









-
Aldehydes serve as intermediates in numerous metabolic pathways within organisms; however, excessive aldehyde accumulation can disrupt normal metabolic processes and potentially become toxic[1]. Aldehyde dehydrogenases (ALDHs) constitute a superfamily of NAD(P)+-dependent enzymes that catalyze the oxidation of diverse aliphatic and aromatic aldehydes to their corresponding carboxylic acids[2]. This process produces NADPH and NADH, which play critical roles in maintaining redox balance and aldehyde homeostasis[3]. These enzymes are frequently termed 'aldehyde scavengers' due to their role in detoxifying harmful aldehydes[4]. In addition, ALDHs are involved in various metabolic pathways such as amino acid metabolism, retinoic acid metabolism, and osmoprotectant synthesis.
ALDHs are ubiquitous across all taxonomic groups, including animals, plants, algae, and others, and exhibit high conservation throughout evolution. According to the nomenclature guidelines established by the Aldehyde Dehydrogenase Gene Nomenclature Committee (AGNC), proteins are classified into 24 distinct families[5]. In plants, the ALDH superfamily encompasses 14 families (2, 3, 5, 6, 7, 10, 11, 12, 18, 19, 21, 22, 23, and 24), with seven families (ALDH10, ALDH12, ALDH19, ALDH21, ALDH22, ALDH23, and ALDH24) being plant-specific. These genes are expressed in multiple tissues and respond to various stressors. Extensive research indicates that upregulated ALDH expression is a common feature of activated stress response pathways.
Most studied plant ALDH exhibit upregulated expression in response to various abiotic stresses, including high salinity, dehydration, elevated temperatures, waterlogging, oxidative stress, and heavy metal exposure[6−8]. Environmental challenges such as drought and high salinity trigger rapid reactive oxygen species (ROS) production, leading to toxic aldehyde accumulation and disrupted metabolic processes. ALDH enzymes detoxify these harmful aldehydes, thereby enhancing plant survival under stress. Recent studies suggest that ectopic overexpression of ALDH enhances plant tolerance to abiotic stresses. Beyond Arabidopsis thaliana, ALDH families have also been characterized in diverse plant species, including moss, algae[9], and economically important crops such as sorghum, soybean, and cotton[10−12]. However, ALDH remain unstudied in orchids.
Cymbidium, a large genus within the orchid family (Orchidaceae), exhibits diverse growth habits, including terrestrial, epiphytic, and saprophytic lifestyles[13]. For a long time, orchids have fascinated people with their exquisite floral morphology[14]. Orchids have a long history of cultivation and ornamental appreciation globally[15]. However, global climate change, including global warming and rising sea levels, poses significant challenges for terrestrial orchids, particularly in terms of soil drought and salinization[16,17]. Notwithstanding China's vast geographical coverage and rich germplasm diversity, the country faces increasingly severe environmental challenges. To address this, we focused on three historically significant Cymbidium species in China—C. sinense, C. ensifolium, and C. goeringii—and conducted whole-genome identification of ALDH genes to study their evolutionary relationships and stress-responsive expression patterns. We identified and classified ALDHs from these three species into ten families and six protein subfamilies according to AGNC standards[5]. Further analysis included genomic organization, sequence homology, and subcellular localization of the identified ALDH genes. We further analyzed ALDHs gene expression in C. sinense leaves under drought and salinity stress, and validated the subcellular localization of stress-responsive genes.
-
The whole-genome sequencing data for C. sinense, C. ensifolium, and C. goeringii were obtained from their respective genomic resources[18−20]. Protein sequences of the A. thaliana ALDH family were sourced from TAIR (
www.arabidopsis.org , accessed on August 10, 2024). For Oryza sativa, ALDH family protein sequences were retrieved from Phytozome (https://phytozome-next.jgi.doe.gov , accessed on August 12, 2024), and previous studies[21]. To conduct BLASTP comparisons using the ALDHs data from the genomes of C. sinense, C. ensifolium, and C. goeringii, we utilized TBtools (Version 2.4.1)[22]. The presence of the ALDH superfamily domain (Pfam00171) in the identified proteins was confirmed using HMMER (www.ebi.ac.uk/Tools/hmmer/search/phmmer , accessed on August 15, 2024). All identified ALDH family members were classified according to the criteria established by the AGNC[5]. The physicochemical properties of the proteins were analyzed using the parameter calculation function in TBtools (Version 2.4.1)[22]. Subcellular localization predictions were performed using Cell-PLoc (www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2 , accessed on November 17, 2024).Phylogenetic analysis and multiple sequence alignment
-
Phylogenetic trees for ALDH genes from C. sinense, C. ensifolium, C. goeringii, A. thaliana, and O. sativa were constructed using Phylosuite (version 1.2.3) with the Maximum Likelihood algorithm and 1,000 bootstrap replicates[23]. The resulting phylogenetic tree was visualized using iTOL (
https://itol.embl.de/personal_page.cgi , accessed on August 20, 2024). Multiple sequence alignments of CsALDHs, CeALDHs, and CgALDHs were conducted using the MAFFT function in Phylosuite. ESPript 3.0 (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi , accessed on August 20, 2024) was used with default parameters to visualize amino acid sequences. Conserved motifs were analyzed using MEME (version 5.5.7,https://meme-suite.org/meme/tools/meme , accessed on August 15, 2024). The NCBI's Conserved Domain Database was utilized to examine conserved domain structures (www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi , accessed on August 15, 2024). TBtools was then used to visualize the results obtained from both MEME and NCBI analyses.Analysis of cis-acting elements and gene structure
-
The upstream 2000 bp regions of the CsALDH, CeALDH, and CgALDH gene coding sequences were captured using TBtools. The PlantPAN (version 4.0,
http://plantpan.itps.ncku.edu.tw/plantpan4/index.html , accessed on October 4, 2024) was utilized to predict and analyze the promoter regions within these 2,000-bp upstream sequences[24]. The numbers of exons, introns, coding sequences (CDSs) and untranslated regions (UTRs) were calculated using GXF stat. Data collection, analysis, and visualization were all performed using Microsoft Excel.Chromosome localization and collinearity analysis
-
TBtools was employed to map ALDH genes onto the chromosomes of C. sinense, C. ensifolium, and C. goeringii. One-Step MCScanX was utilized for collinearity analysis. Following pairwise collinearity analysis among the three species and A. thaliana, FileMerge MCScanX was used to integrate the results. Finally, Dual Synteny Plot was applied for visual presentation and refinement.
Protein-protein interaction (PPI) network, and microRNA binding site prediction
-
All CsALDH protein sequences were uploaded to the STRING database (
https://cn.string-db.org , accessed on September 27, 2024) to predict protein interactions. The resulting interaction data from STRING were imported into Cytoscape, where the Betweenness Centrality (BC) method was used to identify highly interacting proteins. The protein interaction network was then optimized and visualized. Additionally, psRNATarget (www.zhaolab.org/psRNATarget , accessed on November 16, 2024) was used to predict miRNA binding sites[25], with default parameters.Material processing and transcriptome data analysis
-
Healthy C. sinense 'Jinhuashan' plants, cultivated in the nursery of Fujian Agriculture and Forestry University (Fujian, China), were selected and acclimated in an artificial climate chamber for 10 d under the following conditions: photoperiod of 12 h light/12 h dark, light intensity of 8,000 lux, day/night temperature of 28/25 °C, and humidity of 75%. For drought stress treatment, roots of intact plants were immersed in 20% PEG6000 solution for 15 min and then removed[26]. For high salt stress treatment, roots of intact plants were immersed in 300 mmol/L NaCl solution for 15 min and then removed[27]. The first mature leaf from plants was sampled at five time points: 0 h (control, untreated), 6, 12, 24, and 48 h post-treatment. Each time point included three biological replicates (independent plants). Transcriptome data processing followed the method of Yang et al.[18] (NCBI accession: SRR15174943−SRR15174947). Raw reads were quality-filtered using fastp[28], and TPM (Transcripts Per Million) values were calculated with Kallisto (version 0.48.0) with parameters set as: quant -I -b 100. Data were visualized using TBtools.
Fluorescence quantitative test and data analysis
-
Total RNA was isolated from leaves at various time points after drought and salt stress treatments using FastPure Plant Total RNA Isolation Kit (Vazyme, Nanjing, China; Cat. No. RC401-01). Hifair III One-Step RT-qPCR SuperMix with gDNA Digestion (Yeasen, Shanghai, China; Cat. No. 11151ES) was employed to synthesize cDNA. The Actin gene (Mol022529) was chosen as the internal control for its stability[29]. Primers for the Actin gene and 14 candidate genes were designed using Primer Premier 5.0.
Real-time PCR was conducted on the Applied Biosystems QuantStudio system using cDNA labeled with SYBR Green fluorescent dye (Yeasen, Shanghai, China; Cat. No. 10222ES60). The thermal cycling protocol was as follows: an initial denaturation at 95 °C for 1 min, followed by 40 cycles of 95 °C for 10 s and 60 °C for 20 s, with fluorescence signal acquisition during the annealing/extension phase. Dissociation curves were generated using the instrument's default settings, and CT values were recorded for each sample. Gene expression levels were normalized to the Actin internal control gene (Mol022529), and relative expression was calculated using the 2−ΔΔCᴛ method[30]. Expression level correlation analysis was conducted using OmicStudio (
www.omicstudio.cn/home , accessed on September 25, 2024)[31].Cloning and subcellular localization of CsALDH5F1 and CsALDH11A3
-
We selected cDNA samples exhibiting the highest expression levels of target genes in qRT-PCR as templates for cloning. The templates used for cloning CsALDH5F1 and CsALDH11A3 were cDNAs collected at 24 and 48 h after salt stress treatment, respectively. The Super1300 vector was linearized using XbaI and KpnI restriction enzymes. Primers with homologous arms were designed based on the coding sequences (excluding their terminators) of CsALDH5F1 and CsALDH11A3, with reference to the Super1300 vector. Target gene fragments were amplified using homology-arm-containing primers, gel-purified, and ligated with the linearized Super1300 vector (Vazyme, Nanjing, China; Cat. No. DC301-01). The recombinant plasmids were then transformed into Escherichia coli for propagation. Positive clones were selected for plasmid amplification, followed by plasmid extraction (Vazyme, Nanjing, China; Cat. No. DC201-01) and subsequent transformation into Agrobacterium. The working suspension was prepared in deionized water with final concentrations of 1 M MgCl2, 1 M MES buffer, and 20 mg/mL Acetosyringone. The bacterial suspension was adjusted to an OD600 value of 0.8−1.0. Bacterial suspension was injected into interveinal regions of the abaxial surface in newly matured leaves of tobacco (Nicotiana benthamiana). Following injection, plants were maintained in darkness for 24 h and subsequently cultured under normal conditions for 48 h. GFP fluorescence signals on tobacco leaves were observed using confocal laser microscopy (Leica STELLARIS) in complete darkness.
-
We identified 14, 17, and 21 ALDH genes in C. sinense, C. ensifolium, and C. goeringii, respectively. To elucidate their evolutionary relationships and functional correlations within the ALDH gene family, we constructed a phylogenetic tree using ALDH sequences from these three orchids, A. thaliana, and O. sativa. An unrooted phylogenetic tree was generated based on CeALDH, CsALDH, CgALDH, AtALDH, and OsALDH sequences, followed by cluster analysis.
As shown in Fig. 1a, according to the AGNC guidelines, protein sequences showing > 40% similarity to known ALDH sequences were classified into the same family, while sequences with > 60% similarity were assigned to the same subfamily. Sequences showing < 60% similarity (i.e., > 40% divergence) to known ALDH sequences were classified as new gene families. Based on this classification, all identified ALDH family members in the three orchids were grouped into ten main families (Families 2, 3, 5, 6, 7, 10, 11, 12, 18, and 22) and six subfamilies. Subfamilies A, B, C, F, and H correspond to those previously identified in A. thaliana and O. sativa, while subfamily L, newly identified in this study, exhibits 40%−60% amino acid sequence similarity to other known ALDH subfamilies. Among the main ALDH families, ALDH3 contained the most members across the three orchid species, followed by ALDH2. Among subfamilies, the newly identified subfamily L has the most members, followed by subfamily B.
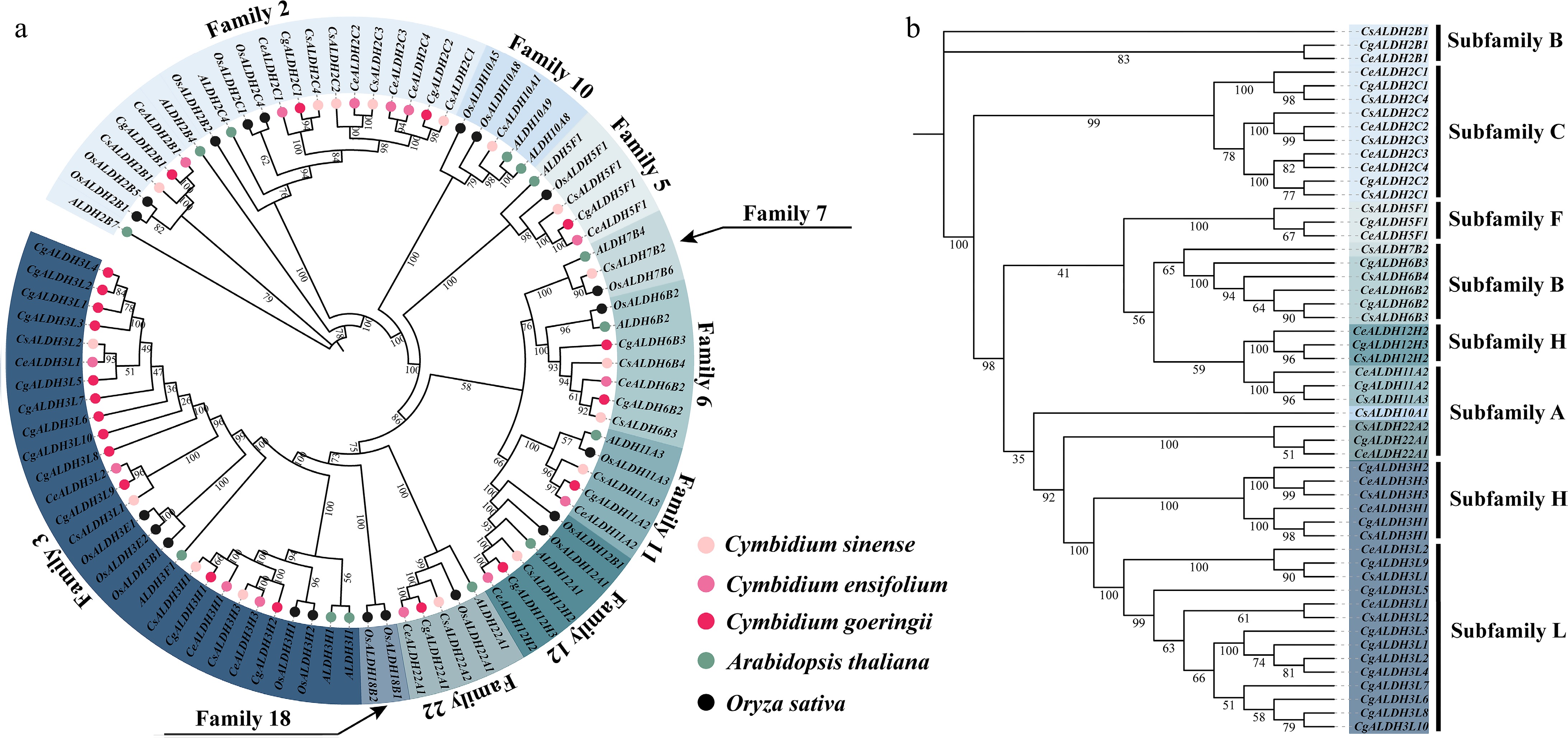
Figure 1.
Phylogenetic analysis of ALDH family members. (a) Phylogenetic tree constructed using ALDH family members from C. sinense, C. ensifolium, C. goeringii, A. thaliana, and O. sativa. (b) In the phylogenetic tree derived from C. sinense, C. ensifolium, and C. goeringii, the text on the right indicates the classification of protein subfamilies. These subfamilies are categorized into seven groups (A, B, C, F, H, and L), with subfamily L being a newly identified group in this study.
The physicochemical properties of the identified ALDH proteins in C. sinense, C. ensifolium, and C. goeringii are summarized as follows. Protein lengths ranged from 117 to 653 amino acids (aa), isoelectric points (pI) ranged between 4.71 and 9.75, and molecular weights (MW) ranged from 13.29 to 71.19 kDa. Additionally, subcellular localization prediction indicated that cytoplasm was the primary localization site for most ALDH proteins, followed by mitochondria, chloroplasts, peroxisomes, and the endoplasmic reticulum (Supplementary Table S1).
Multiple sequence alignment and phylogenetic analysis of ALDH proteins
-
We conducted a conservation analysis on the identified members of the ALDH family. During the identification process, we observed that some genes were relatively short in length. However, searches in the Pfam and NCBI Conserved Domain Databases revealed that all identified ALDH members possessed the ALDH or ALDH-SF superfamily domain (PF00171) (Fig. 2a). Therefore, we concluded that these shorter genes likely represent valid ALDH family members despite potential errors in genome annotation, and thus retained them for further analysis.
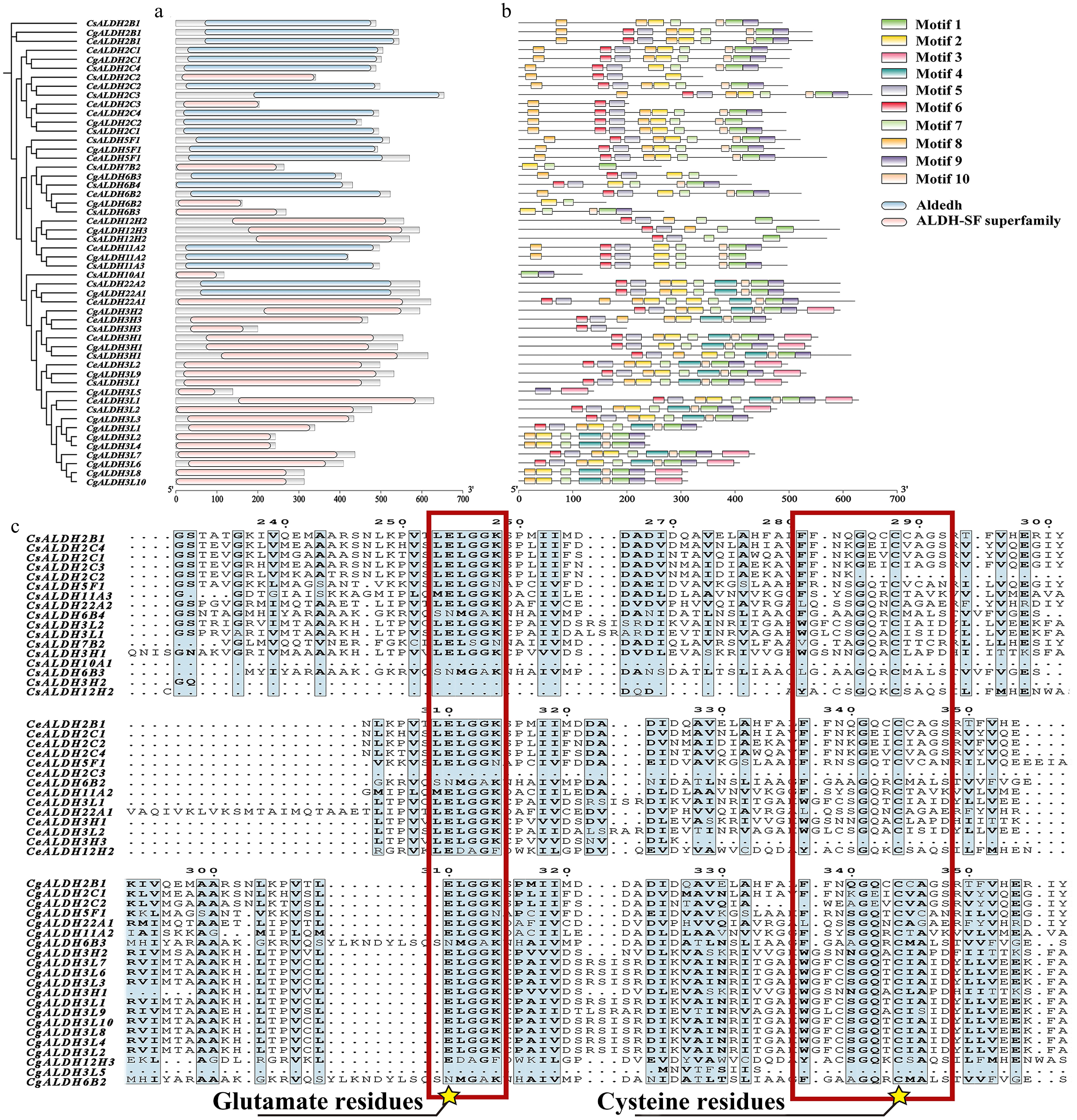
Figure 2.
Conserved domain analysis and multiple sequence alignment of the CsALDH, CeALDH, and CgALDH gene families. (a) Conserved domains, where blue represents the Aldedh domain (PF00171) and pink represents the ALDH-SF superfamily domain (PF00171). (b) Motifs of proteins, with different colors representing distinct motifs numbered from 1 to 10. The scale at the bottom allows for estimation of protein lengths. (c) Multiple sequence alignment of all identified proteins. Conserved motifs and active sites of glutamic acid residues and conserved motifs and active sites of cysteine residues are marked with red squares and yellow star symbols.
Motif analysis (Fig. 2b) indicated no significant differences in motifs across various families. Notably, most members of the ALDH3 family exhibited a unique motif 3, while most members of the ALDH3 and ALDH22 families possessed motif 4, characteristics not observed in other families.
ScanProsite and multiple sequence alignment analyses (Supplementary Table S2; Fig. 2c) revealed that most ALDH proteins contained both glutamic acid (PS00687) and cysteine (PS00070) active sites. Among the three orchid species, all members of the ALDH3 family lacked both PS00687 and PS00070. In C. sinense, 41.18% genes (7/17) contained both active sites; in C. ensifolium, 50.00% genes (7/14) contained both active sites; and in C. goeringii, 23.81% genes (5/21) contained both active sites.
Analysis of cis-elements and gene structure
-
Analysis of the 2,000-bp upstream region of the CsALDH, CeALDH, and CgALDH gene coding sequences (CDS) revealed numerous transcription factor binding sites. Specifically, C. sinense, C. ensifolium, and C. goeringii were found to contain 47, 47, and 43 types of transcription factor binding sites, respectively, with the number of sites ranging from 29 (e.g., CsALDH3L2, CsALDH6B4, CeALDH2C3, CeALDH2C4) to 37 (CsALDH3H3, CsALDH10A1, CgALDH3L9) (Fig. 3).
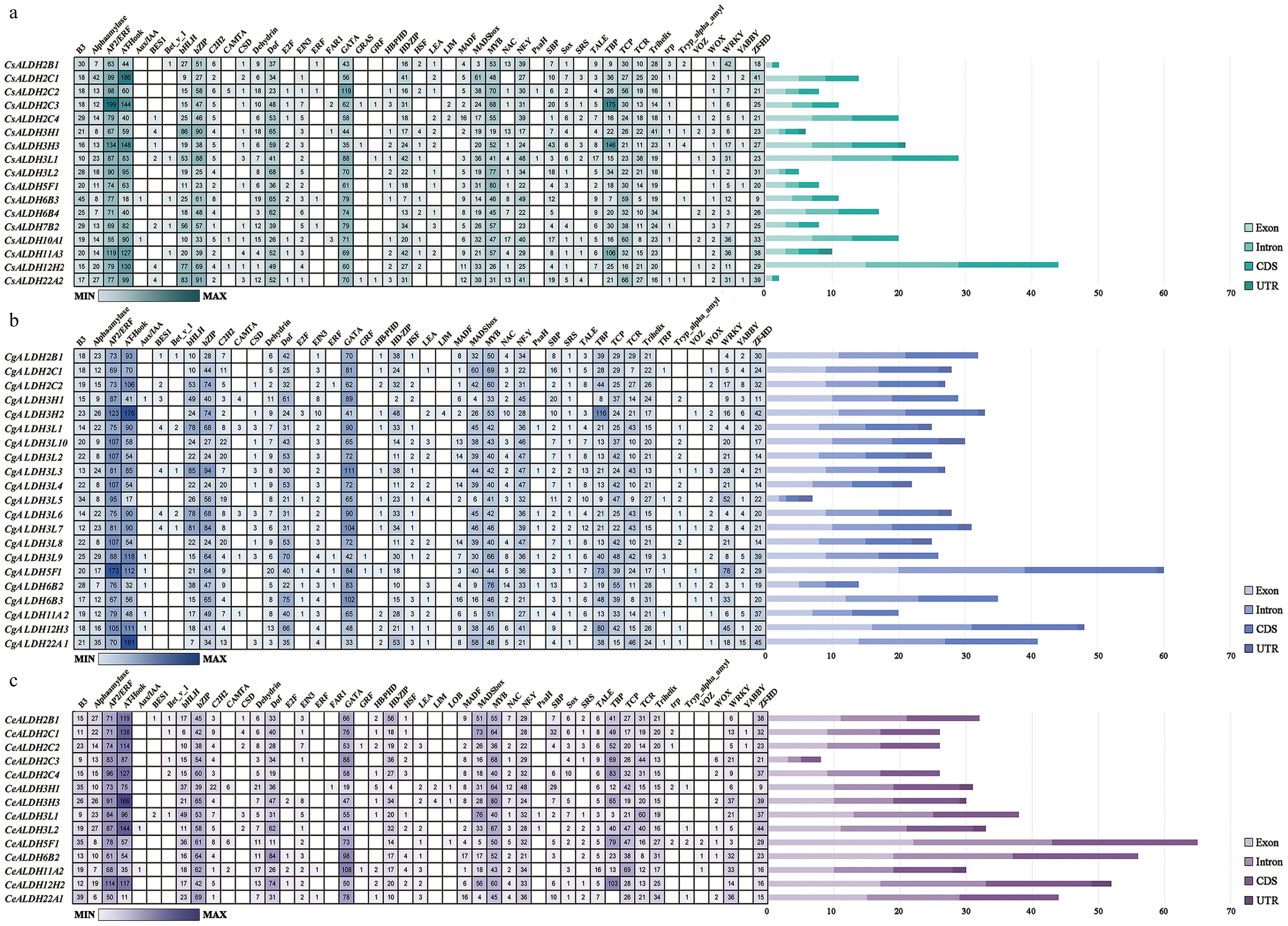
Figure 3.
Cis-element prediction and gene structure analysis. The gene structure of each ALDH family member is visualized, depicting the counts of exons, introns, CDSs, and UTRs. Distinct colors are used to differentiate these structural components. (a) C. sinense. (b) C. goeringii. (c) C. ensifolium. The quantity can be estimated based on the scale at the bottom.
We further analyzed the regulatory functions associated with these transcription factors and found that ALDHs can respond to various plant hormones and abiotic stress signals. For instance, plant hormone-responsive transcription factors (e.g., Aux/IAA, BES1, and EIN3), cold stress-responsive factors (e.g., AP2/ERF and HSF), heat stress-responsive factors (e.g., AT-Hook), drought stress-responsive factors (e.g., NF-Y and LEA), and salt stress-responsive factors (e.g., SRS and C2H2).
Furthermore, we examined the structure and quantity of introns, exons, CDSs, and UTRs (Fig. 3). Most members contain at least one exon and intron, except for CsALDH22A2 and CsALDH2B1, which lack introns. Most members of the CsALDH and CeALDH families do not contain UTRs. Among the CgALDHs, 33.3% members (7/21) lack UTRs, while the remaining members have one to four UTRs.
Chromosomal localization and collinearity analysis of ALDH in three Cymbidium orchids
-
The chromosomal locations of ALDH genes in the three Cymbidium species are shown in Supplementary Fig. S1. The distribution of ALDH genes on the chromosomes of C. sinense and C. ensifolium was relatively uniform, whereas the distribution in C. goeringii is more concentrated. Specifically, both C. sinense and C. goeringii exhibited a similar distribution pattern, with a maximum of three ALDHs per chromosome (e.g., Chr08 and Chr17 in C. sinense; Chr09 and Chr14 in C. ensifolium). In contrast, in C. goeringii, ALDH genes are predominantly located on chromosome Chr07, with no more than three ALDHs present on any single chromosome.
We conducted synteny analysis of ALDHs among the three orchid species and A. thaliana (Fig. 4). The results demonstrated that the three orchids exhibited stronger syntenic relationships among themselves than with the model plant A. thaliana. Through homology analysis of ALDHs across the four species, we identified 13 homologous gene pairs between C. sinense and C. goeringii, 10 between C. goeringii and C. ensifolium, but only one between A. thaliana and C. ensifolium. These findings indicate that ALDHs were more conserved within orchids than between orchids and A. thaliana.
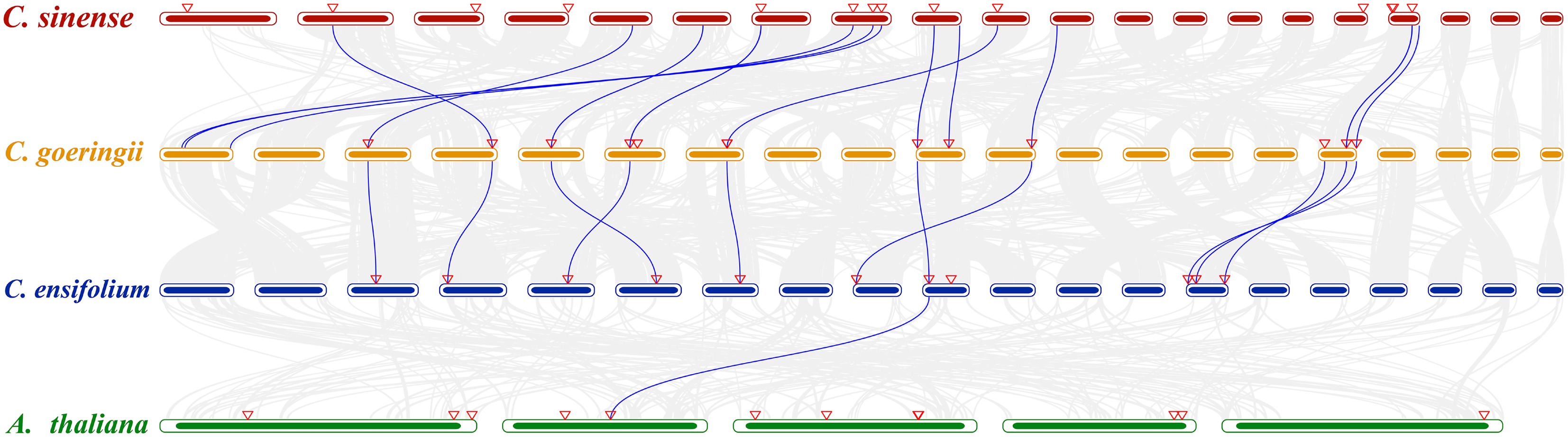
Figure 4.
Collinearity analysis between C. sinense, C. goeringii, C. ensifolium, and A. thaliana. The chromosomes of orchids are arranged from left to right as Chr01 to Chr20. The chromosomes of A. thaliana are arranged from left to right as Chr01 to Chr05. The gray lines in the background indicate collinear blocks within the three genomes. The blue lines highlight syntenic ALDH pairs. Red triangles indicate ALDH localization.
Protein-protein interaction (PPI) network analysis
-
We constructed a PPI regulatory network based on the 17 identified CsALDH proteins (Fig. 5a). Analysis of the proteins interacting with CsALDHs revealed that the majority were associated with abiotic stress responses. The most strongly interacting protein was POP (Polyamine Oxidase). Cytochrome P450 (CYP), polyamine oxidase (PAO), and Alanine-Glyoxylate Transaminase (AGT) also exhibited strong interactions with CsALDH proteins.
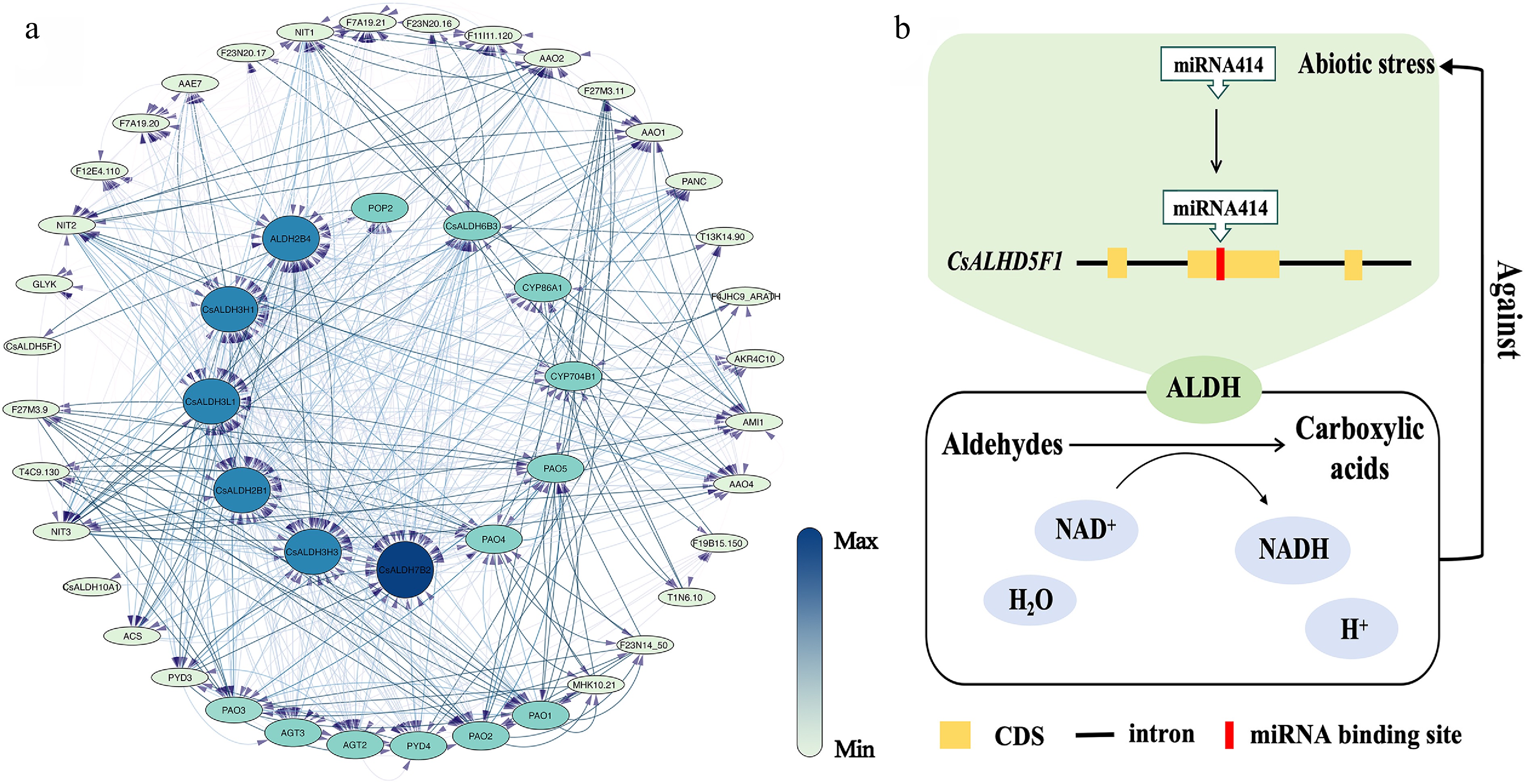
Figure 5.
(a) The PPI network of CsALDH. Each ellipse represents a protein, and lines connect pairs of interacting proteins. The strength of the interactions can be estimated using the color bar on the right. (b) Response pattern diagram of CsALDH5F1 under abiotic stress.
MicroRNA binding sites and stress response mechanism prediction
-
MicroRNAs (miRNAs) were widely present in eukaryotic organisms as endogenous non-coding single-stranded RNA molecules approximately 20−24 nucleotides in length. By analyzing the CsALDH sequences using psRNATarget (Supplementary Table S3), we found that CsALDH3H3 had the highest number of miRNA target sites (30). Among the predicted miRNAs, previous studies had validated that miR414 responded to abiotic stresses in plants, particularly salt stress[32]. We speculate that miR414 may inhibit the translation of its target genes post-transcriptionally, thereby reducing their protein level. This mechanism could enhance the conversion of toxic aldehydes into carboxylic acids as a salt stress response (Fig. 5b).
Expression patterns by transcriptome data and qRT-PCR analysis of CsALDHs
-
As shown in Fig. 6a, a comparative analysis of Transcripts Per Million (TPM) values was conducted across different tissues (Supplementary Table S4). The transcript abundance of each unigene in the C. sinense floral transcriptome was represented by log2(TPM) values. The expression levels of CsALDHs in the root were generally significantly lower than in other tissues. Notably, CsALDH2B1, CsALDH6B3, and CsALDH7B2 showed no expression in roots, stems, leaves, flowers, or fruits. Additionally, CsALDH2C1 was not expressed in the root. In the stem, CsALDH3L1, CsALDH10A1, and CsALDH1A3 showed prominent expression. In leaves, most genes were highly expressed, particularly CsALDH10A1, which had a TPM value as high as 153.32. In flower, the overall expression of CsALDHs was lower than in stem, leaf, and fruit but slightly higher than in root. In fruit, CsALDH3L1 and CsALDH10A1 exhibited high expression levels.
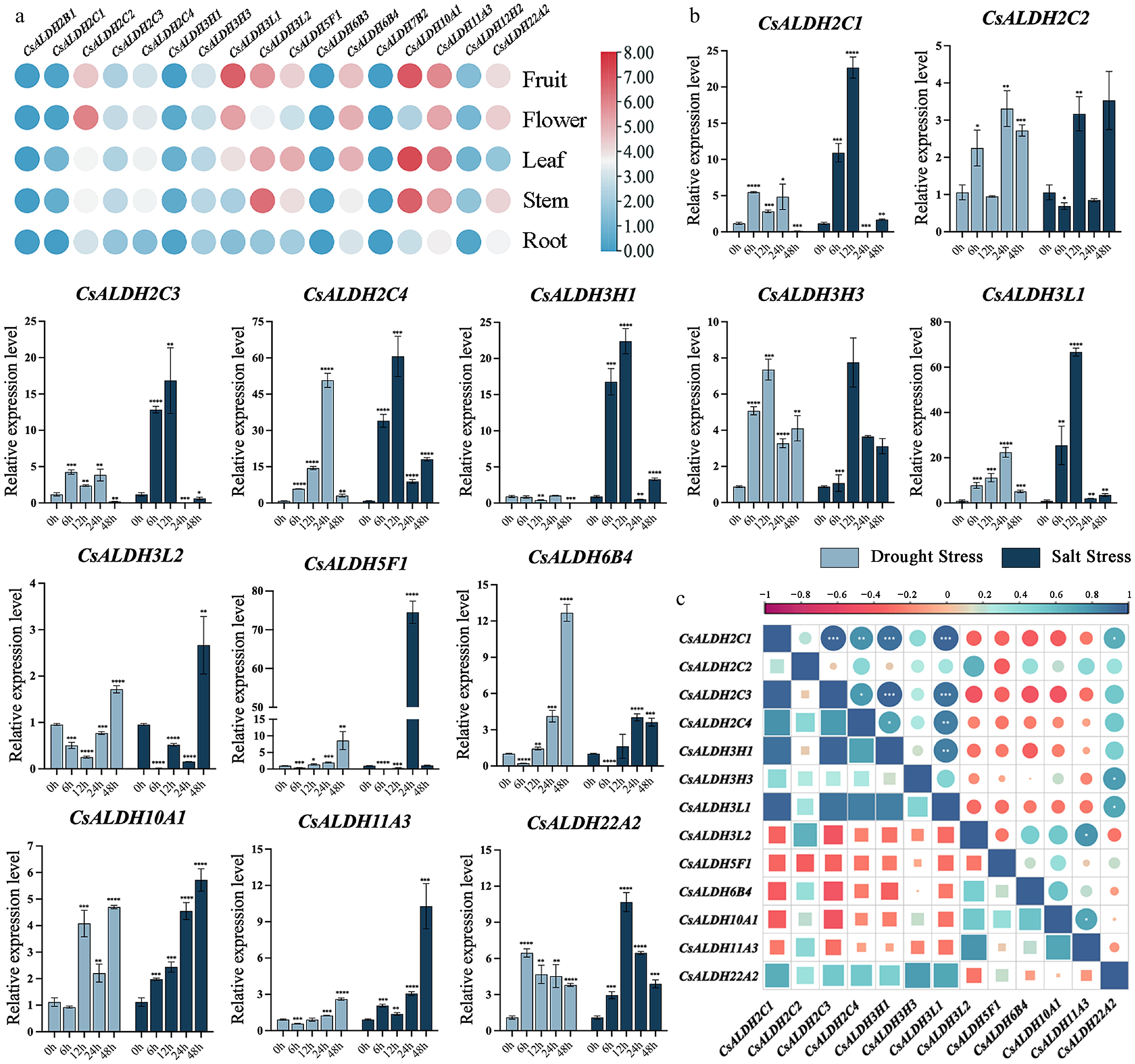
Figure 6.
(a) The heatmap of ALDHs expression in different tissues of C. sinense. (b) Expression patterns of ALDHs in C. sinense leaves at different time points after drought and salt stress. The x-axis represents time points (0 h as the control group, 6, 12, 24, and 48 h as experimental groups). The y-axis indicates the relative expression levels after exposure to stress for different durations. Light blue bars represent drought stress, while dark blue bars represent salt stress. The symbols *, **, ***, and **** denote significant differences at p < 0.05, p < 0.01, p < 0.001, and p < 0.0001, respectively. (c) Correlation analysis of qRT-PCR results. Correlation strengths are indicated by the graduated scale above.
Fourteen differentially expressed CsALDHs were screened, and their expression patterns at various time points after stress were analyzed by qRT-PCR. Figure 6b displayed 13 genes with significant expression changes, and the qRT-PCR data are presented in Supplementary Table S5. The primer sequences used for qRT-PCR are provided in Supplementary Table S6. The qRT-PCR results indicated that the ALDH family actively responded to drought and salt stress. Overall, the response of CsALDHs to salt stress was more pronounced than to drought stress. These results suggest that the ALDH gene family is indeed involved in the response to both drought and salt stress, but different genes exhibit distinct response mechanisms to these stresses. Under drought stress, the expression of most genes significantly increased before or at 12 h, while under salt stress, the expression of all CsALDHs significantly increased starting from 12 h. Some genes also showed significant stress-specific correlations (Fig. 6c).
Cloning and subcellular localization of CsALDH5F1 and CsALDH11A3
-
ALDH was a key element in oxygen-dependent metabolic transformation in cells. CsALDH5F1 and CsALDH11A3 were significantly upregulated at 24 and 48 h under salt stress. In phylogenetic trees, CsALDH5F1 clustered with AtALDH5F1, while CsALDH11A3 clustered with AtALDH11A3. Previous studies have shown that AtALDH5F1 is highly responsive to hypoxia stress, and AtALDH11A3 catalyzes the production of 3-phosphoglycerate and NADPH, which are essential for mannitol biosynthesis in many plant species[33]. Mannitol enhanced plants' ability to withstand stressful conditions, including drought and elevated soil salinity[34].
Therefore, we hypothesized that CsALDH5F1 and CsALDH11A3 also play roles in C. sinense's response to stress. We performed PCR amplification using C. sinense cDNA as a template with specific primers for CsALDH5F1 and CsALDH11A3. Agarose gel electrophoresis showed bands corresponding to the expected sizes of target fragments, and the full-length sequences of CsALDH5F1 and CsALDH11A3 were 780 and 531 bp, respectively (Fig. 7a). Subcellular localization prediction indicated that CsALDH5F1 localized in mitochondria, while CsALDH11A3 is located in the cytoplasm. To verify these predictions, we expressed the fusion proteins 35S::GFP-CsALDH5F1 and 35S::GFP-CsALDH11A3 in N. benthamiana leaves, with 35S::GFP-Super1300 as the control, and DAPI as the nuclear marker. The results confirmed that CsALDH5F1 was indeed localized in the mitochondria, while CsALDH11A3 was localized in the cytoplasm (Fig. 7b).
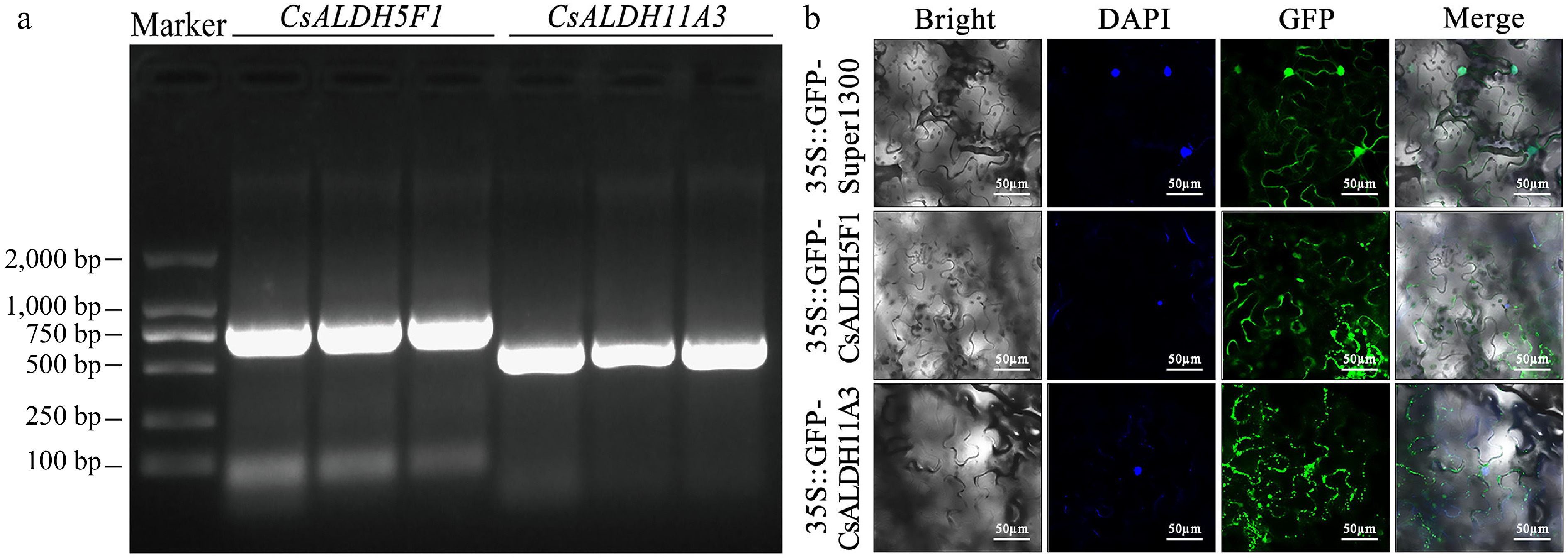
Figure 7.
(a) Agarose gel electrophoresis of PCR-amplified DNA full-length sequences. All samples exhibited a single band at the expected size, confirming the successful amplification of the target genes. (b) Subcellular localization of CsALDH5F1 and CsALDH11A3 in N. benthamiana epidermal cells.
-
ALDH genes are widely present in animals and plants and are involved in various life processes[35]. The genome-wide identification of CsALDHs, CeALDHs, and CgALDHs provides a foundation for further research on the functions of the ALDH superfamily in orchids. In this study, we identified a total of 17 ALDH genes in C. sinense, 14 in C. ensifolium, and 20 in C. goeringii. The numbers are comparable to those in A. thaliana (14)[35], rice (21)[36], pepper (27)[37], and melon (17)[38]. These results suggest that the ALDH family undergoes quantitative changes during long-term environmental adaptation, thereby enhancing plant resistance to environmental stress. As one of the most ecologically diverse plant families, orchids (Orchidaceae) are widely distributed across diverse habitats and exhibit unique life strategies[39]. The differential expansion of ALDH genes across orchid species likely reflects lineage-specific evolutionary responses to habitat-specific stressors. These findings offer new insights into the role of ALDH superfamily genetic adaptations in shaping the ecological success and niche-specific survival strategies of orchids. Multiple sequence alignment confirmed the conservation of key amino acid active sites in the ALDH family, specifically Glutamic (PS00687), and Cysteine (PS00070), findings that align with previous research[40]. Phylogenetic analysis provides a valuable theoretical framework for predicting functions of homologous genes across species. Genes clustered within the same phylogenetic branch exhibited conserved structural and functional characteristics, allowing for the inference of gene function based on prior studies. For instance, AtALDH5F1 and AtALDH11A3 in A. thaliana have been shown to respond to abiotic stress, with their expression levels decreasing under hypoxic conditions and being sensitive to temperature changes[1,33]. Therefore, it is reasonable to hypothesize that genes clustered with these ALDHs may possess similar or identical functions. Studies on OsALDHs suggest that certain ALDH genes involved in abiotic stress responses could contribute to the breeding improvement of rice[36]. Identifying stress-responsive ALDH genes in orchids will facilitate genetic improvement of orchid cultivars, enhancing their adaptability to extreme habitats.
The structural variations observed in ALDH genes are likely associated with the evolution and functional diversification of this gene family across different species. Structural analysis of the CsALDH, CeALDH, and CgALDH superfamilies revealed that while conserved motifs differ among families, they tend to be more similar within the same family. Analysis of intron and exon numbers indicated that most ALDH genes contain multiple introns and exons. Examination of cis-acting elements in the promoter regions showed that the upstream regions of ALDHs in the three orchid species contain numerous binding sites for transcription factors involved in light response, plant hormone signaling, cell development, and abiotic stress responses, such as B3, E2F, SBP, and bZIP[41]. In wheat, TaNF-YA7-5B has been shown to regulate stomatal closure under drought conditions, promoting leaf water retention and maintaining ROS homeostasis[42]. Heterologous expression of LEA proteins from Pinus tabuliformis in Escherichia coli demonstrated that most PtaLEAs enhanced the bacterium's tolerance to salt and heat stress[43].
Prediction of the CsALDHs PPI network revealed its interactions with numerous proteins associated with abiotic stress responses. For instance, Polyamine Oxidase (POP) plays a crucial role in polyamine metabolism, an essential process for plant growth, development, and stress tolerance[44]. Cytochrome P450 (CYP) is involved in suberin synthesis, significantly enhancing plant resistance to salt stress[45]. Alanine-Glyoxylate Transaminase (AGT) participates in glyoxylate metabolism, which is vital for maintaining metabolic homeostasis in plants[46]. Collectively, these proteins play pivotal roles in plant growth, development, stress responses, and metabolic regulation. As important regulatory factors, miRNAs participated in processes including plant growth, development, and stress responses[47]. The analysis of miRNA binding sites indicated that most miRNAs in C. sinense responded to various abiotic stresses; for example, miR169 was responsive to salinity, temperature, and drought stresses[48], while miR399a responded to phosphorus deficiency[49]. In the present study, CsALDH5F1 showed the most significant response under salt stress at the 24 h mark, which may be related to its targeting by miRNA414. Based on these predicted miRNA sites, we constructed a potential stress response pattern diagram. Consequently, we investigated the responses of C. sinense to drought and high salt stress.
The ALDH superfamily represents a large and complex group of enzymes. In this study, we analyzed the expression profiles of the CsALDH family in roots, stems, leaves, flowers, and fruits of C. sinense using transcriptome data. Most CsALDHs (14 out of 17) were expressed in these tissues. The tissue-specific expression patterns provide foundational data for exploring their potential functions. Following drought and high salt stress treatments on C. sinense, qRT-PCR analysis was conducted at different time points post-stress to determine the expression patterns of each gene under abiotic stress. ALDHs generally showed positive responses to both drought and salt stress. Notably, the expression levels of CsALDH5F1 and CsALDH11A3 increased significantly during specific periods following stress exposure. Subcellular localization confirmed that these genes are not localized in the nucleus, as predicted, but function in mitochondria and cytoplasm, consistent with previous studies in A. thaliana[33]. Our findings provided valuable insights and served as a reference for future research on abiotic stress responses in orchid plants.
-
Through genome-wide identification in C. sinense, C. ensifolium, and C. goeringii, we detected 17, 14, and 21 ALDH genes respectively. Gene structure and homology analyses demonstrated that the ALDH family exhibited high evolutionary conservation. Furthermore, prediction of PPI networks and miRNA binding sites indicated close associations between ALDHs and abiotic stress responses. Transcriptome and qRT-PCR analyses revealed that 13 differentially expressed CsALDHs played crucial roles in abiotic stress responses. Subcellular localization experiments in tobacco with CsALDH5F1 and CsALDH11A3 validated prediction accuracy and confirmed these CsALDHs' cellular locations.
The research was funded by the National Key Research and Development Program of China (2023YFD1600504), and the Research Cooperation Program of Fujian Agriculture and Forestry University (KH240047A).
-
The authors confirm contributions to the paper as follows: study conception and design: Wang L, Liu ZJ, Lan S; data collection: Wang L, Zheng R; experimental operation: Wang L, Lu J, Tian Y; analysis and interpretation of results: Wang L, Zhang C, Chen C; draft manuscript preparation: Wang L, Zhao X. All authors reviewed the results and approved the final version of the manuscript.
-
The genomic data of C. sinense are derived from NCBI: PRJNA743748. The genomic data of C. ensifolium are derived from NCBI: PRJNA694815. The genomic data of C. goeringii are derived from NCBI: PRJNA668552. All data generated or analyzed during this study are included in this published article and its supplementary information files, and also available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Protein physicochemical properties of CsALDHs, CeALDHs and CgALDHs.
- Supplementary Table S2 Glutamic (PS00687) and Cysteine (PS00070) active sites in CsALDHs, CeALDHs and CgALDHs.
- Supplementary Table S3 MicroRNA binding site prediction in CsALDHs.
- Supplementary Table S4 TPM values of CsALDHs in different plant tissues.
- Supplementary Table S5 Expression profiles of CsALDHs in leaves under drought and salt stress.
- Supplementary Table S6 The primers of CsALDHs.
- Supplementary Fig. S1 Chromosomal localization of genes. (a) Distribution of CsALDHs across chromosomes in C. sinense. (b) Distribution of CgALDHs across chromosomes in C. goeringii. (c) Distribution of CeALDHs across chromosomes in C. ensifolium.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang L, Zheng R, Zhang C, Chen C, Lu J, et al. 2025. Aldehyde dehydrogenase superfamily of three Cymbidium orchids: genome-wide identification and response models to drought and salt stress. Ornamental Plant Research 5: e033 doi: 10.48130/opr-0025-0035 

Aldehyde dehydrogenase superfamily of three Cymbidium orchids: genome-wide identification and response models to drought and salt stress
- Received: 31 December 2024
- Revised: 26 June 2025
- Accepted: 03 July 2025
- Published online: 28 August 2025
Abstract: Aldehydes serve as intermediates in various metabolic processes, and their excessive accumulation can disrupt normal plant metabolic activities. Environmental stressors, including drought and salinity, can trigger the generation of reactive oxygen species (ROS), which results in elevated levels of aldehydes within plant cells. Aldehyde dehydrogenases (ALDHs) are recognized as key enzymes that act as 'aldehyde scavengers' capable of detoxifying the harmful aldehydes produced during oxidative stress conditions. Orchids constitute one of the largest families within angiosperms and possess significant economic value. However, genetic data for these non-model plants are limited, and the molecular mechanisms underlying orchid responses to abiotic stress remain poorly understood. In this study, we identified 17, 14, and 21 ALDH family members in three Cymbidium orchids: C. sinense, C. ensifolium, and C. goeringii, respectively. We conducted comprehensive analyses of their conserved domains, promoter elements, and collinearity relationships and predicted the interaction networks of ALDH proteins and microRNA binding sites in these three orchid species. Furthermore, we subjected C. sinense to drought and high salt-stress treatments and examined its expression patterns under these conditions using quantitative real-time PCR (qRT-PCR). Subcellular localization experiments were also performed on genes that exhibited significant responses to stress. Our findings provide valuable insights into the molecular mechanisms of stress resistance in orchids and contribute to a deeper understanding of their potential applications as ornamental plants.
-
Key words:
- Aldehyde dehydrogenases /
- Chinese orchid /
- Abiotic stress /
- Response models