









-
The transition from vegetative to reproductive growth marks a critical phase in plant development, culminating in flowering[1]. This process is strictly regulated by the integration of endogenous genetic programs and autonomous timing mechanisms with external environmental cues such as photoperiod and temperature[2,3]. In Arabidopsis thaliana, floral development accelerates markedly following the formation of floral primordia, driven primarily by rapid elongation[4]. Plant morphogenesis, particularly scape elongation, is governed by intricate phytohormonal networks, with gibberellin (GA) biosynthesis gradients acting as pivotal developmental regulators[5]. These gradients trigger spatiotemporally ordered transcriptional cascades that link GA signaling to epigenetic reprogramming, driving stem elongation and developmental phase transitions[6]. The conservation of GA signaling across plant lineages is well established: exogenous GA3 promotes flowering in chrysanthemum[7], lily[8], and Paphiopedilum orchids—where it mediates photoperiod-dependent scape elongation and floral commitment[9]—while in rice, GA3 modulates submergence-adaptive internode elongation via biosynthetic flux and receptor signaling[10]. Molecularly, core GA pathway components—including the GA INSENSITIVE DWARF1 (GID1) receptor, DELLA repressors, and Phytochrome Interacting Factor (PIF) transcription factors—form a conserved regulatory module[11,12], as seen in lettuce, where the DELLA protein LsRGL1 controls scape elongation and flowering time via GA pathway regulation[13], highlighting the evolutionary adaptability of GA-driven growth-development coordination.
Studies in pea (Pisum sativum L.) have demonstrated that the auxin indole-3-acetic acid (IAA) promotes internode elongation through its regulatory effect on bioactive GA biosynthesis[14]. Concurrently, auxin orchestrates diverse developmental processes, such as cell elongation, organogenesis, and floral stem growth, with conserved regulatory roles across species[15]. In flowering Chinese cabbage, IAA coordinates signaling and transport to regulate internode expansion[16], while in peas, auxin directly induces internode elongation[17]. The auxin pathway, comprising TIR1 receptors, AUX/IAA repressors[18], and auxin response factor (ARF) transcription factors[19], dictates phytohormone function: TIR1 and AUX/IAA drive bolting in Saposhnikovia divaricata[20], and arf6/8 mutants exhibit stem growth defects, underscoring their necessity for inflorescence development[21,22]. Notably, the role of auxin in regulating scape elongation and anthesis in Clivia miniata remains uncharacterized.
C. miniata (Amaryllidaceae) is an understory species endemic to South African that has gained commercial prominence as an ornamental plant, especially in China, where it is traditionally associated with prosperity during the Lunar New Year celebrations[23]. However, its unpredictable flowering phenology, which is governed by complex genetic and environmental interactions, poses substantial challenges in horticulture[24]. Current flowering control methods, including exogenous hormone application and temperature manipulation, remain suboptimal[25]. Building on previous work identifying scape elongation as a key developmental checkpoint before anthesis[25], with the aim of screening effective hormones for regulating its flowering anthesis and further exploring effective regulatory measures, this study systematically explored the hormone profile during different stages of scape elongation in C. miniata. It demonstrated the regulatory roles of GA and auxin (IAA) in scape elongation through their systematic application. These findings contribute both to a fundamental understanding of hormonal regulation in monocot flowering and to the development of practical strategies for controlling the reproductive timing of this economically critical ornamental species.
-
C. miniata plants were cultivated in chambers with a controlled environment at Shenyang Agricultural University (41°49' N, 123°34' E). Plants were grown in 27 cm × 25 cm × 25 cm containers filled with an optimized growth medium (3:1:1 leaf mold : coconut chaff : compost, v/v/v; pH 6.0–6.5). The environmental parameters included a 10 h light/14 h dark photoperiod with a 20/10 °C thermoperiod and 65%–75% relative humidity. For metabolomic analyses, scapes with inflorescences were sampled from multiple plants at synchronized developmental time points. Four developmental stages were selected for analysis: the flower bud stage (CM_F), the inflorescence stage (CM_I), the inflorescence emergence stage (CM_E), and the scape stage (CM_S) (Fig. 1a). Triplicate biological replicates (entire scapes with inflorescences) were collected at synchronized time points in each stage, immediately flash-frozen in liquid nitrogen (within 30 s), and stored at −80 °C until liquid chromatography tandem mass spectrometry (LC-MS/MS) analysis.
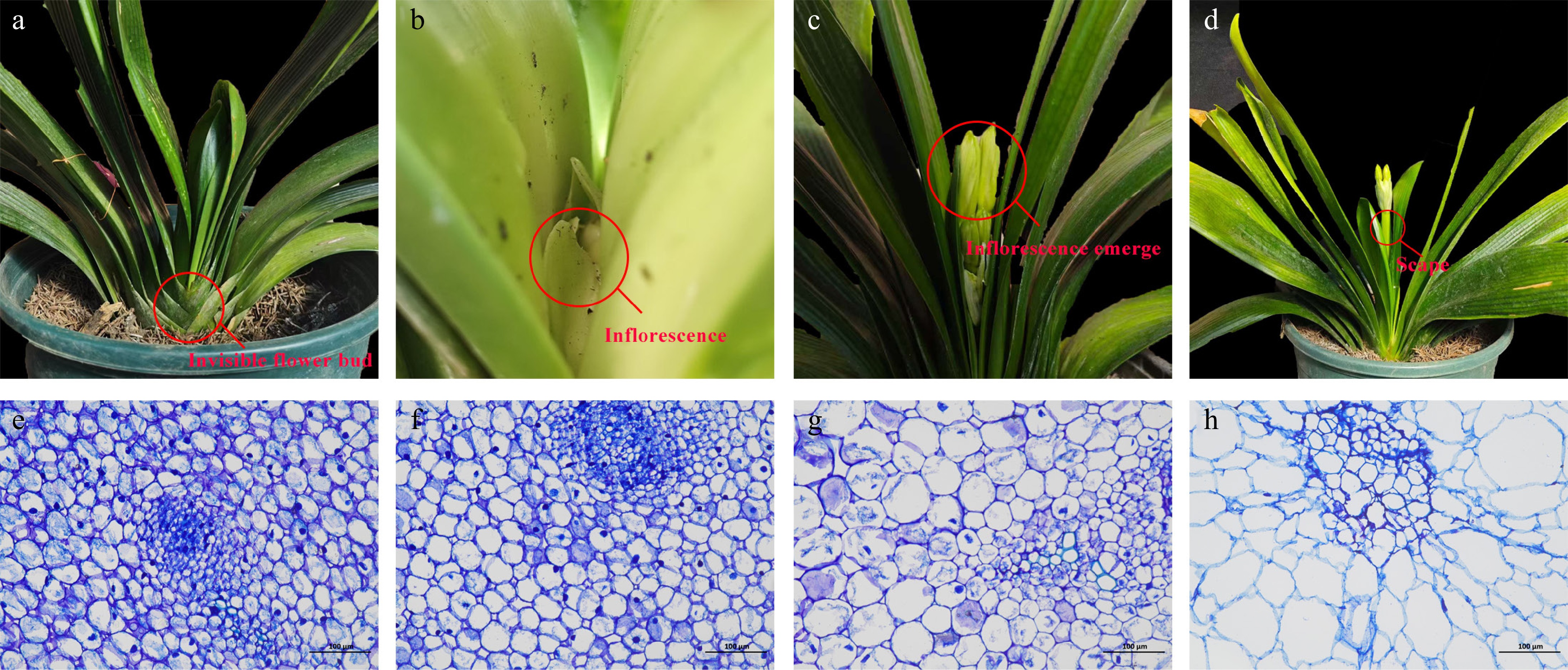
Figure 1.
The four developmental stages before anthesis (a−d) and their corresponding paraffin sections (e−h) of C. miniata. (a), (e) Flower bud stage, CM_F, in which the flower bud is invisible. (b), (f) Inflorescence stage, CM_I, when the flower bud develops into a complete inflorescence but both the flower bud and inflorescence are invisible. (c), (g) Inflorescence emergence stage, CM_E, when the inflorescence is visible but the scape is invisible. (d), (h) Scape stage, CM_S, when the scape elongates and both the inflorescence and scape are visible.
Microscopic observations
-
The scape samples underwent standardized histological processing: aseptic aqueous rinsing followed by formalin-acetic acid-alcohol (FAA) chemical fixation. The embedded specimens were sectioned with a microtome at 8 µm thickness following the standard paraffin protocols. The sections were stained with Mayer's hematoxylin, mounted with neutral gum, and imaged using a Leica RM2255 microtome system (Leica Biosystems) equipped with an Olympus DP27 digital camera. Morphometric measurements were performed on high-resolution digital images.
Metabolome assay
-
Fresh tissue samples (50 mg) from each developmental stage (CM_F, CM_I, CM_E, and CM_S) were cryo-pulverized in 1 mL of an ice-cold extraction buffer (methanol : water : formic acid, 15:4:1 v/v/v) containing 2 μL of stable isotope-labeled internal standards (100 ng/mL). After vortexing (10 min, 25 °C) and centrifugation (14,000 × g, 5 min, 4 °C), the supernatants were filtered (0.22-μm nylon mesh) and analyzed by LC-MS/MS (Agilent 6495C QqQ) with the samples maintained at 4 °C during acquisition.
Metabolite profiling was performed on a hyphenated ultrahigh-performance liquid chromatography–electrospray ionization-tandem mass spectrometry (UHPLC-ESI-MS/MS) platform (ExionLC AD UHPLC coupled with a QTRAP 6500+ hybrid triple quadrupole/linear ion trap, Sciex, Shanghai). Chromatographic separation was performed using an ACQUITY UPLC HSS T3 C18 reversed-phase analytical column (100 mm × 2.1 mm inner diameter, 1.8 μm; Waters) under thermostatic conditions (40 °C), with 2-μL injection volumes. A binary solvent system of 0.04% (v/v) acetic acid in water (Phase A) and 0.04% acetic acid in acetonitrile (Phase B) was used at a flow rate of 350 μL/min. A seven-step multisegment gradient was optimized: 2% B (0–1 min), linear ramping to 95% B (1–8 min), isocratic elution (8–9 min), and re-equilibration to the initial conditions (9–12 min).
The QTRAP 6500+ hybrid triple quadrupole–linear ion trap mass spectrometer (Sciex), equipped with a Turbo V electrospray source and operated in polarity-switching mode at ±5,500 V, was calibrated by conducting iterative assessments of the ion suppression effects. The critical ionization parameters included 35 psi of the curtain gas (N2), 550 °C as the source temperature, and dual nebulizer/auxiliary gases at 35 psi each. Time-scheduled multiple reaction monitoring (MRM) acquisition (dwell time: 50 ms) monitored 214 precursor–product ion pairs for 32 phytohormone subclasses, with the declustering potentials (65–120 V) and collision energies (15–35 eV) dynamically optimized by compound-specific tuning. Raw spectral data processing was performed using Analyst 1.6.3 acquisition software with Multiquant 3.0.3 validation algorithms, implementing peak-area normalization against deuterated internal standards and matrix-matched calibration curves (R2 > 0.998).
Metabolomic data processing was performed using an XCMS-based computational pipeline[26] for chromatographic peak alignment, feature extraction, intensity-based normalization, spectral deconvolution, and putative metabolite annotation. Multivariate pattern recognition via principal component analysis (PCA) was performed using OriginPro 2022 (OriginLab Corporation, USA) with feature scaling to unit variance prior to eigen decomposition. Metabolites were identified using MetaX[27] with mass accuracy thresholds of < 2 parts per million (ppm) against the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and the Human Metabolome Database (HMDB). Differentially abundant metabolites were defined by a fold change of > 2 or < 0.5 with Benjamini–Hochberg adjusted p < 0.05. Correlation networks were visualized using Cytoscape 3.9.1[28] to construct weighted interaction graphs from Pearson's correlation matrices (|r| > 0.95, p < 0.05).
Hormone treatments applied
-
Uniform 4-year-old C. miniata plants were selected for exogenous hormone application in the greenhouse at Shenyang Agricultural University. At the beginning of the treatment, C. miniata plants were at the CM_I stage. Treatment groups included the control (CK, distilled water), GA3 (50, 100, 150, and 200 mg/L), and IAA (50, 100, 150, and 200 mg/L). Each treatment group included 10 biological replicates. Hormone solutions (1 L per treatment) were applied to the soil for root absorption at the treatment's initiation (1 October 2024), followed by subsequent applications every 10 d until 30 November 2024. Phenological stages—specifically inflorescence emergence and anthesis—were monitored throughout the treatment period (Supplementary Table S1).
Quantitative reverse transcription-polymerase chain reaction
-
Total RNA isolated from biological specimens (1 μg input) was subjected to first-strand cDNA synthesis employing a gDNA-eliminating PrimeScript RT reagent kit with gDNA removal (TaKaRa Bio, China). Synthesized cDNA was normalized to 200-μL working stocks through spectrophotometric quantification (ratio of absorbance at 260 nm and at 280 nm [A260/A280] = 1.8–2.0) to ensure the template's uniformity for quantitative reverse transcription–polymerase chain reaction (qRT-PCR) validation across experimental replicates. Conserved regions of selected differentially expressed genes were amplified using primers designed with Primer3Web (v4.1.0,
https://primer3.ut.ee ). All primers used in this study are listed in Supplementary Table S2. A TaqMan Universal SYBR qPCR Master Mix Real-Time PCR system (Vazyme, Nanjing) was used for gene quantitative detection, with 18S rRNA serving as the internal reference gene[29]. Each group included three technical replicates and three biological replicates. Relative gene expression was calculated using the 2−ΔΔCᴛ method[30]. Statistical significance was assessed by one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. Quantitative traits are presented as the mean ± standard deviation (n = 5 biological replicates), and data were visualized using GraphPad Prism 8.0.2 (GraphPad Software, USA) with optimized perceptual color mapping for scientific clarity.Statistical analysis
-
All analyses were visualized with GraphPad Prism version 8.0.2 software (San Diego, California, USA). All data of the treatments are expressed as the mean ± standard deviation. Data were analyzed using one-way ANOVA, and the differences were measured using Duncan's test (p < 0.05) within GraphPad Prism.
-
First, the preflowering developmental process of C. miniata was observed. The flower bud differentiation stage was invisible, as the bud remained embedded at the base of the pseudobulb. In a previous study, we performed detailed observations of the flower bud differentiation stage (CM_F). At this time, the flower bud was invisible (Fig. 1a)[25]. Once floral organ development was completed, the inflorescence was formed, marking the inflorescence stage (CM_I). At this stage, the scape had not yet started to elongate and the inflorescence was invisible. However, when we stripped away the leaves, a fully developed inflorescence could be observed (Fig. 1b). Then the scape began to elongate and ejected the inflorescence of the leaves, which was marked as the inflorescence emergence stage (CM_E). At this stage, the inflorescence was visible and the scape was invisible (Fig. 1c). As elongation continued, the plant entered the scape stage (CM_S) and the leaves emerged, eventually leading to anthesis. At this stage, both the inflorescence and scape were visible (Fig. 1d). Paraffin sections prepared from the four developmental stages (CM_F, CM_I, CM_E, and CM_S; Fig.1e−h) revealed coordinated cellular changes associated with scape elongation. A proportional expansion in both the transverse and longitudinal directions was demonstrated, indicative of regulated anisotropic growth. Microscopic analysis showed progressive enlargement and elongation of the parenchyma cells, consistent with the observed macroscopic growth patterns.
Qualitative and quantitative analyses of hormone metabolomics
-
This study identified and quantified 60 hormones across 8 major classes: 16 cytokinins (CKs), 14 auxins, 12 GAs, 11 jasmonates (JAs), 4 salicylic acids (SAs), 1 ABA, 1 ethylene (ETH), and 1 melatonin (MLT). Normalized hormone content data were visualized via heatmaps (Fig. 2a). Salicylic acid 2-O-β-glucoside, 2-methylthio-cis-zeatin riboside, 6-benzyladenosine, and dihydrozeatin-O-glucoside riboside were exclusively detected in the CM_F stage, coinciding with the completion of flower bud differentiation. These hormones are likely to have a minimal influence on subsequent scape elongation.
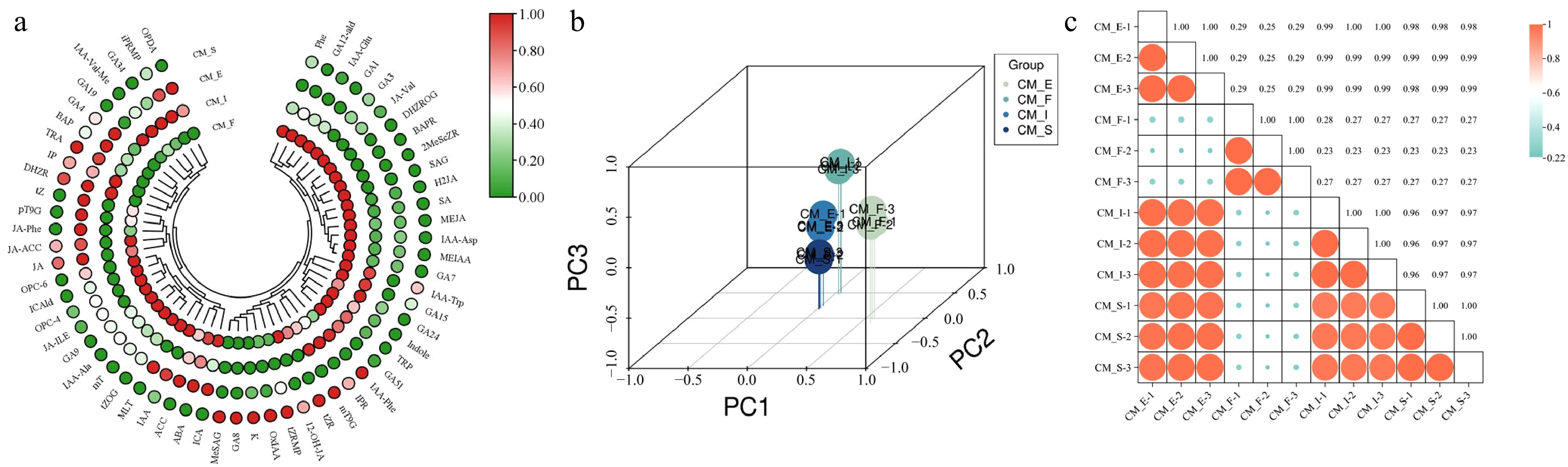
Figure 2.
Quality control analysis of hormone metabolomics in different stages of scape elongation in C. miniata. (a) Heatmap of hormone metabolites. (b) PCA score plot. (c) Pearson correlation matrix of phenotypes.
PCA was used to assess the overall hormonal differences among developmental phases and to evaluate variability within groups to investigate natural variations in hormone profiles across various scape elongation stages. The analysis revealed distinct hormonal profiles among the four stages (CM_F, CM_I, CM_E, and CM_S), with minimal intrastage variation, thus demonstrating strong sample reproducibility (Fig. 2b). Intergroup and intragroup correlation analyses revealed high consistency among biological replicates within each stage. CM_S, CM_I, and CM_E exhibited significant correlation compared with CM_F, possibly reflecting the unique physiological state of CM_F. This represented the flower bud differentiation phase prior to scape elongation (Fig. 2c).
Analysis of differentially expressed hormones
-
Hormones exhibiting a fold change of ≥ 1 or ≤ 0.5, with a false discovery rate (FDR) of < 0.05, were classified as differentially expressed. The analysis identified 52 hormones with significant variations across the four developmental stages, comprising 13 auxins, 12 CKs, 11 GAs, 10 JAs, 3 SAs, 1 MLT, 1 ETH, and 1 ABA. Comparative analysis revealed that 10, 11, and 8 hormones were upregulated, whereas 30, 31, and 35 were downregulated in CM_I vs CM_F, CM_E vs CM_F, and CM_S vs CM_F, respectively (Fig. 3a). A Venn diagram identified 20 common differentially expressed hormones across all comparisons, comprising seven JAs, four auxins, three GAs, three CKs, and three SAs (Fig. 3b). A heatmap illustrating the normalized expression profiles of these 20 hormones is presented in Fig. 3c. Correlation network analysis among the 20 analyzed hormones was performed to systematically explore potential interactions. Nodes represented individual hormones, and edges indicated statistically significant (p < 0.05) metabolic correlations. Positive correlations predominated over negative interactions in these networks. The network analysis revealed that IAA-aspartate (IAA-Asp) emerged as the most interconnected node, suggesting its potential role as a central hormonal regulator. Methyl ester IAA (MEIAA) also showed strong co-regulation patterns, particularly with GA7, indicating its functional association in the hormonal network (Fig. 3d).
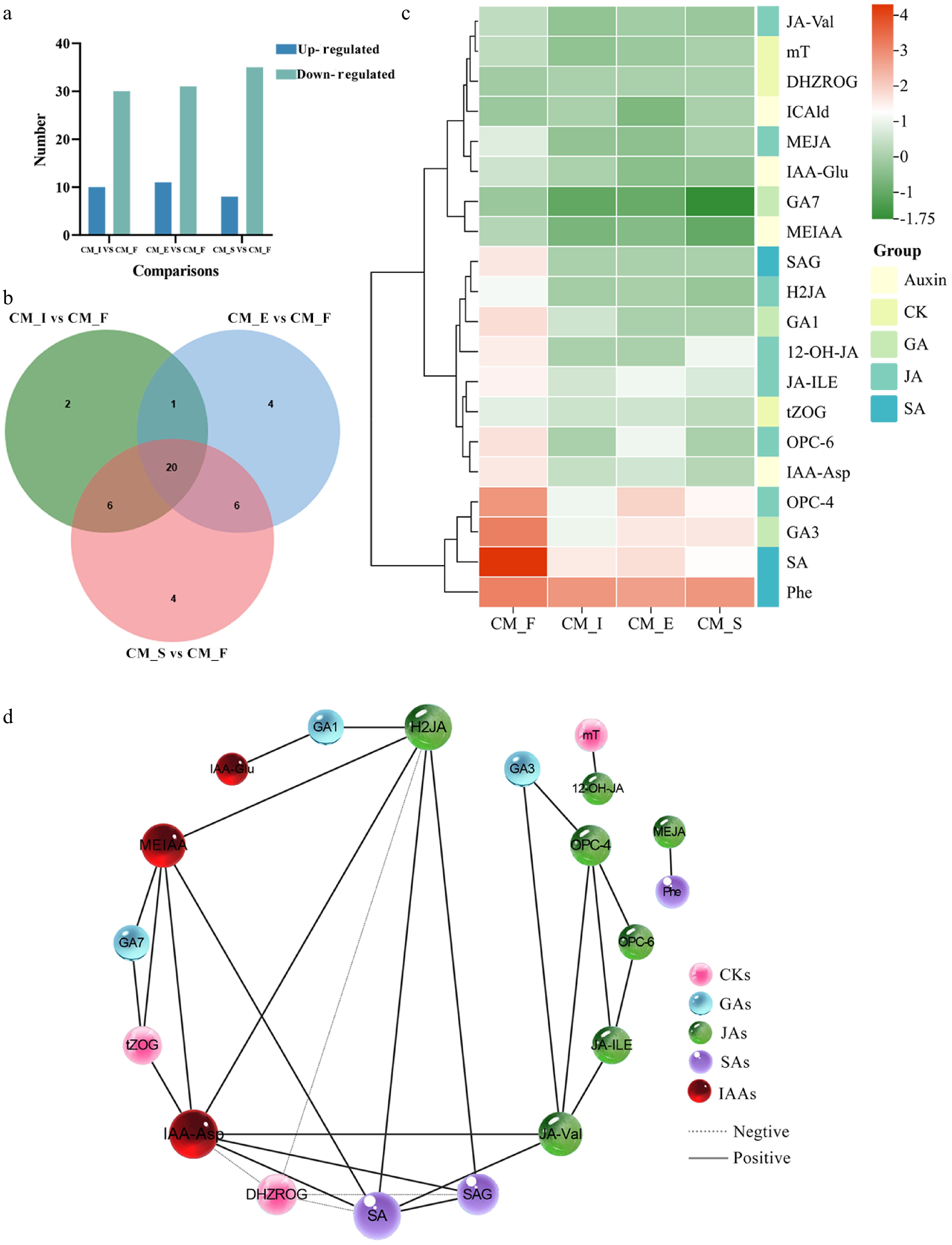
Figure 3.
Overview of differentially expressed metabolites. (a) Pairwise comparisons of different stages. Blue and green bars represent differentially upregulated and downregulated metabolites, respectively. (b) Venn diagrams highlighting different comparisons. (c) A heatmap of 20 differentially expressed hormones. (d) A hormone correlation network analysis of 20 differentially expressed hormones based on Pearson's correlation coefficients (|r| > 0.95, p < 0.05). The dotted lines between two metabolites represent negative correlations, and the solid lines between two metabolites represent positive correlations. The width of the line between two metabolites changes with the size of the correlation r: The thinner the line, the smaller the correlation.
The line chart (Fig. 4) clearly demonstrates the content changes in the four auxins, including indole-3-carboxaldehyde (ICAld), IAA-Asp, indoleacetyl glutamic acid (IAA-Glu and MEIAA); three GAs, including GA1, GA3, and GA7; three CKs, including dihydrozeatin-O-glucoside riboside (DHZROG), trans-zeatin-O-glucoside (tZOG), and meta-topolin (mT); seven JAs, including 3-oxo-2-(2-[Z]-pentenyl) cyclopentane-1-butyric acid (OPC-4), jasmonoyl-L-isoleucine (JA-ILE), dihydrojasmonic acid (H2JA), methyl jasmonate (MEJA), 12-hydroxyjasmonic acid (12-OH-JA), N-[(-)-Jasmonoyl]-(L)-valine (JA-Val), 3-oxo-2-(2-[Z]-pentenyl)cyclopentane-1-hexanoic acid (OPC-6); and three SAs, including L-phenylalanine (Phe), salicylic acid (SA), and salicylic acid 2-O-β-glucoside (SAG) during the scape elongation stage. The findings revealed significant upregulation of IAA-Asp, GA3, OPC-4, JA-Ile, H2JA, JA-Val, OPC-6, and SA levels from the CM_I to the CM_E stage, which coincided with the rapid elongation of the scape.

Figure 4.
The expression trend charts of five hormone categories. (a) IAA, auxins; (b) GAs, gibberellins; (c) CKs, cytokinins; (d) JAs, jasmonates; (e) SAs, salicylic acids.
Promotion of anthesis by exogenous GA3 and IAA application
-
GAs and auxins (IAA) play pivotal regulatory roles in floral transition, with exogenous applications of GA3 and IAA extensively employed in anthesis research. Treatment with both GA3 and IAA significantly advanced the inflorescence emergence stage, scape elongation, and anthesis date of C. miniata, with varying degrees of effect (Tables 1, 2 & Fig. 5). Notably, treatment with 150 mg/L IAA and 100 mg/L GA3 exhibited the most pronounced promotion of individual inflorescence emergence, advancing the event by 31 and 29 d relative to the control, respectively. Notably, the earliest scape emergence and flowering phenology were induced by 150 mg/L IAA and 100 mg/L GA3, with 200 mg/L IAA and 150 mg/L GA3 as suboptimal treatments. However, scape emergence rates were universally decreased across all treatments except 50 mg/L IAA, with 150 mg/L IAA and 150 mg/L GA3 exhibiting the highest rate at 90%. Further, 100 mg/L IAA and 100 mg/L GA3 advanced anthesis by 34 and 38 d, respectively. Collectively, our analyses identified 150 mg/L IAA and 200 mg/L GA3 as the optimal treatment concentrations for promoting early anthesis in C. miniata.
Table 1. Statistics of critical developmental stages for C. miniata treated with different concentrations of IAA in 2025.
IAA concentration (mg/L) Date of inflorescence emergence (month/day) Date of scape elongation
(month/day)Date of anthesis (month/day) Days of advanced anthesis (days) Rate of scape emergence (%) 0 (control) 2/2 ± 0.5 2/15 ± 0 2/26 ± 0.5 0c 100.0 50 1/17 ± 0.5 1/26 ± 0.5 2/12 ± 0.5 14.21 ± 3.59b 100.0 100 1/16 ± 0.5 1/25 ± 0.5 1/23 ± 0.5 34.19 ± 0.97a 20.0 150 1/1 ± 0.5 1/14 ± 0.5 1/28 ± 0.5 29.81 ± 3.52a 90.0 200 1/6 ± 0.5 1/22 ± 0.5 2/2 ± 0.5 24.15 ± 2.12a 60.0 Different lowercase letters in the column indicate statistically significant differences (p < 0.05) as determined by Duncan's multiple range test. Table 2. Statistics of critical developmental stages in C. miniata treated with different concentrations of GA3 in 2025.
GA concentration (mg/L) Date of inflorescence emergence (month/day) Date of scape elongation (month/day) Date of anthesis (month/day) Days of advanced anthesis (days) Rate of scape emergence (%) 0 (control) 2/2 ± 0.5 2/15 ± 0 2/26 ± 0.5 0c 100.0 50 1/5 ± 0.5 1/18 ± 0.5 1/22 ± 0.5 35.15 ± 3.59a 50.0 100 1/3 ± 0.5 1/9 ± 0.5 1/19 ± 0.5 38.31 ± 0.97a 50.0 150 1/9 ± 0.5 1/18 ± 0.5 2/1 ± 0.5 25.33 ± 1.52b 90.0 200 1/4 ± 0.5 1/10 ± 0.5 1/22 ± 0.5 35.81 ± 3.52a 80.0 Different lowercase letters in the column indicate statistically significant differences (p < 0.05) as determined by Duncan's multiple range test. 
Figure 5.
Timeline of inflorescence emergence, scape elongation, color onset, and anthesis in C. miniata under different treatments.
Validation of related genes' expression through GA and IAA treatments
-
RT-qPCR analysis was performed to quantify the transcriptional dynamics of key components in GA and auxin signaling pathways across scape tissues treated with various concentrations of phytohormones to elucidate the molecular mechanisms underlying hormonal regulation (Fig. 6). Specifically, the key genes involved in GA signal transduction were examined, including CmGID1 (a nuclear receptor of GA signalling[29]), CmSLR1 (a DELLA protein, repressor of GA signalling[30]), and CmPIF3 (a transcription factor downstream from DELLA proteins[31]), as well as those in the IAA signal transduction pathway, such as CmAUX/IAA10 (a auxin transcription factor [TF] repressor), CmARF1, CmARF4, CmARF11, and CmARF18 (auxin signal TFs downstream of AUX/IAA). The coding DNA sequence data were derived from transcriptome sequencing of C. miniata performed in the authors' laboratory and presented in previously unpublished findings.

Figure 6.
Relative expression of scape elongation-related genes after different treatments. Error bars = standard deviation. **** p < 0.01 (Student's t-test).
RT-qPCR analysis revealed a significant induction of auxin response factors, with CmARF1, CmARF4, CmARF11, and CmARF18 exhibiting marked upregulation in response to both moderate (50 mg/L) and elevated (150 mg/L) IAA concentrations. Similarly, CmAUX/IAA10 expression was upregulated after exposure to 50 and 100 mg/L IAA, although only the change observed under the 50 mg/L IAA treatment was statistically significant. In contrast, treatment with 150 and 200 mg/L GA resulted in substantially upregulated expression of CmGID1 and CmPIF3. The expression of CmSLR1, a DELLA protein, was downregulated following treatment with 150 mg/L GA.
These findings collectively suggest that the exogenous application of GA and IAA triggers distinct responses in their respective signal transduction pathways. The differential expression patterns of CmAUX/IAA10, CmARF1, CmARF4, CmARF11, CmARF18, CmGID1, CmSLR1, and CmPIF3 suggest their potential roles in regulating the differential accumulation of IAA and GA during scape elongation.
-
Cellular differentiation has been established as a crucial determinant of stem elongation in the family Brassicaceae, including economically important species such as Brassica juncea and B. oleracea[32]. Building on this foundation, the present study systematically characterized four consecutive developmental stages in C. miniata: (1) the flower bud differentiation stage (CM_F), (2) the inflorescence stage (CM_I), (3) the inflorescence emergence stage (CM_E), and (4) the scape elongation stage (CM_S). The histomorphometric analysis of paraffin-embedded scape sections demonstrated stage-dependent cellular elongation patterns directly correlating with macroscopic scape growth (Fig. 1). These findings revealed a conserved mechanism coordinating cellular differentiation with organ-level growth dynamics during scape elongation.
Plant metabolomics has emerged as a powerful tool for elucidating the metabolic networks governing developmental processes[33]. Specifically, scape growth and flowering are precisely controlled by dynamic phytohormone interactions. Comparative metabolomics of early-bolting Angelica sinensis cultivars revealed ABA accumulation as a crucial factor regulating flowering time[34]. Complementary studies in Hylomecon japonica demonstrated that vernalization-induced floral transition was accompanied by a decrease in IAA levels and an increase in the GA3/ABA ratio[35]. These findings underscore the conserved role of hormone homeostasis in plants' reproductive development. This study profiled hormones across four developmental stages (CM_F, CM_I, CM_E, and CM_S) in C. miniata to decipher the regulatory network governing scape elongation. For this purpose, 52 differentially expressed hormones were identified, elucidating the underlying mechanisms of scape elongation in this ornamental species. This study detected 20 conserved differentially expressed hormones among the three pairwise comparisons (CM_I vs CM_F, CM_E vs CM_F, and CM_S vs CM_F). The predominant hormonal classes included JAs, auxins, GAs, CKs, and SAs, suggesting their central regulatory functions. An especially remarkable finding was the consistently high expression of L-Phe throughout flower bud differentiation and scape elongation, indicating its potential involvement in hormone biosynthesis, which is crucial for developmental progression. The network analysis of hormone correlations identified IAA-Asp as a central hub within the network. The strong correlation between MEIAA and GA7 implies substantial crosstalk between the auxin and GA pathways, potentially coordinating growth processes during scape elongation. Overall, these findings significantly enhance the understanding of the hormonal regulation of scape elongation.
The transition to flowering in plants is precisely regulated by complex molecular networks integrating internal developmental cues with external environmental signals[36]. GAs serve as central regulators of this process, coordinating reproductive timing across diverse species[37,38]. The key findings establish the regulatory role of GA in stem elongation. GA3-induced stem elongation is reversibly inhibited by chlorocholine chloride-mediated suppression of GA biosynthesis in Brassica napus[39]. Exogenous GA application directly modulates stem elongation kinetics in Triticum aestivum[40]. Previous studies demonstrated the fundamental role of GA in regulating the developmental progression to flowering through stem growth modulation. In the present study, the expression dynamics of GA3 exhibited a distinct temporal pattern in C. miniata. The expression levels began to rise in the CM_I stage and peaked in the CM_E stage. This expression pattern indicated the involvement of GA3 in the early phases of scape development, particularly in scape elongation. A controlled dose-response experiment was conducted using 10 plants per treatment group to further clarify the role of GA3. This was followed by foliar applications of 50–200 mg/L GA3. The 100 and 200 mg/L treatments led to the earliest scape emergence, compared with the control plants, which showed a 1-month delay. The 100 mg/L treatment advanced anthesis by 38 d, corroborating the authors' previous findings[25] and validating the role of GA3 in promoting flowering in C. miniata. Disruptions in GA signaling components often result in abnormal flowering phenotypes. The GA signaling pathway is regulated by an evolutionarily conserved GID1-DELLA module. However, F-box proteins display lineage-specific divergence, with AtSLY1 functioning in A. thaliana and OsGID2 in O. sativa[40]. Bioactive GA isoforms form ternary complexes with GID1 and DELLA proteins. This interaction triggers the polyubiquitination of DELLA proteins by Skp, Cullin, F-box containing (SCF) complex E3 ligases, followed by their proteasomal degradation, thereby activating the expression of GA-responsive genes[41−43]. This process regulates stem elongation and reproductive transitions through hierarchical cascades of phosphorelay signaling[13]. PIFs regulate flowering in both Phalaenopsis and Chinese pine by integrating GA signaling through direct interactions with DELLA proteins[12,44]. An investigation of GA signaling components in C. miniata revealed a significant upregulation of CmGID1 expression at the onset of scape elongation in all GA-treated groups compared with the controls. Additionally, treatment with 150 and 200 mg/L of GA led to substantially upregulated expression of CmPIF3. The consistent induction of CmGID1 and CmPIF3 suggests their potential role as positive regulators of scape elongation in C. miniata.
The auxin class of phytohormones orchestrates essential developmental processes, from embryogenesis to tropic responses[45]. IAA, the primary active auxin synthesized in apical meristems, regulates stem elongation in various species, including Narcissus pseudonarcissus and Paeonia suffruticosa[46]. Although exogenous IAA accelerates bolting in Lactuca sativa[47], the metabolomic analysis of C. miniata in this study identified two distinct auxin derivatives as the key scape elongation regulators: MEIAA and IAA-Asp. These findings revealed species-specific adaptations in auxin-mediated growth regulation while maintaining the conserved role of hormones in developmental control.
Systematic dose-response experiments were conducted, applying 50–200 mg/L IAA. Specifically, the 150 mg/L IAA treatment proved most effective in promoting early inflorescence emergence. In contrast, control plants exhibited a delay in scape emergence of approximately 1 month compared with the plants in the treatment groups. Although the 50 mg/L IAA treatment yielded emergence rates equivalent to those of the controls (100%), all treatment concentrations accelerated anthesis. The 100 mg/L IAA treatment had the most pronounced effect, advancing anthesis by 34 d. These results establish a concentration-dependent function of exogenous IAA in regulating the anthesis time of C. miniata. The findings can significantly enhance the understanding of auxin-mediated flowering control, particularly in monocot species.
In A. thaliana, the auxin signaling pathway is regulated by the TIR1/AFB receptor family. This family facilitates the ubiquitin-dependent degradation of Aux/IAA repressors, thereby initiating the subsequent activation of ARF transcription factors[48−49]. Once activated, the ARFs regulate the expression of auxin-responsive genes, including members of the Aux/IAA and GH3 families as their transcriptional targets[50]. Similarly, in L. sativa, several ARF homologs, such as LsARF2, LsARF3, and LsARF8a, mediate auxin responses[51,52]. This conservation of the auxin signaling pathway across various plant species underscores its fundamental significance in plant biology. The present study on auxin signaling in C. miniata revealed that IAA upregulated AUX/IAA10 expression. Additionally, the exogenous application of 50 and 150 mg/L IAA led to increased expression of multiple CmARF genes (CmARF1, CmARF4, CmARF11, and CmARF18) in scape tissues. These results suggest a role of the components of the auxin signaling pathway in regulating scape elongation. Thus, IAA likely promotes stem growth by activating the transcription of key signaling components.
-
This study comprehensively elucidated the regulatory mechanisms underlying scape elongation and anthesis in C. miniata. By systematically analyzing four developmental stages, the study revealed that scape elongation is a pivotal determinant of anthesis timing, operating independently of flower bud differentiation. The histological analysis clearly linked cellular elongation to scape growth. The metabolomic profiling in this study identified 52 differentially expressed hormones, including 20 conserved regulators such as JAs, auxins, GAs, CKs, and SAs, which together formed a complex regulatory network. The consistent expression of L-Phe throughout development and the identification of IAA-Asp as a central hub in the network underscored their crucial regulatory roles. Functional experiments demonstrated that GA3 promoted scape emergence and anthesis in a dose-dependent manner. This effect was corroborated by the upregulation of GID1, a key component of the GA signaling pathway. Moreover, the present study revealed that auxin exerted concentration-specific effects on scape development by regulating AUX/IAA10 and ARF genes' expression. Collectively, these findings significantly enhance our understanding of scape development. They also provide a solid framework for future mechanistic studies and offer valuable insights for horticultural strategies to precisely control the time of anthesis.
This study was supported by the Basic Scientific Research Project for Universities of the Education Department of Liaoning Province in 2022 (LJKMZ20222130).
-
The authors confirm contributions to the paper as follows: study conception and design, data collection, analysis, and interpretation of results: Ji Y; draft manuscript preparation: Ji Y, Lei J, Xue L; manuscript revision: Lei J, Xue L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 IAA, GA, and CK treatments.
- Supplementary Table S2 qRT-PCR primers.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ji Y, Xue L, Lei J. 2025. Regulation of scape elongation through gibberellin and auxin detected by hormone metabolomic profiling in Clivia miniata. Ornamental Plant Research 5: e041 doi: 10.48130/opr-0025-0037 

Regulation of scape elongation through gibberellin and auxin detected by hormone metabolomic profiling in Clivia miniata
- Received: 05 May 2025
- Revised: 15 July 2025
- Accepted: 28 July 2025
- Published online: 27 October 2025
Abstract: Clivia miniata Regel is a globally prized ornamental plant with significant economic value in China, as a traditional decoration during Lunar New Year celebrations. However, its natural anthesis time in the greenhouse often fails to coincide with the Spring Festival, thereby necessitating precise control over anthesis through a better understanding of its scape elongation mechanism. Phenological observations identified four key stages (flower bud stage, CM_F; inflorescence stage, CM_I; inflorescence emergence stage, CM_E; and scape stage, CM_S) before anthesis. Observations revealed progressive elongation of parenchyma cells in the developing scape. Hormone profiling across these stages was conducted to explore the hormonal regulation of this process, identifying 60 distinct hormones, 52 of which showed significant differential expression. Among these, auxins and gibberellins (GAs) emerged as primary regulators of both scape elongation and anthesis time. Building on these insights, exogenous hormone application experiments were performed with remarkable practical outcomes. All phytohormone treatments significantly accelerated anthesis. Specifically, applying 50 mg/L indole-3-acetic acid (IAA) resulted in complete inflorescence emergence with 100% efficiency and accelerated anthesis by 14 d. The application of 150 mg/L IAA resulted in complete inflorescence emergence with 90% efficiency and accelerated anthesis by 29 d. Moreover, the application of 150 mg/L GA3 resulted in complete inflorescence emergence with 90% efficiency and accelerated the anthesis by 25 d. The application of 200 mg/L GA3 resulted in complete inflorescence emergence with 80% efficiency and accelerated anthesis by 35 d, demonstrating unprecedented potential for manipulating anthesis timing. These findings elucidate the physiological and molecular mechanisms underlying anthesis time in C. miniata.
-
Key words:
- Clivia miniata /
- Scape elongation /
- Hormone profiling /
- Anatomical observation /
- Anthesis regulation