






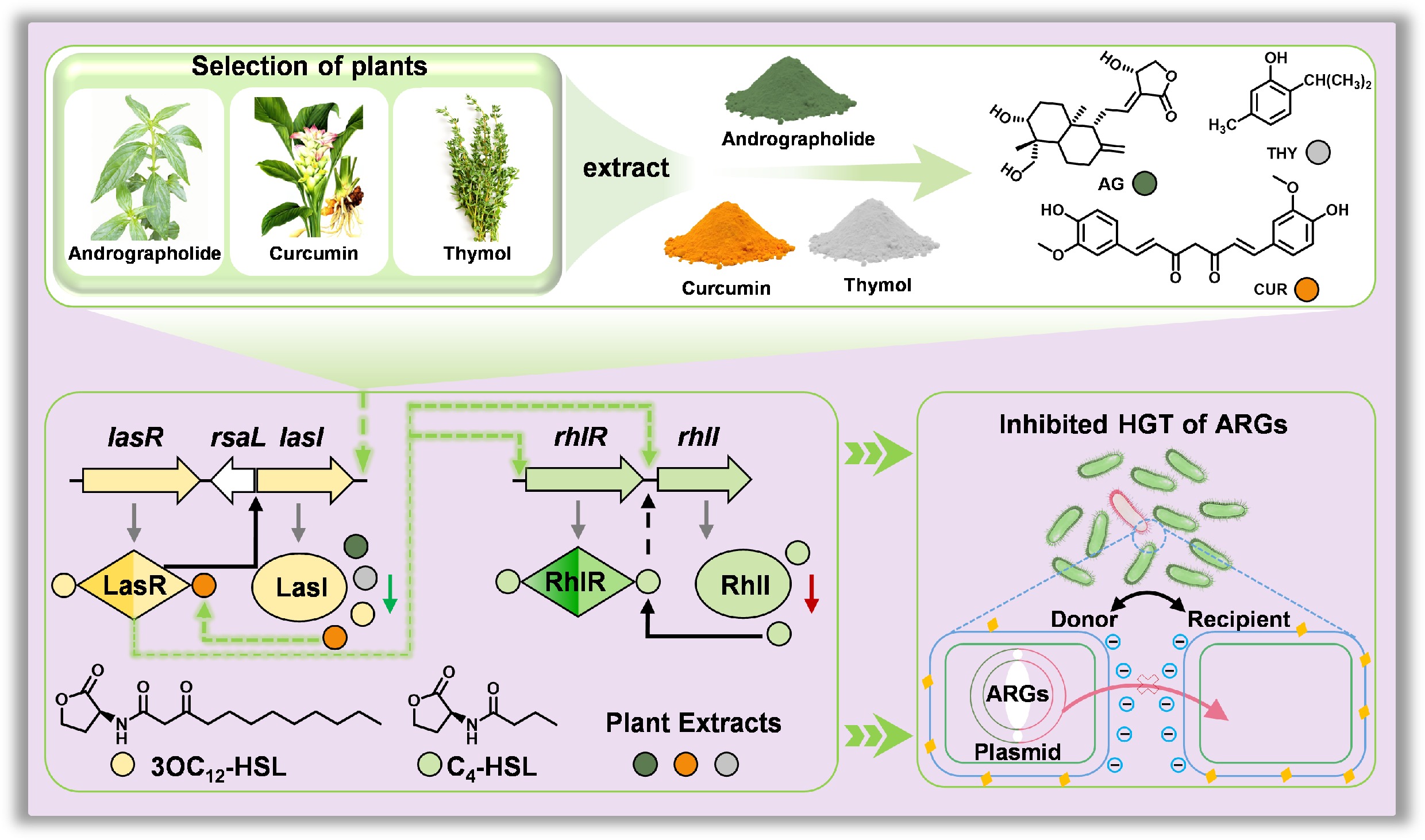



-
The application of livestock manure is a highly significant strategy for fertilizing the land and promoting crop growth[1]. However, a large variety of human bacterial pathogens (HBPs) have been inevitably introduced into the soil when applying livestock manure, raising a high concern about their risks to the ecosystem and humans[2]. Especially in recent years, the extensive use of antibiotics in livestock industries has extensively promoted the occurrence and spread of antibiotic resistance genes (ARGs) and virulence factor genes (VFGs)[3,4]. Moreover, the presence of mobile genetic elements (MGEs) such as plasmids, transposons, and integrons further accelerates the horizontal gene transfer (HGT) of environmental ARGs and VFGs among HBPs, leading to antibiotic-resistant HBPs and even multidrug-resistant superbugs[5]. Previous studies have demonstrated that soil HBPs can migrate into plant endophytes and the phyllosphere, thus entering the food chain and causing a potential threat to human health[6,7]. Hence, it is necessary to develop effective strategies to mitigate the risks posed by HBPs and the associated ARGs, VFGs, and MGEs in agricultural soils, with a focus on food safety and human health.
Currently, several strategies have been developed to address this issue. For example, Cheng et al. found that biochar significantly reduced the abundance of HBPs and ARGs in soils[8]. Some studies also demonstrated that nanoparticles (including nano zero-valent iron, Fe2O3 nanoparticles) attenuated the abundance of ARGs via the damage of genomic and plasmid DNA or the inhibition of HGT[9,10]. The above studies may give important clues to help us alleviate the HBPs and the associated ARGs, VFGs, and MGEs in agricultural soils; however, these nanoparticles are often expensive to manufacture and use, hindering their practical application at a large scale[11]. Additionally, some environmental risks of using nanomaterials should not be neglected[12]. Therefore, it is still a challenge to develop efficient technological, green, economic, and behavioral interventions to mitigate the risks of HBPs from agricultural soils.
Plant extracts derived from active ingredients in roots, stems, leaves, flowers, and other parts have been reported as a promising soil health management practice for soil remediation[13]. However, these recent studies mainly investigate their roles in reducing heavy metals and organic pollutants, improving soil physicochemical properties, and promoting plant growth[14,15]. Few studies have investigated the remediation effects of plant extracts on soil biological pollution. There is no denying that some plant extracts have antibacterial properties[16]. For example, prenylated flavonoids from plants exhibit robust antibacterial activities against important pathogens, including gram-positive pathogens and gram-negative pathogens[17,18]. These studies mainly use plant extracts as antibacterial agents in the clinical treatment of diseases associated with microbial infections. Recent studies also demonstrated the potential of natural plant extracts to protect sugar beet and wheat against soil-borne plant viruses[19,20]. Most existing studies focus on the bactericidal effects of extracts on pathogenic bacteria; however, little is known about whether they inhibit the HGT of ARGs and VFGs. Thus, the regulatory potential and mechanisms of plant extracts on soil HBPs, and the related risk factors, remain unclear.
Quorum sensing (QS), a cell-cell signaling process in bacteria, is critical for coordinated group behavior[21]. Previous studies have demonstrated that many root-associated bacteria require QS for colonization of the rhizosphere, thereby forming biofilms[22], which can harbor pathogenic microbes and accelerate the propagation of antimicrobial resistance, as HGT tends to occur within dense bacterial populations[23]. Moreover, QS also contributes to the production of a variety of virulence factors, including protease, hemolysin, pyocyanin, and hydrogen cyanide, which are closely related to the risks of HBPs[24]. Thus, disturbing QS may be responsible for the attenuation of the abundance and risks of HBPs. In fact, many studies have reported the capacity of plant extracts to interfere with intra- and inter-species QS communication systems. Several studies have shown that conjugative transfer of ARGs can be effectively inhibited by blocking QS in drinking water systems[25,26], however, few studies have investigated the attenuation of antibiotic resistance in agricultural soils amended with manure by disturbing QS. Moreover, whether plant extracts can interfere with QS and reduce the abundance of soil microbial contaminants remains unclear.
Herein, the objective of this study is to explore the effect of plant extracts on the attenuation of microbial contaminants in agricultural soils amended with manure and to reveal the underlying mechanism. Curcumin (CUR), andrographolide (AG), and thymol (THY) were selected as representative components of plant extracts because previous studies have shown they may inhibit QS and thereby eradicate bacterial biofilms[27−29]. The results demonstrated that plant extracts significantly reduced the abundance of HBPs in manure-amended agricultural soils, thereby reducing their risks, as evidenced by reductions in ARGs, VFGs, and MGEs. Specifically, the plant extracts disturbed the QS by reducing the concentration of signal molecules and down-regulating QS-related genes, thereby inhibiting biofilm formation and reducing the virulence factors secreted by HBPs. Moreover, in vitro assays further verified that plant extracts inhibited the spread of ARGs and VFGs by disturbing the QS. At last, molecular docking was applied to more deeply uncover the mechanism by which different plant extracts disturb bacterial QS. This study will not only help understand the attenuation of microbial contaminants in manure-amended agricultural soils mediated by plant extracts, but also provide strong support for the development of promising technologies to control soil-borne microbial contaminants.
-
The soil was sourced from Zhejiang University (120°09'10" E, 30°30'57" N) in Hangzhou, China, subsequently air-dried, crushed, and sieved through a 2.0 mm sieve. The physicochemical characteristics of the soil are presented in Supplementary Table S1. Curcumin (CUR) and andrographolide (AG) were purchased from Aladdin Reagent (Shanghai) Co., Ltd, while thymol (THY) was obtained from Merck Company (Darmstadt, Germany). Swine manure was obtained from Huiren Organic Fertilizer (Hangzhou, China).
Experimental design
-
Manured soil microcosms were prepared in a plastic pot (35 cm × 24 cm × 11 cm), into which 3 kg of air-dried soil with a moisture content of 18%–20% was added. The plastic pots were covered with plastic film and incubated at 22 ± 2 °C. Following a seven-day acclimation period, swine manure was added to the soil at a concentration of 100 g/kg, referring to previous studies[4,30]. The swine manure was thoroughly mixed with the soil for one week, and the prepared aqueous plant extracts, including CUR (0.1 mg/kg), AG (0.75 mg/kg), and THY (1.25 mg/kg), were applied once a week. Notably, the doses of CUR, AG, and THY used in the experiment were determined based on preliminary data (Supplementary Fig. S1), which showed that they had no significant effect on bacterial growth at these exposure levels. Soil microcosms without plant extracts were used as control groups (CK). Maintaining soil moisture was achieved by spraying sterile water onto it every 3 d. At sampling times (0, 1, 4, 6, 8, 10, and 12 weeks), soil samples were collected and stored at −80 °C for metagenomic sequencing and the detection of soil signal molecules. The detailed bioinformatics analysis processes, including DNA extraction, metagenomic sequencing, data processing, and analysis, were displayed in the Supplementary Text S1 & Supplementary Table S2.
Analysis on the composition and diversity of HBPs
-
To obtain bacterial functional annotations and taxonomic classification, sequences in the redundant gene set were aligned against the NCBI Non-Redundant (NR) database with an E-value threshold of 1 × 10−5. The composition of potential and high-risk HBPs was identified by cross-referencing lists of potential/dangerous HBPs reported in previous studies[4,8]. Species-level annotations were retrieved from the taxonomic database associated with the NR repository, and species abundance was quantified as the sum of the abundances of genes corresponding to each species. Subsequently, the abundance of each bacterial species and HBP-related species was determined across all samples at each taxonomic rank (Domain, Kingdom, Phylum, Class, Order, Family, Genus, Species), enabling the construction of taxon-specific abundance tables. Notably, because the absolute abundance of microorganisms in soil is easily affected by soil physical and chemical properties, sampling time, and spatial heterogeneity, and fluctuates widely, the relative abundance can more accurately reflect the proportional change in the target pathogen within the entire microbial community. Therefore, the relative abundance was used to calculate the removal efficiency of pathogens rather than absolute abundance.
Analysis of ARGs, VFGs, and MGEs abundances
-
The distribution patterns of ARGs and MGEs derived from metagenomic sequencing datasets were analyzed using the Structured Antibiotic Resistance Genes (SARGs v2.0) module integrated into ARG-OAPs software[8], with an identity threshold of 80% and an E-value cutoff of 10−7. For each sample, ARG abundance in the metagenome was quantified as 'copies per cell'. This quantification was achieved by normalizing the read abundance of individual genes and calibrating the gene copy number against that of essential single-copy genes in the metagenome, a normalization approach that facilitates more precise assessment of the relative abundance of antibiotic resistance genes. The quantification and analysis of all MGEs were conducted following the same protocol as for ARGs, utilizing an MGE-specific reference database[8]. For the screening of VFGs, high-quality reads were aligned against the Virulence Factor Database (VFDB;
www.mgc.ac.cn/VFs/main.htm ) via BLASTX searches, with an E-value threshold of 10−7 and an identity threshold of 80%[4,8]. The gene abundance of VFGs was calculated using the identical methodology applied to ARG abundance analysis.Detection of QS signals
-
The concentration of QS signals in soil was measured by high-performance liquid chromatography-tandem mass spectrometry (LC-MS; UPLC/Xevo TQD, Waters, USA), and the details are presented in Supplementary Text S2, Supplementary Table S3 & S4.
Effects of plant extracts on the virulence factors
-
After incubation, 1 mL of the bacterial solution was removed and centrifuged at 12,000 rpm for 10 min at 4 °C to obtain the supernatant. Then, the concentrations of elastase, pyocyanin, and rhamnolipid in the supernatant were determined. The specific methods were described in Supplementary Text S3.
Effects of plant extracts on the formation of biofilm
-
P. aeruginosa and A. baumannii were selected to perform the experiments in which the formation of biofilm was determined using the crystal violet staining assay. The details were placed in Supplementary Text S4.
Effects of plant extracts on the horizontal gene transfer
-
Given that the RP4 plasmid, which confers resistance to ampicillin, kanamycin, and tetracycline, has been widely used to investigate the horizontal transfer of ARGs in soil, it was selected to explore the effects of plant extracts on horizontal gene transfer. Briefly, E. coli HB101 carrying plasmid RP4 resistant to tetracycline, kanamycin, and ampicillin was used as the donor strain, and Pseudomonas putida KT2442 carrying the same plasmid was used as the recipient strain. These strains were applied to perform the intraspecies conjugative transfer of ARGs. For inter-species conjugative transfer of ARGs, the recipient strain was replaced with an E. coli NK5449-resistant strain. The details about the effects of plant extracts on the conjugative transfer of ARGs were presented in Supplementary Text S5.
Molecular docking
-
The LasR (ID: 3IX3) protein was searched from the PBD protein database (
www.rcsb.org ). The PubChem database (www.pubchem.ncbi.nlm.nih.gov ) was used to search for and download the molecular structures of the root extracts, including CUR (ID: 969516), AG (ID: 5318517), and THY (ID: 6989). Molecular docking software (AutoDock Vina) was used to predict which amino acid residues on LasR were bound to 3OC12-HSL, CUR, AG, and THY. To verify the theoretical results, validation experiments were performed using the Biacore X100 molecular interaction instrument to determine the combined affinity of LasR with 3OC12-HSL, CUR, AG, and THY.Statistical analysis
-
Data are presented as mean ± standard deviation (SD). Pearson's correlation coefficients were calculated using SPSS statistical package version 21 (SPSS Inc., Chicago, USA), and a one-way ANOVA was conducted to assess significance. The statistical difference was determined at p < 0.05, unless otherwise indicated.
-
Previous studies have shown that organic fertilizer made from pig manure contains a wide variety of pathogens[31−33]. In this study, a total of 323 HBPs were screened based on the HBPs database. After application of plant extracts, the total abundance of HBPs decreased significantly (Fig. 1a). For instance, the total abundance of HBPs was reduced by 25.71%, 27.59%, and 27.88% after the treatment of CUR, AG, and THY, respectively. Similarly, the number of HBPs was also reduced (Fig. 1b). Notably, a greater reduction in the number of HBPs was observed with the addition of CUR, followed by AG and THY (Fig. 1b). Next, the composition of HBPs was further analyzed, and the results showed that HBPs were mainly composed of Firmicutes, Actinobacteria, Proteobacteria, and Bacteroidetes, similar to a previous study[5]. Compared with Firmicutes and Bacteroidetes, the abundance of Actinobacteria and Proteobacteria in HBPs was significantly decreased with the addition of plant extracts (Fig. 1c; Supplementary Fig. S2), implying that plant extracts may selectively act on Actinobacteria and Proteobacteria but had little effect on Firmicutes and Bacteroidetes, consistent with a previous study where they found plant extracts from the peanut selectively inhibited or facilitated certain communal bacterial, with decreases in the relative abundance of the bacterial taxa Gelria glutamica, Mitsuaria chitosanitabida, and Burkholderia soli and increases in the relative abundance of the bacterial taxon Desulfotomaculum ruminis[19]. The bacterial diversity of HBPs was further analyzed, and the results demonstrated that plant extracts changed the community structure of the screened HBPs in soil, evidenced by a clear separation of Bray-Curtis distance determined by principal coordinate analysis (PCoA) (Supplementary Fig. S3a). The Shannon and Simpson indices, representing the α diversity of HBPs, showed no significant difference among all groups (Supplementary Fig. S3b, c). However, the richness indices decreased significantly in soil supplied with CUR, AG, and THY, suggesting that plant extracts reduced the abundance of HBPs (Supplementary Fig. S3d). Taken together, these results demonstrated that the plant extracts used in this study had great potential to reduce the abundance of HBPs in soil.
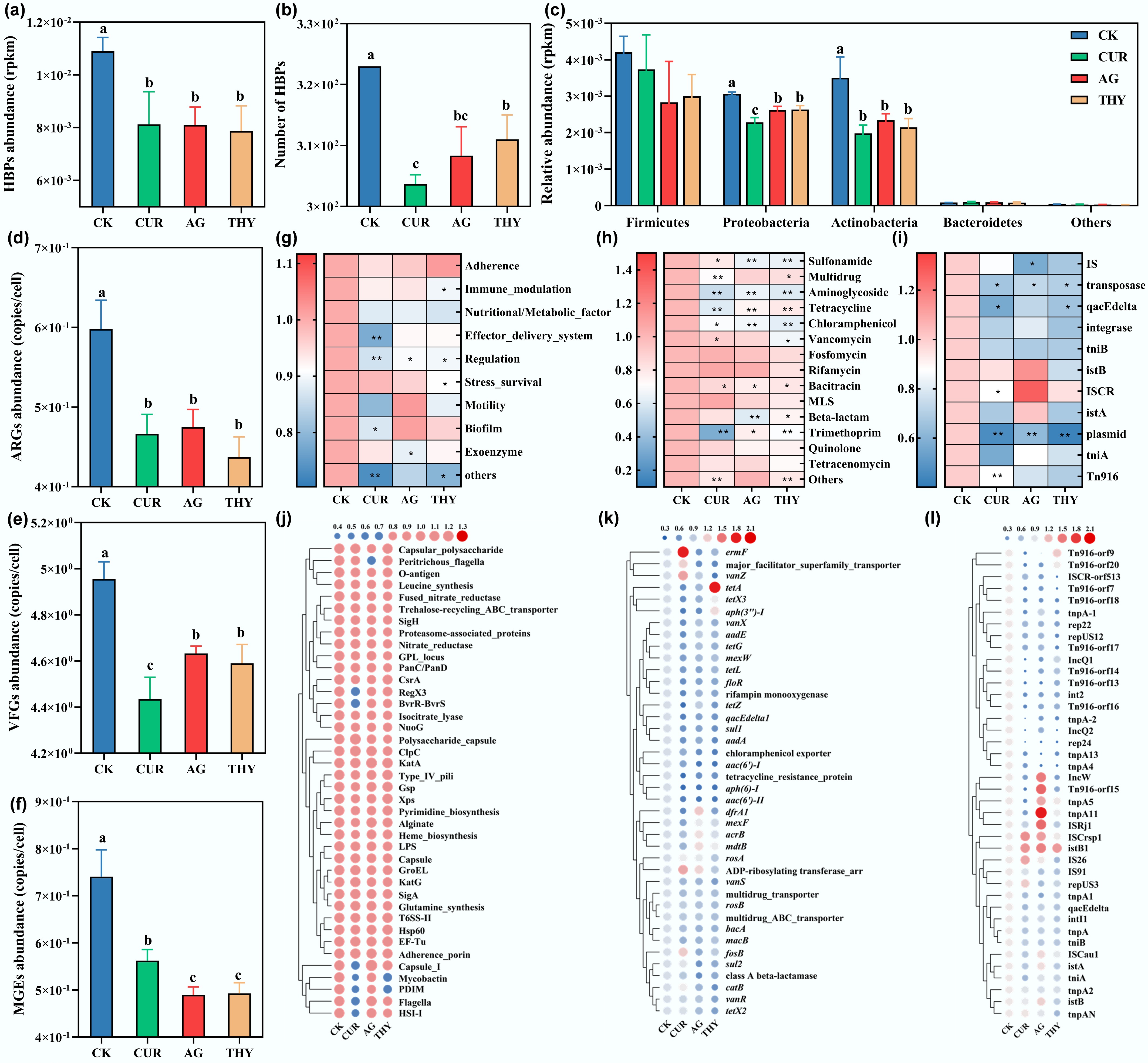
Figure 1.
Effects of plant extracts on the relative abundance and risk of HBPs in soil. (a) Total abundance, and (b) number of HBPs in soil treated with CUR, AG, and THY. (c) Effects of CUR, AG, and THY on the major bacterial phylum of HBPs. The total abundance of (d) ARGs, (e) MGEs, and (f) VFGs in soil treated with plant extracts. Effects of CUR, AG, and THY on the main categories of (g) ARGs, (h) MGEs, and (i) VFGs. The abundance of the top 40 subclasses of (j) ARGs, (k) MGEs, and (l) VFGs in soil treated with plant extracts. Data points with the same letter suggest no significant difference (p > 0.05).
Whether a decrease in the abundance of HBPs reduces their risk remains unclear. Considering that VFGs are an important factor reflecting the pathogenicity of HBPs, changes in the abundance of VFGs in soil were first analyzed. The result showed that the abundance of VFGs was decreased by 10.52%, 6.51% and 7.39% under the treatment of CUR, AG and THY, respectively (Fig. 1d). Fourteen categories of VFGs were screened, and it was observed that CUR reduced the VFGs related to effector delivery system, regulation and biofilm; AG decreased the VFGs related to regulation and exoenzyme; THY decreased the VFGs related to immune modulation, regulation and stress survival (Fig. 1g). At the same time, the VFG subclasses that ranked in the top 40 in abundance were also analyzed, and most VFGs were inhibited by CUR, AG, and THY (Fig. 1j). Collectively, these results demonstrated that plant extracts effectively reduced the pathogenicity of HBPs.
Another factor that cannot be ignored is the transmission risks of HPBs. Next, the influence of plant extracts on the transmission risks of HBPs by analyzing ARG abundance was investigated. Compared with CK, the abundance of ARGs in soil decreased by 21.94%, 20.50%, and 26.87% after treatment with CUR, AG, and THY, respectively (Fig. 1e). A total of 14 major ARG categories were screened. It was mainly composed of Sulfonamide, Multidrug, Aminoglycoside, Tetracycline, Chloramphenicol, Vancomycin, Fosmidomycin, Rifamycin, Bacitracin, and Macrolide-lincosamide-streptogramin (MLS), Beta-lactam, Trimethoprim, Quinolone, and Tetracenomycin, which had also been detected in chicken manure, bovine manure, and sheep manure[34,35]. Compared with CK, the abundance of most ARGs was reduced (Fig. 1h), consistent with previous studies where the abundance of ARGs was lower in the rhizosphere of lettuce, endive, and ryegrass grown in manure-amended soils[31,36]. Similarly, the application of CUR, AG, and THY also reduced the abundance of ARGs subclasses ranked in the top 40 (Fig. 1k). The above results showed that plant extracts effectively reduce the transmission risk of HBPs. Therefore, it was speculated that the decrease in ARGs and VFGs mediated by plant extracts was responsible for the decreased risk of HBPs. However, different results were observed in previous studies. For example, the relative abundances of sul1, ermB, tetW, or int1 genes in soils planted with lettuce, radish, and broccoli were not significantly affected[37]. Alternatively, some studies observed that the relative abundances of ARGs, such as tetX genes, in the rhizosphere were higher than those in bulk soils[36]. These discrepant phenomena may be ascribed to the different environmental factors, such as soil types, plant species, antibiotic contents, and plant extract types, which influence the proliferation and transfer of ARGs, thus leaving a significant gap to be filled to acquire comprehensive knowledge of the effects of the plant extracts on the distribution and composition of ARGs[30].
VFGs and ARGs can be transferred among different bacterial hosts via HGT mediated by MGEs, thereby increasing the likelihood that antibiotic-sensitive pathogens acquire antibiotic resistance[38,39]. Therefore, the effects of plant extracts on the abundance of MGEs in soil were then analyzed. The abundance of MGEs was significantly decreased with the addition of plant extracts (Fig. 1f). A total of 11 categories of MGEs, including insertion sequences (IS), transposon elements (Tn916, TniA, and TniB), integrases, plasmids, and transposon enzymes, were screened. The abundance of most MGEs decreased (Fig. 1i), in line with a previous study that found the relative abundance of MGEs was lower in the rhizosphere of maize, rice, and soybean than in bulk soils[3]. Similarly, analysis of the top 40 MGE subclasses in abundance also showed that plant extracts decreased the abundance of MGE subclasses such as tnpA1, TNPA-2, tnpA5, tnpA13, rep22, rep24, Inc, repUS12 and Tn916 (Fig. 1l). Previous studies have demonstrated that the presence of MGEs can promote the dissemination and proliferation of ARGs and VFGs, thereby increasing the health risks posed by HBPs[40,41]. The results also showed that ARGs were significantly correlated with MGEs (r = 0.909, p < 0.01) (Supplementary Fig. S4a), VFGs and MGEs were also significantly correlated (r = 0.711, p < 0.01), showing a linear positive correlation (Supplementary Fig. S4b), consistent with previous studies[42,43]. Collectively, these results confirmed that the reduction of MGEs was closely associated with decreases in VFGs and ARGs.
To further verify the reduced risks of HBPs mediated by plant extracts, co-occurrence analysis was performed using HBPs ranked in the top 100, ARGs ranked in the top 40, and VFGs ranked in the top 40. The network consisted of 157 nodes and 352 edges, and HBPs had a strong correlation with ARGs and VFGs (r ≥ 0.70, p < 0.05) (Supplementary Fig. S5c), which was further evidenced by a good linear positive correlation between HBPs and ARGs (Supplementary Fig. S5a), as well as VFGs (Supplementary Fig. S5b), consistent with previous studies[42,43]. Specifically, multidrug resistance gene, tetracycline resistance gene, aminoglycoside resistance gene, nutritional/metabolic_factor related gene, immune_modulation related gene, and regulation related gene were dominant in the network, and Proteobacteria (n = 30), Actinobacteria (n = 27), Firmicutes (n = 14) and Bacteroidetes (n = 6) are the primary hosts of ARGs and VFGs, in line with previous studies[2,30,44] Previous studies have demonstrated that co-hosting of ARG-VFGs by HBPs has the most significant potential risk, compared to single hosting of ARGs or VFGs[45,46]. In this study, a total of 56 high-risk HBPs co-hosting ARGs and VFGs, such as B. cereus, B. subtilis, S. maltophilia, C. xerosis, and M. tuberculosis, were screened. Their abundances decreased by ~29.85% under plant extract treatment, while no significant difference was observed among the treatment groups (Supplementary Fig. S5d). Specifically, it was found that CUR reduced the abundance of C. xerosis by 68.00% and S. maltophilia by 29.49%, AG reduced the abundance of B. subtilis and C. xerosis by 38.02% and 45.31%, respectively, THY reduced the abundance of B. subtilis and C. xerosis by 55.29% and 47.48% (Supplementary Fig. S5e), respectively. Taken together, the above results demonstrated a significant correlation among ARGs, VFGs, and HBPs, and concluded that the decrease in ARGs and VFGs mediated by plant extracts reduced the risk of HBPs.
Plant extracts disturb the QS of HBPs in soil
-
Previous studies have demonstrated that the risks of HBPs are closely related to their QS systems[4,24]. Therefore, the next investigated was the influence of plant extracts on the QS systems of HBPs. Based on metagenomic data and QS-related genes on the KEGG website, QS-related genes are screened, and they mainly include synthetic and receptor genes. As shown in Fig. 2a, the application of CUR, AG, and THY significantly reduced the relative abundance of QS genes by 12.60%, 7.02%, and 9.83%, respectively, consistent with previous studies[27,47,48]. The relative abundance of synthesis-related and receptor-related genes decreased in the CUR, AG, and THY treatment groups (Fig. 2b, c), suggesting that the expression of these genes was affected by plant extracts. Furthermore, a total of 57 QS-related genes, including 20 synthesis-related genes and 37 receptor-related genes, were screened (Fig. 2d), and the results showed that CUR mainly inhibited the expression of rhlI, agrB, pqsE, nisC, cepR, cciR, agrA, and agrC genes; AG mainly inhibited the expression of rhlI, cviI, agrB, and nprB genes; THY mainly inhibited the expression of avsI, cviI, nisC, nisB, nprB, cepR, agrC, and nprR genes (Fig. 2d). Collectively, these results demonstrated that the QS system of HBPs was disturbed by plant extracts.
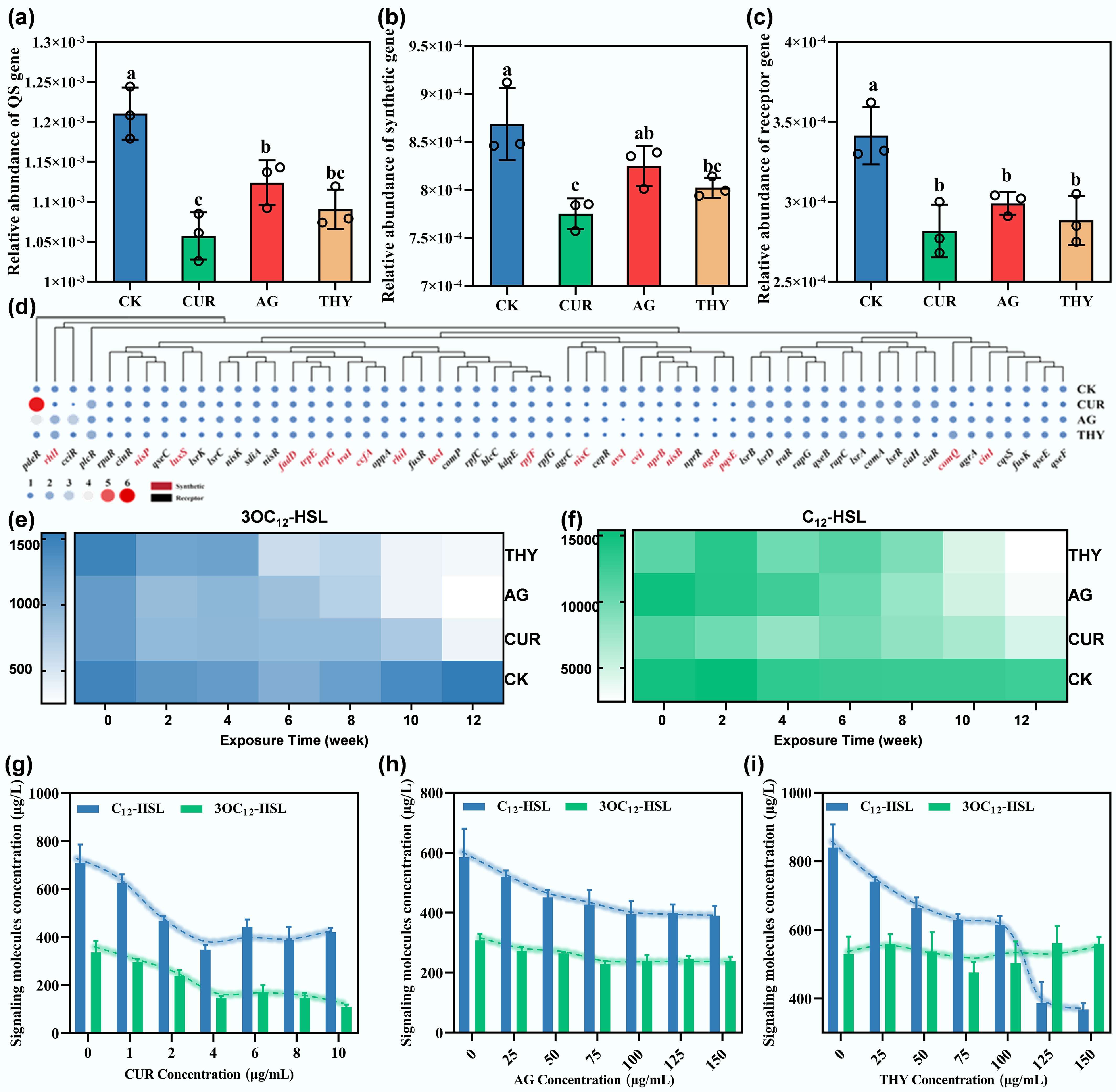
Figure 2.
Effects of plant extracts on the QS of HBPs in soil. Changes in the abundance of (a) total QS genes, (b) QS-related synthetic genes, and (c) QS-related receptor genes in the soil after application of different types of plant extracts. Different lowercase letters indicate significant differences among the treatments at p < 0.05. (d) Effects of plant extracts on soil-specific QS-related genes. Effect of plant extracts on the concentrations of (e) 3OC12-HSL, and (f) C12-HSL in the soil microcosms at different incubation times. Effects of different concentrations of (g) CUR, (h) AG, and (i) THY on the secretion of signaling molecules by PAO1.
The QS system works through signal molecules. According to the different types of synthesized signal molecules, QS genes can be divided into five categories, including AHL, AI-2, NisP, AI-3, and HHQ/PQS, and it was clearly observed that AHL-related genes are the most common identified QS genes, accounting for more than 24% of the total (Supplementary Fig. S6). Considering that AHLs are the most common QS signal molecules and that more than 50 Gram-negative bacteria use them for bacterial communication[21], the next focus was on analyzing AHL-related genes in detail. Under the treatment of CUR, AG, and THY, the abundance of AHL genes was decreased by 27.74%, 20.99%, and 15.37%, respectively (Supplementary Fig. S6). Next, the concentration of AHL signal molecules (3OC12-HSL and C12-HSL) was determined. As shown in Fig. 2e, f, the concentrations of 3OC12-HSL and C12-HSL in CK did not change significantly over time. In contrast, the concentrations of 3OC12-HSL and C12-HSL in the plant extract-supplemented treatment group gradually decreased over time. To confirm this result, further investigation was done on the influence of different concentrations of plant extracts on the secretion of signals by PAO1, a key HBPs species. The results showed that plant extracts also reduced the concentration of 3OC12-HSL and C4-HSL secreted by PAO1, and the higher the concentration, the stronger the inhibition effect (Fig. 2g–i). In addition, as expected, QS-related gene expression in PAO1 was also downregulated. For example, CUR and AG significantly inhibited the expression of lasI, lasR, and rhlI of PAO1 by 56.50%, 39.15% and 38.94%, respectively. At the same time, THY inhibited the expression of lasI, lasR, rhlI, and rhlR by 28.53%, 32.39%, 44.29% and 36.98%, respectively (Supplementary Fig. S7). Taken together, it was concluded that plant extracts disturb the QS in HPBs by reducing the concentration of AHLs in soil.
Whether disturbed QS in HBPs mediated by plant extracts is associated with reduced risk remains unclear. To answer this question, the relationship between the abundance of HBPs, ARGs, MGEs, VFGs, and the total abundance of QS genes was further analyzed. It was found that the total abundance of QS genes was positively correlated with the abundance of HBPs (r = 0.779, p < 0.01), ARGs (r = 0.85, p < 0.05), VFGs (r = 0.891, p <0.01), and MGEs (r = 0.7, p < 0.05) (Supplementary Fig. S8). Similarly, the abundances of synthesis-related and receptor-related genes were positively correlated with those of HBPs, ARGs, MGEs, and VFGs (Supplementary Fig. S8). These results demonstrated that the plant extracts significantly reduced QS-related genes in soil, which may inevitably disturb communication within or among bacterial species, thereby contributing to decreases in ARGs, VFGs, MGEs, and HBPs in abundance.
Mechanism of the reduced risk of HBPs mediated by plant extracts by disturbing QS
Plant extracts reduced the virulence factor secreted by HBPs
-
Given that QS regulates the secretion of virulence factors, disruption of the QS system may also affect virulence factor secretion. To test this hypothesis, the concentration of several virulence factors was measured. The results showed that, compared with CK, the concentrations of rhamnoolide, elastase, and pyocyanin in the plant extract treatment groups decreased by 37.76%–51.35% (Fig. 3a), 52.64%–83.56% (Fig. 3b), and 50.00%–50.76% (Fig. 3c), respectively. Moreover, expression of genes encoding virulence factors was also decreased (Fig. 3d). For example, lasB, a gene related to rhamnolipid regulation, decreased by 34.70%–43.65% in the presence of plant extracts. The genes rhlA and rhlB encoding elastase decreased by 37.46%–57.15% and 41.74%–57.05%, respectively. The genes phzS and phzM encoding pyocyanin decreased by 17.42%–47.79% and 24.00%–63.49% in the presence of plant extracts, respectively. Taken together, these results confirmed that plant extracts reduced the secretion of virulence factors of PAO1 by disturbing its QS system, as illustrated in Fig. 3e.
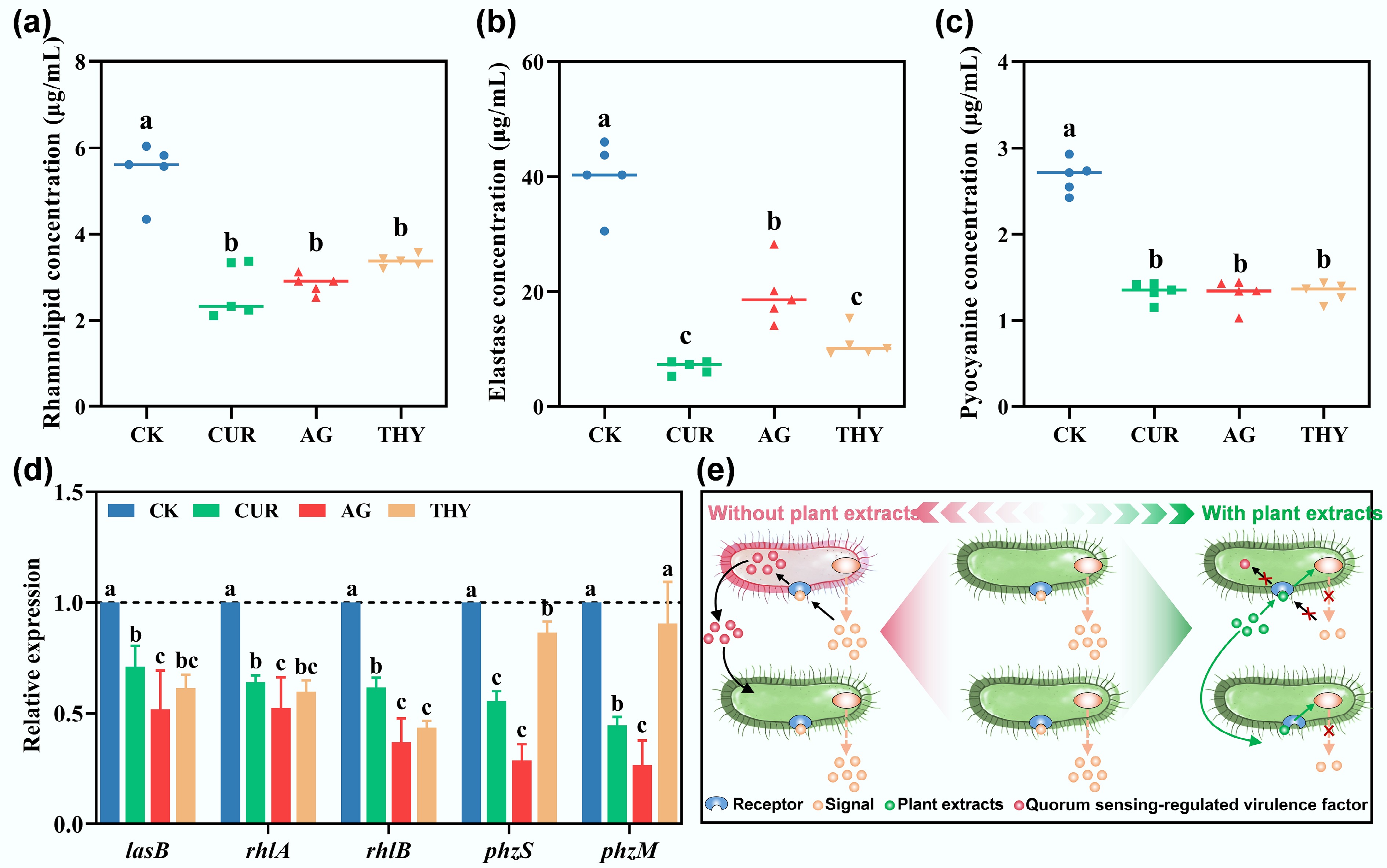
Figure 3.
Effects of different plant extracts on virulence factors (a) rhamnolipid, (b) elastase, and (c) pyocyanin secreted by PAO1. (d) Effects of different plant extracts on virulence factor-related genes of PAO1. (e) Schematic illustration of the reduction of virulence factors of PAO1 via disturbing their QS system mediated by plant extracts. Data points with the same letter indicate no significant difference (p > 0.05).
Plant extracts reduced the formation of biofilms of HBPs
-
The formation of biofilm enables bacteria to remain in EPS for longer, which promotes the exchange and recycling of nucleic acids[49]. ARGs and VFGs can be transmitted through HGT, and the biofilm itself is also a virulence factor[50]. Previous studies have demonstrated that biofilms formed by HBPs can accelerate the rate of colonization of HBPs and cause serious diseases and infections[51]. PAO1, one of the typical HBPs, was selected to investigate the influence of plant extracts on the formation of biofilms. The results showed that the biofilm of PAO1 was reduced by 26.54%, 38.13% and 39.41% under the treatment of CUR, AG, and THY, respectively, suggesting that plant extracts effectively reduced the formation of PAO1 biofilm (Fig. 4a). A. baumannii was further applied to verify the above result, this is because it also uses AHLs as signaling molecules[52]. As expected, the formation of A. baumannii biofilm was also inhibited after the treatment of plant extracts (Fig. 4b). Taken together, these results indicate that plant extracts can effectively reduce biofilm formation by bacteria, potentially reducing the transmission of ARGs and VFGs through HGT.
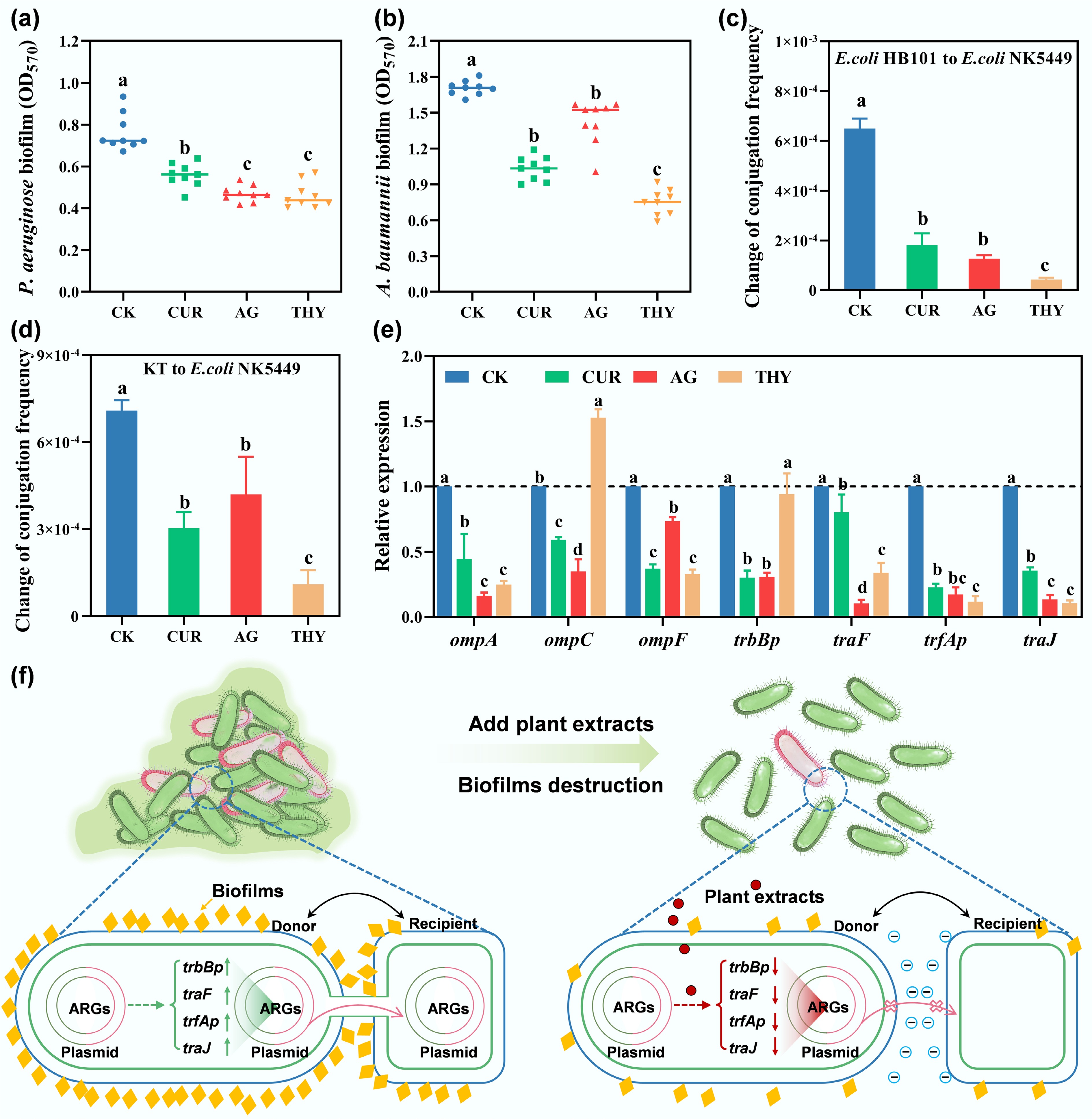
Figure 4.
Effects of different plant extracts on the formation of biofilms of (a) PAO1 and (b) Acinetobacter baumannii (A. baumannii). Effects of plant extracts on (c) the intra-species, (d) inter-species transfer conjugation frequencies, and (e) genes associated with conjugative transfer. Data points with the same letter indicate no significant difference (p > 0.05). (f) Illustrative diagrams for the inhibition of the conjugative transfer of ARGs and VFGs with the addition of plant extracts.
Plant extracts inhibited the conjugative transfer of ARGs and VFGs
-
Conjugative transfer is the most common pathway in HGT. To assess the influence of plant extracts on the transmission of ARGs and VFGs, donor strains E. coli HB101, P. putida KT2442, and recipient strains resistant to rifampicin containing the multi-resistant plasmids RP4 (tetracycline, ampicillin, and kanamycin), NK5449 were used for intra-species and inter-species transfer[53]. The results showed that plant extracts effectively reduced the frequency of interspecies transfer by 72.19%–93.56% (Fig. 4c) and indirect co-transfer by 41.13%–84.64% (Fig. 4d), respectively, suggesting that plant extracts have the potential to reduce the spread of ARGs and VFGs.
To further verify the above conclusion, the relative expression of several typical genes, including ompA, ompC, and ompF53, was determined. This is because previous studies have shown that bacteria can control the flow of foreign DNA and other substances by regulating membrane permeability through outer membrane proteins (OMPs)[54]. As shown in Fig. 4e, under the treatment of CUR, AG, and THY, the expression of ompA, ompC, and ompF genes decreased by 55.62%–83.92%, 41.06%–65.16%, and 26.59%–67.32%, respectively. This suggested that plant extracts altered membrane permeability by disturbing the OMPs, thereby inhibiting the transfer of ARGs. Conjugative pairing system (Mpf) and plasmid transfer replication system (Dtr) are directly related to the conjugative transfer system, and trbBp is the key promoter in Mpf, while traF is involved in the DNA processing and pairing formation. After the addition of plant extracts, the expression of trbBp and traF genes decreased by 58.43%–70.56% and 64.66%–89.56%, respectively. In the Dtr system, trfAp was considered to be a potent promoter of junction transfer. It was observed that trfAp expression decreased by 77.50%–88.56%, and traJ expression in the Dtr system decreased by 64.66%–89.56%. The inhibition of the conjugative transfer of ARGs and VFGs with the addition of plant extracts was illustrated in Fig. 4f. Taken together, these results demonstrated that the inhibition of plant extracts on the spread of ARGs was mainly attributed to the disturbance of membrane permeability, conjugative pairing system, and plasmid transfer replication system[55].
Plant extracts disturbed QS by competitively binding to LasR with signal molecules
-
Considering the impact of plant extracts on regulatory genes and signal molecules of the QS system, molecular docking was applied to deeply uncover the structural basis underlying plant extracts binding to receptor proteins of the QS system, as changes in the protein structure due to ligand binding may affect their activity. Based on the soil-borne signal molecule monitoring data in Fig. 2, the concentration of 3OC12-HSL showed the most significant reduction. As widely recognized, the binding of LasR to 3OC12-HSL plays a pivotal role in initiating the transcriptional activation of downstream target genes in the QS system. Consequently, in the present study, the primary focus was on exploring the interaction between plant extracts and LasR via molecular docking. The three-dimensional structure of the LasR protein was obtained from the Protein Data Bank (PDB) in PubChem. As shown in Fig. 5a, four amino acid sites bind 3OC12-HSL to LasR; that is, 3OC12-HSL forms hydrogen bonds with His-258, Thr-224, and Try-218, and an alkyl interaction with Arg-225, consistent with a previous study[56]. Six amino acid sites bind CUR to LasR, and it forms a hydrogen bond with Lys-109, Ala-146, Asn-127, Phe-145, Ala-200, exhibiting Pi-alkyl interaction with Arg-199 (Fig. 5b). AG binds with the LasR protein via alkyl and Pi-alkyl interactions on Phe-145, Ile-129, Arg-199, Ala-200, and Try-163 (Fig. 5c). THY formed a hydrogen bond with Lys-246, exhibited Pi-alkyl interaction with Try-243, and had Pi-Sima with Phe-247 (Fig. 5d). Generally, hydrogen-bond interactions in the LasR protein-3OC12-HSL ligand complex stabilize and orient the 3OC12-HSL lactone ring, thereby triggering its transcriptional activity[56]. However, compared with 3OC12-HSL, plant extracts were superior to interact with LasR protein, which was evidenced by number of binding sites and the binding energy (Fig. 5, left panel), consistent with previous studies where they found the interaction among flavones, furanones, 6-Gingerol and the LasR protein were stronger than that of 3OC12-HSL[56,57]. Considering that the lower the binding energy, the more stable the ligand binding to the receptor protein will be, therefore, it was concluded that CUR had a higher affinity than that of AG, THY and 3OC12-HSL. Finally, the binding affinity of 3OC12-HSL and plant extracts for the LasR protein was measured using a molecular interaction analyzer to verify the molecular docking results further. The results showed that the binding affinity of 3OC12-HSL to LasR protein was 2.884e−7 J/mol. The binding affinities of CUR, AG, and THY to the LasR protein were 2.764e−8, 4.835e−8, and 6.474e−7 J/mol, respectively, consistent with the molecular docking results. Based on the above discussion, it was concluded that changes in protein structure due to ligand binding may cause changes in the LasR protein's activity and function, then disturbing the QS system.
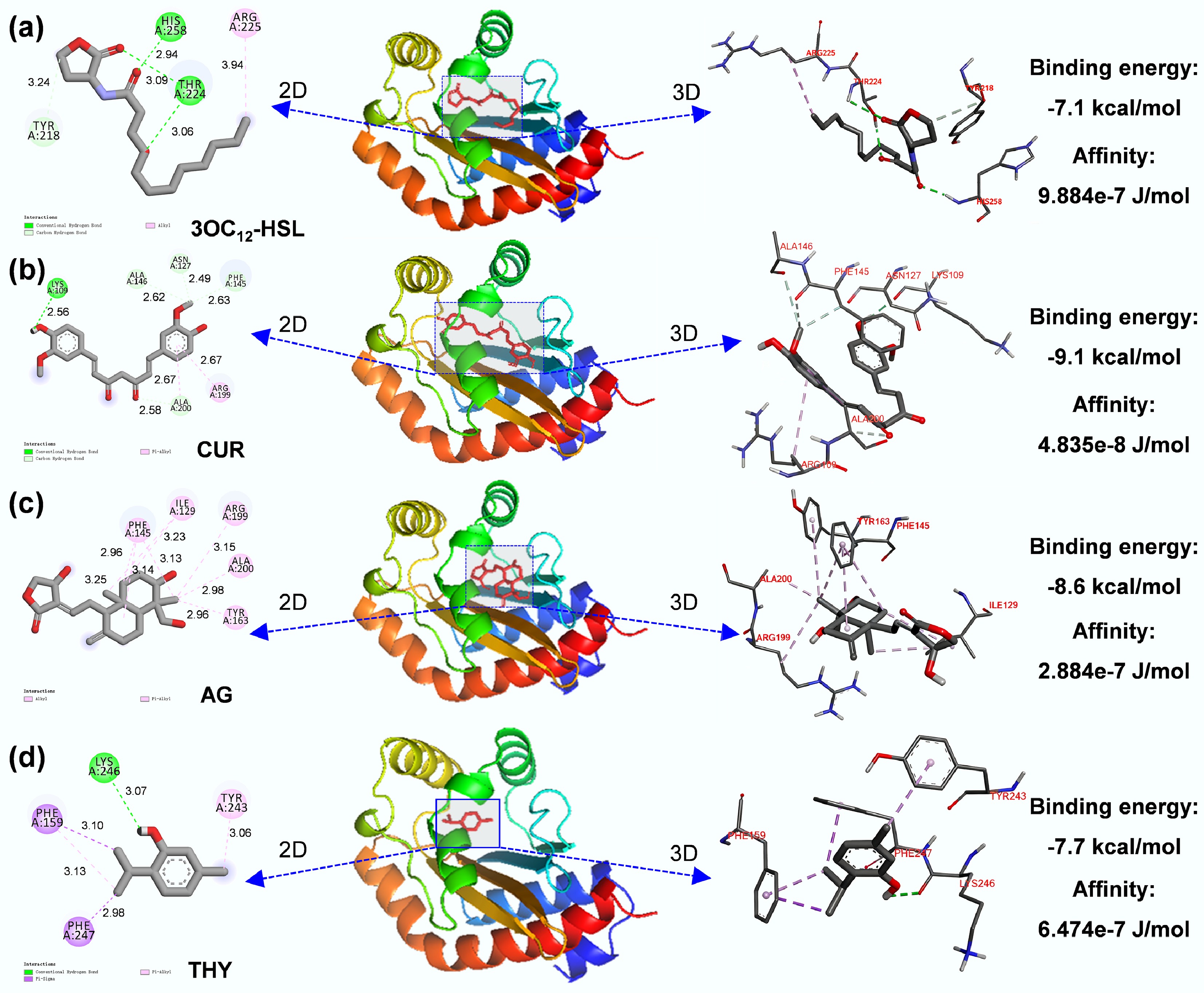
Figure 5.
Molecular docking predicted amino acid sites for the binding of (a) 3OC12-HSL, (b) CUR, (c) AG, and (d) THY and LasR protein.
-
In conclusion, the results demonstrate that plant extracts bind strongly to QS-related protein sites, thereby inhibiting signal generation and QS gene expression, ultimately disturbing the QS system. Once the QS system of HBPs is disturbed, their communication is blocked, as is biofilm formation, inhibiting the HGT of ARGs and VFGs among HBPs and leading to reduced abundance of ARGs, VFGs, and HBPs, as well as increased abundance and risks of HBPs. Collectively, this study uncovers the important role of plant extracts on the attenuation of microbial contaminants (HBPs, ARGs, VFGs, and MGEs) in soil, which not only gives new insights into understanding the environmental behavior of microbial contaminants in the agricultural soil, but also provides an important support for developing plant-derived substrates for the remediation of microbial contaminants in soil.
However, numerous studies have confirmed that these plant extracts possess antibacterial properties. While they had no significant effect on bacterial growth at the dosage used in this study, the complexity of the soil environment is far greater than that of the laboratory pure culture system. Factors such as soil pH, organic matter content, and microbial community structure may alter the activity and stability of plant extracts, thereby affecting the intensity of their antibacterial effects. Meanwhile, the cumulative effect of plant extracts in the soil over long-term application may also enhance their direct bactericidal activity, thereby forming a synergistic or additive effect with the QS interference mechanism. Therefore, future studies need to accurately quantify the relative contributions of direct antibacterial effects and the QS interference mechanism. Additionally, it is necessary to explore the regulatory laws of soil physicochemical properties on these two mechanisms, thereby providing a more comprehensive theoretical basis for the precise application of plant extracts in the remediation of soil biological pollution.
-
It accompanies this paper at: https://doi.org/10.48130/biocontam-0025-0009.
-
Not applicable.
-
The authors confirm contributions to the paper as follows: study conception and design: Guo F, Lu K, Zhang J, Zhu L, Li N, Wang M; material preparation, data collection and analysis: Guo F, Lu K, Zhang J, Zhu L, Li N, Wang M; draft manuscript preparation: Guo F, Lu K, Wang M; manuscript review and revision: Zhu L, Li N. All authors reviewed the results and approved the final version of the manuscript.
-
All data analyzed to evaluate the conclusions are available from the authors upon reasonable request. Moreover, the sequence data is publicly available: NCBI-PRJNA1356285.
-
We acknowledge the financial support from the 'Leading Goose' R&D Program of Zhejiang (Grant No. 2024C03131), the National Key R&D Program of China (Grant No. 2022YFC3704600), and the National Natural Science Foundation of China (Grant Nos 22122607, U21A20292, 22376097, and 22306164).
-
The authors declare that they have no conflict of interests.
-
# Authors contributed equally: Fangjie Guo, Kun Lu
Full list of author information is available at the end of the article. - Supplementary Table S1 Soil moisture, available nitrogen (AN), available phosphorus (AP), pH, and electrical conductivity (EC).
- Supplementary Table S2 Primer sequences tested in this study.
- Supplementary Table S3 HPLC–MS/MS operating conditions.
- Supplementary Table S4 The optimum parameters of liquid chromatography–tandem mass spectrometry.
- Supplementary Text S1 DNA Extraction, metagenomic sequencing, data processing, and analysis.
- Supplementary Text S2 Detection of QS signals.
- Supplementary Text S3 Effects of root extracts on the virulence factors.
- Supplementary Text S4 Effects of root extracts on the formation of biofilm.
- Supplementary Text S5 Effects of root extracts on the horizontal fene transfer.
- Supplementary Fig. S1 Effects of (a) CUR, (b) AG and (c) THY on the growth and the secreted signals (C12-HSL and 3OC12-HSL) of PAO1.
- Supplementary Fig. S2 Effects of different plant exudates on soil bacterial composition. (a) Bray-Curtis distance-based PCoA analysis of soil bacterial composition after 12 weeks of incubation. Changes in bacterial diversity were determined using (b) the Shannon index and (c) the Simpson index. (d) Effects of adding different plant exudates on the relative abundance of soil microorganisms.
- Supplementary Fig. S3 Effects of root exudates on the diversity of HBPs. (a) PCoA based on Bray-Curtis distance analysis of the composition of HBPs after 12 weeks of incubation. (b) Shannon index, and (c) Simpson index. (d) Richness index of the HBPs community.
- Supplementary Fig. S4 (a) Linear regression analysis of MGEs and ARGs. (b) Linear regression analysis of MGEs and VFGs.
- Supplementary Fig. S5 (a) Linear regression analysis of ARGs and HBPs. (b) Linear regression analysis of VFGs and HBPs. (c) Effects of different root exudates on the abundance of high–risk HBPs in soil. Different lowercase letters indicate significant differences among the treatments at p < 0.05. (d) Relationship between ARGs, VFGs, and HBPs. (e) Species and abundance of high-risk HBPs.
- Supplementary Fig. S6 (a) The main QS gene categories in the soil. Effects of applying root exudates on the abundance of (b) AHL–class, (c) AI–2 class, (d) NISP–class, (e) AI–3 class, and (f) HHQ/PQS class genes. Different lowercase letters indicate significant differences among the treatments at p < 0.05.
- Supplementary Fig. S7 Effects of different root extracts on QS–related genes of PAO1. Data points with the same letter indicate no significant difference (p > 0.05).
- Supplementary Fig. S8 Linear regression analysis of QS–related gene abundance with (a) HBPs, (b) ARGs, (c) VFGs, and (d) MGEs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Guo F, Lu K, Zhang J, Zhu L, Li N, et al. 2025. Plant extracts alleviate the risks of human bacterial pathogens from agricultural soil by disturbing quorum sensing. Biocontaminant 1: e008 doi: 10.48130/biocontam-0025-0009 

Plant extracts alleviate the risks of human bacterial pathogens from agricultural soil by disturbing quorum sensing
- Received: 16 September 2025
- Revised: 05 November 2025
- Accepted: 12 November 2025
- Published online: 26 November 2025
Abstract: Essential roles for plant extracts in shaping rhizosphere bacterial communities have been demonstrated. However, few studies have investigated the effects of plant extracts on the risks posed by human bacterial pathogens (HBPs) in agricultural soil, and the underlying mechanisms remain unclear. Herein, by conducting soil microcosms and bacterial pure culture experiments, it was found that the plant extracts significantly reduced the abundance and risks of HBPs in soils, evidenced by a 20.50%–26.87%, 6.51%–10.52%, and 24.60%–34.23% reduction in the abundance of antibiotic resistance genes (ARGs), virulence factor genes (VFGs), and mobile genetic elements (MGEs). More importantly, plant extracts inhibited the conjugative transfer of ARGs and VFGs among HBPs. Mechanistically, the plant extracts disturbed the quorum sensing (QS) of HBPs by reducing the concentration of signal molecules and down-regulating QS-related genes, an effect attributed to favorable intermolecular interactions between the plant extracts and QS proteins. Molecular docking results further showed that the binding energy and affinity of plant extracts to QS-related proteins were the main factors accounting for their QS interference effect. Collectively, this study will be beneficial for understanding the attenuation of microbial contaminants in soils mediated by plant extracts.
-
Key words:
- Plant extracts /
- Human bacterial pathogens /
- Risks /
- Quorum sensing /
- Molecular docking