recorded at 26%, 25%, and 76%. Under 200 mM NaCl treatment, K+ content increased in both leaves and roots, though the overall variation remained relatively minor. The K+/Na+ ratio consistently remained higher in roots than in leaves, with a slight but statistically insignificant increase in roots under 200 mM NaCl treatment (Fig. 3c). These findings suggest that roots possess a greater capability to absorb K+ from the external environment while restricting Na+ uptake, thereby contributing to an improved internal metabolic environment.
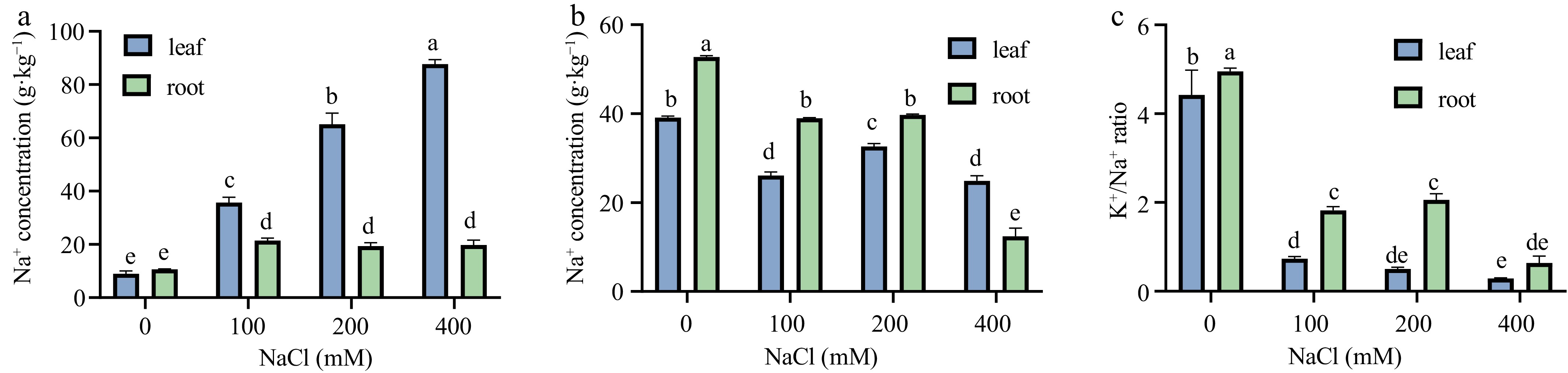
Figure 3.
Effects of salt stress on Na+ and K+ contents of Sansha Gynura divaricata. The leaves and roots of Sansha Gynura divaricata were harvested after salt treatment, and the contents of (a) Na+, and (b) K+ were determined by flame spectrophotometer. (c) The ratio of Na+ to K+. The data is expressed as mean ± SE of three replicates. The different letters at the top of the column indicate significant differences at p < 0.05.
Effects of salt stress on photosynthetic pigments and antioxidant enzyme activities in Sansha Gynura divaricata
-
The impact of salt stress on photosynthetic pigments in Sansha Gynura divaricata seedling leaves is illustrated in Fig. 4a. Following 23 d of exposure to varying salt concentrations, a declining trend in photosynthetic pigments was observed, with reductions differing across various indicators. The levels of chlorophyll a, chlorophyll b, and total chlorophyll exhibited the smallest decline within the 100−200 mM range, whereas carotenoid content showed a slight increase at 100 mM. When salt stress reached 400 mM, all photosynthetic pigment parameters decreased to their lowest levels. Experimental findings demonstrated that salt stress contributed to a reduction in photosynthetic pigment content. However, the decline was less pronounced at 200 mM NaCl, suggesting that this concentration exerted only limited adverse effects on Sansha Gynura divaricata. In contrast, exposure to 400 mM NaCl markedly suppressed the synthesis of photosynthetic pigments.
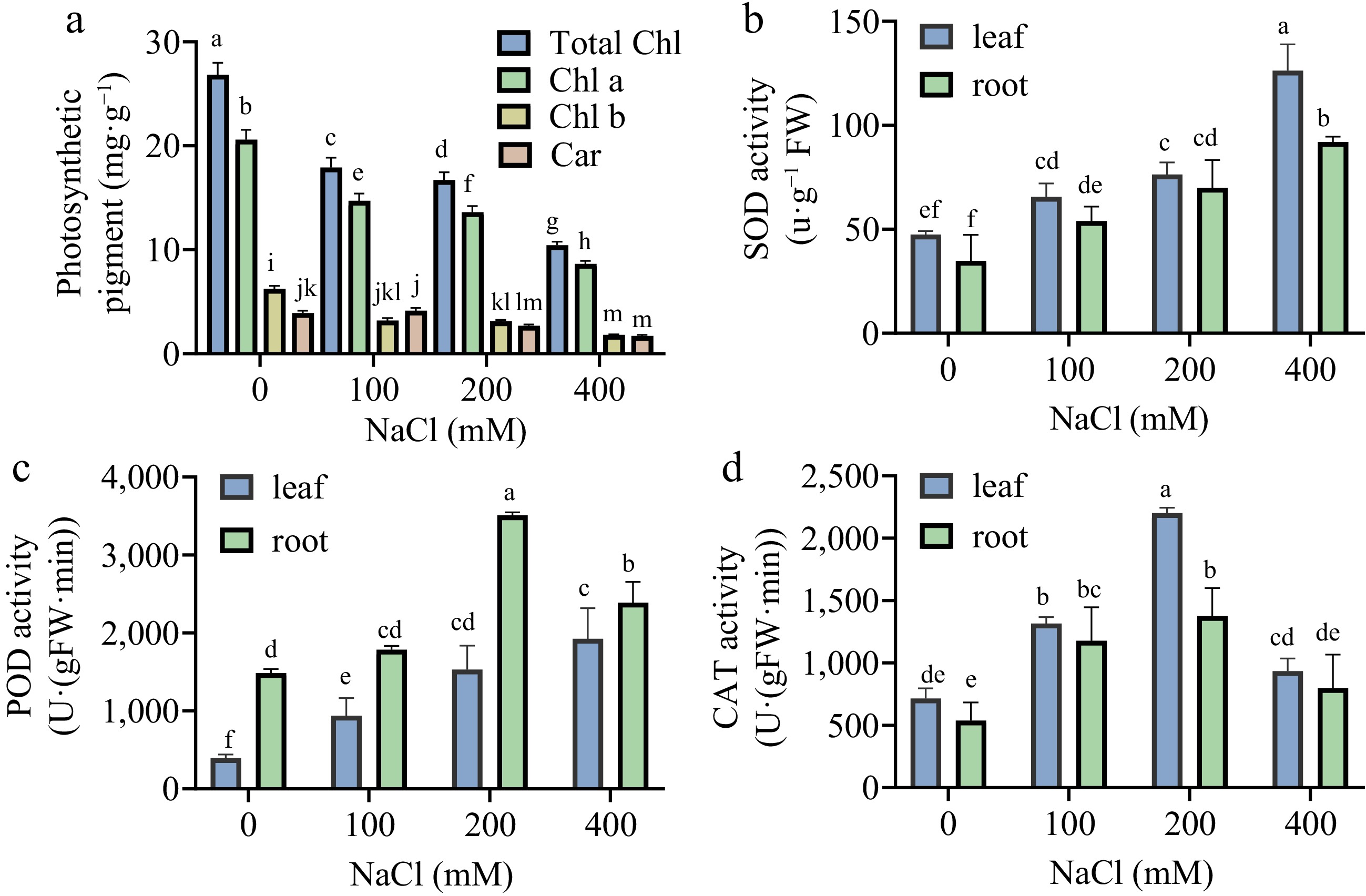
Figure 4.
Effects of salt stress on the activities of photosynthetic pigments and antioxidant enzymes of Sansha Gynura divaricata. The leaves and roots of Sansha Gynura divaricata were harvested after salt treatment, and the activities of (a) photosynthetic pigment, (b) SOD, (c) POD, and (d) CAT were determined by enzyme labeling instrument. The data is expressed as mean ± SE of three replicates. The different letters at the top of the column indicate significant differences at p < 0.05.
The responses of antioxidant enzymes in Sansha Gynura divaricata seedlings under salt stress are depicted in Fig. 4b−d. SOD activity exhibited a significant increase with rising salt concentrations in both leaves and roots (p < 0.05), with higher activity detected in leaves than in roots, while both demonstrated comparable growth rates (Fig. 4b). This suggests that SOD contributes to maintaining redox homeostasis in Sansha Gynura divaricata under salt stress conditions. Additionally, CAT and POD activities were also upregulated in response to salt stress, functioning in a coordinated manner. In leaves, POD activity displayed an upward trend from 0 to 200 mM, reaching its peak, followed by a decline at 400 mM, though levels remained higher than those in CK. In contrast, POD activity in roots exhibited a continuous increase up to 400 mM, where it attained its maximum value (Fig. 4c). CAT activity followed a similar pattern in both leaves and roots, initially increasing and subsequently declining, with peak values recorded at 200 mM, while levels at 400 mM remained elevated compared to CK (Fig. 4d). These findings suggest that SOD, POD, and CAT collectively contribute to mitigating oxidative stress in Sansha Gynura divaricata by scavenging reactive oxygen species. Specifically, SOD facilitates the conversion of superoxide anions into hydrogen peroxide, which is then further decomposed into water through the catalytic actions of POD and CAT.
Effects of salt stress on MDA content and osmotic regulators in Sansha Gynura divaricata
-
MDA, a byproduct of membrane lipid peroxidation, intensifies membrane damage and functions as an indicator of cellular injury severity. With increasing salt concentrations, structural degradation of cells and peroxidation levels are exacerbated[17]. The MDA content in both leaves and roots of Sansha Gynura divaricata exhibited a progressive increase, reaching its peak at 400 mM (Fig. 5a). The extent of this increase was generally more pronounced in leaves than in roots, suggesting that cell membrane damage was more severe in the leaves of Sansha Gynura divaricata. SP serves a pivotal function in modulating cellular osmotic potential. As illustrated in Fig. 5b, under salt concentrations ranging from 0 to 400 mM, SP content in leaves initially increased before subsequently declining, reaching its highest value at 200 mM, whereas in roots, an upward trend was maintained. SP content in leaves was markedly greater than that in roots (p < 0.05), with the maximum value in leaves being 1.34 times that observed in roots.
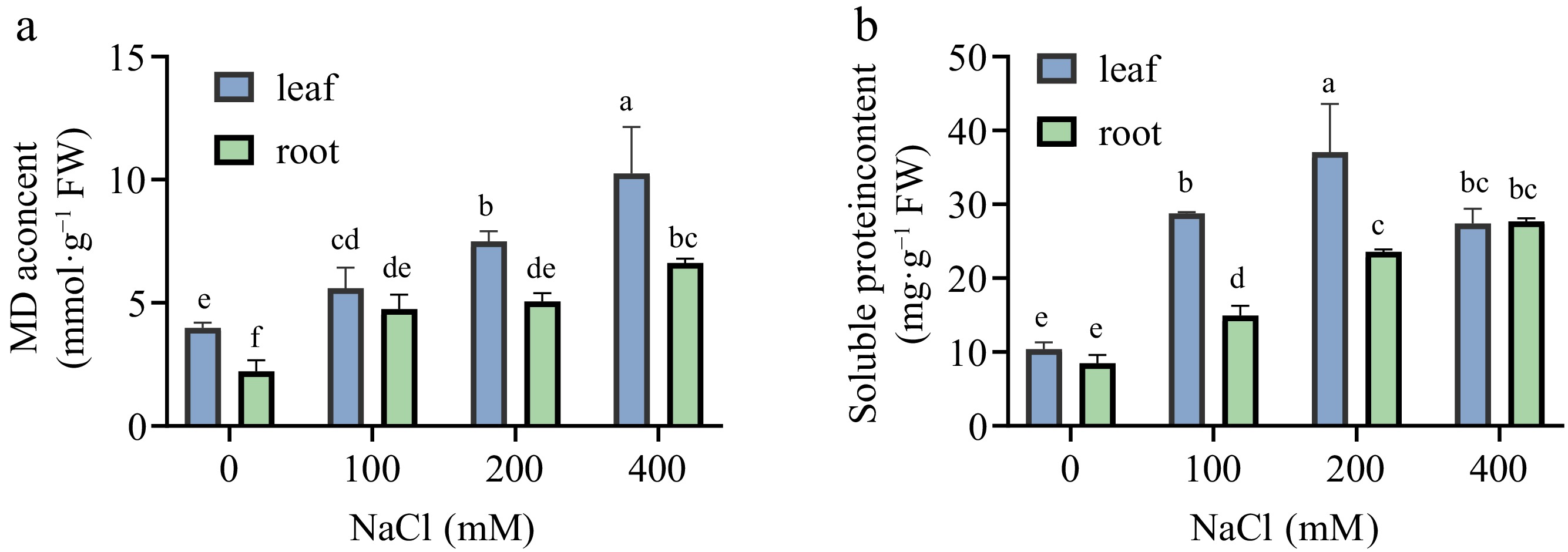
Figure 5.
Effects of salt stress on MDA content and osmotic regulation substances of Sansha Gynura divaricata. The leaves and roots of Sansha Gynura divaricata were harvested after salt treatment. The contents of (a) MDA, and (b) soluble protein were determined by enzyme labeling instrument. The data were expressed as mean ± SE of three replicates. The different letters at the top of the column indicate significant differences at p < 0.05.
Transcriptome responses of Sansha Gynura divaricata leaves and roots to salt stress
-
To further explore the salt tolerance mechanism behind Sansha Gynura divaricata, transcriptome sequencing was performed on the leaf and root tissues of Sansha Gynura divaricata seedlings before and after treatment with 200 mM NaCl. A total of 599,893,414 clean reads were obtained across the four treatment groups, with Q30 ranging from 94.59% to 95.72%, indicating good sequencing quality. Further correlation analysis showed that biological replicates within the same treatment group clustered well, and there were significant differences between treatment groups. This suggests that there are significant differences in gene expression between different tissues of Sansha Gynura divaricata, and salt treatment further triggered transcriptional changes in root and leaf tissues (Fig. 6a). Using |log2(FoldChange)| ≥ 1 and padj ≤ 0.05 as the screening criteria, 3,508 differentially expressed genes were identified in the leaves of Sansha Gynura divaricata after salt treatment, of which 2,608 DEGs were upregulated and 900 DEGs were downregulated (Fig. 6b). KEGG analysis results showed that differentially expressed genes in leaves under salt treatment were significantly enriched in metabolic pathways such as plant hormone signal transduction, cutin, suberine and wax biosynthesis, phenylpropanoid biosynthesis, flavonoid biosynthesis, galactose metabolism, valine, leucine and isoleucine biosynthesis, MAPK signaling pathway-plant, isoquinoline alkaloid biosynthesis, tyrosine metabolism, and diterpenoid biosynthesis (Fig. 6d). Among them, 53 DEGs were related to the plant hormone signal transduction pathway, and these differentially expressed genes mainly participated in ABA and auxin signaling pathways. Notably, most genes in the ABA signal transduction pathway were significantly upregulated after salt treatment, including one PYL (Pyrabactin resistance), nine PP2C (protein phosphatase 2C), two SnRK2 (serine/threonine-protein kinase SRK2), and one ABF (ABA responsive element binding factor) encoding gene; while the differentially expressed genes involved in the upstream of auxin signal transduction were significantly downregulated in the leaves of Sansha Gynura divaricata after salt treatment, including six AUX/IAA (auxin-responsive protein) and two ABF (auxin response factor) encoding genes. In the downstream of this pathway, the expression levels of six SUAR (SAUR family protein) and three GH3 (auxin-responsive GH3 gene family) family members changed under salt treatment conditions (Fig. 6f). In root tissues, the number of downregulated DEGs (1,792) was greater than the number of upregulated DEGs (975) (Fig. 6c). Differentially expressed genes were mainly enriched in nitrogen metabolism, phenylpropanoid biosynthesis, monoterpenoid biosynthesis, MAPK signaling pathway - plant, zeatin biosynthesis, cutin, suberine and wax biosynthesis, dary metabolites, biosynthesis of various plant secondary metabolites, cysteine and methionine metabolism, and brassinosteroid biosynthesis. Plant hormone signal transduction and nitrogen metabolism were the most significantly enriched metabolic pathways for differentially expressed genes in the roots of Sansha Gynura divaricata under salt treatment (Fig. 6e). The DEGs encoding NRT (4 DEGs), NIR (1 DEG), NR (2 DEGs), and GS (1 DEG) were significantly downregulated in the roots of Sansha Gynura divaricata under NaCl treatment, while a gene encoding GDH was significantly upregulated after salt treatment (Fig. 6g). These results suggest that NaCl treatment inhibits the absorption and utilization of nitrogen by Sansha Gynura divaricata, while ABA and auxin signal transduction pathways may play a positive role in the response and adaptation of Sansha Gynura divaricata to salt stress.
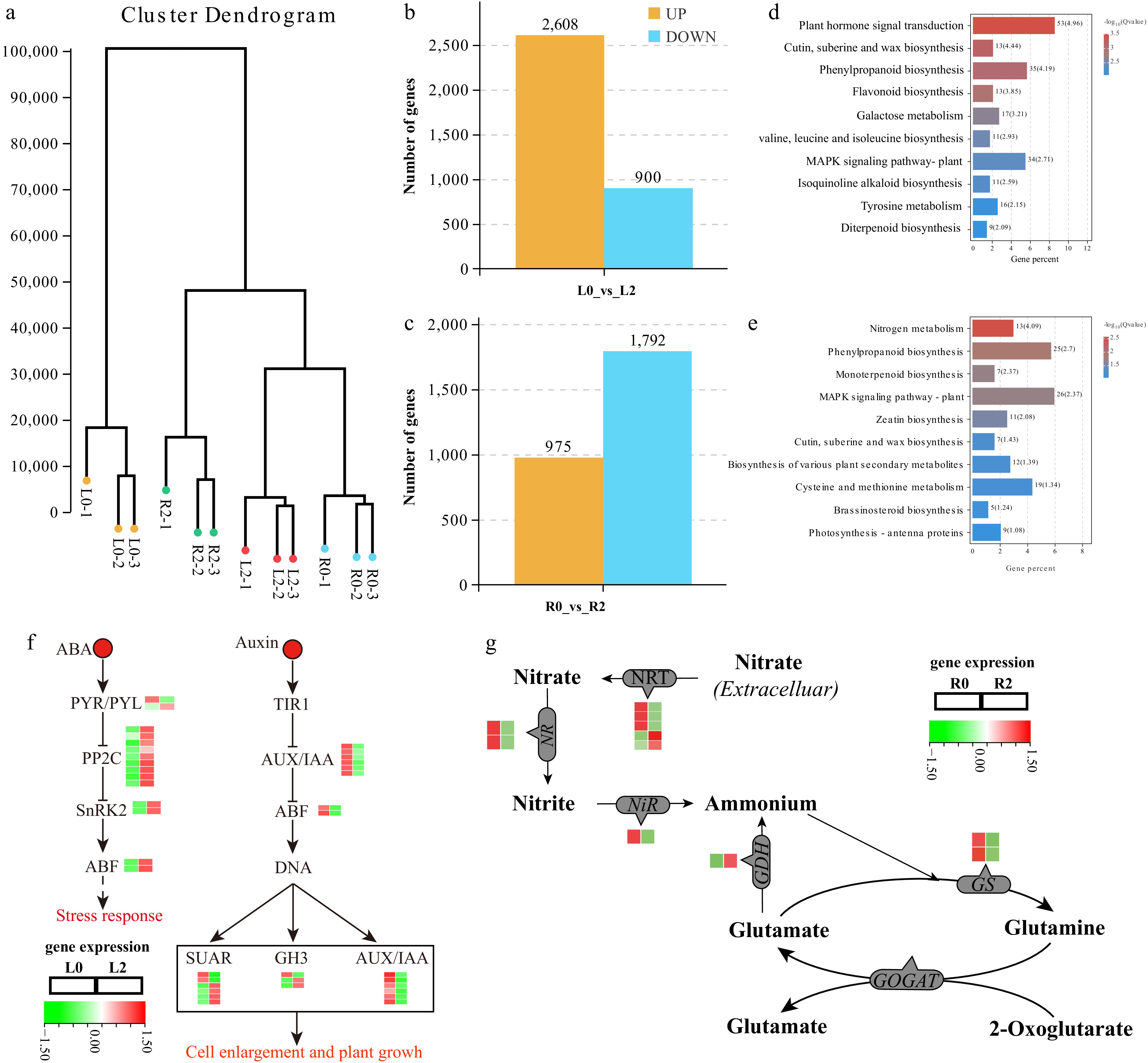
Figure 6.
RNA-Seq data expression profile of Sansha Gynura divaricata leaves and roots under salt stress. (a) Correlation analysis of transcript expression between all samples. (b) Number of DEGs in L2_vs_L0 comparison groups. (c) Number of DEGs in R2_vs_R0 comparison groups. (d) KEGG enrichment analysis of DEGs in leaves. (e) KEGG enrichment analysis of DEGs in roots. (f) Expression patterns of genes involved in plant signal transduction in leaves. (g) Expression patterns of genes involved in nitrogen metabolism in roots.
Metabolomic responses of Sansha Gynura divaricata leaves and roots to salt stress
-
To elucidate the effects of salt stress on the accumulation of metabolites in the leaves and roots of Sansha Gynura divaricata, non-targeted metabolomics analysis was conducted on four groups of samples, detecting a total of 2,298 metabolites in both tissues. Principal component analysis (PCA) results showed that salt treatment caused changes in the accumulation of metabolites in the leaves and roots of Sansha Gynura divaricata, with each treatment's replicates exhibiting good clustering effects.
The first principal component explained 55.6% of the metabolite differences between different tissues, while the second principal component explained 19.5% of the metabolite differences caused by NaCl treatment in both tissues (Fig. 7a). Using VIP > 1, p value < 0.05, and FC ≥ 2 or FC ≤ 0.5 as selection criteria, 219 differentially accumulated metabolites (85 upregulated and 134 downregulated) were identified in the L2_vs_L0 comparison group, and 148 differentially accumulated metabolites (89 upregulated and 59 downregulated) were identified in the R2_vs_R0 comparison group (Fig. 7b). To display the top 20 differentially accumulated metabolites (DAMs) with the greatest up or downregulation in both comparison groups, bar charts were created. In the R2_vs_R0 comparison group, Glu-Gln showed the greatest upregulation (Log2FC = 7.326195), while 4-amino-2-(4-chlorophenyl)-6-(methylthio) pyrimidine-5-carbonitrile showed the greatest downregulation (Log2FC = −5.971868). In addition, the top-ranked up-regulated metabolites mainly included amino acids and secondary metabolites, such as D-(−)-glutamine, arginine, asparagine, and scopolamine, while the top-ranked downregulated metabolites included a large number of organic acid compounds, including 2-oxoglutaric acid, DL-malic acid, and fumaric acid (Fig. 7c). In the L2_vs_L0 comparison group, sodium danshensu had the highest upregulation (Log2FC = 5.601281), while Cer 12:0; 2O/4:0 had the greatest downregulation (Log2FC = −7.213428) (Fig. 7d). Further enrichment analysis of the differentially accumulated metabolites in both comparison groups was performed using the KEGG database. The results indicated that under salt stress, differentially accumulated metabolites in the leaves of Sansha Gynura divaricata were significantly enriched in metabolic pathways such as monoterpenoid biosynthesis, 2-oxocarboxylic acid metabolism, arginine biosynthesis, carbon metabolism, alanine, aspartate and glutamate metabolism, butanoate metabolism, microbial metabolism in diverse environments, biosynthesis of amino acids, glycine, serine and threonine metabolism, and biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid (Fig. 7e), while the differentially accumulated metabolites in the roots were involved in metabolic processes such as alanine, aspartate and glutamate metabolism, carbon metabolism, biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid, arginine biosynthesis, histidine metabolism, butanoate metabolism, biosynthesis of amino acids, biosynthesis of terpenoids and steroids, 2-oxocarboxylic acid metabolism, and citrate cycle (TCA cycle) (Fig. 7f).
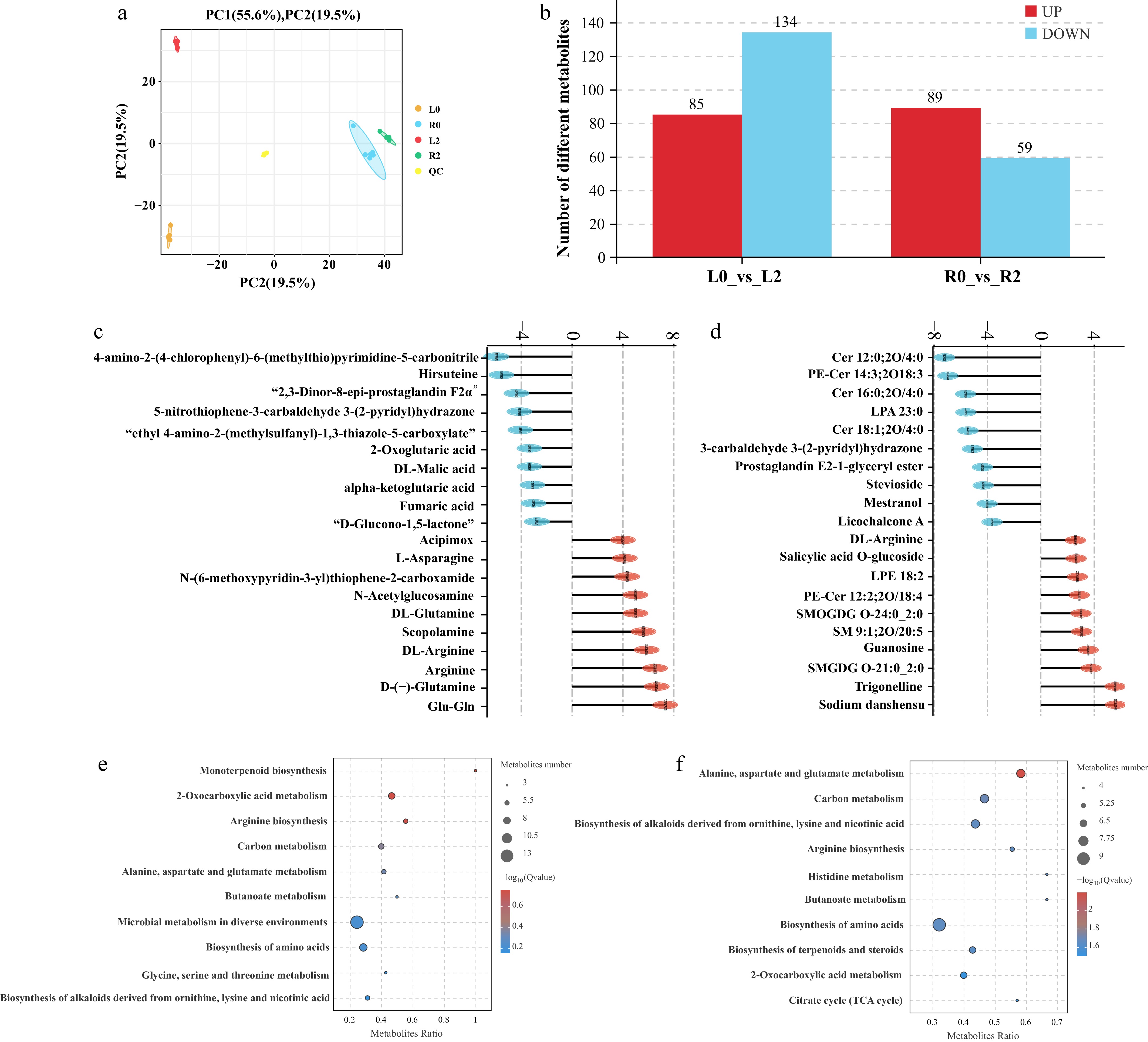
Figure 7.
Metabolomic analysis of Sansha Gynura divaricata leaves and roots under salt stress. (a) Principal component analysis (PCA) plots for all samples. (b) Number of differentially accumulated metabolites in different comparison groups. (c) The top 20 metabolites with the largest upregulated or downregulated fold changes in Sansha Gynura divaricata roots under salt stress. (d) The top 20 metabolites with the largest upregulated or downregulated fold changes in Sansha Gynura divaricata leaves under salt stress. (e) Bubble chart of KEGG pathways for differentially accumulated metabolites in roots. (f) Bubble chart of KEGG pathways for differentially accumulated metabolites in leaves.
-
Gynura divaricata is native to Wuzhishan city, Hainan province. Its leaves and stems possess a succulent texture, offering both palatable flavor and high nutritional value, making it a widely consumed wild vegetable in Hainan with recognized medicinal and breeding significance. The protein content of Gynura divaricata surpasses that of celery[18] and cabbage and is comparable to that of asparagus. It is abundant in vitamins, including folic acid, as well as essential mineral elements such as Ca, P, K, Na, and Mg, with concentrations markedly exceeding those found in Chinese cabbage, rape, and celery[19]. These nutrients provide critical dietary support for island residents and stationed troops, aiding in the prevention of osteoporosis, rickets, and potassium deficiency, while contributing to immune system enhancement. Moreover, Sansha Gynura divaricata is a rich source of flavonoid compounds, which exhibit antioxidant, anti-inflammatory, antiviral, and antidepressant properties[20], in addition to their potential to lower blood lipid and cholesterol levels[21]. Among these compounds, gallocatechin is the most abundant, serving a pivotal function in lipid metabolism regulation[22], thereby classifying the plant as a functional vegetable with both nutritional and medicinal benefits. Although Sansha Gynura divaricata features an extended growth cycle, extensive branching, and strong stress resistance, it typically enters the flowering stage between March and April. Following flowering, the plant undergoes aging, resulting in a decline in the quality of edible portions, rendering them unsuitable for consumption. However, a morphologically similar vegetable has been identified in Sansha city, growing naturally on South China Sea reefs, exhibiting a non-flowering phenotype (Fig. 1). This distinction may be attributed to geographical variations, as environmental factors exert a profound influence on plant adaptability. The distinctive island climate and ecological conditions of the South China Sea may have driven substantial genetic divergence in Sansha Gynura divaricata, leading to the emergence of unique genetic traits.
In this experiment, as salt concentration increased, a decline in both biomass and chlorophyll content of Sansha Gynura divaricata was observed. However, at 100−200 mM salt concentrations, photosynthetic pigment levels remained relatively stable, with carotenoid content exhibiting an unexpected increase rather than a decrease at 100 mM (Fig. 4a). This stability may be attributed to the synergistic involvement of antioxidant enzymes and chlorophyll in photosynthesis and photoprotection, which contributed to maintaining growth, development, and internal metabolic homeostasis in Sansha Gynura divaricata[23]. The antioxidant enzyme system comprises SOD, CAT, and ascorbate peroxidase (APX)[24]. As the primary defense mechanism within the antioxidant system, SOD facilitates the rapid reaction of O2− with H+, generating H2O2 and O2. Subsequently, POD and CAT function cooperatively to catalyze reactions involving H2O2 and organic peroxides, such as lipid peroxides, thereby mitigating H2O2 accumulation[25]. Additionally, these enzymes participate in the conversion of tissue carbohydrates into lignin, regulating plant growth and development while ensuring the structural integrity and stability of the cell wall[26,27]. Under salt stress conditions, increased SOD, POD, and CAT activities were detected in both roots and leaves of Sansha Gynura divaricata (Fig. 4b−d). Although POD and CAT activities exhibited a slight reduction at 400 mM, their levels remained higher than those in the CK. SOD activity was the most elevated under salt stress, whereas the decline in POD activity at 400 mM may have resulted from disruptions in biosynthetic pathways or reduced enzyme stability. At 200 mM salt concentration, CAT activity increased, likely due to the role of H2O2 as a signaling molecule that enhances CAT function, thereby maintaining low H2O2 concentrations and improving salt tolerance. However, the reduction in CAT activity at 400 mM may be attributed to its limited protective capacity, low affinity for H2O2, and susceptibility to inactivation or degradation[28,29]. A similar pattern of increased SOD, POD, and CAT activities under 200 mM salt stress has been reported in Brassica napus seedlings[30]. Furthermore, the observed decline in enzymatic activity at elevated salt concentrations aligns with findings from Cavalcanti's study on cowpea leaves[31], suggesting that although plants enhance salt stress adaptation through antioxidant enzyme systems, these protective mechanisms have inherent limitations. When salt concentrations exceed the plant's tolerance threshold, these defense responses become ineffective. Additionally, MDA content, an essential indicator of membrane lipid peroxidation damage, was examined[32]. A continuous increase in MDA content was recorded in both leaves and roots as salt concentrations rose (Fig. 5a), reflecting progressively severe membrane damage. Regarding other osmotic regulators, SP content in leaves initially increased but subsequently declined at 400 mM (Fig. 5b), possibly due to stress conditions surpassing the plant's osmotic regulation capacity, thereby impairing photosynthesis, respiration, and metabolic functions. Conversely, the gradual accumulation of SP in roots suggests that mechanisms were activated to alleviate salt stress-induced osmotic pressure, facilitate cellular ion compartmentalization, and sustain cell turgor pressure, ultimately protecting metabolic pathways[33].
When exposed to saline environments, Sansha Gynura divaricata undergoes a substantial increase in Na+ accumulation due to soil salinity, which disrupts ionic equilibrium within the plant and impairs the uptake of water and essential nutrients[34]. Under salt stress conditions ranging from 0 to 200 mM, no significant inhibition was detected, with only minimal reductions observed in plant height, root length, and fresh weight, while water content remained relatively stable. However, at 400 mM salt concentration, a marked inhibitory effect was noted compared to the CK, characterized by a reduction in leaf number, significant shortening of primary roots, decreased lateral root formation, and lower water content. Despite these effects, the plant sustained favorable growth conditions (Fig. 2a, b) and retained a normal external morphology, indicating strong salt tolerance. Although roots are generally the primary organs exposed to salt stress in soil, with Na+ content typically higher in roots than in leaves in most crops, this study demonstrated that Na+ levels in the leaves of Sansha Gynura divaricata consistently surpassed those in roots. This suggests active Na+ transport from roots to aerial parts, leading to greater stress accumulation in above-ground tissues. Despite the elevated Na+ content within plant tissues, K+ levels remained relatively stable. The most significant antagonistic interaction was observed between Na+ and K+, highlighting K+ homeostasis as a key factor in mitigating Na+ toxicity. Treatment with 200 mM NaCl resulted in reduced Na+ content and increased K+ accumulation in both leaves and roots, whereas at 400 mM, Na+ levels increased while K+ concentrations declined (Fig. 3a, b). Additionally, the K+/Na+ ratio increased at 200 mM but exhibited a slight decrease at 400 mM (Fig. 3c). These findings suggest that under 200 mM NaCl stress, Sansha Gynura divaricata sustains ionic homeostasis by enhancing K+ uptake to counteract salt stress. Although metabolic activities were somewhat affected at 400 mM NaCl, functional integrity was largely preserved. Previous studies have reported that succulent-stemmed and leaf-bearing cells of Suaeda salsa can actively transport Na+ against concentration gradients into extracellular spaces and vacuoles through Na+/H+ antiporter proteins[35], thereby sequestering Na+ within succulent tissues. This mechanism enhances osmotic regulation and water retention, allowing plants to mitigate salt stress through ionic balance regulation and improved water absorption capacity. Similarly, Sansha Gynura divaricata exhibits halophytic characteristics, with its succulent stems and leaves potentially facilitating the sequestration of toxic Na+ within vacuoles of green tissue cells. This stored Na+ may function as an osmotic regulator, contributing to water balance maintenance and reducing cytoplasmic Na+ toxicity[36]. Moreover, the presence of succulent stems and leaves aids in water storage, effectively diluting salt ion concentration per unit volume, thereby preventing toxic ion accumulation[37] and safeguarding the plant from salt-induced damage. In summary, the succulence of Sansha Gynura divaricata stems and leaves appear to represent a key adaptive strategy for enhancing salt tolerance.
Transcriptomics and metabolomics can analyze the effects of environmental stress on plant gene expression and metabolites at high throughput. In this study, Sansha Gynura divaricata exhibited extensive transcriptional changes after salt treatment. Overall, the impact of salt stress on gene expression in Sansha Gynura divaricata leaves was greater than in the roots (Fig. 6c, d). However, more genes were downregulated in the roots of Sansha Gynura divaricata after salt treatment, suggesting that different tissues of Sansha Gynura divaricata have distinct responses and adaptation mechanisms to salt stress. Compared to the absence of salt solution, the differentially expressed genes in Sansha Gynura divaricata leaves under 200 mM NaCl treatment were significantly enriched in the plant hormone signal transduction pathway (Fig. 6f). Genes involved in the ABA and auxin pathways within this pathway were most affected by salt stress. Abscisic acid (ABA) plays a crucial role in various plant growth and developmental processes, including seed germination, dormancy, root elongation, leaf senescence, and fruit ripening[38−40]. Moreover, ABA plays a vital role in plant responses to biotic and abiotic stresses, enabling plants to adapt and survive in harsh ecological environments. The ABA receptors Pyrabactin resistance (PYR)/PYR1-like (PYLs), protein phosphatase 2C (PP2Cs), serine/threonine-protein kinase SRK2 (SnRK2s), and the ABA-responsive element binding factor (ABF) are key components of the ABA signaling pathway[41]. In this study, differentially expressed genes encoding PP2Cs, SnRK2s, and ABF were significantly upregulated after NaCl treatment, suggesting that Sansha Gynura divaricata may enhance the plant's adaptability to NaCl stress by activating the ABA signaling pathway. Auxin is another important plant hormone that plays a significant role in plant growth and development, and stress responses. Studies have shown that auxin signaling can regulate plant salt tolerance by modulating the accumulation of flavonols or the ability to scavenge reactive oxygen species[42]. In this study, six genes encoding AUX/IAA were significantly downregulated after NaCl treatment, indicating that these genes play an important role in the auxin-mediated stress response of Sansha Gynura divaricata. Whether they participate in flavonol accumulation or ROS scavenging remains to be further studied. Unlike the leaves, the differentially expressed genes identified in the roots of Sansha Gynura divaricata under NaCl treatment were significantly enriched in the nitrogen metabolism pathway. Interestingly, these differentially expressed genes are related to the absorption and assimilation of nitrate, and most were downregulated in the roots of Sansha Gynura divaricata under salt treatment. In addition, the analysis of the metabolome also indicated that salt stress caused changes in amino acid metabolism and carbon metabolism pathways in Sansha Gynura divaricata plants (Fig. 7). These findings indicate that sodium chloride treatment affects the absorption and utilization of nitrate by Sansha Gynura divaricata. However, whether this effect directly leads to inhibited growth under salt stress requires further investigation. This is crucial for rational fertilization in the practical production of Sansha Gynura divaricata in saline-alkali soils.
-
Sansha Gynura divaricata, recognized as a widely consumed vegetable, possesses dual attributes encompassing both nutritional and medicinal properties. While its protein and vitamin levels are comparable to those of conventionally cultivated vegetables, it exhibits a higher concentration of essential mineral elements and is abundant in flavonoids. Its consumption has been associated with multiple health benefits, including antioxidant activity, free radical scavenging, antimicrobial and anti-inflammatory effects, and immune system enhancement. Notable geographical and climatic variations exist between the South China Sea reef, where Sansha Gynura divaricata thrives, and Wuzhi Mountain. Environmental factors such as fluctuations in light exposure, temperature, air humidity, and soil salinity may influence its genetic traits, resulting in physiological modifications and the inability to undergo flowering and fruiting. The plant's growth cycle is not restricted by seasonal changes, exhibiting remarkable stress resistance and adaptability. Furthermore, its ability to be cultivated year-round with multiple harvests on South China Sea reefs contributes to reduced production costs and enhanced yield. Salt tolerance analysis of Sansha Gynura divaricata indicates that normal growth is sustained within a NaCl concentration range of 0−200 mM. Although exposure to 400 mM NaCl exerts significant effects, the plant retains its normal morphological characteristics, further demonstrating its strong salt tolerance. These findings suggest that Sansha Gynura divaricata is nutritionally rich, highly resistant to salt stress, and easily cultivable. Consequently, it is well suited for growth in saline environments such as coastal tidal flats and salt-affected lands, offering a promising vegetable option to support large-scale cultivation efforts on South China Sea reefs.
-
The authors confirm their contributions to the paper as follows: conceptualization: Jiang X , Xie Q; formal analysis: Tu Y; funding acquisition: Jiang X, Xie Q; investigation: Tu Y , Zhang H; methodology: Wang Y, Chu J; supervision: Jiang X, Xie Q; writing − original draft preparation: Tu Y, Jiang X, Xie Q; writing − review and editing: Xie Q. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author upon reasonable request.
-
We thank Bullet Edits for English language editing. We also thank the reviewers and editors for helpful comments on earlier drafts of the manuscript. This study is supported by the Natural Science Foundation of Guangdong province (2023A1515012295), the Program for Guangdong Provincial Innovative Team for Development and Utilization of Germplasm Resource of Saline-Alkali Tolerant Plants, National Key R&D Program of China (2018YFE0207203-2), the National Natural Science Foundation of China (31660253), the Foundation of Zhanjiang city, Scientific Research Start-up Funds of Guangdong Ocean University.
-
The authors declare that they have no conflict of interest.
-
Received 31 January 2026; Accepted 5 May 2026; Published online 30 June 2026
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tu Y, Zhang H, Wang Y, Chu J, Xie Q, et al. 2026. Analysis of nutritional value and salt tolerance of Sansha Gynura divaricata. Tropical Plants 5: e023 doi: 10.48130/tp-0026-0018 

Analysis of nutritional value and salt tolerance of Sansha Gynura divaricata
- Received: 31 January 2026
- Revised: 09 April 2026
- Accepted: 05 May 2026
- Published online: 30 June 2026
Abstract: Sansha Gynura divaricata (L.) DC., recognized as a salt-tolerant vegetable endemic to the islands and reefs of the South China Sea, has been the subject of limited investigation regarding its salt tolerance and nutritional attributes. In this study, potted Gynura divaricata plants were subjected to four NaCl concentration gradients (0, 100, 200, and 400 mM). After 23 d of exposure to salt stress, key growth and physiological parameters, along with K+ and Na+ concentrations, were quantified and evaluated. Additionally, nutritional properties were assessed in the edible tender stems and leaves. Finally, this research undertook a comprehensive analysis of gene expression and metabolite responses of Gynura divaricata seedlings under a 200 mM NaCl treatment. The findings demonstrated that Gynura divaricata possesses significant salt tolerance. Under 200 mM NaCl treatment, Gynura divaricata exhibits remarkable salinity resilience—maintaining Na+/K+ homeostasis and robust antioxidant capacity, while simultaneously modulating hormone signal transduction and regulating the expression of genes associated with nitrogen metabolism. Furthermore, alterations in key metabolites within the nitrogen metabolism pathway contribute to their stress tolerance. This multifaceted response enables Gynura divaricata plants to resist and adapt to salt-induced damage, providing valuable insights into the mechanisms underlying salt tolerance in plants. Nutritional analyses highlighted that Gynura divaricata is enriched with essential mineral elements and flavonoid compounds. Consequently, Gynura divaricata is recognized as an exceptional vegetable with substantial nutritional value, integrating both medicinal and dietary applications, and is well-suited for cultivation in the high-salinity environments of the South China Sea islands and reefs.
-
Key words:
- Sansha Gynura divaricata /
- Nutritional value /
- Salt tolerance /
- RNAseq