









-
Flavanone 3-hydroxylase (F3H), catalyzing the conversion of naringenin into dihydroflavonols, is a key enzyme involved in plant flavonoid/anthocyanin biosynthesis pathways[1,2]. The expression levels of genes encoding F3Hs have been identified to be associated with flavonoid/anthocyanin accumulation in many plants. For example, in anthocyanin-rich 'Zhongshen 1' mulberry fruits, the expression level of F3H is positively correlated with the anthocyanin content[3]; the expression of CitF3H is positively correlated with the anthocyanin accumulation in juice sacs of blood orange 'Moro' during fruit ripening[4]; the anthocyanin reduce (are) tomato mutant, caused by the mutation in the F3H gene, has much higher accumulation of naringenin but significantly lower levels of downstream products including anthocyanins[5]; transgenic rice overexpressing OsF3H showed upregulated expression of downstream flavonoid biosynthesis related structural genes (OsDFR and OsFLS), and promoted accumulation of kaempferol, quercetin, delphinidin, and cyanidin[6]. The naringenin content in transgenic Arabidopsis seedlings overexpressing the Pohlia nutans F3H showed significantly lower accumulation of naringenin but much higher levels of downstream flavonoids, and enhanced salt resistance and antioxidant ability[7].
Evidence revealed that the expression of F3H genes could be greatly influenced by many environmental factors[8]. Blue light can promote the anthocyanin biosynthesis by upregulating the expression of CitF3H[4]. In grapes, glucose, fructose, and sucrose treatments can all induce F3H transcription and promote the biosynthesis and accumulation of anthocyanins in a hexokinase-dependent pathway[9]. Similarly, in Arabidopsis, sucrose treatment can activate the expression of AtF3H, thereby promoting the anthocyanin accumulation in seedlings[10,11]. Moreover, it was found that additional JA and ABA treatments can further enhance the sucrose-induced AtF3H expression and anthocyanin accumulation[11]. The expression of Dendrobium officinale F3H was reported to be salt- and cold-inducible, and its upregulation resulted in increased anthocyanin and flavonol accumulation and conferred plants with a tolerance to these stresses[12]. Tobacco plants overexpressing an F3H from Lycium chinense showed upregulated expression of DFR and ANS genes, improved flavon-3-ol accumulation, and enhanced antioxidant ability[13].
Recent research demonstrated the transcriptional regulation of transcription factors (TFs) on F3H genes. The mulberry ERF5 can bind to the 'GCCGAC' sequences on the promoters of anthocyanin-related MYBA and F3H genes, activating their expression and promoting anthocyanin biosynthesis in fruits[14]. The rice OsC1PLSr functions in regulating anthocyanins biosynthesis and accumulation by activating the expression of anthocyanin biosynthesis structural genes, including F3H[15]. Erigeron breviscapus EbMYBP1 can directly bind to the promoter of EbF3H, activate the gene's transcription, and promote flavonoid biosynthesis and accumulation in leaf[16].
Blueberry, a perennial woody plant, is known for its anthocyanin-rich fruits. Its fruits have very high health-care values, with anthocyanins as the main contributors[17,18]. Currently, there are some reports on the functions of blueberry F3H genes in anthocyanin biosynthesis. For example, Zhang et al.[19] cloned a F3H gene (named VcF3H) from blueberry leaf, and verified its anthocyanin accumulation promoting effect through heterologous expression in Arabidopsis. Lin et al.[20] identified several anthocyanin biosynthesis structural genes, including a F3H, from blueberry transcriptome data, and found that the expression levels of these genes are positively correlated with anthocyanin content. Xu et al.[21] studied the influences of dielectric barrier discharge cold plasma (CP) on anthocyanin accumulation, and found that CP treatment could upregulate the expression of F3H and some other structural genes in post-harvested blueberry fruits. To date, however, there is no report focusing on the systematic identification, characterization, and functional analysis of anthocyanin-related F3H genes in blueberry. In this study, we first identified and characterized all the anthocyanin-related F3Hs in the blueberry genome, and investigated their expression patterns under different abiotic stress treatments in fruits at five different ripening stages using transcriptome data and quantitative real-time PCR (qRT-PCR) analysis. Furthermore, the function of VcF3H2, an anthocyanin-related VcF3H showing high expression in fruits and the most significant differential expression changes during fruit ripening, was studied using transient overexpression assays in apple and blueberry fruits. It is worth noting that, many binding sites for TFs, such as MYB and bHLH, were found in the promoters of VcF3H2 and some other anthocyanin-related VcF3Hs. To reveal the possible transcriptional regulation of TFs on VcF3Hs, the binding and activation abilities of two anthocyanin biosynthesis regulatory TFs, VcMYB-1[22] and VcAN1[23,24], on the VcF3H2 promoter were verified using dual-luciferase assays (LUC) and yeast one-hybrid (Y1H). Our study will be helpful for understanding the roles of F3H in blueberry anthocyanin biosynthesis and can provide a basis for clarifying the transcriptional regulation of TFs in anthocyanin biosynthesis.
-
The blueberry plants used in this study were healthy and unique two-year-old 'Legacy' and 'FL03' seedlings cultivated in our lab. The green, pink, red, purple, and blue blueberry fruits used for gene cloning and expression analysis were harvested from two-year-old 'FL03' plants. Bagged 'Gala' apples used for transient overexpression analysis were harvested from the apple cultivation base at the Institute of Horticulture, College of Horticulture, Shanxi Agricultural University (Shanxi, China). For tobacco leaf transient overexpression, 6-week-old Nicotiana benthamiana plants were used.
Identification and characterization of anthocyanin-related F3H genes in blueberry
-
The genome data files of southern highbush blueberry (Vaccinium corymbosum) were downloaded from GDV (GENOMEDATABASE FOR VACCINIUM,
www.vaccinium.org ). The protein sequence of anthocyanin-related AtF3H (AT3G51240.1)[25] was downloaded from TAIR (www.arabidopsis.org ), and sequences of other anthocyanin-related F3Hs, including VvF3H (NM001281105.1)[9], OsF3H (Os04g0662600)[26], GmF3H (ACM62745.1)[27], LcF3H (AID50182.1)[28], PeF3H (AIS35916.1)[29], TfF3H (AGL98422.1)[30], were downloaded from NCBI (www.ncbi.nlm.nih.gov ). By using AtF3H, VvF3H and OsF3H as query sequences, local blastp was used to identify anthocyanin-related F3Hs in blueberry (E-value ≤ l ×10−5, identity ≥ 60%). Then, CDD (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ) was used to verify the presence of the PLN02515 superfamily domain in identified VcF3Hs.MEGAX was used for the alignment and phylogenetic analysis of VcF3Hs and flavonoid/anthocyanin-related F3Hs reported from other plants. A phylogenetic tree for these F3Hs was constructed using Neighbor-Joining method (Bootstrap = 1,000 and other parameters were set as default values), ITOL (
https://itol.embl.de/ ) was used for figure drawing. For the prediction of conserved motifs in anthocyanin-related VcF3Hs, MEME (http://meme-suite.org/tools/meme ) was used. TBtools software was used to visualize the motif distribution, and to predict the conserved domains in VcF3Hs and gene structures of their corresponding genes[31]. The Advanced Circos module of TBtools was used for the synteny analysis of anthocyanin-related VcF3Hs, and the Simple Ka/Ks Calculator (NG) program was used to calculate the duplication time among duplicated VcF3H gene pairs[32].The Protein Parameter Calc (ProtParam-based) module of TBtools and Plant-mPLoc (
www.csbio.sjtu.edu.cn/bioinf/plant-multi )[33] was used to predict the physiochemical proprieties and subcellular localization of VcF3Hs, respectively.Gene expression analysis
-
In our previous studies, we sequenced the transcriptomes of ABA (treated with 100 mM ABA), MeJA (100 mM MeJA), salt (200 mM NaCl and Na2SO4), alkali (200 mM NaHCO3 and Na2CO3), 4 °C low temperature (LT), and 40 °C high temperature (HT) treated blueberry leaves and 'FL03' fruits at five ripening stages. In this study, the FPKM (Fragments Per Kilobases per Million reads) values of anthocyanin-related VcF3Hs were extracted from these transcriptome data for heatmap drawing using TBtools. To further verify their expression patterns in these samples, quantitative real-time PCR (qRT-PCR) analysis of three selected VcF3Hs (VcF3H2, VcF3H6 and VcF3H8) was also performed with three replications. Primers used for the qRT-PCR verification were designed using primer3 (
http://primer3.ut.ee/ ) and listed in Supplementary Table S1. Amplifications were performed on a Bio-Rad CFX96™ real-time fluorescent quantitative PCR machine with a reaction procedure as follows: pre-denaturation at 95 °C for 30 s; denaturation at 95 °C for 15 s, annealing at 60 °C for 30 s, and extension at 72 °C for 30 s, 45 cycles. By using the GAPDH (AY123769)[34] as an internal reference gene, the relative expression of selected VcF3Hs in different blueberry samples was calculated using the 2−ΔΔCᴛ method.Prediction of VcF3Hs interacting proteins
-
Based on the Arabidopsis protein database, STRING v12.0 (
https://string-db.org/ ) was used to predict proteins interacting with VcF3Hs (minimum required interaction score > 0.700)[35]. Then, the Cytoscape software was used to draw the protein-protein interacting network figure of VcF3Hs[36].Promoter analysis
-
The 2,000 bp sequences upstream the start codon (ATG) of anthocyanin-related VcF3Hs were extracted from blueberry genome data using TBtools and considered as promoter sequences. By using PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ), and PlantRegMap (https://plantregmap.gao-lab.org/binding_site_prediction.php ), the cis-acting elements and transcription factor binding sites in promoters of anthocyanin-related VcF3Hs were predicted (p-value ≤ l × 10−5), respectively.Gene cloning and overexpression vector construction
-
The VcF3H2 coding sequence (CDS) was amplified through reverse transcription PCR by using 'FL03' blueberry fruit cDNA as template. After sequencing verification, the gene was subcloned and introduced into the pBI121 vector using the Ready-to-use Seamless Cloning Kit produced by Songon Biotech (Shanghai, China). Then, the obtained p35S::VcF3H2 recombinant vectors were transformed into Agrobacterium GV3101. Primers used for gene cloning and vector construction are listed in Supplementary Table S1.
Transient expression analysis of VcF3H2 in apple and blueberry fruits
-
According to the method described by Lv et al.[37], Agrobacteria GV3101 carrying p35S::VcF3H2 and empty pBI121 vectors (EV) were cultured to OD600 ≈ 1.5 in 50 mL of Luria-Bertani (LB) medium. Then,the Agrobacteria solution was centrifuged at 5,000 rpm for 4 min. After discarding the supernatant, the Agrobacteria pellet was re-suspended using MES solution, adjusted to OD600 = 0.8, shake-cultured at 200 rpm at 28 °C for 30 min, and kept still at 28 °C in the dark for 1 h to obtain the Agrobacteria infiltration solution used for apple and blueberry fruit injection.
For apple fruit transient overexpression analysis, approximately 30 μL Agrobacteria infiltration solution was injected into each site around the equatorial regions of apple fruits, and each fruit was injected at two opposite sites. Infiltrated fruits were kept in the dark at 24 °C for 2 d, followed by light conditions with a light density of 1,500 lx and photoperiod of 16 h light/ 8h dark. For blueberry fruit transient overexpression, green fruits on two-year-old 'Legacy' blueberry plants were used, and each fruit was injected with 10~15 μL Agrobacteria infiltration solution at one site. After agrobacteria infiltration, blueberry plants were then kept in the dark at 24 °C for 2 d, and removed to normal light condition. Five days later, CR8 colorimeter (3nh, Guangzhou, China) was used to measure the color parameters of apple and blueberry fruits. Then, the anthocyanins contents in apple and blueberry peels around the injected regions were determined according to the method of Zhang et al.[38].
Apple and blueberry fruit peel total RNA was isolated using Trizol (Invitrogen, Waltham, CA, USA), and reversed transcribed into cDNA using PrimeScript RT Master Mix (Perfect Real Time) kit (Takara, Dalian, China). These cDNA samples were used as templates for qRT-PCR analysis of anthocyanin biosynthesis structural genes (MdCHI, MdCHS, MdF3H, MdDFR, MdUFGT, and MdANS in apple[39], and VcCHS, VcDFR, VcF3H, VcUFGT, and VcANS in blueberry[40], Supplementary Table S1) in fruit peels.
Yeast one-hybrid (Y1H)
-
By using 'FL03' gDNA as a template, a 1,960 bp long VcF3H2 promoter sequence was amplified. Then, the promoter sequences were introduced into the pAbAi vector to obtain the recombinant vector pVcF3H21960, and transformed into the yeast strain Y1HGold. Then, pGADT7-VcAN1 and pGADT7-VcMYB-1 were individually transformed into the yeast strain carrying pVcF3H21960 to verify the binding activity of these two anthocyanin-related TFs on the VcF3H promoter. The pGADT7-VcAN1 and pGADT7-VcMYB-1 vectors were provided by our laboratory[23,24].
Dual-luciferase assay (LUC)
-
The 1,960 bp VcF3H2 promoter sequences were introduced into pNC-Green-LUC vector to obtain the LUC-pVcF3H21960 recombinant vector. The stop codon removed VcMYB-1/VcAN1 CDS sequences were introduced into the pNC-GreenII-SK vector to obtain the SK-MYB-1 and SK-AN1 recombinant vectors. Vectors were transformed into Agrobacteria GV3101 (pSoup-p19), and cultured till OD600 reached approximately 1. Agrobacteria carrying SK-MYB-1/SK-AN1 were equal volume mixed with the LUC-pVcF3H21960 Agrobacteria suspension, kept still at room temperature for 30 min, and then infiltrated into the abaxial side of N. benthamiana leaves. Simultaneously, pNC-Green-LUC + pNC-Green-SK, SK-VcMYB-1/VcAN1 + pNC-Green-LUC, and pNC-Green-SK + LUC-pVcF3H21960 Agrobacteria suspension mixture was also infiltrated into the same tobacco leaf at different parts. For each experiment, at least 20 tobacco leaves were used as replications. After infiltration, tobacco plants were cultured in dark at 25 °C for 2 d, and an additional 3 d under normal light conditions[41]. Then, infiltrated tobacco leaves were harvested, smeared with 1.5 mM luciferin substrate solution, kept in the dark for 5 min, and observed under a Tanon 5200 system (Tanon Science & Technology Co., Ltd., Shanghai, China).
-
In total, we identified eight candidate anthocyanin-related VcF3Hs from blueberry, which were named VcF3H1~8 according to their amino acid (AA) number and scaffold localization information of their corresponding encoding genes. Of them, VcF3H1~4 all have 359 AA, VcF3H5 and VcF3H6 have 366 AA, and VcF3H7 and VcF3H8 have 373 AA (Fig. 1a, Supplementary Table S2). Protein similarity analysis revealed that these eight VcF3Hs shared more than 90% similarities. The protein sequences of VcF3H2, VcF3H3, and VcF3H4 are the same, and there is only one amino acid difference between VcF3H2/3/4 and VcF3H1. The protein similarities among VcF3H5~8 are all higher than 96%. In consistence with their high sequence similarities, the motifs identified in anthocyanin-related VcF3Hs are the same in both numbers and orders (Fig. 1b). Conserved domain analysis results showed that they all contained a PLN02515 superfamily domain (Fig. 1c). Gene structure analysis revealed that all the eight VcF3Hs have two introns in their corresponding gDNA, and one additional intron was found in the 5'UTR of VcF3H1 (Fig. 1d). All the VcF3Hs were predicted to be localized in cytoplasm (Supplementary Table S2), which was consistent with the biosynthesis site of anthocyanins.
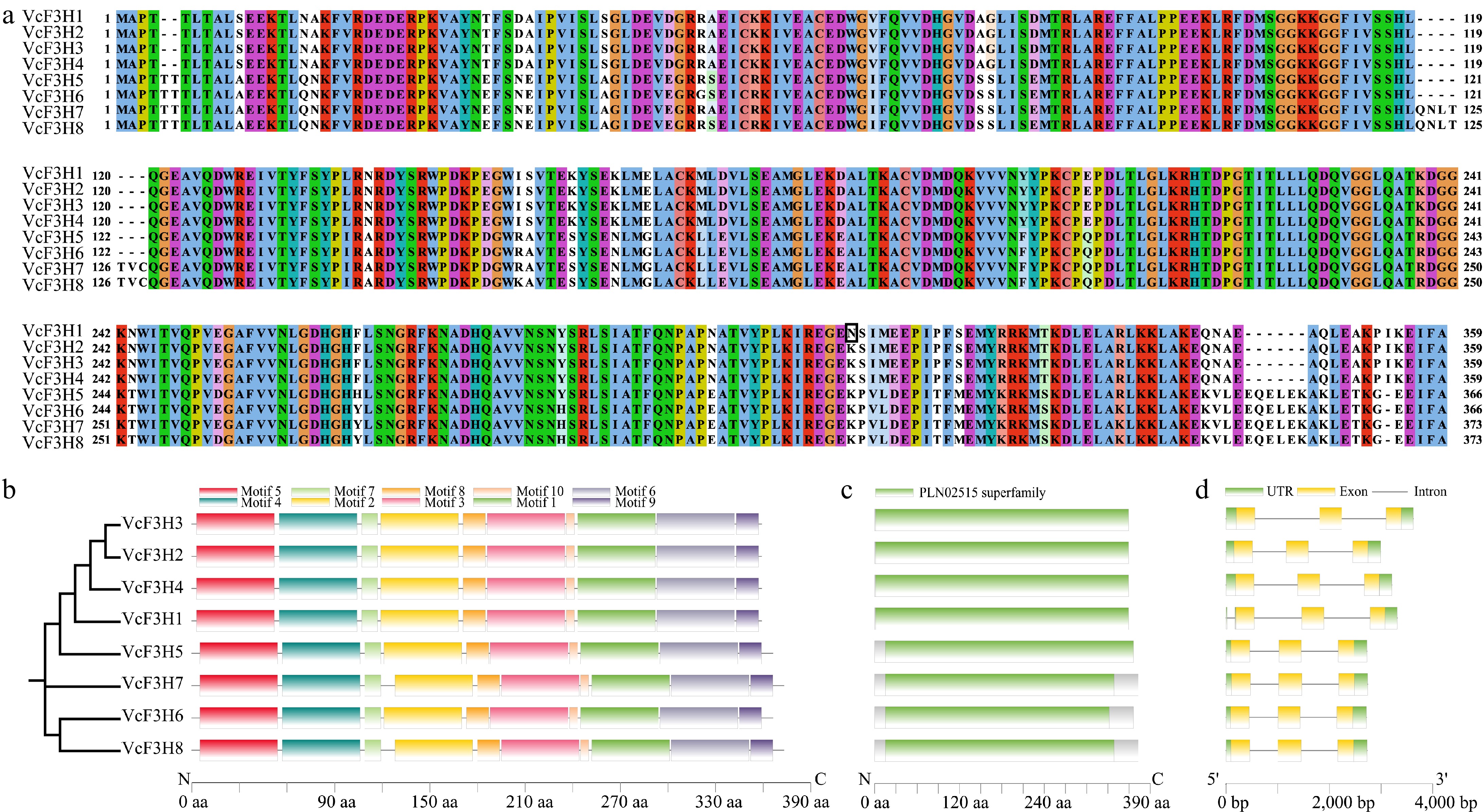
Figure 1.
(a) Sequence alignment, (b) conserved motifs, (c) domain, and (d) gene structure analysis results of the eight anthocyanin-related VcF3Hs identified in blueberry.
Phylogenetic analysis of flavonoid/anthocyanin-related F3Hs from blueberry, Arabidopsis, grape, rice, and some other plants revealed that VcF3H1~4 are close to each other, and VcF3H5/6/7/8 shared close relationships (Fig. 2a). The eight anthocyanin-related VcF3Hs shared greater more than 81% similarities with other reported flavonoid/anthocyanin-related F3Hs (Fig. 2b).
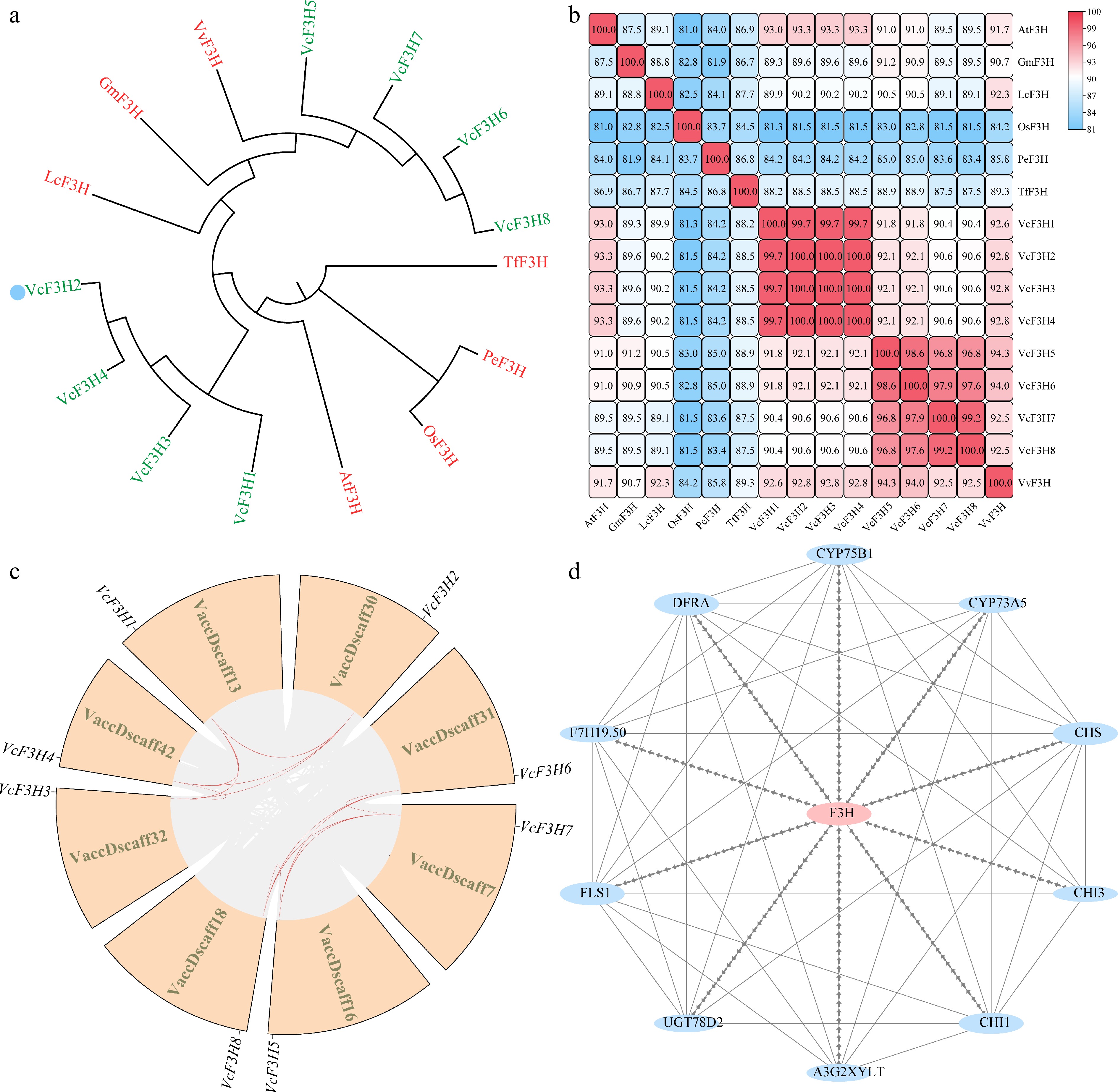
Figure 2.
(a) Phylogenetic, (b) similarity, (c) synteny, and (d) interacting protein analysis results of VcF3Hs.
Synteny analysis showed that there were 12 pairs of duplicated genes among the eight anthocyanin-related VcF3Hs, including six pairs among VcF3H1~4 and six pairs among VcF3H5~8 (Fig. 2c, Supplementary Table S3). The Ka/Ks values of all replicated gene pairs ranged from 0 to 0.1475, indicating that the anthocyanin-related VcF3Hs experienced strong purification selection pressure during evolution. By calculating the time of their duplication events, it was found that these gene replication events occurred 0 million years ago (Mya) ~0.28 Mya.
Based on the Arabidopsis protein database, the interacting proteins of anthocyanin-related VcF3Hs were predicted using STRING (Fig. 2d). They were all identified as homologous proteins of AtF3H, and their interacting proteins included many key enzymes involved in the flavonoids/anthocyanin biosynthesis pathway, including C4H, CHI, CHS, F3'H, FLS, DFR, UFGT, and so on. It is suggested that these VcF3Hs are involved in blueberry anthocyanin metabolism.
Promoter analysis results
-
The distribution of cis-acting elements of the VcF3Hs promoters was analyzed. Besides the common CAAT-box and TATA-box elements and some functional unknown elements, 39 kinds of elements (in amount of 334) are identified on their promoters. These elements can be further classified into four categories: light responsive, growth, and development-related, phytohormone responsive, and stress responsive (Fig. 3a). All VcF3Hs promoters contained light responsive G-box element, ABA-responsive related ABRE element, drought-inducibility related MYC element, defense and stress-related MYB element, high-temperature responsive STRE elements, and endosperm expression related GCN4_motif elements.
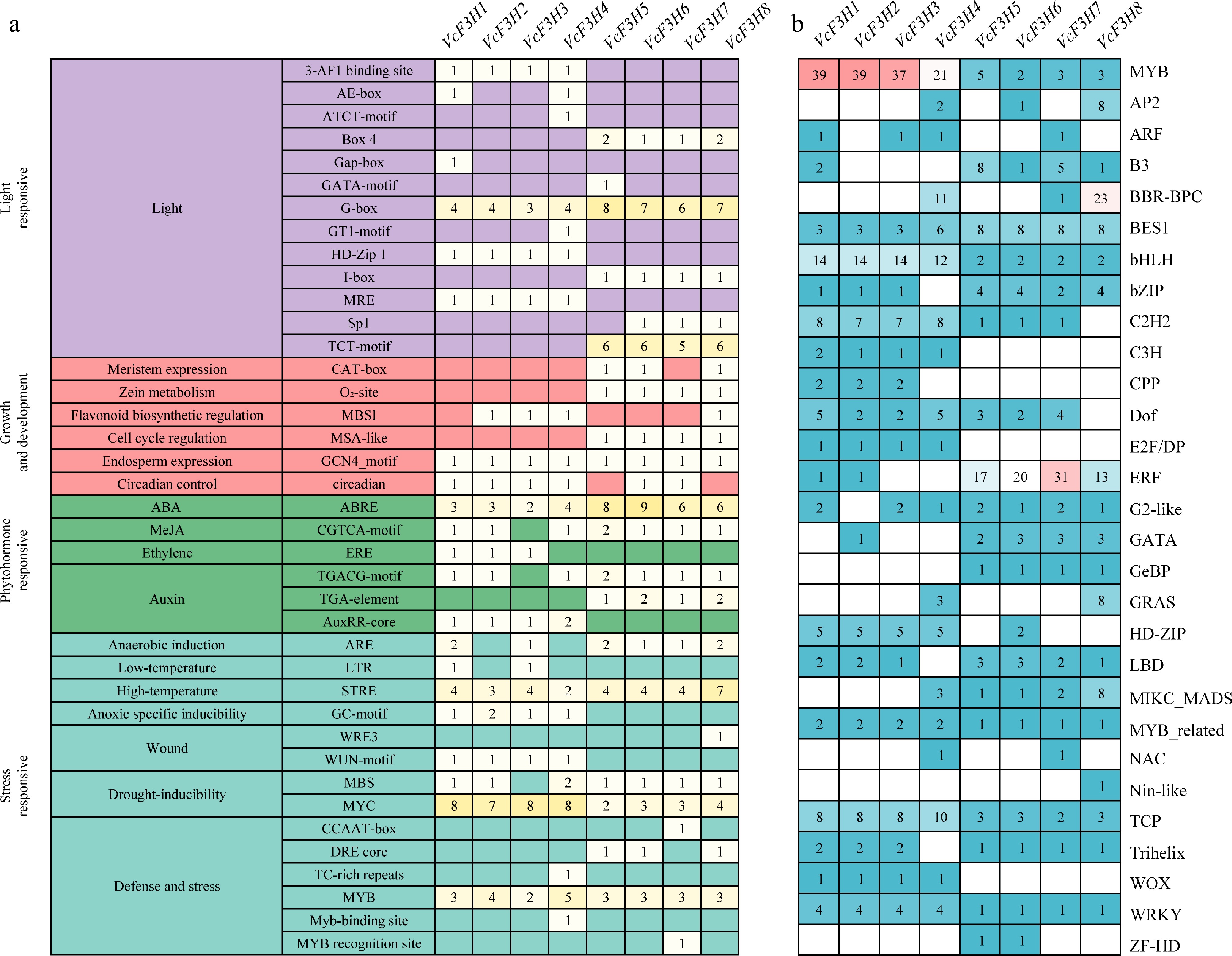
Figure 3.
Predicted (a) cis-acting elements, and (b) transcription factor binding sites in promoters of VcF3H.
We also identified binding sites for TFs from 29 families (Fig. 3b), such as MYB, bHLH, ERF, TCP, and NAC, in promoters of anthocyanin-related VcF3Hs. Among them, binding sites for MYB accounted for the largest (149), followed by ERF (83), and bHLH (62). The number of TFBSs on VcF3H1 promoter was the highest (105), and the number of binding sites on VcF3H6 promoter was the lowest (59). There are MYB, bHLH, MYB_related, BES1, WRKY, and TCP binding sites on all VcF3Hs promoters. It is worth noting that each VcF3H promoter contains at least two binding sites for MYB, bHLH, BES1, and TCP. In addition, the MYB and bHLH binding sites amounts on VcF3H1~4 promoters were much higher than that on VcF3H5~8 promoters.
Gene expression analysis results
-
According to our blueberry fruit transcriptome data, the expression patterns of anthocyanin-related VcF3Hs in fruits at five ripening stages were first studied (Fig. 4a). Results showed that VcF3H1~4 all exhibited a 'fall-rise' expression pattern during fruit ripening, and their expression levels all peaked in purple fruits (PF). The expression of VcF3H7~8 increased as fruit ripened. However, the VcF3H5~6 expression levels in fruits at different ripening stages followed the order: green fruit (GF) < pink fruit (PiF) < PF < red fruit (RF) < blue fruit (BF). The expression of VcF3H2, VcF3H6, and VcF3H8 were further verified by qRT-PCR (Fig. 4b). Results showed that the expression pattern of VcF3H2 was consistent with our transcriptome results. Its expression level in PiF, RF, PF, and BF was about 5.93-, 33.58-, 38.96-, and 20.00-fold of GF, respectively. Although the qRT-PCR results for VcF3H6 and VcF3H8 were not the same as our transcriptome data. Their relative expression levels in PiF, RF, PF, and BF were significantly higher than those in GF.
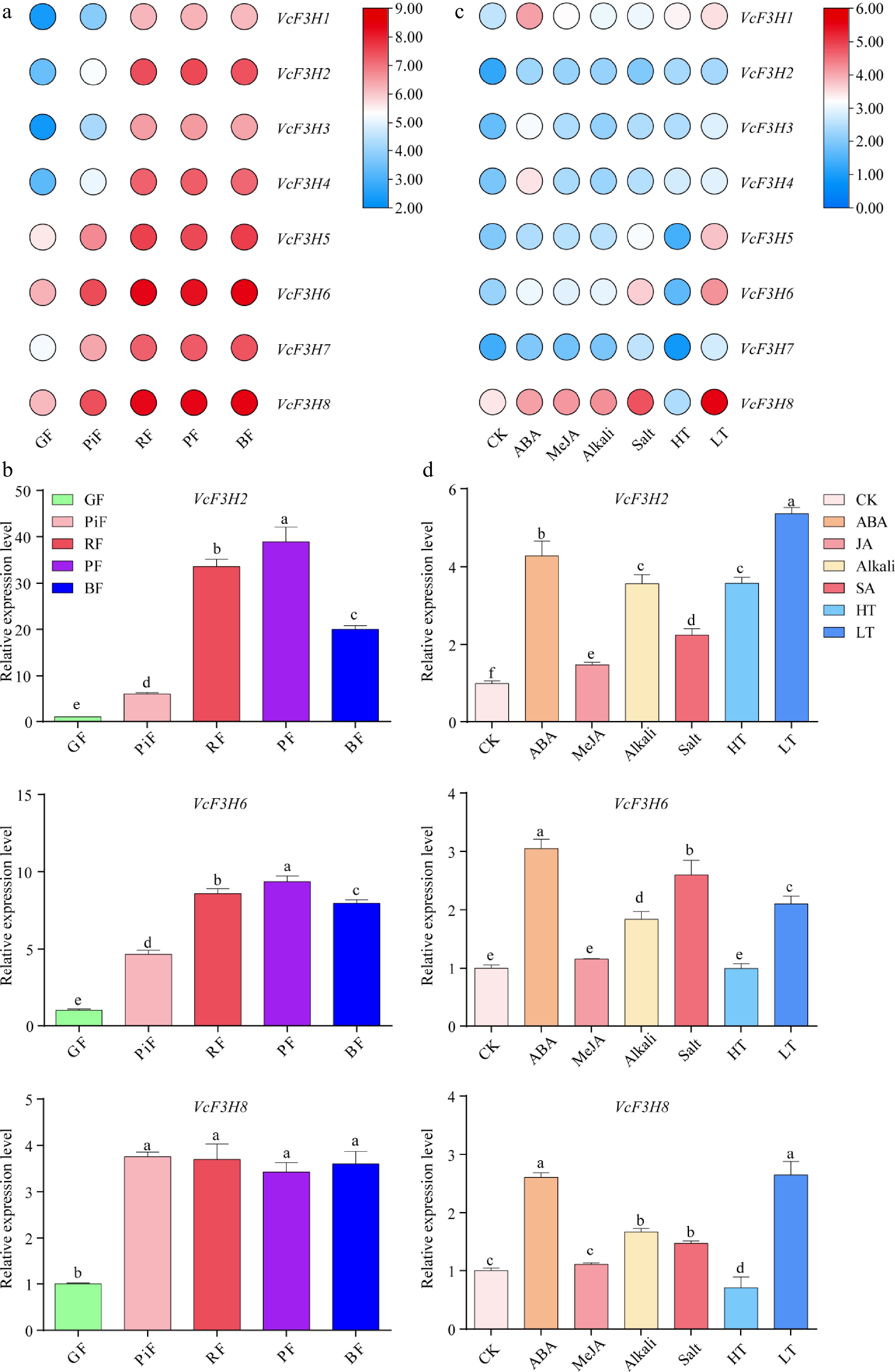
Figure 4.
Gene expression analysis results of VcF3Hs in (a), (b) fruits at five different ripening stages, and (c), (d) in leaves treated with different phytohormones and abiotic stresses. (a) and (c) are transcriptome data analysis results. The Log2 (FPKM + 1) value is used for heatmap drawing. The redder the color, the higher the gene expression, and the bluer the color, the lower the gene expression. (b) and (d) are qRT-PCR analysis results. Different letters above columns represent a significant difference at the p < 0.05 level. GF: green fruit; PiF: pink fruit; RF: red fruit; PF: purple fruit; BF: blue fruit; CK: nontreated control leaf; ABA, MeJA, Alkali, Salt, HT, and LT represents leaf treated with 100 mM ABA, 100 mM MeJA, 200 mM NaCl, and Na2SO4, 200 mM NaHCO3 and Na2CO3, 4 °C low temperature and 40 °C high temperature, respectively.
There are many stress- and phytohormone-responsive elements in promoters of anthocyanin-related VcF3Hs, suggesting that their expression may be significantly affected by various stresses and phytohormone treatments. Consistently, our leaf transcriptome data showed that ABA, MeJA, alkali, salt, and LT treatments significantly upregulated the expression of all anthocyanin-related VcF3Hs (Fig. 4c). Interestingly, HT treatment significantly upregulated the expression of VcF3H1~4 but downregulated the expression of VcF3H5~8. The expression of VcF3H2 was significantly upregulated by ABA, MeJA, alkali, salt, HT, and LT (Fig. 4d), accounting for 4.28-, 1.48-, 3.57-, 2.24-, 3.58-, and 5.36-fold of CK, respectively. ABA, alkali, salt, and LT treatments respectively upregulated VcF3H6 expression by 3.05-, 1.84-, 2.60-, and 2.10-fold compared to CK, while HT and JA treatments did not significantly influence its expression. VcF3H8 expression was upregulated by 2.61-, 1.11-, 1.67-, 1.48-, and 2.65-fold by ABA, MeJA, alkali, salt, and LT treatment, respectively. However, HT downregulated its expression to approximately 71% of CK, and MeJA treatment slightly upregulated its expression.
Transient overexpression analysis of VcF3H2 in apple and blueberry fruits
-
VcF3H2 expressed highly in blueberry fruit and exhibited the most significant upregulation during fruit ripening compared to other anthocyanin-related VcF3Hs. This suggested that it might play a key role in anthocyanin biosynthesis in blueberry fruits during ripening. By using transient overexpression assays in apple and blueberry fruits, its functions were further studied (Fig. 5). Results showed that its overexpression resulted in obvious pigment accumulation in apple peels (Fig. 5a & b). The anthocyanin content in apple peels overexpressing VcF3H2 was found to be 1.53-fold of EV (Fig. 5c) QRT-PCR analysis showed that its overexpression significantly upregulated the expression levels of MdCHI, MdCHS, MdDFR, MdANS, and MdUFGT in apple peels (Fig. 5d), accounting for 2.65-, 1.40-, 1.61-, 1.83-, and 2.03-fold of EV, respectively.
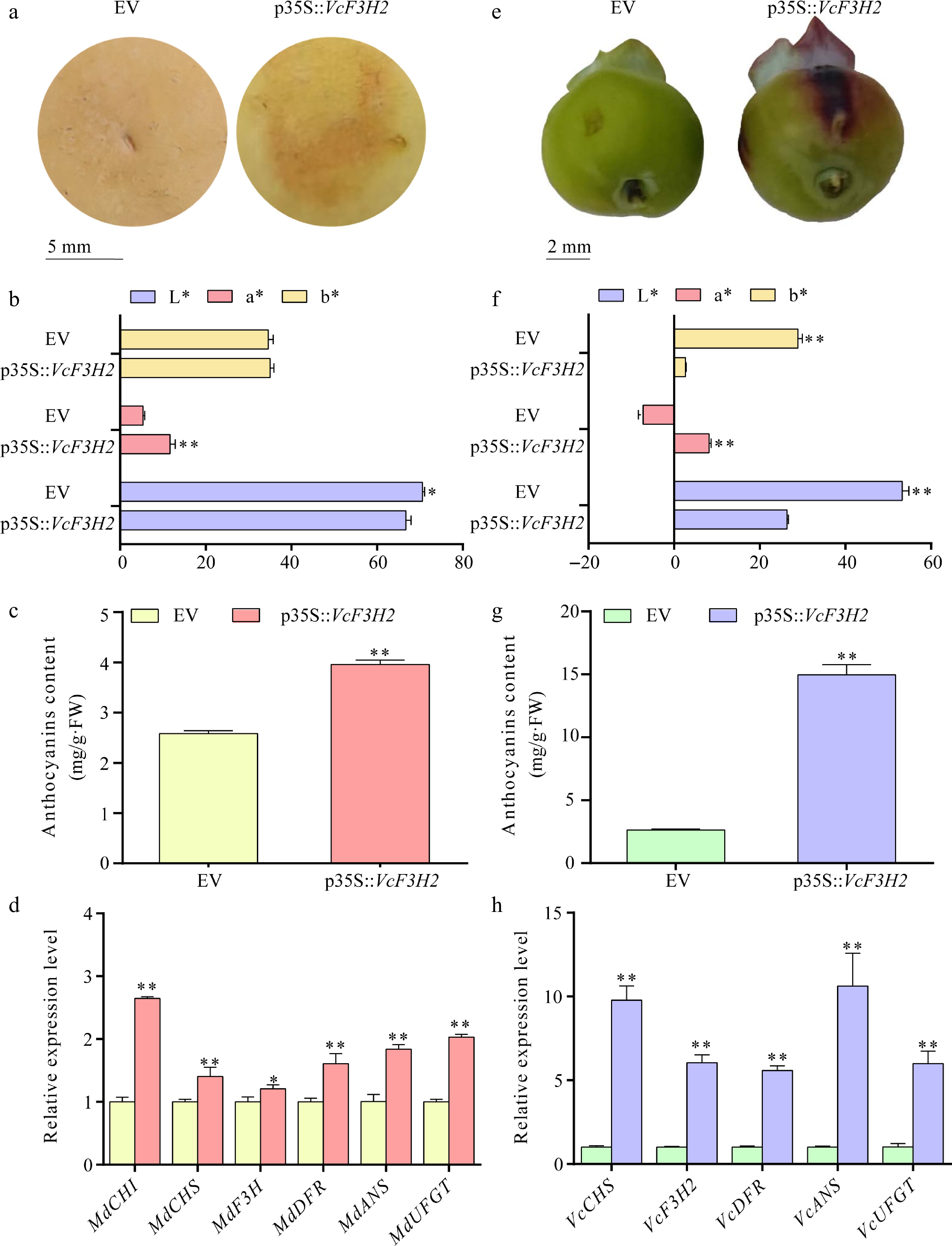
Figure 5.
The influences of VcF3H2 overexpression in anthocyanins biosynthesis and expression of corresponding structural genes in apple and blueberry fruits. (a) Transient overexpression of VcF3H2 promoted anthocyanins accumulation in apple peel; (b)−(d) Effects of overexpression of VcF3H2 on color parameters, anthocyanins content and expression of anthocyanins biosynthesis structural genes in apple peels. (e) Transient overexpression of VcF3H2 promoted anthocyanins accumulation in blueberry fruit peels; (f)−(h) Effects of overexpression of VcF3H2 on color parameters, anthocyanin content, and expression of anthocyanin biosynthesis structural genes in blueberry fruit peels. Data were presented as the means ± standard deviations of three biological replicates, with asterisks showing significant differences (*, p < 0.05; **, p < 0.01). In (b) and (f), L* indicates lightness from 0 (black) to 100 (white); a* indicates red/green (+a* = redder, −a = greener); b* indicates yellow/blue (+b* = yellower, −b* = bluer).
The overexpression of VcF3H2 also led to obvious pigmentation in blueberry fruit peels (Fig. 5e & f). The anthocyanin content in blueberry fruit peels overexpressing VcF3H2 was about 5.68-fold of EV (Fig. 5g), and the expression level of VcF3H2 accounted for 6.04-fold of EV (Fig. 5h). Its overexpression significantly upregulated the expressions of VcCHS, VcDFR, VcANS, and VcUFGT in blueberry peel, accounting for 9.78-, 5.58-, 5.99-, and 10.62-fold of EV (Fig. 5h), respectively.
The binding and activation effects of VcMYB-1 and VcAN1 on VcF3H2 promoter
-
Promoter analysis showed that all anthocyanin-related VcF3Hs contained G-box, Myb, and MYC elements, as well as the binding sites for MYB and bHLH TFs in their promoters. These indicated that the expression of VcF3Hs might be regulated by anthocyanin-regulatory MYBs and bHLHs. By using Y1H, we investigated the binding ability of VcAN1 and VcMYB-1 to the VcF3H2 promoter. Results showed that the growth of yeast carrying pVcF3H21960 was inhibited on SD/-Ura media containing 250 mM Aureobasidin A (AbA) (Fig. 6a), indicating that pVcF3H21960 exhibited no self-activation. Yeast co-transformed with pVcF3H21960 and pGADT7 could not grow on SD/-Leu medium containing 250 mM AbA (Fig. 6b), while strains co-transformed with pVcF3H21960 and pGADT7-VcAN1/VcMYB-1 grew well. These results indicate that both VcAN1 and VcMYB-1 can bind to the VcF3H2 promoter.
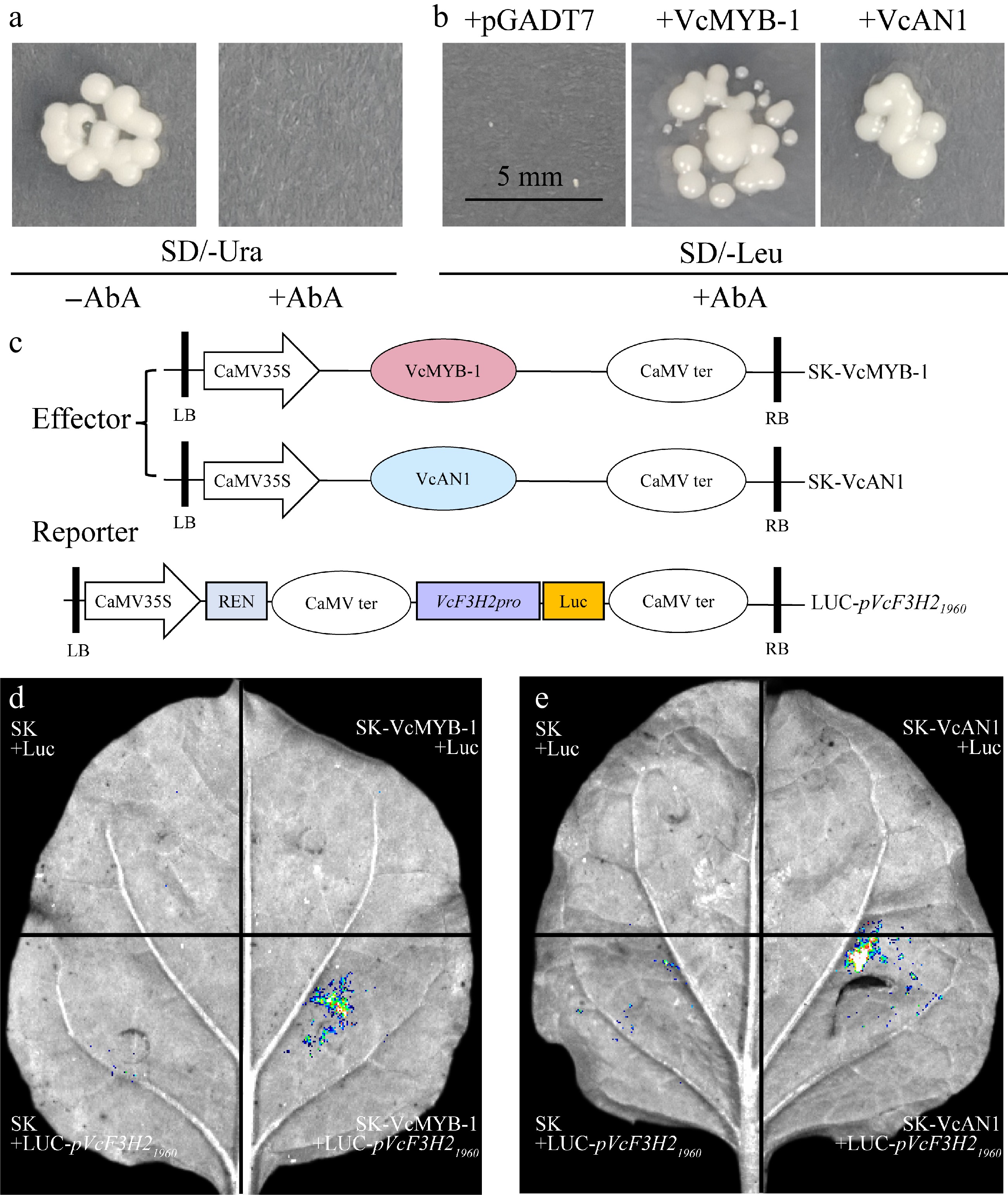
Figure 6.
Regulation of VcMYB-1 and VcAN1 on VcF3H2 promoter. (a) Self-activation verification of the VcF3H2 promoter. AbA: Aureobasidin A. (b) Y1H validation results. (c) Schematic diagrams for the effector and reporter constructs used for LUC; REN: Renilla luciferase. The stop codon removed CDS sequences of VcMYB-1 and VcAN1 were inserted into the pNC-Green-SK vector to generate effector constructs. The 1960 bp VcF3H2 promoter sequence was cloned into pNC-Green-Luc vector to generate the reporter construct (LUC-pVcF3H21960). LB: left border; RB: right border. (d) and (e) LUC imaging results for the binding and activation abilities of VcMYB-1 and VcAN1 on the VcF3H2 promoter, respectively.
To further investigate the binding ability of VcMYB-1 and VcAN1 to the VcF3H2 promoter, dual-luciferase assays (LUC) were performed (Fig. 6c). Both SK-MYB-1 and SK-AN1 were found to significantly enhanced the fluorescence of LUC-pVcF3H21960 in tobacco leaves (Fig. 6d & e), indicating that both VcMYB-1 and VcAN1 can promote anthocyanins accumulation by directly binding to and activating the promoter of VcF3H2.
-
F3H, catalyzing naringenin into dihydroflavonols, intermediates in the biosynthesis of both flavonols and anthocyanidins, is a key enzyme of the plant flavonoids/anthocyanins biosynthesis process. In this study, we identified eight anthocyanin-related F3Hs from blueberry. All the identified VcF3Hs shared more than 80% similarity with several reported flavonoid/anthocyanin-related F3Hs from other plants, such as AtF3H[25], OsF3H[26], VvF3H[9], PeF3H[29], GmF3H[27], and so on. This indicated that plant anthocyanin-related F3Hs were highly conserved. Several key enzymes in the flavonoid/anthocyanin biosynthesis pathway, including C4H, CHI, CHS, F3H, FLS, DFR, and UFGT, were predicted to be interacting proteins of these VcF3Hs, indicating again that they play important roles in blueberry flavonoid/anthocyanin biosynthesis. The protein sequences of VcF3H2~4 were the same, with only one amino acid difference from VcF3H1. Moreover, their expression patterns in fruits at different ripening stages were almost the same. The other four VcF3Hs shared similarity greater than 96.78% with each other, but only slightly over 90% with VcF3H1~4. All these eight VcF3Hs were highly expressed in fruits at late ripening stages, but VcF3H5~8 had different expression patterns from VcF3H1~4. Additionally, among the 12 duplicated gene pairs identified among the eight VcF3Hs, none were found between VcF3H1~4 members and VcF3H5~8 members. Collectively, it can be concluded that the eight anthocyanin-related VcF3Hs can be further categorized into two groups that play somewhat different roles in anthocyanin biosynthesis.
F3Hs play important roles in plant stress responses and their expression could be significantly affected by various stresses, and phytohormones[42,43]. The promoters of all anthocyanin-related VcF3Hs had ABA-responsive, drought-inducibility-related, defense, and stress related, and high-temperature responsive elements. Consistently, gene expression analysis revealed that ABA, alkali, salt, and LT, especially ABA, significantly upregulated the expression of almost all VcF3Hs. The upregulation of D. officinale F3H could increase anthocyanin and flavonol accumulation and confer plants with tolerance to salt and cold[12]. The overexpression of L. chinense F3H greatly enhanced the antioxidant ability of transgenic tobacco plants[13]. The ABA- and stress-inducible nature of these VcF3Hs may enhance the stress-resistance of blueberry in an anthocyanin-dependent way. Interestingly, high temperatures upregulated the expression of VcF3H1~4, but downregulated the expression of VcF3H5~8, suggesting that the two VcF3Hs groups play different roles in response to heat stress.
The upregulation or overexpression of F3H genes have been frequently reported to help in increasing anthocyanin accumulation[44]. Similarly, our study revealed that the transient overexpression of VcF3H2, the most differentially expressed VcF3H member during fruit ripening, resulted in significantly promoted anthocyanin accumulation and greatly upregulated expression of anthocyanin biosynthesis-related structural genes in apple and blueberry peels. Many studies have shown that TFs, such as MYB and bHLH, play important regulatory roles in anthocyanin biosynthesis by directly or indirectly influencing the expression of corresponding structural genes[45−47]. The rice MYC2 can activate the promoters of several anthocyanin biosynthesis genes, including CHI, CHS, F3H, F3'H, DFR, and 3GT[48]. Ginkgo biloba MYB can directly bind to the promoters of F3H and some other structural genes, and influence the flavonol biosynthesis and accumulation in leaves[49]. In our study, binding sites for MYB and bHLH TFs were identified on the promoters of all eight anthocyanin-related VcF3Hs. Notably, MYB and bHLH binding sites on the VcF3H2 promoter made up more than half of all TFBSs (accounting for 57.3%), suggesting that VcF3H2 may be regulated by these two kinds of TFs. Consistently, our Y1H and LUC assay results showed that both the anthocyanin regulatory factors VcMYB-1 and VcAN1 could bind to and activate the VcF3H2 promoter. Thus, the 'VcMYB-1/VcAN1-VcF3H2' module is concluded to play an important role in blueberry anthocyanins biosynthesis (Fig. 7).
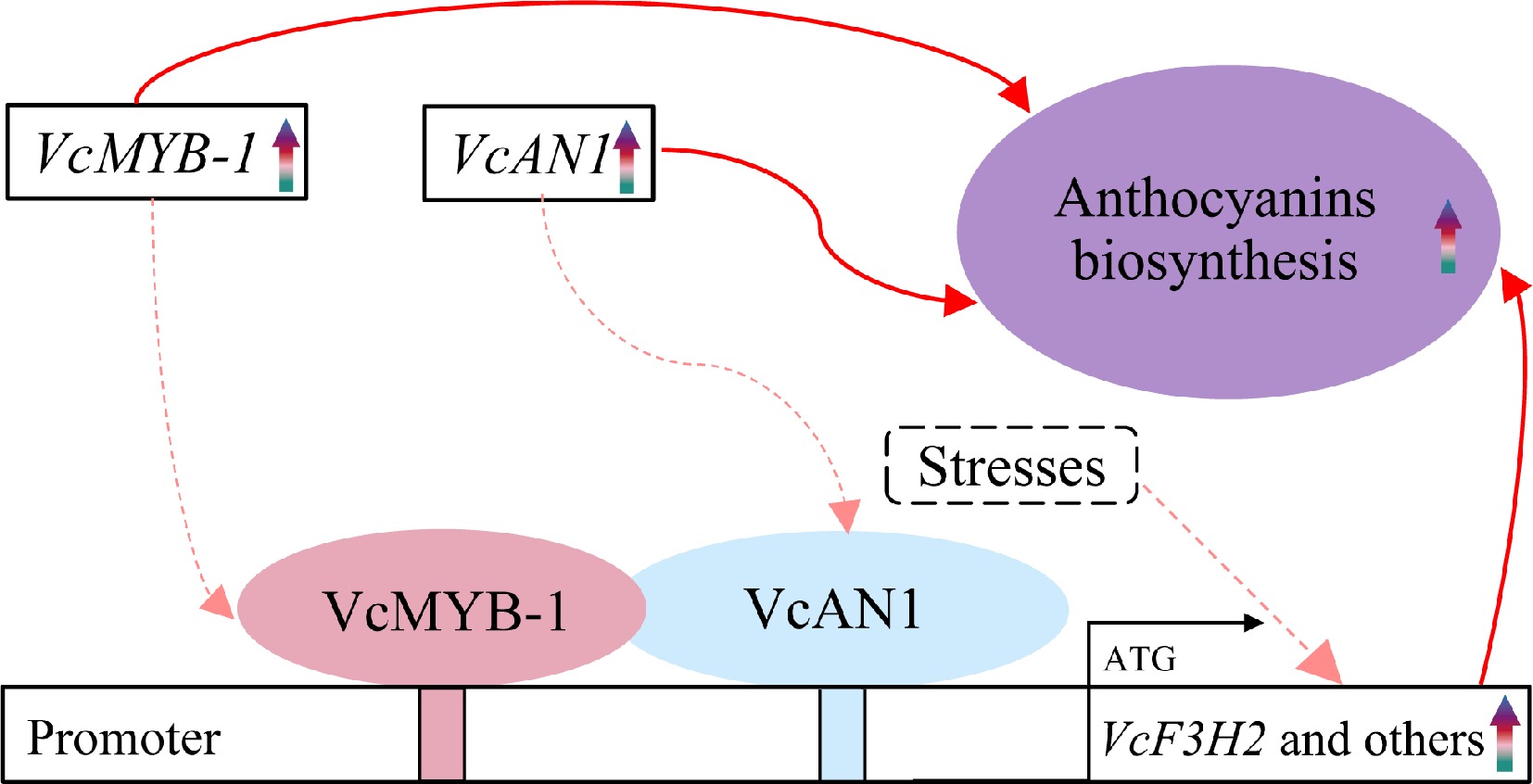
Figure 7.
The 'VcMYB/VcAN1-VcF3H2' module in regulating anthocyanin biosynthesis in blueberry. Rainbow arrows represent upregulation or increase during fruit ripening. Red arrows represent promoting effects. Solid arrows represent results have been validated in this study or previous studies, and dotted arrows represent results that need to be further confirmed.
In summary, we identified and characterized the eight anthocyanin-related F3H genes in blueberry, and demonstrated the contribution of VcF3H2 to anthocyanin biosynthesis. By using Y1H and LUC assays, we validated the binding activities of two anthocyanin regulatory TFs (VcMYB-1 and VcAN1) on the VcF3H2 promoter. These results will help elucidate the anthocyanin metabolism in blueberry. VcF3H2 promoter has many stress-responsive elements and the gene's expression could be significantly influenced by multiple abiotic stresses and phytohormones, implying VcF3H2 might be involved in stress-induced anthocyanin accumulation. Additionally, many binding sites for some other TFs were also identified in the VcF3H2 promoter. In the future, further research should be conducted to verify the roles of VcF3H2 in blueberry stress responses and to reveal the regulatory roles of other TFs on VcF3H2 and other anthocyanin biosynthesis-related structural genes.
This research was supported by the Natural Science Basic Research Program of Shanxi Province (202403021212072, 202203021211267), the PhD Introduction Research Start-up Project of Shanxi Agricultural University (2023BQ117), the Fund for High-level Talents of Shanxi Agricultural University (2021XG010) and the Reward Fund for PhDs and Postdoctors of Shanxi Province (SXBYKY2022004). We sincerely thank Dr. Pu Yan at ITBB, CATAS, for providing the SK and LUC vectors.
-
The authors confirm contribution to the paper as follows: study conception and design: Cheng C; data collection: Li R, Guo S, Hao S, Du S, Zhang J; analysis and interpretation of results: Li R, Guo S, Hao S, Du S, Liu R, Li J; technical assistance: Cheng C, Zhang Y; draft manuscript preparation: Li R, Cheng C. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Information for the primers used in this study.
- Supplementary Table S2 Physiochemical properties analysis results for the identified anthocyanin-related VcF3Hs.
- Supplementary Table S3 The identified duplicated gene pairs among the eight anthocyanin-related VcF3Hs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li R, Guo S, Hao S, Du S, Zhang J, et al. 2025. Characterization of the anthocyanin-related flavanone 3-hydroxylase (F3H) genes and functional analysis of VcF3H2 in blueberry. Fruit Research 5: e022 doi: 10.48130/frures-0025-0012 

Characterization of the anthocyanin-related flavanone 3-hydroxylase (F3H) genes and functional analysis of VcF3H2 in blueberry
- Received: 20 December 2024
- Revised: 10 March 2025
- Accepted: 21 March 2025
- Published online: 04 June 2025
Abstract: Flavanone 3-hydroxylase (F3H) is a key enzyme involved in plant flavonoid/anthocyanin biosynthesis pathways. In this study, we identified eight candidate anthocyanin-related F3H genes from blueberry (Vaccinium corymbosum), and characterized their sequences and expression patterns. These anthocyanin-related VcF3Hs are conserved in sequences and their encoding genes express highly in blueberry fruits at late ripening stages. By using apple and blueberry fruits transient overexpression analysis, the function of VcF3H2 was studied. Its overexpression significantly improved the anthocyanin accumulation and upregulated the expression levels of anthocyanin biosynthesis-related structural genes in fruit peels of both apple and blueberry. Numerous binding sites for MYB and bHLH transcription factors (TFs) were identified in promoters of VcF3Hs, especially VcF3H1~4. Consistently, our yeast one-hybrid and dual-luciferase assay results showed that both the anthocyanin-related VcMYB-1 and VcAN1 could bind to and activate the promoter of VcF3H2, indicating that its roles in anthocyanins biosynthesis were regulated by anthocyanin-related TFs. Our study characterized the anthocyanin-related F3Hs in blueberry and demonstrated the role of the 'VcMYB-1/VcAN1-VcF3H2' module in regulating blueberry anthocyanin biosynthesis.